
Abstract
The concept of rewilding should not only be applied to recovering habitat and vertebrate populations but also to the restoration of complex ecological processes. Large avian scavengers are the target of restoration programs including conservation measures linked to the manipulation of food resources but we lack of a general approach to understanding how scavengers and the ecosystem services they provide will fit into a rewilded Europe. Carcasses play an important role in ecosystem functioning and in the energy flux within food webs. Large ungulates carcases availability, in particular, has, through the course of evolution, given way to the appearance of “true” scavenger strategies, displayed by large body-sized avian organisms (vultures) whose guilds are structured by complex interspecific relationships. Yet, livestock raised in traditional agro-grazing systems have historically replaced wild ungulates as the main food source for vultures. More recently, modern farm intensification, stricter European Union legislation that banned the abandonment of carcasses, and increasing human-vulture conflicts contributed to plunging vulture populations, leading to an unprecedented crisis. Consequently, supplementary feeding became a management tool used worldwide to aid in the recovery of their decimated populations. These so-called vulture restaurants, however, alter the spatial-temporal nature of trophic resources with strong consequences at individual, population, community and ecosystem levels. The conservation of these charismatic species in rewilded European landscapes should rely on wild ungulate expansion, the recovery of large carnivore populations and, in more humanized areas, the promotion of traditional extensive agro-grazing systems limiting artificial feeding activities. In this way, it may be possible to combine both the historically recognized ecosystem services provided by vultures (elimination of undesirable remains, nutrient cycling) with new recreational services (conferring aesthetical value to the environment) while providing economic benefits to rural societies. Vultures and other scavengers, because they exploit space at a huge scale, are singular actors within a rewilded Europe. Their conservation, and that of the ecological processes in which they are involved, requires large-scale approaches surpassing those limits imposed by administrations, habitats and even biomes.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction: Rewilding Ecosystem Services, Not Only Vertebrate Populations
Although the concept of “rewilding” is usually associated to restoring populations of symbolic species, it is obvious that it cannot be understood without the parallel amendment to the structure and functioning of ecosystems (see Chap. 1). In this sense, there are some ecological processes transverse to habitat structure, ecosystems and biomes, which are key to maintaining both complex food webs and the viability of populations of organisms. The large avian scavengers in Europe could serve as a paradigmatic case in this regard. Due to the early and rapid transformation of European landscapes , the decline of large scavenger populations occurred prior to that on other continents (Bijleveld 1974), thus breaking the ancient alliance between traditional agro-pastoral practices and the existent large populations of these birds. However, since the late 20th century to today, it is precisely in Europe that some of the most representative and healthiest populations of vultures and other Old World avian scavengers likely reside, in comparison to other locations where a massive population declines have occurred, such as in Africa and Asia. It is also in Europe where there are likely more active recovery programmes devoted to conserving scavenger’ populations mainly based on reintroductions and renewed recognition of the ecosystem, cultural and economic services that scavengers may provide.
Thus, it may be deduced that the rewilding of Europe, specifically in the case of avian scavenger species, has already begun. This process is often accompanied by profound changes in the carrying capacity of the environment (e.g., through supplementary feeding programs). While undeniably profound changes are occurring that may make the maintenance of populations of birds of prey and the services they provide unstable, it is expected that in a few decades large areas of the European continent will abandon traditional grazing activities (see Chap. 1). On the other hand, other regions will continue the intensive occupation and use of land, which may impose potentially greater impacts on natural systems (e.g., agricultural intensification, growth of urban areas) (Deinet et al. 2013).
In this chapter, our goal is to explore, on the basis of existing information, how top scavengers fit into a wilder Europe. To do this, we first examine the role of availability of carrion resources in the maintenance of ecosystem functioning. Then, we examine the implications of the creation of supplementary feeding stations (so-called vulture restaurants). In addition, we will describe how the relationship between humans and vultures has evolved, identifying ecosystem services provided by these charismatic species from the past to the current date. Finally, we propose that the conservation of top scavenger species and the maintenance of natural complex ecological process linked to the exploitation of unpredictable carrion resources in rewilding the European landscape s should rely on favouring wild ungulates expansion, the recovery of predator populations and the promotion of traditional extensive grazing practices.
2 The Role of Carcasses Within Ecosystems
Carcasses are resources whose role is often minimized in ecological theory (DeVault 2003). Since animals can die at any place and time, a carcass is considered a “prize” for many species that obtain some benefit. From carnivores and avian scavengers to the microcosm, a large number of species, including plants, can benefit from the appearance of carcasses. Consumers can exhibit numeric and/or functional responses including behavioural changes, and increases in their reproductive rates in response to carcasses. A possible increase in the populations of consumers of carrion may trigger changes in the interactions between them and their prey, predators, competitors and parasites. In addition to these direct effects, indirect effects can occur that can elicit “cascading” effects through food chains, both bottom-up and top -down, ultimately affecting the community structure the ecosystem functioning (Cortés-Avizanda 2011; Yang et al. 2008; Fig. 5.1).

Conceptual model of the energy flow and the ecological relationships that are established with the appearance of a carcass in the wild. The arrows indicate the well-known effects of some trophic groups on the others. Here it is also presented the example of the avian scavengers. The “?” indicate hypothetical effects that have not yet been studied in detail. (Based on Selva and Cortés-Avizanda 2009)
From an abiotic point of view, it is noteworthy that the appearance of a carcass represents a key natural “disturbance” in the composition of soils and plant communities, because it means a sudden availability of nutrients (Towne 2000; Melis et al. 2007). In fact, based on the high levels recorded of soil nutrients deposit, biomass production and activity of edaphic fauna, the experts have defined the carcasses as “islands of decomposition” (Carter et al. 2007; Selva and Cortés-Avizanda 2009; Fig. 5.1) This increase in soil nutrient concentration in the vicinity of a carcass shows a gradient decreasing towards the periphery and which may persist for several years (Towne 2000; Danell et al. 2002; Melis et al. 2007). For vegetation, the changes are more drastic and can occur both in terms of biomass and community structure. The “islands of decomposition” represent a resource of high quality and low competition, favouring the establishment of pioneer species. In general, it has been well-described that one year after the occurrence of a carcass the richness, the diversity of species, the biomass and the vegetation cover can increase significantly in the vicinity of the carcass (Towne 2000). This effect depends on the region where it occurs and other numerous factors such as the climate, temperature and soil type. For example, if the carcass occurs in a homogeneous system or an unproductive one, then carcasses can represent a refuge for many plant species leading to an increase in the spatial diversity of the plant community (Towne 2000; Danell et al. 2002; Selva and Cortés-Avizanda 2009).
Relative to direct consumers, it has been reported that the diversity and complexity of microorganisms and invertebrates exploiting dead animals can acquire an extraordinary value (Sikes 1994). More than 500 species of arthropods (and of these, 422 species of insects) have been described in a carcass during different stages of decomposition (Payne 1965), varying by geographic region and environmental conditions (Amendt et al. 2004; Selva and Cortés-Avizanda 2009; Fig. 5.1). Many organisms show a relative specialization in relation to the type of carrion and its decomposition phase. Those organisms may respond to carcass appearance and further trophic cascades may elicit (Payne 1965; Amendt et al. 2004; and see examples in Selva and Cortés-Avizanda 2009). Among vertebrates, carcasses may be consumed by over 30 species of birds and mammals (Houston, 1979; DeVault et al. 2003; Selva 2004; Selva and Fortuna 2007). Most of these organisms are facultative scavengers , which are opportunistic carrion eaters, especially when their main prey become scarce. For instance, it is well-known that in temperate forests during harsh wintering conditions (low temperatures, thick snow cover and low availability of small mammals) predators significantly increase feeding on carcasses (Heinrich 1988; Jedrzejewski et al. 1993; Selva 2004; Selva et al. 2003, 2005; Cortés-Avizanda et al. 2009a).
Despite the importance of these many scavenger species, the avian scavengers are the organisms that have attracted the most attention because of their spectacular nature and their close relationship with humans as effective providers of ecosystem services (see below). These species have evolved different behavioural skills and morphology, allowing their coexistence through the sharing of trophic resources (Kruuk 1967; Root 1967; König 1983; Hertel 1994; Hertel and Lehman 1998; Blondel 2003; Cortés-Avizanda et al 2014; Fig. 5.2). The functioning of the guild is driven by both positive ecological processes (facilitation) and the most obvious competitive (negative) interspecific relationships (Donázar 1993; Cortés-Avizanda et al. 2012). Facilitatory processes have been proposed to follow two opposite paths within this guild: small-body-sized facultative scavengers landing earlier at carcasses can increase the chances of carcass detection by larger (specialist) vultures (local enhancement) whereas large vultures dismember the carcass thus allowing smaller scavengers to profit from the resource (trophic advantage; Kruuk 1967; König 1974, 1983; and see details in Cortés-Avizanda et al. 2012). As a consequence of this efficient process, carcasses of medium-sized animals are consumed in a very short time (Selva 2004; Cortés-Avizanda et al. 2012; Cortés-Avizanda et al 2014 ).

a) Egyptian vulture (Neophron percnopterus) and b)Black kite (Milvus migrans) in the Mauritanian Sahel. In Africa the availability of trophic resources is not limiting and therefore scavengers show a homogeneous spatial distribution. (Cortés-Avizanda et al. 2011;Photo credit: Jose Ramón Benitez)
3 Vultures and Humans: An Unstable Alliance
Top scavengers (“true” vultures, Accipitridae) have been evolutionarily dependent on carcasses of large animals, mainly ungulates, grazing in open areas of southern Europe, Africa and central and southern Asia (Houston 1974, 1979; Donázar 1993). They have in common extreme adaptations and skills aimed to locate scarce and unpredictable sources of food (Houston 1979; Cortés-Avizanda et al. 2014). However, because of the rampant humanization of ecosystems, this natural scenario no longer exists, apart from some strictly protected areas, especially in African countries where large herds of ungulates subsist (Cortés-Avizanda et al. 2011; Fig. 5.2). Thus, following the progressive eradication of most native populations of wild ungulates (e.g. Chap. 8), guilds of avian scavengers have become largely dependent on livestock carcasses associated with human activities (Mundy et al. 1992; Donázar 1993). This scenario has likely remained almost unchanged since the so-called “Neolitic revolution” (i.e., a process in which agrarian societies began to substitute hunting around 8500 BC with agricultural practices including the domestication of the herbivores) in many regions of the Old World, and certainly in southern Europe, where the agro-grazing traditional economies remained unchanged (Donázar et al. 1996a, 2009; Olea and Mateo-Tomas 2009; Fig. 5.3).

Avian scavengers and traditional extensive grazing. After the eradication of native populations of wild ungulates, avian scavengers have become dependent of traditional livestock carcasses. (Photo credit: Iosu Anton)
After the industrial revolution this scenario changed dramatically. During the 19th and the first half of the 20th century, a utilitarian view of nature became common. “Harmful” species were more efficiently prosecuted and raptor populations, including scavengers, declined steeply across Europe and in many regions to full extinction (Bijleveld 1974). In parallel, a key resource for carrion-eaters, the wild ungulates, were also decimated by hunting and poaching (the main decline of ungulate populations in Europe took place during the last centuries after the generalized use of firearms), disappearing from most of the European landscapes (see e.g. Chapman and Buck 1910; Deinet et al. 2013 and references therein). Carcasses of domestic ungulates were probably still plentiful until well into the 20th century but the transformation of farming to intensive practices and the abandonment of extensive grazing reduced their availability in the last decades (Donázar et al. 1996a). This dramatic scenario began to improve from the 1960s onwards. Given the high public profile of many large avian scavengers, local administrations and conservationist groups created a number of feeding stations (also known as “vulture restaurants”; Fig. 5.4) to supply food and help re-establish the decimated populations of these species (Bijleveld 1974 and see below; Fig. 5.4). But perhaps much more importantly, legal protection curbed non-natural mortality allowing scavenger populations to quickly recover, mainly in the Iberian Peninsula and Southern France (Donázar and Fernández 1990; Donázar et al. 1996a, b; Slotta-Bachmayr et al. 2004). The griffon vulture (Gyps fulvus) was the most favoured species, its populations having dramatically increased in numbers in the Iberian Peninsula and France (BirdLife International 2004). Populations of Cinereous vulture (Aegypius monachus) and Bearded vulture (Gypaetus barbatus) have also increased in numbers, albeit more moderately, in some European regions (Margalida and Heredia 2005; Moreno-Opo 2007; Dobado et al. 2012). By contrast, the numbers of small-sized scavengers such as Egyptian vultures (Neophron percnopterus) and Red (Milvus milvus) and Black (Milvus migrans) kites continue to decline (Viñuela et al. 1999; Del Moral and Martí 2001, 2002).

Supplementary feeding stations devoted for vultures. The supplementary feeding stations are a worldwide conservation tool to recover endangered populations of avian scavengers. Usually they are fenced sites where local farmers and rangers dispose the carcasses. a: A vulture restaurant devoted to Egyptian vulture in Fuerteventura (Canary Island) where the dominant griffons are not presented. b: A group of Egyptian vulture feeding at a vulture restaurant also in Fuerteventura. (Photo credit : a) José Antonio Donázar and b) Manuel de la Riva)
A decade ago the appearance of the Bovine Spongiform Encephalopatie (BSE) changed the picture abruptly. New sanitary regulations driven by the European Union banned the abandonment of livestock carcasses in the field, and consequently, the availability of food resources declined in some regions by more than 80 %. The consequences for vulture populations are still being evaluated, but spatial distribution, breeding success and survival seem to have been affected. This is particularly noticeable in those species with greater dependence on carcasses of large animals such as griffon vultures (see reviews in Donázar et al. 2009a). To complicate things even more, after these sanitary measures a conflict between farmers and vultures developed in Mediterranean regions (particularly Spain and southern France). Griffon vultures are known to occasionally kill and consume diminished livestock but the number of cases reported increased sharply after the implementation of sanitary regulations (Margalida et al. 2014). No doubt that this trend was largely the result of a social contagion driven by misinformed media, but regardless, a true conflict arose compromising decades of conservation measures aimed to restore avian scavenger populations. The consequence was that hundreds of individuals of griffon and other species of vultures and facultative scavengers perished by poison, while public opinion called for a quick fix consisting of artificially feeding vultures to distract their attention from live prey (Margalida et al. 2010).
After these events and due to the widespread pressures coming from researchers, policy conservation managers and farmers, recent European regulations have increasingly allowed the disposal of livestock carcasses for consumption by vultures (Donázar et al. 2009a, b; Margalida et al. 2010). The adoption of this new legislation takes time, especially for the transposition of EU laws to individual country governments and then in many cases from the country level to local government departments. In general, the legal framework is cumbersome such that administrations are faced with complex and time-intensive processes. Consequently, the European regional governments (e.g. in Spain), have adopted a common strategy whereby just a few widespread sites -in theory under strict veterinary control- are supplied with carcasses for scavengers . Thus, vultures have access to restaurant networks with large amounts of food (Cortés-Avizanda et al. 2010, 2012). The effects of this conservation strategy, which is not new and has now become widespread, may be substantial from both population and ecological points of view.
4 Vulture Restaurants and the Loss of a Pulsed Resource
Supplementary feeding stations or vulture restaurants have been considered for decades to be a key management tool for the conservation of scavenger bird populations (Bijleveld 1974; Houston 1987; Piper 2006). Species managed by supplementary feeding include: the California Condor (Gymnogyps californianus) in Western North America; the King Vulture (Sacorramphus papa) in Belize; and the Cape Vulture (Gyps coprotheres) and bearded vulture in Southern Africa (Wilbur and Jackson 1983; Houston 1987; Brown 1990; Mundy et al. 1992). In Eurasia, vulture restaurants have mainly targeted populations of bearded, Egyptian, griffon, and cinereous vultures (Donázar 1993; Donázar et al. 2009a, 2010; Cortés-Avizanda et al. 2010; Fig. 5.4). Supplementary feeding stations have also been considered as key tools in the recovery of the critically endangered Asian vulture populations after the catastrophic declines caused by the veterinary treatment of livestock with diclofenac (i.e., NSAID, a non-steroidal anti-inflammatory drug, see Gilbert et al. 2007; Prakash et al. 2012 and references therein). In general, it is assumed that this management tool provides a number of benefits for the conservation of the target species: an increase of food availability, reduction of the risk of poisoning and persecution, and an increase in the availability of micronutrients (calcium) (Piper 2006).
Supplementary feeding stations, however, change the spatio-temporal distribution of a food resource that is otherwise unpredictable and ephemeral. As we have described above, in natural systems, carcasses resemble other trophic pulsed resources such as tree-masting or insect explosions (Ostfeld and Keesing 2000; Rose and Polis 1998). Currently, however, carcasses have become predictable and clumped via this human intervention and the consequences of this deep alteration of habitat quality have been largely ignored, under the assumption that only positive effects would occur. We are now increasingly aware that these positive effects exist but that there are also other negative effects that may override the positive conservation effects on the target species. For instance, it has been demonstrated that proximity to vulture restaurants promotes communal roosts and long-term territory maintenance in the Egyptian vulture increasing the probability of frequent visits by breeding adults from nearby territories (Grande et al. 2009; Benítez et al. 2009; García-Heras et al. 2013; Lopez-Lopez 2014). In the case of the Pyrenean bearded vultures, supplementary feeding stations have led to high rates of immature survival thus dampening the effects of indirect persecution (poisoning) and increasing population viability (Oro et al. 2008). Conversely, negative population effects are also apparent. Negative density-dependent decreases of productivity and appearance of unusual breeding units (polyandrous trios) have been detected in the vicinity of bearded vulture feeding places (Carrete et al. 2006a, b). Unwanted effects have also been detected in the structure and functioning of guilds . Vulture restaurants favour the gathering of individuals of the dominant species (e.g., the griffons), which monopolize food to the detriment of small and less competitive and often more endangered scavenger species (Cortés-Avizanda et al. 2010; 2012). The predictable nature (in space and time) of carrion disposed in these places disrupts ecological processes provoking a reduction of guild diversity and the loss of intraguild facilitatory processes because dominant specialist species (griffons) arrive early and in larger numbers (Cortés-Avizanda et al. 2012). Less competitive species may congregate at feeding places but they may be unsuccessful at feeding, whereby the feeding place would become an ecological trap (Cortés-Avizanda et al. 2012 and authors’ unpublished data; Fig. 5.5).

Randomly distributed vs. predictable resources. Unpredictable trophic resources allow the occurrence of facilitatory processes promoting the biodiversity and the coexistence of species within an Old World avian scavenger guild: E.g. a: A Bearded vulture (Gypaetus barbatus) sharing a carcass with a common raven (Corvus corax) and b: A griffon vulture (Gyps fulvus) sharing a carcass with two Egyptian vultures (Neophron percnopterus). c: Group of griffon vultures at supplementary feeding stations. Large amount of carrion clumped at these predictable sites favours the aggregations of hundreds of individuals (Photo Credit: Jordi Bas (a) and Antonio Atienza (b) and c)))
Finally, the aggregation of food resources at scavenger feeding places may have consequences on non-scavenging species, permeating to other ecological levels. Facultative species consume carcasses less efficiently and more slowly than the specialists or strict carrion-eaters and most importantly, they do not feed only on carcasses but also rely on small prey. Therefore, predation pressure can increase on passerine species breeding in the vicinities of vulture restaurants and carcass accumulations can change the spatial distribution of herbivore mammals (Cortés-Avizanda et al. 2009a, b; Wilmers et al. 2003). This phenomenon may be pronounced at high latitudes, where cold temperatures during long winters slow the activity of microorganisms and invertebrates and where there is a lack of specialist carrion-eaters that would otherwise quickly deplete the carrion (Selva and Cortés-Avizanda 2009; Cortés-Avizanda 2011; Cortés-Avizanda et al. 2009a).
5 How do Vultures Fit into a Rewilding Continent?
According to Navarro and Pereira (see Chap. 1), the decline in the number of extensive livestock in Europe was 25 % between 1990 and 2010. From this assessment two interpretations are possible: on the one hand, the abandonment of traditional grazing involves a significant reduction in livestock and domestic food sources for vultures, but on the other hand, the landscape abandonment may contribute positively to the expansion of wild ungulate populations in many rural areas of Europe, notably in mountain ranges. Blázquez-Alvarez and Sánchez-Zapata (2009) showed that the number of wild ungulates hunted in Spain went from 60,000 in around 1980 to 200,000 in 2005. The current area of distribution occupied by wild ungulate species has also spread (e.g., 70–75 % of the Spanish territory, Sánchez-Zapata et al. 2010) such that up to six different species of ungulates can be found in some mountain areas. Similar trends can be found in other Mediterranean and temperate regions of Europe (Milner et al. 2006).
Increasingly larger populations of wild ungulates are allowing a return to natural diets of specialists and facultative scavenger species (see Moreno-Opo et al. 2007; Sánchez-Zapata et al. 2010). The availability of wild ungulate carcasses for vultures would increase even more if future rewilding processes lead to the expansion of large carnivores which would contribute a regular supply of random carcasses (Selva 2004; Blázquez-Alvarez and Sánchez-Zapata 2009). More important than the global availability of carcasses may be the ways in which the existence of large carnivores can change the temporal and spatial distribution of the resource and the associated consequences. It has been argued that predation can add stability to trophic networks buffering those oscillations linked to temporally-pulsed events of carrion availability such as those determined by climatic events, diseases and hunter-kills (Wilmers and Getz 2005; Wilmers and Post 2006). Currently, and because of persecution, wolves in European landscapes are restricted to forests and mountains, and therefore most of the species benefiting from predated animals are forest-living facultative scavengers (Selva 2004; Selva et al. 2003; 2005). It is expected, however, that in a more relaxed scenario carcasses of killed ungulates would also be available in open biomes, thus being available to large avian scavengers, and restoring the stability and the insurance of food chains and ecological processes (Wilmers and Getz 2005; Tylianakis et al. 2010).
A key question arises from a scenario of substitution of domestic by wild ungulates : can vulture populations survive in a future wilder scenario where most of the carcasses are provided by wild herbivores? Recent research performed in North-Eastern Spain by Margalida et al. (2011) based on the application of bio-inspired computational models revealed that this may be the case for high mountain areas where large and diverse populations of wild ungulates subsist. On the contrary, in low-altitude areas with higher humanization densities, wild herbivores are scarce and their populations would be insufficient to guarantee the long-term maintenance of avian scavenger populations such as those of griffon, Egyptian and bearded vultures. Therefore, these authors also suggest that in lowland areas of European Mediterranean regions, carrion-eaters will still be dependent on resources provided by traditional extensive livestock and/or those supplies offered by conservation managers in vulture restaurants. From these results arise new questions for future research about how extensively rewilding should be facilitated, especially in those low-altitude humanized areas, in order to decrease vulture dependence on networks of large and fixed supplementary feeding stations.
The consumption of wild ungulate carcasses derived from hunting is not exempt of risk for avian scavengers . Birds ingesting pieces of hunting ammunition are exposed to lead intoxication (see review in Fisher et al. 2006; Mateo et al. 2007) to the extent that its incidence can be a serious threat to large-scale reintroduction projects such as that of the California condor in North America (Finkenlstein et al. (2012 and references therein). Lead levels are also very high in Europe, for instance Iberian griffon vultures show seasonal and spatial variations according to the rate of consumption of wild ungulates (García-Fernández et al. 2005). Moreover, carcasses resulting from hunting activities frequently accumulate at a few points at the end of hunting activities (Wilmers et al. 2003), which mimics the predictability offered by vulture restaurants and may result in similar negative effects (see above). Under a future scenario of a wilder Europe, we may promote two lines of action to curb the poisoning of scavengers: the consideration of a wilder Europe with zones limiting or lacking in hunting activities and in parallel the encouragement of the traditional extensive grazing currently under decline.
6 New Services Provided by Vultures
Although there is a strong competition “between” and “within” guilds (bacteria, invertebrates, carnivores, birds) for pulsed carrion resources (see reviews of Root 1967; Jaksic 1981; Schluter and Ricklefs 1993; Blondel 2003), this does not diminish the fact that potentially harmful species (like rats and feral dogs) prosper with local food abundance (Markandya et al. 2008), carrying potentially infectious diseases (Blount et al. 2003). Therefore, a historical scavengers/humans relationship was built, resulting in a sort of mutualism: humans provide trophic resources and vultures eliminate undesirable remains and control the concentration of disease (Deygout et al. 2009; Margalida et al. 2011; Cortés-Avizanda et al. 2012). Currently, and because both sanitary regulations impose the efficient elimination of livestock remains, and modern societies require that industrial activities are performed in a way that minimizes their ecological footprint, these services have acquired a new dimension: vultures can remove carcasses at zero cost whereas the elimination of livestock carcasses by means of industrial procedures entails high expenditures (transport: 20 €/animal; destruction: 96 €/t; see Donázar et al. 2009a, b). Moreover, the CO2 emissions derived from the transport and burning of carcasses are not negligible. Consequently, modern farming economies may still find vultures to be useful allies in sustaining traditional uses of Mediterranean landscapes (Deygout et al. 2009).
In modern societies, vultures, like other large birds, are highly attractive wildlife, providing several recreational services: the delight experienced from the beauty of a vulture in flight or soaring in large groups, or the observation of feeding behaviour and interactions, among other, bring great value to the environment (Fig. 5.6). These services may be translated into significant incomes derived from “ecotourism” (see Chap. 3). For instance, it is estimated that visitors for griffon vulture watching in Israel provides around US$ 1.1–1.2 million per year (Becker et al. 2005) and the park “La falaise aux vautours” in the French Pyrenees receives 15,000–20,000 visitors per year whose primary motivation is wildlife viewing activities. Overall, the observation of scavenger breeding areas and vulture restaurants is an increasingly common activity within specialized touristic tours. This can improve the income of those southern European rural societies subject to profound environmental and socioeconomic changes.

Cultural services provided by birds benefit local communities. The delight with the beauty of vultures alone or soaring in large group at their breeding areas and/or feeding at vulture restaurants confer a value to environment, and attract birdwatchers from all over the world translating this cultural service into significant incomes via “ecoturism”. (Photo credit: Jordi Bas)
To summarize, we can conclude that scavenging directly contributes to human well-being . Avian scavengers provide regulation/maintenance and cultural services due to their valuable role in the decomposition of carcasses, pest control, biodiversity maintenance, and tourism attraction (Haines-Young and Potschin 2013; Maes et al. 2013)
7 Discussion and Conclusion
Land abandonment is widespread in many European areas with poor agricultural development. In the face of this situation, the rewilding of abandoned landscapes (defined as the passive management of ecological succession, see Chap. 1; Fig. 5.7) is viewed as an opportunity to recover native biodiversity and ecological processes and provide a range of ecosystem services (Cramer et al. 2008; and see Chaps. 1, 3). In contrast, active management is aimed at the maintenance of low-intensity agriculture in order to conserve specific organisms and particular habitats, often working against succession processes (e.g. Pain and Pienkowski 1997). This dichotomy is subject to a heated debate with clear implications for European agricultural policies (see details in Merckx and Pereira in press). Within this scenario, the conservation of top scavengers, not only as charismatic species but also as key actors in complex ecological processes, imposes particular challenges. Vultures, as well as a panoply of facultative predatory-scavenger vertebrates, arthropods, and microorganisms, depend largely on the existence of ungulate and medium-size vertebrate carcasses, a resource that in natural conditions is unpredictable in space and time (i.e., by chance, see above) . The importance of ecological processes linked to carcass decomposition and consumption has been historically neglected (DeVault 2003). Instead, carcasses have been considered undesirable and insanitary residuals whose common destination has been industrial destruction. Consequently, a key resource in European ecosystem functioning was sent to the incinerator following sanitary regulations dictated by an exaggerated precautionary principle (Donázar 2009b). Rewilding may now favour the increase of larger populations of wild ungulates as well as the expansion of large carnivores (see Chap. 4), which would contribute to the random nature of carcasses and uphold ecosystem functioning and community structures.

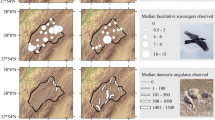
Hotspots of abandonment and rewilding and distribution of the most important breeding areas for top scavenger: Giffon vultures and the endangered Egyptian, bearded and cinereous vultures. The map in the centre, relative to rewilding, shows areas categorized as “agriculture” in 2000 that are projected to become rewilded or afforested in 2030 with the CLUE model (see Chap. 1 for mapping method). The maps showing the scavenger distributions are based on information available in the Spanish Atlas of Breeding Birds (Del Moral and Martí 2004). (Species drawings: Juan Varela)
Other distinctive aspects of top scavengers , and vultures in particular, is that these species exploit landscapes at a scale much larger than humans do. Vultures’ home ranges and foraging displacements may cover as many as hundreds of thousands of hectares. For instance, modern monitoring techniques based on GPS and satellite tracking have demonstrated continental-scale movements of individual birds (e.g. bearded vultures in Margalida et al. 2013; Egyptian vultures in Carrete et al. 2012; among others). This is the result of adaptations to searching for unpredictable resources (Hertel 1994; Cortés-Avizanda et al. 2014), originally ungulate herds moving across changing landscapes (Houston 1974), something that still occurs in modern Europe with transhumant livestock husbandries (i.e., movement of livestock between winter and summer pastures, Olea and Mateo 2009), but on a very limited basis today. Thus, vultures have space requirements that can scarcely be reduced to the small-scale of administrative limits prevailing in European landscapes . In this context the question that arises is how can we manage to fit vultures into the projected rewilded Europe? As mentioned above, recent studies related to vulture populations in the Spanish Pyrenees show that carcasses of wild ungulates are able to maintain vulture populations in the long-term (Margalida et al. 2011 and see above for details). However, the expansion of wild herbivores into the abandoned lands of many European mountains is already occurring and likely to increase in the future (see Chap. 8). However, to date, according to Margalida et al. (2011), although avian scavengers make large-scale seasonal movements from mountains to lowlands the recovery of wild ungulates in mountain areas would be insufficient to support the populations. At best, it is evident that the long-term viability of vulture populations restrained to mountain areas would be affected by constraints derived from small population size (Margalida et al. 2011; Donázar et al. 2009a). On the other hand, many healthy vulture populations thrive in very diverse lowland agro-grazing systems where most of the carcasses are provided by extensive livestock and wild medium-sized prey (notably wild rabbits Oryctolagus cuniculus) (Donázar et al. 1996b; Moreno-Opo et al. 2010; Carrete et al. 2007). Therefore, under the scenario of a future wilder Europe and in conjunction with the expansion of wild ungulates and predators, the encouragement of traditional extensive grazing should be prioritised, especially in those lowland humanized areas of agro-grazing systems where arrival of wild ungulates may be no possible.
Overall, the conservation of vultures and other avian scavengers requires a necessary equilibrium between the recovery of wild ungulates in remote (and rewilded) areas, the existence of traditional agro-grazing systems in lowland regions and, the modifications/adaptations of sanitary laws in order to allow the abandonment of livestock carcasses freely in the wild (even outside of protected and defined areas). Recent European regulations are opening the way for this approach and within this context, the maintenance of supplementary feeding stations (vulture restaurants) is not a desirable conservation strategy because of the negative population, community and ecosystem effects that appear to surpass the positive effects linked to the mere improvement of demographic parameters (see above for references). Therefore, this scenario represents a great opportunity to reclaim future conservation strategies considering community and ecosystem perspectives. New policies should focus on: (1) the conservation of foraging and breeding behaviour of hundreds of scavenger as well as the ecosystem restoring by promoting the availability of natural random carcasses; (2) highlighting the importance of the regulating services derived from those feeding behaviours such as the reduction of transmission of animal diseases; (3) the new ecosystem services not related directly to species conservation but that may imply greater incomes to local economies from ecotourism and associated with recreational activities in natural areas; and (4) the consideration of non-economic value that these species may provide such as the existence values and use them to educate society on conservation.
The scenario described thus far applies to those regions of Western Europe (the Iberian Peninsula, Southern France and other Mediterranean regions) where large scavenger populations have persisted (Donázar et al. 2009a). A totally different picture exists in central and Eastern Europe, where the populations of vultures and other carrion-eaters were virtually extirpated during the course of the twentieth century. There, passive rewilding of large regions probably will not lead to short or medium-term recovery of top scavenger/vulture populations and the associated scavenging processes because of the extremely low rate of spatial expansion of populations of these long-lived organisms that is explained by extreme natal philopatry. In this case, active measures like reintroduction are required and of course, the elimination of those limiting factors, mainly direct and indirect persecution, that once determined the populations’ demise. Rewilding also considers the reintroduction of species and initial support with supplementary feeding stations. In this case, we suggest that those programs be based on rigorous scientific population viability studies and that food supplies, if necessary, are provided under careful adaptive management for greater effectiveness of rewilding conservation decisions (Cortés-Avizanda et al. 2010; McCarthy and Possingham 2007; Possingham et al. 2001)
Finally, while many mammal species and other vertebrates such as large ungulates and carnivores, may be viable in large mountain areas subjected to passive management (Fig. 5.7; and see details in Chap. 1), populations of flying organisms (covering large distances on a daily basis) would nevertheless require broader approaches dealing with both conservation aims and common policies in rewilded regions and in areas where traditional agro-grazing activities are maintained. Moreover, it must not be forgotten that top scavenger breeding in remote cliffs are providing key ecological services not only in high-mountain pasturelands but also in lowland grazing areas. These reflections suggest that new questions may arise along with new challenges, especially those related to how the maintenance of ecologically functional populations of large body-sized and long-lived organisms fits within the current rewilding concept (see also Chaps. 4 and 8). To attend to these targets and requirements effectively is to ensure the persistence of the alliance between humans and vultures that has allowed the survival of these charismatic birds for millennia.
Acknowledgments
We wish to thank Jose Antonio Sánchez Zapata, Roger Jovani, and Nuria Selva who kindly discussed with us some of the ideas presented here and provided helpful comments on the earlier versions of the chapter. ACA was funded by a PostDoc grant from the Fundação para a Ciência e a Tecnologia (FCT) (SFRH/BPD/91609/2012), and JAD by the project CGL2012-40013-C02-01. Isa Afan from the Laboratorio de SIG y Teledetección, Estación Biológica de Doñana, CSIC (LAST-EBD), built the maps of species distribution. We also thank the photographers, Jose Ramón Benitez, Iosu Anton, Jordi Bas, and Antonio Atienza, Manuel de la Riva for their contribution to this chapter.
References
Amendt, J., Krettek, R., & Zehner, R. (2004). Forensic entomology. Die Naturwissenschaften, 91, 51–65.
Becker, N., Inbar, M., Bahat, O., Choresh, Y., Ben-Noon, G., & Yaffe, O. (2005). Estimating the economic value of viewing griffon vultures Gyps fulvus: A travel cost model study at Gamla Nature Reserve, Israel. Oryx, 39, 429–434.
Benítez, J. R., Cortés-Avizanda, A., Ávila, E., & García, R. (2009). Effects of the creation of a supplementary feeding station for the conservation of Egypttian vulture (Neophron percnopterus) population in Andalucia (southern Spain), in: Donázar, J.A., Margalida, A. & Campión, D. (Eds.),Vultures, feeding stations and sanitary legislation: a conflict and its consequences from the perspective of conservation biology. Sociedad de Ciencias Aranzadi San Sebastián, Spain, pp. 276–291.
Bijleveld, M. (1974). Birds of prey in Europe. London: Macmillan Press.
BirdLife International (2004). Birds in the European Union: A status assessment. BirdLife International. Wageningen, The Netherlands.
Blázquez-Alvarez, M., & Sánchez-Zapata, J. A. (2009). The role of wild ungulates as a resource for the community of vertebrate scavengers. In J. A. Donázar, A. Margalida, & D. Campión,(Eds.). Vultures, feeding stations and sanitary legislation: A conflict and its consequences from the perspective of conservation biology (pp. 308–327). Munibe 29 (suppl.), Sociedad de Ciencias Aranzadi San Sebastián, Spain.
Blondel, J. (2003). Guilds or functional groups: Does it matter? Oikos, 100, 223–231.
Blount, J. D., Houston, D. C., Moller, A. P., & Wright, J. (2003). Do individual branches of immune defence correlate? A comparative case study of scavenging and non-scavenging birds. Oikos, 102, 340–350.
Brown, C. J. (1990). An evaluation of supplementary feeding for Bearded Vultures and other avian scavengers in the Natal Drakensberg. Lammergeier, 41, 30–36.
Carrete, M., Donázar, J. A., & Margalida, A. (2006a). Density-dependent productivity depression in Pyrenean Bearded Vultures: Implications for conservation plans. Ecological Applications, 16, 1674–1682.
Carrete, M., Donázar, J. A., Margalida, A., & Bertran, J. (2006b). Linking ecology, behaviour and conservation: Does habitat saturation change the mating system of bearded vultures? Biology Letters, 2, 624–627.
Carrete, M., Grande, J. M., Tella, J. L., Sánchez-Zapata, J. A., Donázar, J. A., Díaz-Delgado, R., & Romo, A. (2007). Habitat, human pressure, and social behaviour: Partialling out factors affecting territory extinction in the Egyptian vulture. Biological Conservation, 136, 143–154.
Carrete, M., Bortolotti, G. R., Sánchez-Zapata, J. A., Delgado, A., Cortés-Avizanda, A., Grande, J. M., & Donázar, J. A. (2012). Stressful conditions experienced by endangered Egyptian vultures on African wintering areas. Animal Conservation, 16, 353–358.
Carter, D. O., Yellowlees, D., & Tibbett, M. (2007). Cadaver decomposition in terrestrial ecosystems. Die Naturwissenschaften, 94, 12–24.
Chapman, A., & Buck, G. (1910). Unexplored Spain. London: Arnold.
Cortés-Avizanda, A., Selva, N., Carrete, M., & Donázar, J. A. (2009a). Effects of carrion resources on herbivore spatial distribution are mediated by facultative scavengers. Basic and Applied Ecology, 10, 265–272.
Cortés-Avizanda, A., Carrete, M., Serrano, D., & Donázar, J. A. (2009b). Carcasses increase the probability of predation of ground nesting birds: A caveat regarding the conservation value of vulture restaurants. Animal Conservation, 12, 85–88.
Cortés-Avizanda, A., Donázar, J. A., & Carrete, M. (2010). Managing supplementary feeding for avian scavengers: Guidelines for optimal design using ecological criteria. Biological Conservation, 143, 1707–1715.
Cortés-Avizanda, A. (2011). The ecological and conservation effects of trophic resource predictability: carcasses and vertebrate communities. PhD Thesis. Universidad Autónoma Madrid, Spain.
Cortés-Avizanda, A., Jovani, R., Carrete, M., & Donázar, J. A. (2012). Resource unpredictability promotes species diversity and coexistence in an avian scavenger guild: A field experiment. Ecology, 93, 2570–2579.
Cortés-Avizanda, A., Jovani, R., Donázar, J. A. & Grimm, V. (2014). Birds sky networks: How do avian scavengers search for carrion resource. Ecology, 95, 1799–1808.
Cramer, V. A., Hobbs, R. J., & Standish, R. J. (2008). What’s new about old fields? Land abandonment and ecosystem assembly. Trends in Ecology & Evolution, 23, 104–112.
Danell, K., Berteaux, D., & Braathen, K. A. (2002). Effect of muskox carcasses on nitrogen concentration in tundra vegetation. Arctic, 55, 389–392.
Deinet, S., Ieronymidou, C., McRae, L., Burfield, I. J., Foppen, R. P., Collen, B., & Böhm, M. (2013). Wildlife comeback in Europe: The recovery of selected mammal and bird species. Final report to Rewilding Europe by ZSL, BirdLife International and the European Bird Census Council. London: ZSL.
Del Moral, J. C., & Martí, R., (2001). El Buitre leonado en la Península Ibérica. Monografía nº7. SEO/BidLife, Madrid.
Del Moral, J. C., & Martí, R. (2002). El Alimoche en España y Portugal. Monografía nº8. SEO/BidLife, Madrid.
Del Moral, J. C., & Martí, R. (2004). Atlas de aves reproductoras de España. Dirección General de Conservación de la Naturaleza-SEO. Madrid. Spain.
DeVault, T. L., Rhodes, O. E., & Shivik, J. A. (2003). Scavenging by vertebrates: Behavioral, ecological, andevolutionary perspectives on an important energy transferpathway in terrestrial ecosystems. Oikos, 102, 225–234.
Deygout, C., Gault, A., Sarrazin, F., & Bessa-Gomes, C. (2009). Modeling the impact of feeding stations on vulture scavenging service efficiency. Ecological Modelling, 220, 1826–1835.
Dobado, P. M., Díaz, F. J., Díaz-Portero, M. A., García, L., Luque, E., Martín, J., Martínez, P., & Arenas, R. M. (2012). El buitre negro Aegypius monachus en Andalucía (España). In P. M. Dobado & R. M. Arenas (Eds.), The Black Vulture: Status, conservation and studies (pp. 13–37). Consejería de Medio Ambiente de la Junta de Andalucía, Sevilla, Spain.
Donázar, J. A. (1993). Los buitres ibéricos. In J. M. Reyero (Ed.), Biología y Conservación. Madrid, Spain.
Donázar, J. A., & Fernández, C. (1990). Population trends of Griffon Vultures (Gyps fulvus) in northern Spain between 1969 and 1989 in relation to conservation measures. Biological Conservation, 53, 83–91.
Donázar, J. A., Naveso, M. A., Tella, J. L., & Campión, D. (1996a) Extensive grazing and raptors in Spain. In D. J. Pain & M. W. Pienkowski (Eds.), Farming and birds in Europe (pp. 117–149). Cambridge: Academic.
Donázar, J. A., Ceballos, O., & Tella, J. L. (1996b). Communal roost of Egyptian vultures (Neophron percnopterus): dynamics and implications for the species conservation. In J. Muntaner, & J. Mayol (Eds.), Biology and conservation of mediterranean raptors (1994) (pp. 189–202). SEO/BirdLife, Monography 4, Madrid.
Donázar, J. A., Margalida, A., & Campión, D. (2009a). Vultures, feeding stations and sanitary legislation: A conflict and its consequences from the perspective of conservation biology. Sociedad de Ciencias Aranzadi San Sebastián, Spain.
Donázar, J. A., Margalida, A., Carrete, M., & Sánchez-Zapata, J. A. (2009b). Too sanitary for vultures. Science, 326, 664.
Donázar, J. A., Cortés-Avizanda, A., & Carrete. M. (2010). Dietary shifts in two vultures after the demise of supplementary feeding stations: Consequences of the EU sanitary legislation. European Journal of Wildlife Research, 56, 613–621.
Finkelstein, M. E., Doak, D. F., George, D., Burnett, J., Brandt, J., Church, M., Grantham, J., & Smith, D. R. (2012). Lead poisoning and the deceptive recovery of the critically endangered California Condor. Proceedings of the National Academy of Sciences of the United States of America, 109, 11449–11454.
Fisher, I. J., Pain, D. J., & Thomas, V. G. (2006). A review of lead poisoning from ammunition sources in terrestrial birds. Biological Conservation, 131, 421–432.
García-Fernández, A. J., Martinez-Lopez, E., Romero, D., Maria-Mojica, P., Godino, A., & Jimenez, P. (2005). High levels of blood lead in griffon vultures (Gyps fulvus) from Cazorla Natural Park (southern Spain). Environmental Toxicology, 20, 459–463.
García-Heras, S., Cortés-Avizanda, A., & Donázar, J. A. (2013). Who are we feeding? Asymmetric individual use of surplus food resources in an insular population of the endangered Egyptian vulture Neophron percnopterus. PLoS ONE, 8, e80523.
Gilbert, M., Watson, R. T., Ahmed, S., Asim, M., & Johnson, J. A. (2007). Vulture restaurants and their role in reducing diclofenac exposure in Asian vultures. Bird Conservation International, 17, 63–77.
Grande, J. M., Serrano, D., Tavecchia, G., Carrete, M., Ceballos, O., Díaz-Delgado, R., Tella, J. L., & Donázar, J. A. (2009). Survival in a long-lived territorial migrant: Effects of life-history traits and ecological conditions in wintering and breeding areas. Oikos, 118, 580–590.
Haines-Young, R., & Potschin, M. (2013).Common International Classification of Ecosystem Services (CICES): Consultation on Version 4, August-December 2012. EEA Framework Contract No EEA/IEA/09/003.
Heinrich, B. (1988). Winter foraging at carcasses by three sympatric corvids, with emphasis on recruitment by the raven, (Corvus corax). Behavioral Ecology and Sociobiology, 23, 141–156.
Hertel, F. (1994). Diversity in body size and feeding morphology within past and present vulture assemblages. Ecology, 75, 1074–1084.
Hertel, F., & Lehman, N. (1998). A randomized nearest neighbor approach for assessment of character displacement: The vulture guild as a model. Journal of Theoretical Biology, 190, 51–61.
Houston, D. C. (1974). Food searching in griffon vultures. East African Wildlife, 12, 63–77.
Houston, D. C. (1979). The adaptations of scavengers. In A. R. E. Sinclair & M. N. Griffiths (Eds.), Serengeti: dynamics of an ecosystem (pp. 236–286). Chicago: University of Chicago Press
Houston, D. C. (1987). Management techniques for vultures—feeding and releases. In D. J. Hill (Ed.), Breeding and management of birds of prey (pp. 15–29). Bristol: University of Bristol.
Jaksic, F. M. (1981). Abuse and misuse of the term “guild” in ecological studies. Oikos, 37, 397–400.
Jedrzejewski, W., Zalewski, A., & Jedrzejewska, B. (1993). Foraging by pine marten (Martes martes) in relation tofood resources in Białowieza National Park, Poland. Acta Theriologica, 38, 405–426.
König, C. (1974). Zum verhalten spanischer Geier an Kadavern. Journal für Ornithologie, 115, 289–320.
König, C. (1983). Interspecific and intraspecific competition for food among old world vultures. In S. R. Wilbur & J. A. Jackson (Eds.), Vulture biology and management (pp. 153-171). Berkeley: University of California Press.
Kruuk, H. (1967). Competition for food between vultures in east Africa. Ardea, 55, 171–193.
López-López,P. García-Ripollés, C., & Urios, V (2014). Food predictability determines space use of endangered vultures: implications for management of supplementary feeding. Ecological Applications 24, 938–949.
Maes, J., Teller, A., Erhard, M., Liquete, C., Braat, L., et al. (2013). Mapping and Assessment of Ecosystems and their Services. An analytical framework for ecosystem assessments under action 5 of the EU biodiversity strategy to 2020. Publications office of the European Union, Luxembourg.
Margalida, A., & Heredia, R. (2005). Biología de la conservación del quebrantahuesos (Gypaetus barbatus) en España. Organismo Autónomo Parques Nacionales. Madrid: Organismo Autónomo De Parques Nacionales.
Margalida, A., Donázar, J. A., Carrete, M., & Sánchez-Zapata, J. A. (2010). Sanitary versus environmental policies: Fitting together two pieces of the puzzle of European vulture conservation. Journal of Applied Ecology, 47, 931–935.
Margalida, A., Colomer, M. A., & Sanuy, D. (2011). Can Wild Ungulate Carcasses Provide Enough Biomass to Maintain Avian Scavenger Populations? An Empirical Assessment Using a Bio-Inspired Computational Model. PLoS ONE, 6(5), e20248.
Margalida, A., Carrete, M., Hegglin, D., Serrano, D., Arenas, R., & Donázar, J. A. (2013). Uneven large-scale movement patterns in wild and reintroduced pre-adult bearded vultures: Conservation implications. PLoS ONE, 11(8), e65857.
Margalida, A. Campión, D., & Donázar, J.A. (2014). Vultures vs livestock: conservation relationships in an emerging conflict between humans and wildlife . Oryx, 48, 172–176.
Markandya, A., Taylor, T., Longo, A., Murty, M. N., Murty, S., Dhavala, K. (2008). Counting the cost of vulture decline-An appraisal of the human health and other benefits of vultures in India. Ecological Economics, 67, 194–204.
Mateo, R., Rodríguez-de la Cruz M., Vidal, D., Reglero, M., & Camarero, P. (2007). Transfer of lead from shot pellets to game meat during cooking. Science of the Total Environment, 372, 480–485.
McCarthy, M. A., & Possingham, H. P. (2007). Active adaptive management for conservation. Conservation Biology, 21, 956–963.
Melis, C., Selva, N., Teurlings, I., Skarpe, C., Linnell, J. D. C., & Andersen, R. (2007). Soil and vegetation nutrient response to bison carcasses in Białowieza Primeval Forest, Poland. Ecological Research, 22, 807–813.
Merckx, T., & Pereira, H. M. (in press). Reshaping agri-environmental subsidies: From marginal farming to large-scale rewilding. Basic and Applied Ecology. doi: 10.1016/j.baae.2014.12.003
Milner, J. M., Bonenfant, C., Mysterud, A., Gaillard, J-M, Csányi, S., & Stenseth, N. C. (2006). Temporal and spatial development of red deer harvesting in Europe: biological and cultural factors. Journal of Applied Ecology, 43, 721–734.
Moreno-Opo, R. (2007). El buitre negro. In: R. Moreno-Opo, F. Guil (Coords.). Manual de gestión del hábitat y las poblaciones de buitre negro en España (pp. 25–45, 404 pp.). Dirección General para la Biodiversidad. Ministerio de Medio Ambiente. Madrid.
Moreno-Opo, R., Arredondo, A., & Guil, F. (2010). Foraging range and diet of Cinereous vulture Aegypius monachus using livestock resources in Central Spain. Ardeola, 57, 111–119.
Mundy, P., Butchart, D., Ledger, D., & Piper, S. (1992) The vultures of Africa. San Diego: Academy.
Olea, P. P., & Mateo-Tomás, P. (2009). The role of traditional farming practices in ecosystem conservation: The case of transhumance and vultures. Biological Conservation, 142, 1844–1853.
Oro, D., Margalida, A., Carrete, M., Heredia, R., & Donázar, J. A. (2008). Testing the goodness of supplementary feeding to enhance population viability of an endangered vulture. PLoS ONE, 3(12), e4084.
Ostfeld, R. S., & Keesing, F. (2000). Pulsed resources and community dynamics of consumers in terrestrial ecosystems. Trends in Ecology and Evolution, 15, 232–237.
Pain, D. J., & Pienkowski, M. W. (Eds.). (1997). Farming and birds in Europe: The Common Agricultural Policy and its implications for bird conservation. London: Academic.
Payne, J. A. (1965). A summer carrion study of the baby pig (Sus scrofa, Linnaeus). Ecology, 46, 592–602.
Piper, S. E. (2006). Supplementary feeding programs: How necessary are they for the maintenance of numerous and healthy vultures populations? In D. C. Houston & S. E. Piper (Eds.), Proceedings of the international conference on conservation and management of vulture populations (pp. 41–50). Natural History Museum of Crete WWF Greece.
Possingham, H. P., Andelman, S. J., Noon, B. R., Trombulak, S., & Pulliam, H. R. (2001). Making smart conservation decisions. In G. Orians & M. Soulé (Eds.), Research priorities for conservation biology (pp. 225–244). Washington, DC: Island Press.
Prakash, V., Bishwakarma, M. C., Chaudhary, A., Cuthbert, R., Dave, R., Kulkarni, M., Kumar, S., Paudel, K., Ranade, S., Shringarpure, R., & Green, R. (2012). The population decline of Gyps vultures in India and Nepal has slowed since the veterinary use of diclofenac was banned. PLoS ONE, 7, e49118.
Root, R. B. (1967). The niche exploitation pattern of the blue-grey gnatcatcher. Ecological Monographs, 37, 317–350.
Rose, M. D., & Polis, G. A. (1998). The distribution and abundance of coyotes: The effect of allochthonous food subsidies from the sea. Ecology, 79, 998–1007.
Sánchez-Zapata, J. A., Eguía, S., Blázquez, M., Moleón, M., & Botella, F. (2010). Unexpected role of ungulate carcasses in the diet of Golden Eagles Aquila chrysaetos in Mediterranean mountains. Bird Study, 57, 352–360.
Schluter, D., & Ricklefs, R. E. (1993). Species diversity. An introduction to the problem. In R. E. Ricklefs & D. Schluter (Eds.), Species diversity in ecological communities (pp. 1–10). Chicago: The University of Chicago Press.
Selva, N. (2004). The Role of Scavenging in the Predator Community of Bialowieza Primeval Forest. Tesis doctoral, Universidad de Sevilla, Sevilla, Spain.
Selva, N., Cortés-Avizanda, A. (2009). The effects of carcass and carrion dump site on communities and ecosystems. In J. A. Donázar, A. Margalida, & D. Campión (Eds.), Vultures Feeding Stations and Sanitary Legislation: a Conflict and its Consequences from the Perspective of Conservation Biology (pp. 452–473) Munibe 29 (suppl.), Sociedad de Ciencias Aranzadi, San Sebastián, Spain.
Selva, N., & Fortuna, M. A. (2007). The nested structure of a scavenger community. Proceedings of the Royal Society of London B, 274, 1101–1108.
Selva, N., Jedrzejewska, B., Jedrzejewski, W., & Wajrak, A. (2003). Scavenging on European bison carcasses in Białowieza Primeval Forest (eastern Poland). Ecoscience, 10, 303–311.
Selva, N., Jedrzejewska, B., Jedrzejewski, W., & Wajrak, A. (2005). Factors affecting carcass use by a guild of scavengers in European temperate woodland. Canadian Journal of Zoology, 83, 1590–1601.
Sikes, D. S. (1994). Influence of ungulate carcasses on coleopteran communities in Yellowstone National Park, USA. MSc Thesis, Montana State University, Montana, USA.
Slotta-Bachmayr, L., Bögel, R., & Camiña Cardenal, A. (Eds.). (2004). The Eurasian Griffon Vulture (Gyps fulvus); in Europe and the Mediterranean: Status report and action plan. East European/Mediterranean Vulture Working Group.
Towne, E. G. (2000). Prairie vegetation and soil nutrient responses to ungulate carcasses. Oecologia, 122, 232–239.
Tylianakis, J. M., Laliberté, E., Nielsen, A., & Bascompte, J. (2010). Conservation of species interaction networks. Biological Conservation, 143, 2270–2279.
Viñuela, J., Martí, R., & Ruiz, A. (1999). El milano real en España. Monografía n_6. SEO/BidLife. Madrid.
Wilbur, S. R., & Jackson, J. A. (1983). Vulture biology and management. Berkeley: University of California Press.
Wilmers, C. C., & Getz, W. M. (2005). Gray wolves as climate change buffers in Yellowstone. PLoS Biology, 3(4), e92.
Wilmers, C. C., & Post, E. (2006). Predicting the influence of wolf-provided carrion on community dynamics under climate change scenarios. Global Change Biology, 12, 403–409.
Wilmers, C. C., Stahler, D. R., Crabtree, R. L., Smith, D., & Getz, W. M. (2003). Resource dispersion and consumer dominance: scavenging at wolf and hunter-killed carcasses in Greater Yellowstone, USA. Ecology Letters, 6, 996–1003.
Yang, L. H., Bastow, J. L., Spence, K. O., & Wright, A. N. (2008). What can we learn from resource pulses? Ecology, 89, 621–634.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2015 The Author(s)
About this chapter
Cite this chapter
Cortés-Avizanda, A., Donázar, J., Pereira, H. (2015). Top Scavengers in a Wilder Europe. In: Pereira, H., Navarro, L. (eds) Rewilding European Landscapes. Springer, Cham. https://doi.org/10.1007/978-3-319-12039-3_5
Download citation
DOI: https://doi.org/10.1007/978-3-319-12039-3_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-12038-6
Online ISBN: 978-3-319-12039-3
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)