
Summary
We have studied the visually triggered orienting behavior of frogs following complete unilateral transection of the neuraxis at the junction of the medulla and spinal cord, as well as after smaller lesions at the same level. Complete transection produces the same behavioral deficit as previously reported (Kostyk and Grobstein 1982, 1987a) for a similar lesion at the junction between midbrain and medulla. Lesioned frogs failed to turn toward stimuli at all locations in the ipsilateral visual hemifield, responding instead with forwardly directed movements in which there was a persistance of variations related to stimulus elevation and distance. Responses to stimuli in the contralateral visual hemifield were normal. Similar deficits were seen after smaller lesions restricted to a medial white tract. Partial damage to the tract resulted in turns of reduced amplitude for stimuli throughout the ipsilateral hemifield. Lesions to adjacent tissue were without effect on the behaviors studied. In all animals, we observed a strong correlation between turn amplitude for lateral stimuli and the distance at which the animals switched from snapping to hopping. These observations provide new evidence that a transformation from a retinocentric to a lateralized and parcellated form of spatial representation occurs in going from the retinotectal projection to the descending tectofugal pathway in the caudal midbrain, and that this form of representation remains stable until the spinal cord. A second transformation involved in determining the actual movement to be triggered must occur subsequently. Our findings also suggest that the signals underlying orienting turns may not descend into the spinal cord on tectospinal axons, and suggest that the lateralization of descending signals probably occurs conicidentally with a synaptic relay in the midbrain tegmentum.
Similar content being viewed by others
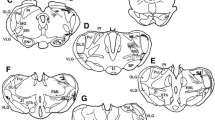

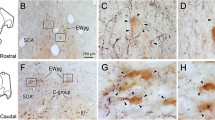
Abbreviations
- CG:
-
Central grey
- DH:
-
Dorsal horn
- inf R:
-
Inferior reticular nucleus
- Ris:
-
Isthmic reticular nucleus
- Rmed:
-
Medial reticular nucleus
- Rsup:
-
Superior reticular nucleus
- SF:
-
Solitary fasciculus
- VH:
-
Ventral horn
- 9:
-
Glossopharyngeal motor nucleus
- 10:
-
Vagus motor nucleus
- 12:
-
Hypoglossal motor nucleus
References
Akert K (1949) Der visuelle Greifreflex. Helv Physiol Acta 7: 112–134
Beevor CE (1909) Remarks on paralysis of the movement of the trunk in hemiplegia. Br Med J 1: 881–885
Comer C, Grobstein P (1981) Tactually elicited prey aquisition behavior in the frog. Rana pipiens, and a comparison with visually elicited behavior. J Comp Physiol 142: 141–150
Dacey DM, Ulinski PS (1986) Optic tectum of the eastern garter snake, Thamnophis sirtalis. I. Efferent pathways. J Comp Neurol 245: 1–28
Ewert J-P (1984) Tectal mechanisms that underlie prey-catching and avoidance behaviors of the optic tectum. In: Vanegas H (ed) Comparative neurology of the optic tectum. Plenum, New York
Ewert J-P (1987) Neurethology of releasing mechanisms: prey catching in toads. Behav Brain Sci 10: 337–405
Ewert J-P, Burghagen H, Schurg-Pfeiffer E (1983) Neuroethological analysis of innate releasing mechanism for prey-catching behavior in toads. In: Ewert J-P, Capranica RR, Ingle D (eds) Advances in vertebrate neuroethology. Plenum, New York
Graham-Brown T (1913) On the postural and non-postural activities of the midbrain. Proc R Soc B 87: 145–163
Grobstein P (1988a) Between the retinotectal projection and directed movement: topography of a sensorimotor interface. Brain Behav Evol 31: 34–48
Grobstein P (1988b) Organization in the sensorimotor interface: a case study with increased resolution. In: Ewert J-P, Arbib MA (eds) Visuomotor coordination: amphibians, comparisons, models, and robots. Plenum, New York
Grobstein P, Comer C, Kostyk S (1980) The potential binocular field and its tectal representation in Rana pipiens. J Comp Neurol 190: 175–185
Grobstein P, Comer C, Kostyk SK (1983) Frog prey capture behavior: between sensory maps and directed motor output. In: Ewert J-P, Capranica RR, Ingle D (eds) Advances in vertebrate neuroethology. Plenum, New York
Grobstein P, Masino T (1986) Sensorimotor circuitry underlying directed movement in the frog: evidence for an intermediate representation of space in the tectofugal pathways. Soc Neurosci Abstr 12: 684
Grobstein P, Reyes A, Zwanzigger L, Kostyk SK (1985) Frog prey orienting: variations in output with stimulus distance. J Comp Physiol A 156: 775–785
Grobstein P, Crowley K, Spiro J (1988) Neuronal organization for directed movement in the frog: similarities in visual and tactile prey orienting. Soc Neurosci Abstr 14: 1236
Hinsey JC, Ransom SW, Dixon HH (1930) Responses elicited by stimulation of the mesencephalic tegmentum in the cat. Arch Neurol Psychiat 24: 966–977
Ianonne AM, Gerber AM (1982) Brown-Sequard syndrome with paralysis of head turning. Ann Neurol 12: 116
Ingle D (1970) Visuomotor functions of the frog optic tectum. Brain Behav Evol 3: 57–71
Ingle D (1972) Depth vision in monocular frog. Psychon Sci 29: 37–38
Ingle D (1973) Two visual systems in the frog. Science 181: 1053–1055
Ingle D (1983) Brain mechanisms of visual localization by frogs and toads. In: Ewert J-P, Capranica RR, Ingle D (eds) Advances in vertebrate neuroethology. Plenum, New York
Ingle D, Sprague JM (1975) Sensorimotor functions of the midbrain tectum. Neurosci Res Prog Bull 13: 173–288
Kostyk SK, Grobstein P (1982) Visual orienting deficits in frogs with various unilateral lesions. Behav Brain Res 6: 379–388
Kostyk SK, Grobstein P (1987a) Neuronal organization underlying visually elicited prey orienting in the frog. I. Effects of various unilateral lesions. Neuroscience 21: 41–55
Kostyk SK, Grobstein P (1987b) Neuronal organization underlying visually elicited prey orienting in the frog. II. Anatomical studies on the laterality of central projections. Neuroscience 21: 57–82
Kostyk SK, Grobstein P (1987c) Neuronal organization underlying visually elicited prey orienting in the frog. III. Evidence for the involvement of an uncrossed descending tectofugal pathway. Neuroscience 21: 83–96
Lázár G (1969) Efferent pathways of the optic tectum in the frog. Acta Biol Acad Sci Hung 20: 171–183
Masino T, Grobstein P (1986) Sensorimotor circuitry underlying directed movement in the frog: organization of tectofugal pathways and likely involvement of the nMLF. Soc Neurosci Abstr 12: 684
Masino T, Grobstein P (1989) The organization of descending tectofugal pathways underlying orienting in the frog. Rana pipiens. II. Evidence for the involvement of a tecto-tegmento-spinal pathway. Exp Brain Res 75: 245–264
McIlwain JT (1982) Lateral spread of neural excitation during microstimulation in intermediate gray layer of cat's superior colliculus. J Neurophysiol 47: 167–178
Middlebrooks JC, Knudsen EI (1984) A neural code for auditory space in cat's superior colliculus. J Neurosci 4: 2621–2634
Nieuwenhuys R, Opdam P (1976) Structure of the brainstem. In: Llinás R, Precht W (eds) Frog neurobiology: a handbook. Springer, Berlin Heidelberg New York
Patton P, Grobstein P (1984) Forebrain involvement in preyforienting in the frog. Soc Neurosci Abstr 10: 61
Raphan T, Cohen B (1971) Brainstem mechanisms for rapid and slow eye movements. J Neurophysiol 34: 920–936
Rubinson K (1968) Projections of the tectum opticum of the frog. Brain Behav Evol 1: 529–561
Sparks DL (1986) Translation of sensory signals into commands for control of saccadic eye movements: role of primate superior colliculus. Physiol Rev 66: 118–171
Sparks DL, Holland R, Guthrie BL (1976) Size and distribution of movement fields in the monkey superior colliculus. Brain Res 113: 21–34
Wurtz RH, Albano JE (1980) Visual-motor function of the primate superior colliculus. Ann Rev Neurosci 3: 189–226
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Masino, T., Grobstein, P. The organization of descending tectofugal pathways underlying orienting in the frog, Rana pipiens . Exp Brain Res 75, 227–244 (1989). https://doi.org/10.1007/BF00247931
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00247931