
Abstract
Drylands are the largest biomes on Earth, yet also one of the most vulnerable to climate change and human activities. Dryland ecosystems in the world are characterized by unique and distinctive features and are known to be particularly sensitive to natural and anthropogenic disturbances. Understanding the dynamics of dryland ecosystems and their direct and indirect drivers in socio-economic and natural terms will not only provide facts and knowledge about the dynamics and drivers of future trajectories, but also provide scientific guidance for the development of appropriate measures to improve the resilience of dryland ecosystems in response to human-driven climate change. We first provide an overview of the peculiar nature of dryland land cover, which features spare and patterned vegetation, soil biocrust, and man-made solar energy surface. We specifically highlight new opportunities for remote sensing observations and discuss their potential to provide new insights into dryland ecosystem functions and services. We next discuss the importance of and trends in water availability with emphasis on the different plant water utilization strategies found across global drylands, non-rainfall water absorption, water availability estimation, and hydrological impact of land cover changes. Together these factors determine the development and degradation of drylands across global gradients of water availability. We then outline the role of climate change, population increase, and human activities in driving dryland changes. We end with a forward-looking perspective on future dryland research.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Dryland are the largest biome on Earth (Schimel 2010), yet one of the most vulnerable to climate change and human activities (Smith et al. 2019; Reynolds et al. 2007). The basic dryland landscape has long been sculpted by the interaction between low but highly variable annual rainfall, high frequency of droughts and heat waves (Huang et al. 2017), and human activities (e.g., soil cultivation, livestock grazing, and fire use). Thus, drylands are characterized by unique and distinctive features (Wu et al. 2021b), including nutrient-poor soils (Ci and Yang 2010), spare vegetation cover (Tarnita et al. 2017), biocrust (Antoninka et al. 2020), and distinct water utilization strategy (Wang et al. 2017c). These traits further influence ecosystem functions and services and reduce the resilience of ecosystem to changes in specific drivers, by providing low annual productivity (Smith et al. 2019), and regulating atmospheric carbon dioxide concentrations (Biederman et al. 2017; Ahlström et al. 2015). As a result, dryland ecosystem is regarded as a complex coupled human-environmental system (Reynolds et al. 2007). Drylands are a major component of the land surface and play an important role in global environmental change and ecological sustainability (Maestre et al. 2016; Lian et al. 2021; Li et al. 2021a), and a better understanding about drylands will help develop appropriate measures that can address anthropogenic climate changes.
Abiotic factors (e.g., precipitation and temperature), biome attributes (e.g., diversity, spatial patterns, and species invasion), and human activities (e.g., grazing, farming, and urbanization) are widely considered as the main drivers of dryland ecosystem dynamics, which represent a sophisticated social-ecological system (Maestre et al. 2012, 2016; Lian et al. 2021; Li et al. 2021a). Specifically, reduction of dryland resilience can partly be attributed to the negative impacts of extreme climate events, as it could lead to declines in vegetation diversity and cover (Delgadobaquerizo et al. 2013; Dannenberg et al. 2019), thereby disrupting species interaction networks (Burkle et al. 2013). CO2 is an important dryland change driver as well, which produces important feedbacks to the local and regional hydrological cycles by promoting plant growth and ameliorating plant water stress (Lian et al. 2021; Gonsamo et al. 2021; Donohue et al. 2013). As the most important sources of livelihoods in drylands and the direct impact of human activities on dryland ecosystems, grazing and soil cultivation are major contributors to land degradation and desertification in drylands (Evans and Geerken 2004; Reid et al. 2005). Rapid urbanization and ecological conservation and restoration are also human activities that have impacted drylands, with the former ordinarily causing a loss of species diversity, carbon stocks, and ecosystem services (Tian and Qiao 2014; Liu et al. 2019b), but also having a positive effect on poverty alleviation, and the latter enhancing greening and ecosystem services (especially in the drylands of northern and northwestern China), but also generating considerable local water stress (Li et al. 2021a).
The intensifying variability of precipitation in drylands and the risk of global warming increase the threat to ecosystem recovery in drylands compared to other humid areas (Huang et al. 2016; Berdugo et al. 2020), as modelled by the future climate scenarios, which predicts the dryland area will increase by 11–23% by the end of this century (Huang et al. 2016; Prăvălie 2016). High variability of precipitation would reduce the soil moisture and suppress the growth of vegetation in dry season, and the increasing air temperature accompanied by abundant solar radiation result in high potential evapotranspiration (Reynolds et al. 2007) and further intensify local water stress, increasing the risk of land degradation. As the nature of low fertility of dryland soils, both tillage and grazing could cause quick and major impacts to dryland ecosystem. Thus, human resource extraction usually exacerbates land degradation in drylands as well (Li et al. 2021a; Evans and Geerken 2004). Considering the increase of global temperature and population, the risk of land degradation and desertification in dryland regions is rising, as drylands are particularly sensitive to rapid rates of physical and social change. Effective government policies are particularly important due to the growing economic and social demand for rangelands and irrigated farmlands, which will influence the attributes of drylands and the functional interactions in dryland landscapes (García-Palacios et al. 2018; McCollum et al. 2017). Regional decision-makers need to consider rapid changes in precipitation, water scarcity status, and temperature changes when proposing adaptation strategies for local ecosystems and socioeconomic development (Zhang et al. 2021). The dynamic process of dryland ecosystems and its direct and indirect driving forces in both socioeconomic and natural aspects should be studied, because these factors play a critical role in revealing the changing trends of dryland ecosystems at macro scales and thus provide the facts and knowledge about the future trajectories of dryland ecosystems dynamics.
In this chapter, peculiar dryland land cover and water availability and their changes and drivers are reviewed, synthesized, and discussed, particularly in relation to remote sensing application, for an understanding of research progress and future directions to cope with anthropogenic climate change. Diversity is absolutely the significant feature of drylands in the world, which cover about 41% of Earth’s land surface and hold to more than 38% of total global population. Although large space has been devoted to discussing vegetation pattern, biocrust, photovoltaic black surfaces and plant water strategies in this chapter, we do not try to find their common features, but rather to integrate diversity into the whole description.
2 Peculiar Dryland Land Cover and Changes
One of the most distinctive features that distinguishes dryland ecosystems from other ecosystems is their unique and diverse land cover, which is the dynamic mixture of herbaceous, shrubs, trees, biological soil crusts (biocrusts), and bare ground. Land use/cover and its changes (LUCC) have been explored extensively (Liu et al. 2020; Li et al. 2017; Chen et al. 2015; Wu et al. 2017), but with little attention given to unique land cover types and their characteristics in dryland regions. In this section, the peculiar nature of dryland land cover is reviewed with a focus to highlight new opportunities for remote sensing observations and their potentials to provide new insights into the functions and services of dryland ecosystems.
To adapt to the harsh arid environment and water scarcity, the vegetation in dryland areas has evolved self-organizing patterns, the special spatial pattern ranging from patches to stripes to labyrinths (Mander et al. 2017; Tarnita et al. 2017), as the result of mutual compromise between dryland vegetation and the environment. Researchers have studied the self-organizing patterns for their potential value in indicating the transition of dryland ecosystems toward desertification (Konings et al. 2011; Ludwig et al. 2007, 2002), however, recent study points out that the self-organizing pattern should be regarded as a signal of resilience instead of evade tipping point (Rietkerk et al. 2021).
Biocrusts, a kind of photochemical soil commonly existing on the surface of drylands worldwide, are another uniquely prominent feature in drylands (Belnap 2003; Ferrenberg et al. 2017; Smith et al. 2019). Biocrusts are mainly formed by the interaction of bacteria, fungi, and algae with soil particles to develop a thin, dense, shell-like community of organisms on the soil surface (Ngosong et al. 2020; Ferrenberg et al. 2017). Biocrusts reflect the unique form of non-rainfall water use strategy by dryland organisms (Wu et al. 2021b), which largely changes the redistribution of surface water in deserts and sandy lands, and plays an important role in carbon and nitrogen cycling and soil organic matter formation (Reed et al. 2012; Darrouzet-Nardi et al. 2015; Rodriguez-Caballero et al. 2018).
In addition, as one of the most important man-made surfaces in dryland ecosystems, photovoltaic panels (PVs) are highly valued and rapidly expanding in drylands due to their ability to provide large amounts of green energy to humans. PVs alter the albedo in deserts and change the radiation balance at the surface, indirectly affecting local hydrological cycle and climate change (Arnds et al. 2017). However, the current feedback mechanisms of PVs on land–atmosphere interactions, and the impact of PVs on local environment and ecology are yet understood (Barron-Gafford et al. 2016). As a power means of earth observation, remote sensing has been able to accurately identify the distribution of PVs nationwide and worldwide (Yu et al. 2018; Kruitwagen et al. 2021), providing strong support for the exploration of the impact mechanism.
2.1 Vegetation Pattern and Changes
Vegetation in many drylands is patterned regularly. These spatial patterns are prominent in dryland ecosystems, where they are often manifested as bare soil embedded in patches of vegetation (Okin et al. 2015). These patterns include regular vegetation strips alternating with bare ground, vegetation spots and labyrinths, and regular bare ground gaps within contiguous vegetated areas (Couteron and Lejeune 2001; Klausmeier 1999), such as tiger bushes in Sahel (Rietkerk et al. 2004; Klausmeier 1999), fair circles in the Namib Desert (Juergens 2013) of Africa, cyclic vegetation patterns in southern Australian deserts (Fatchen and Barker 1979), desert shrub in north-western China (Fig. 2.1), and sparse desert scrub vegetation in the tropical deserts of Mexico. Although some common understandings have been reached that large-scale regular spatial pattern may result from local biological interactions in homogeneous landscapes (Hassell et al. 1991), the specific interaction mechanism in different dryland ecosystem is still controversial. Thus, different hypotheses and models have been proposed and simulated to explain the existence of these patterns (Tarnita et al. 2017; Zhao et al. 2019; Juergens 2013; von Hardenberg et al. 2001).

a Desert shrub in Tengger Desert, China. b Fair circle in Namibia desert. c Tiger bush in northern Sahel
The spatial pattern of vegetation self-organizes under harsh environmental conditions, related to the amount of rainfall provided to the surface according to some studies (Mander et al. 2017; Tarnita et al. 2017). If environmental conditions deteriorate, the ecosystem may tip to a barren degraded state. Therefore, the spatial self-organization of vegetation in drylands can be used as a warning signal for tipping toward an alternative stable state (Kefi et al. 2007). Early warning signals based on spatial patterns are thus highly important (Scheffer et al. 2009) as indicators for imminent tipping (Rietkerk et al. 2004) where degradation may become irreversible (or difficult to reverse). However, recent researches are prone to consider these spatial patterns as signal of ecosystem resilience instead of warning signal of critical transitions, because they are observed to stay stable for a wide range of conditions, allowing complex systems to persist beyond a tipping point (Rietkerk et al. 2021). In any case, the vegetation pattern is important for the evolution of complex ecosystems.
These patterns may positively affect essential ecosystem functions, such as ecosystem productivity in Savana (Pringle et al. 2010). The vegetation pattern is self-organized through scale dependent feedback, associated with the modification of a range of plant functional traits (Zhao et al. 2019). During the formation of spatial self-organization, vegetation can optimize nutrient utilization and enhance individual competitiveness by regulating the root-to-shoot ratio and other traits. Furthermore, vegetation could create better microhabitats for benthos through the formation of self-organization, increasing their total abundance and species richness, thus improving ecosystem productivity and stability. In addition, these patterns could enhance the landscape function in arid and semi-arid rangeland regions from the aspect of landscape as well (Bastin et al. 2002). Through the trapping and retaining of rain water, soil particles, and organic matter from vegetation patch, these patterns provide more favorable habitat for vegetation and fauna. Thus, the landscape with such vegetation patterns is considered more functional and healthier and these vegetation patches’ spatial pattern can be regarded as indicators to measure the health of arid and semi-arid landscapes (Bastin et al. 2002; Ludwig et al. 2002, 2007).
The composition, structure, and function of dryland ecosystems often vary greatly over short lateral distances, reflecting the high spatial heterogeneity of moisture, which typically varies with elevation, soil type, and distance from water sources (Biederman et al. 2017). Vegetation on the soil surface both intercepts and redistributes surface water and promotes water infiltration, and it also enhances the direct and potential evaporation of stored water from soil. Thus, its spatial distribution has a strong influence on the spatial variation of moisture, which has been used to explain the formation of spatial self-organization of vegetation in drylands. On the one hand, vegetation enhances the infiltration of water into the soil and promotes the growth of vegetation to a certain spatial extent. On the other hand, competition for water among vegetation inhibits its further expansion (Rietkerk et al. 2002). In addition, the structure and composition of vegetation are influenced by seasonal to annual variations in water availability (Dakos et al. 2011; Gremer et al. 2015). Frequent or intense droughts can fundamentally alter the structure of vegetation ecosystems because long-term limited water availability inhibits further vegetation growth (van der Molen et al. 2011), while high-intensity fluctuations in water in space and time make vegetation ecosystems highly vulnerable to global environmental changes and anthropogenic disturbances (Safriel and Adeel 2008).
However, changes between the formation or disappearance of these patterns and water availability are not in real time, with hysteresis phenomenon, in fact, pervasive (van de Koppel et al. 2002). Such changes imply that both spotted vegetation patterns and bare ground at very low rainfall levels in the drylands represent the steady state of the respective ecosystems (von Hardenberg et al. 2001). The disappearance of spotted vegetation indicates a complete loss of effective root networks and enhanced water infiltration mechanisms, while bare ground may be re-covered with vegetation only when rainfall levels greatly exceed the formation level of spotted vegetation patterns (Scheffer 2020).
Therefore, the spatial patterns of the surface and patch-size distributions are interesting elements to be observed by high-resolution satellite data (Xu et al. 2015) or multi-angular data since they determine the partition of water and allow for a diagnosis of the state of ecological functioning. In addition, the spatial distribution of vegetation in the form of spaced clumps (clumped vegetation) produces anisotropic radioactive reflectance that significantly alters the surface albedo. The specific bidirectional reflectance distribution functions they present are thus beneficial to differentiate surfaces with clumped vegetation from others (He et al. 2012).
However, multi-angle satellite data such as Multi-angle Imaging Spectro Radiometer (MISR) (Chopping et al. 2008) and Compact High Resolution Imaging Spectrometer onboard the Project for On-board Autonomy (CHRIS-PROBA) (Verrelst et al. 2008) have dropped out of use, and only a few satellites such as ZY-3 are currently available to provide this type of measurement (Wang et al. 2021). ZY-3 was launched in January 2021 and carries three high-resolution panchromatic cameras and an infrared multispectral scanner (IRMSS). Positioned in the forward, longitudinal, and rearward views, respectively, these cameras allow three-dimensional mapping and can be used to map spare vegetation distributions. Recently, there has been a sharp increase in high-resolution satellite data with short payback periods taken from different viewing angles, allowing for multiple perspectives of the same target. Thus, they can be used for the observation of spare vegetation patterns. In addition, by obtaining old aerial photographs, such as declassified photographs from military satellites like the Corona series, it has become possible to track changes in vegetation patterns in drylands since the 1960s (Andersen 2006), which might provide surprising information (Li et al. 2021a).
2.2 Biocrust and Changes
Biological soil crust (BSC) is a photoautotrophic community composed of algae, bacteria, lichens, mosses, and other microorganisms that widely grow and develop in vegetation interspaces. They are the most characteristic pioneer organisms in drylands, accounting for 40–70% of the living cover (Belnap et al. 2016), and developed a thin layer of shell organisms on the surface of soil or rock by the interaction between fungi, green algae, cyanobacteria, lichens, and mosses and soil particles (Fig. 2.2) (Lan et al. 2017). BSC is often used as one of the major indicators of ecosystem stability and degraded ecosystem restoration evaluation and plays an important role in improving soil physical properties and promoting ecological restoration (Root et al. 2017; Couradeau et al. 2019; Zhou et al. 2019). Therefore, it is necessary to avoid its degradation or disappearance (Zhao et al. 2021; Giraldo-Silva et al. 2020; Antoninka et al. 2020). Some researchers found that the interaction mode between BSCs and soil will change the physical and biological environment, determine the soil texture, nutrient composition, and soil surface morphology, and affect the hydrological cycle and the capture of soil, organic matter, seeds, and nutrient-rich dust (Zhou et al. 2019; Lehnert et al. 2018; Pointing and Belnap 2012). These changes to the environment will affect the whole ecosystem.

Different kinds of biological soil crusts (BSCs)
The formation of BSCs is generally divided into five stages: sand, physical crust, algal crust, lichen, and moss. With the different stages of BSCs, their biomass and other physical properties also change. Due to their unique physiological structure and function, BSCs have a strong ability to adapt to the environment (Zhou et al. 2020). They present important ecological functions, such as enrichment of soil nutrients and nitrogen fixation through photosynthesis (Wang et al. 2017b; Ngosong et al. 2020), and affect key desert ecosystem processes in arid and semiarid areas to a great extent, and are related to Earth system functions through potential impacts on global biology and climate (Williams et al. 2016). In the 1980s and early 1990s, researchers conducted considerable research on BSCs, especially in Australia, Israel, and the western United States (Jeffries et al. 1992; Bolton et al. 1993; Tueller 1987; Bonell and Williams 1986). The study of BSCs involves many directions, such as the impact of BSCs on soils, runoff and hydrology, and the ecological role of BSCs in surface restoration. BSCs in China are mainly distributed in the Taklimakan Desert, Gurbantünggüt Desert, Tengger Desert, Mu Us Sandy Land, Kubuqi Desert, Ulan Buh Desert, etc. (Fig. 2.3) and have been studied extensively (Yang et al. 2019b; Weber et al. 2008).

BSCs covering drylands worldwide are an important functional vegetation unit and play an important role in the carbon and nitrogen cycling of desert ecosystems (Chamizo et al. 2012; Weber et al. 2015; Swenson et al. 2018; Klopatek 1992; Ferrenberg et al. 2017). On a global scale, cryptogamic covers absorb approximately 3.9 Pg/yr C and 49 Tg/yr N, which account for approximately 7% of the net primary production of terrestrial vegetation and nearly half of the terrestrial biological nitrogen fixation, respectively (Elbert et al. 2012). However, BSC occurrence is mainly driven by a combination of precipitation, temperature, and land management, and land use and climate change might cause a loss of BSC coverage by approximately 25–40% within the next 65 years, which will substantially affect ecosystem functioning, including reducing the microbial contribution to nitrogen cycling and enhancing the emissions of soil dust (Rodriguez-Caballero et al. 2018). Therefore, as one of the important species in dryland ecosystems and desertification areas, BSC is of great importance for regional management. However, we lack accurate data on the spatial distribution and change in BSCs, which also leads to great limitations in our understanding of BSC coverage and functions (Smith et al. 2019). Therefore, how BSCs respond to climate change and how BSCs change the nutrient cycle (Darrouzet-Nardi et al. 2018) and spectrum of arid land (Rutherford et al. 2017) need further exploration.
BSCs can change the color and spectral characteristics of bare soil. Spectrum bands between 600 and 700 nm have absorption characteristics associated with the specific pigments in the components of BSCs, which may be used for remote sensing detector identification (Chen et al. 2005; Ustin et al. 2009; Karnieli et al. 1999). Accordingly, remote sensing offers an opportunity to detect and monitor the distribution of BSCs. However, several factors hamper the detection of BSCs, as summarized by Smith et al. (2019). The spectral reflectance of BSCs dominated by algal crust, determines the spectral difference between vascular plants and sandy soil, and proposed the crust index (CI) (Karnieli 1997). The phycocyanin in cyanobacteria has higher reflectivity in the blue spectral band than in similar substrates without BSCs. According to the normalized difference of the red and blue spectral bands, the CI image is more sensitive to ground objects than the original image. The application of the proposed CI index can be performed by any image obtained by a sensor containing a blue band. Other indices, such as Biological Soil Crust Index (BSCI), Continuum Removal Crust Identification Algorithm (CRCIA), and Crust Development Index (CDI) (Chen et al. 2005; Weber et al. 2008; Rodríguez-Caballero et al. 2017) were proposed for BSC detection with multispectral satellite images. Evaluation of the CI and BSCI with the field based on spectral and hyperspectral images shows that these two methods are not suitable for accurately distinguishing BSCs from bare soil and plants in heterogeneous areas (Weber et al. 2008). The CI is based on the normalized difference between blue and red bands, which makes it more suitable for detecting cyanobacteria dominated BSCs. The BSCI can better detect lichen dominated BSCs in the Gurbantünggüt Desert (China) based on the red, green, and near-infrared bands of the Landsat ETM + sensor. BSCs in arid areas are mainly sensitive to moisture, greatly different in dry and wet conditions (Smith et al. 2019).
Additionally, the study of BSCs coverage is very important because the difference in spectral response between BSCs, bare soil, and vegetation observed on a detailed scale leads to the difference in spectral response in heterogeneous mixing areas, which depends on the relative coverage of bare soil, vegetation, and BSCs observed in most dryland ecosystems. Random forest model was used to optimize BSCs extraction by using band combinations similar to that of the CI and BSCI, and it tested the coverage of BSCs in Mu Us sandy Land in North China using multispectral datasets (Landsat-8 and Sentinel-2 datasets). The findings showed that applying the random forest algorithm to Sentinel-2 dataset can accurately calculate the distribution of BSCs (Chen et al. 2019b). However, there are still many deficiencies in monitoring the temporal and spatial distribution of BSCs based on remote sensing, and the existing biocrust indices have limitations. When BSCs are soaked in dew and rain, their physical characteristics such as color will change rapidly, thus changing the spectral response. In addition, in the rainy season, the time span of BSCs is long when chlorophyll is quickly formed, and has an impact on the spectral response. The above problems increase the uncertainties to estimate BSCs cover and surface feature extraction, which are usually ignored in vascular plant research (Ferrenberg et al. 2017). Smith et al. (2019) suggested that a key first step to remote sensing monitoring of BSCs is their explicit incorporation into existing remote sensing algorithms. Most existing land cover algorithms do not include a BSC classification, although BSCs account for 12% of the terrestrial Earth surface (Rodriguez-Caballero et al. 2018) and 40–70% of the living cover in drylands (Belnap et al. 2016). New China cover products have taken this important step forward and added a BSC classification at a resolution of 10 m for years 2015 and 2020 in areas with less than 20% vegetation cover.
2.3 Photovoltaic Black Surfaces
As one of the important factors leading to global warming and climate change, the burning of fossil fuels, such as petroleum and coal, is also an important factor that many countries rely on for development (Mohsin et al. 2019). Energy demand continues to rise, which is likely to further increase carbon dioxide emissions worldwide. Studies have shown that photovoltaic electricity generation accounts for 10% of the grid, which will cause a 12.3% reduction in the global CO2 emissions volume (Zhai et al. 2012). Therefore, governments around the world are committed to achieving the goal of energy savings and emission reduction through the use of renewable resources (Zhu et al. 2019). Renewable energy comes from abundant natural resources, such as solar energy, wind energy, and biomass. Therefore, it is considered to be an eco-friendly energy source with zero to minimum carbon dioxide emissions (Malahayati 2020).
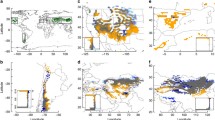
Drylands are the main places to host green energy, such as solar and wind energy. Studies have shown that if 4% of the Earth’s desert areas were fitted with PVs, the energy provided could meet the consumption needs of the entire world (Prăvălie 2016). According to the climatic definition of the sum of ultragrid and arid regions, there are currently approximately 30 million square kilometers (approximately 20% of terrestrial land) of desert area (Ezcurra 2006). Photovoltaic (PV) solar power generation has grown 41% annually since 2009, and the trend is still expanding. It is estimated that by 2040, PV solar power generation capacity will increase nearly ten times. Geospatial data describing the energy system are needed to manage the intermittent power generation, mitigate climate change risks, and determine biodiversity, conservation, and land conservation priorities due to land use and land cover changes required for trade-PVs deployment. Globally, there are approximately 68,661 facility footprints for spatial positioning (Fig. 2.4), with 423 GW (−75/+77 GW) generated from the end of 2017 to September 2018 (Kruitwagen et al. 2021). In the International Energy Agency’s (IEA) Sustainable Development Scenario, it is estimated that 4,240 GW of PV solar power generation capacity will be deployed by 2040, which is a 10,000-fold increase from 385 MW in 2000 (Zhu et al. 2019) and a tenfold increase from 2018.

Photovoltaic black surface in a Zhongwei, China, b Centre solar Ouarzazate, Morocco, and c Arizona, United States
The largest proportion of PV solar panels is located in farmland, followed by dryland and grassland (Kruitwagen et al. 2021). PVs system installed in farmland can achieve significant power generation without potentially reducing the crop yield (Miskin et al. 2019). Some countries, such as China and the United States (Yu et al. 2018; Kruitwagen et al. 2021), have deployed a large amount of photovoltaic solar energy in arid areas. Although the black surface of the PV solar panel absorbs most of the sunlight, which changes the distribution of solar energy, only a small part (approximately 15%) of the input energy is converted into electrical energy. The rest is converted into thermal energy in the form of heat. At the same time, the color of solar panels is usually darker than the color of the ground they cover, resulting in considerable extra energy absorbing and releasing in the form of heat into the surrounding environment, and thus leading to climate change. Meanwhile, the deployment of large-scale PV power plants by changing the amount of albedo affects the absorption and storage of energy on the ground (Arnds et al. 2017). The surface runoff and percolation potential are significantly increased at the local scale (Pisinaras et al. 2014). If this effect only occurs in local areas, then it may not affect desert areas and less-populated areas. However, in the context of carbon neutrality and carbon peaks worldwide, countries are laying PV solar panels over a large area of thousands of square kilometers. From such a large area, the heat reradiated by PV solar facilities will pass through the atmosphere. The circulation effect redistributes energy, which will have an impact on regional and even global climates.
With the large-area use of PV solar panels, environmental problems and climate change have attracted increasing attention. In fact, whether ground observations or satellite remote sensing observations are performed, the ecosystem and microclimate of the area where the PV solar panel is installed have undergone complex changes (Barron-Gafford et al. 2016).
The change mechanism in local areas is mainly the feedback of land–atmosphere interaction, at the same time, it’s also the main mechanism of drought in the Sahara due to overgrazing. Studies have found that similar land cover changes can trigger ecological and local climate responses, especially in arid/semiarid regions. Interestingly, for the first time, research has linked this land–atmosphere interaction feedback mechanism with solar PVs in the Sahara Desert. The results show that the installation of large-scale PV solar panels has improved the vegetation conditions in the area and increased precipitation (Li et al. 2018b). When a global atmospheric model with a dynamic surface is used for simulation, the PV black surface mask will cause higher land surface temperature and convergent currents compared to the desert surface, which will lead to more rainfall and promote vegetation growth. The expansion of vegetation coverage further reduces the surface albedo, and this positive feedback mechanism further expands the initial temperature and humidity conditions in the area. The construction of a PV station changes the original surface roughness, which affects the ground reception and reflection longwave radiation, wind field type, turbulence intensity, atmospheric boundary layer height, etc. which in turn change the ventilation and heat dissipation conditions of the PV station. These changes will change the local temperature and change the radiation balance (Millstein and Menon 2011). Based on the RCP2.6 scenario, with the installation of PV modules in the Northern Hemisphere, the temperature decreases by as much as 1 °C in the eastern region; however, in the Southern Hemisphere, due to the limited installation area of PV modules, the cooling effect is significantly reduced, and the change in temperature has led to changes in the global precipitation pattern (Li and Gao 2021).
However, these local-scale changes are expected to have larger-scale effects via ocean dynamics and atmospheric remote correlation. At the same time, these effects may significantly alter the assessment of the mitigation potential of solar farms, but existing models are unable to fully capture these effects due to the assumption of a constant ocean temperature and heat transport.
Large-scale PV solar farms constructed in the Sahara Desert are expected to meet the world’s energy demand while also increasing the rainfall and vegetation cover in the region. However, in other locations far from the region, such impacts may offset this regional benefit. It has been shown that the redistribution of precipitation has led to drought and forest degradation in the Amazon, global surface temperature increases and sea ice disappearance, especially in the Arctic, due to increased polar heat transport and northward expansion of deciduous forests in the Northern Hemisphere (Lu et al. 2021). These remote impacts through atmospheric teleconnections and ocean dynamics by other large-scale PV farms in the rest of the world have not been addressed.
3 Dryland Water Availability and Changes
Another unique feature of dryland ecosystems is their low rainfall but high variability and high frequency of extreme weather events (Huang et al. 2017; Zhang et al. 2021). Water is the main influencing factor for dynamic changes in dryland ecosystems (Hoover et al. 2020), both for ecosystem sustainability and livelihood (D’Odorico and Bhattachan 2012). The greatest feature of dryland is the lack of water, which affects natural and managed ecosystems, restricts the production of livestock and crops, wood, fodder, and other plants, and affects the provision of environmental services. In arid and semiarid regions, strong coupling occurs between ecological and hydrological balance. This coupling demonstrates the central role of hydrological balance in dryland areas (Verstraete et al. 2009). In this section, we synthesize the importance of and trends in water availability with the emphasis on different plant water utilization strategies found across global drylands, non-rainfall water absorption, water availability estimation, and hydrological impact of land cover changes. Together these factors determine the development and degradation of drylands across global gradients of water availability.
Drylands are highly sensitive to strong daily, seasonal, and interdecadal perturbations of water availability (Sloat et al. 2018; D'Odorico et al. 2006). On a short time scale, such as from daily to interannual, dryland changes are dominated by climate variability. However, on a longer time scale of ten to one hundred years, the physiological effects of carbon dioxide-induced vegetation fertilization have important feedbacks on the local and regional hydrological cycles (Lian et al. 2021). Plants can evolve physiological and developmental processes to cope with unfavorable growth conditions. In the context of a rapidly growing population with a continuous increase in demand for water and food, the role of plant physiological mechanisms in coping with water stress and promoting their own growth will become even more important (Lian et al. 2021).
Renewable water from drylands is estimated at only 8% of the world’s total, which is insufficient to support ecosystems at optimal functioning (Ma 2005). Moreover, water competition for humans and environmental demand has caused water scarcity and constrained the economic development in many regions around the world including the Colorado River Basin of Argentina (Wild et al. 2021). The Murray Darling River, one of the major food production basins in Australia, is in a similar state and recently a plan was established to help protect and restore regional water resources (Leblanc et al. 2012). Similarly, China has established a ‘‘Stringent Water Resources Management System”, or the ‘‘Three Red Lines” as a long-term framework for addressing key water challenges (Wu et al. 2021a). Rather than an environmental challenge, achieving water security is a governance issue that requires political will, resources, and leadership (Stringer et al. 2021) to develop a synergy approach that considers the needs of humans, the environment, and ecosystems in drylands.
3.1 Plant Water Utilization Strategy
Natural vegetation has ecological significance in inhibiting drought and maintaining the stability of riparian ecosystems in arid and semiarid regions (Ye et al. 2010). The ability of plants to tolerate and recover from periodic water stress affects their competitiveness, survival, and distribution, thus leading to shifts in plant communities as environmental conditions change (Kilgore et al. 2021). Drought stress is a serious adverse factor that limits plant growth and productivity (Reddy et al. 2004). Drought stress induces a range of physiological and biochemical responses in plants. In arid and semiarid regions, plants have formed many adaptive mechanisms and strategies in response to water deficit through long-term natural selection and coevolution (Bacelar et al. 2006; Dichio et al. 2006). When encountering drought, the production of the phytohormone abscisic acid is triggered in plants through the accumulation of stress tolerance-related permeates and proteins, which in turn leads to the closure of stomata and induces the expression of related stress genes (Shinozaki and Yamaguchi-Shinozaki 2007). Plant root arbuscular mycorrhizal symbionts are the oldest and most common strategy to improve plant nutrient access and environmental stress response (Klironomos 2003). For example, inoculation of arbuscular mycorrhizal fungi under water deficit conditions is an effective measure to ensure or increase corn yield (Celebi et al. 2010).
Research on the relationship between climate and plants dynamics at large or regional scales has mainly focused on the response of the different vegetation types under a climate gradient to different climate types (Thuiller et al. 2004). Climate factors generally include temperature (annual average temperature, accumulated growth temperature, highest monthly average temperature, etc.), water (precipitation amount and timing, potential ET, vapor pressure deficit, air humidity), light (solar radiation), and other factors, which often determine the distribution of plants (Reich and Oleksyn 2004). Precipitation is one of the factors that determine the plant species distribution and community composition. Especially in areas with rapid declines in water availability, precipitation is the limiting factor which related to the plant diversity. Therefore, along the precipitation gradient, the plant water strategy becomes a matter of choosing whether to absorb water quickly or store water effectively. Plants can implement water-saving strategies to avoid drought through the effective use of limited water resources (Liu and Ma 2015). In dryland ecosystems, plants have developed many distinct strategies to allow the use of fog water through the canopy (Wang et al. 2017c). More than 80% of plant distribution patterns in Western Africa are significantly related to annual rainfall (Maharjan et al. 2011). Wood density and deciduousness are the determinants of plant drought tolerance. The plant species in this area are mainly tall trees that present slender and straight trunks, branching near the top platy roots, smooth and thin bark, and large dark green leaves and leathery texture. These characteristics allow plants to quickly absorb water when the rainy season comes.
In Asia, Populus euphratica mainly grows in extremely arid desert regions. It is mainly distributed around the Taklimakan Desert in China. Populus euphratica is a typical abiotic stress-resistant woody species. Biologists discovered in the Populus euphratica genetic spectrum that small molecules and noncoding microRNAs play an important role in growth, development, and drought stress response (Li et al. 2011). To adapt to the arid environment, the leaves on the shoots of young trees are narrow and long, while the leaves on large trees are round. After a long period of evolution, Populus euphratica is tolerant to light and resists heat, drought, salt, and alkali conditions. Populus euphratica will follow wherever the desert river flows. They can survive well by relying on the protection of well-developed root systems and a groundwater level which is not lower than 4 m (Fig. 2.5a).

Populus euphratica forest in a Xinjiang Uyghur, China, b Saguaro cactus in Western USA, c Cavanillesia arborea in South America, and d Baobabs in Mozambique
In North America, Carnegiea gigantea (Saguaro cactus) (Fig. 2.5b) is a long-lived columnar cactus endemic to the Sonoran Desert, found in Mexico and southern Arizona in the United States. Among their many adaptations, Saguaro utilizes a distinctive photosynthetic pathway evolved to minimize water loss in hot and arid environments known as Crassulacean acid metabolism (CAM) photosynthesis (Bräutigam et al. 2017). Saguaro, such as all CAM plants, absorbs CO2 through stomatal pores at night when air temperatures are relatively low and then keeps the stomata closed throughout the day when radiation and air temperatures are relatively high, which greatly reduces water loss (Knauft and Arditti 1969). Saguaro also has shallow root systems to capture rainwater from the slightest of rainfall events, spines as modified leaves that protect the plant and prevent water loss associated with dry winds, and pleats that allow the plant to expand to hold more water during wet periods and contract during extended dry periods (Lajtha et al. 1997).
In South America, Cavanillesia arborea (bottle tree) (Fig. 2.5c) is a plant with distinctive water storage strategy. Bottle trees are native to the Brazilian Plateau of South America. The water is stored in the bottle tree, which has a very sturdy trunk, but few branches and leaves. This shape, different from the shape of other trees, is affected by the drought environment. In the area where bottle trees are found, the dry season and the rainy season alternate throughout the year, but the rainy season is short and the dry season is long. To survive in such an environment, they only grow sparse leaves during the rainy season, and not in the dry season. This growth form of the bottle tree reduces the transpiration and loss of water. Some bromeliads in Mexico develop specialized trichomes (Andrade 2003), and several Crassula species located in the Namib Desert take water up through hydathodes into their leaves (Martin and von Willert 2000).
In the African mainland, Madagascar, and Australia, Baobabs (Adansonia) (Fig. 2.5d) are native to the dry and hot savannah region, characterized by their massive size and multiple uses (Sanchez et al. 2010). Although all baobab trunks are thick, the woodwork is very sparse, a characteristic evolved to survive the dry season. During the rainy season, the thick body and loose wood are used to absorb and store large volume of water to withstand the long dry spells, which is why elephants, eland, and other animals chew the bark during the dry seasons. Whenever the dry season comes, it will quickly shed all its leaves to reduce water loss. Not only the trunk, but also the leaf characteristics of baobab trees reflect the species’ wisdom in coping with long droughts. Studies have shown that the stomatal density of the leaves has a high correlation with climatic characteristics, positively correlated with local temperature and negatively correlated with precipitation (Sanchez et al. 2010; Abrams et al. 1990).
Organism responses in environments where rainfall is intermittent, and where the amplitude and longevity of soil moisture pulses are more important than the mean soil water levels, have long been characterized as a pulse—reserve systems (Smith and McAllister 2008). In Oceania, mainly in Australia, the stem succulent strategy can cope with very dry conditions but depends on reliable re-charge every year, a condition that is safe in the north and central American deserts but not met in central Australia (Smith and Morton 1990). Some organisms create niches for others, incidentally or mutualistically, and it seems that this occurs to protection from resource limitations more frequently in arid biomes than other ecosystems. For example, it’s more common in arid environments that the presence of one plant facilitates the establishment or growth of another, of the same or, usually, different species (Flores and Jurado 2003).
3.2 Nonrainfall Water
Nonrainfall water includes dew, fog, and water vapor adsorption, which is an important surface water input to dryland ecosystems in arid and semi-arid zones (Gao et al. 2020). It is believed that nonrainfall water inputs (NRWIs) are extremely important for water-limited dryland ecosystems and play an important role in the dynamics of dryland ecosystems (Kaseke et al. 2017). NRWIs represent a particular water source especially for biocrusts, insects, and plants in desert regions (Kaseke et al. 2012; Zhang et al. 2015). More significant for dryland areas where precipitation is low relative to water demand (e.g., potential ET) than other non-dryland areas, NRWIs also occur in non-dryland areas, such as the Caribbean Islands and New England, USA (Wang et al. 2017c).
NRWIs are mainly provided to the soil surface through three levels: the adsorption of fog, dew, and water vapor. Each level has its own formation mechanism, and formation occurs under certain meteorological conditions and/or surface conditions (Wang et al. 2017c; Zhang et al. 2015; Meng and Wen 2016). In some inland deserts, precipitation in the form of rain or snow is the main water resource while dew and fog are considered ancillary moisture resources (Jia et al. 2014).
Dew in the land surface process model affects the microwave bright temperature and backscattering coefficient of vegetation (Kabela et al. 2009). Dew evaporation contributes 5% of the total water vapor flux measured above the canopy with a microwave radiometer (Schneebeli et al. 2011). It has been shown that the formation of dew is mainly influenced by the intensity of radiative cooling, water vapor pressure, and wind speed (Yokoyama et al. 2021).
Dew water may play an important role in keeping plants hydrated by absorbing water through the leaf surface. Dew in the semiarid desert valley area of northeastern Nevada contributes approximately 14 mm to the total annual water content which represents approximately 10% of the annual rainfall (Malek et al. 1999), and in the Taklimakan Desert of China, the average daily amount is 0.13 mm for over 77% of the growing season days, with a cumulative amount of dew for those days 16.1 mm (Zhuang and Zhao 2017). Dew even increases the CO2 assimilation rate and leads to the complete recycling of plant water status and leaf pigment content, which is significant in the hydration and activation metabolism of water stressed in summer (Munné-Bosch and Alegre 1999).
Fog can be detected and mapped with both geostationary and polar satellite data (Amani et al. 2020; Wu and Li 2014). In some coastal locations, vegetation appears to use fog-drop water year-round. Seasonal advective and orographic fog supply the only significant annual moisture along 3000 km long in the hyperarid coastal belt of Peru and Chile (Moat et al. 2021). In addition to providing water sources, fog water also helps to change the energy balance of vegetation, reduce transpiration, increase stomatal conductance, and increase the CO2 absorption rate (Martin and von Willert 2000).
These NRWI components, typically very small but important for determining the magnitude of water and energy flux (e.g., latent heat) during the dry season (Gao et al. 2020; Wang et al. 2017c; Uclés et al. 2013), may become a major water source that helps reduce various water pressures for living things in a particular environment. Studies have shown that canopy dew at the Mizhi Experimental Station in Shaanxi Province significantly reduces night sap flow, and the contribution of NRWIs to ET can reach 18.4% (Gao et al. 2020). In some coastal desert areas, NRWIs may exceed precipitation (Henschel and Seely 2008).
In recent years, NRWI research has again become a research focus, arising largely from the state of human-water tensions in the world. There is growing evidence that these small but important NRWIs have a significant impact not only on vegetation growth but also on the survival of other microorganisms and the maintenance of the dynamic balance of geo-biochemistry under arid and low rainfall conditions (McHugh et al. 2015). However, NRWI research currently mostly uses traditional observations to focus on local areas and small-scale units to explore its magnitude and ecological effects on local areas. The method to use remote sensing for performing large-scale quantitative estimations of NRWIs and their ecological effects on drylands is still lacking.
3.3 Water Availability
The major challenge for managing water resources in modern, developed river basins is to determine the safe, sustainable limit for water utilization. In many river basins around the world, water is overallocated and over extracted for use in agriculture, cities, or industry. In fact, irrigated agriculture accounts for roughly 80% of global freshwater consumption and 40% of global crop production, used to stabilize food and feed production across dryland regions (Döll and Siebert 2002; Siebert and Döll 2010; McCabe and Wolock 2007). As a result, insufficient water is available for environmental and ecosystem flows, and in many places, rivers no longer reach the sea, groundwater tables have dropped, and lakes and natural habitats have become dry and degraded (Grafton et al. 2013).
Global, regional, and basin-wide estimates of water availability rely on models (Masood et al. 2015; Trenberth et al. 2007; Hanasaki et al. 2008), subject to large uncertainties due to soil hydraulic parameters, weather conditions, and land cover changes, and how these factors correlate with water availability (Hanasaki et al. 2008). Human activities and climate change have greatly influenced the natural hydrological cycle and changed the availability of water resources (Grafton et al. 2013). In recent decades, the natural landscape and associated hydrological characteristics have changed considerably worldwide as well as in China. In China’s Loess Plateau, evidence has shown that revegetation intensity has been approaching regional sustainable water resource limits (Zastrow 2019; Feng et al. 2016). There is a huge decrease in the ratio of annual runoff to precipitation in many catchments because of the expansion of forests which consume more precipitation (Zastrow 2019). The overexpansion of cropland and ecological shelterbelt is reported as the major causes for the shrinkage of the Ebinur Lake basin (Zeng et al. 2019). Likewise, the Colorado River Basin (CRB) in the United States has experienced an increasing demand for water due largely to agricultural intensification, driving more frequent periods of water shortages as precipitation didn’t increase while temperature increase, and leading to failures in meeting water allocation demand (Woodhouse and Pederson 2018).
All of these interventions change the spatiotemporal component development of the hydrological cycle; however, they are not adequately reflected in the hydrologic models on which estimates of sustainable water use are based. These conditions continue to simulate water availability under natural conditions (Hanasaki et al. 2013). It is complex to precisely assess the available water amount for human use in a highly developed basin using available hydrological methods. It is also difficult to set up full coverage monitoring system through metering facilities due to the high cost of both building and maintenance (Berbel and Esteban 2019). Thus, objective estimates of the available consumable water (ACW) for human use, i.e., the consumption cap at the basin or subbasin scale require a water consumption balance approach (Wu et al. 2018). Such an approach would provide a solid basis for analyzing the influences of climate change, cropland expansion, and large-scale revegetation programs.
ACW is the total amount of water in the basin sustainably available for human activities after accounting for natural inflows and outflows and the requirements of environments and ecosystems. To guarantee sustainable water resource management, ACW is calculated with the following principles: (1) groundwater overexploitation is forbidden; (2) enough water is saved for sustainable natural ecological systems; (3) basic environmental flow is considered in river systems; and (4) the water cycle between ground and surface water systems is conserved (Wu et al. 2014).
Essentially, the ACW provides water managers with a maximum allowable human activities cap or the upper limit on water consumption in basins or watersheds, which can also be called the water boundary at basins (Zipper et al. 2020). The ACW water balance equation can be rewritten as follows:
In this equation, System Inflows include upstream river inflows and inter-basin transfers, both of which are monitored and measured with stream gauges. System Outflows include the outflow of water dedicated for environmental needs (for habitat and aquatic ecosystem services, etc.), as well as any outflows unusable by humans, such as flood runoff exceeding designed dam or water bank holding capacity, groundwater recharge to saline aquifers, and rivers and lakes sewage discharge. Precipitation is the average annual precipitation in the basin, a key parameter for the ACW. Precipitation can be accurately measured at individual rainfall station sites, but at the basin scale, station-based monitoring may produce large errors due to the insufficient number of stations, especially in upstream mountainous areas where rainfall can be relatively high. New remote sensing technology can now be combined with rainfall station monitoring to produce high-precision precipitation data sets across all land-use types in the basin. Natural ET (ETnatural) is the uncontrollable ET from natural forests, grasslands, wetlands, etc. Traditional water resource accounting methods cannot accurately measure ETnatural which is critical to calculate the realistic, sustainable cap on human water consumption in the basin—i.e., the ACW. In the water balance equation, the basin ACW can be calculated by subtracting ETnatural from the other measurable inflows and outflows. Remote sensing can be used to determine ETnatural (see below). Satellite remote sensing is a revolutionary technology that allows, for the first time, comprehensive and accurate measurement of the three key dimensions in the ACW sustainable water balance equation: land-use, precipitation, and ETnatural.
Remote sensing to classify land-use—Satellite imagery is used to optically classify land-use into categories relevant to consumptive water use amounts and patterns. Land cover throughout the basin is divided into two categories: (1) natural land cover is the land without human intervention, including natural forests, grasslands, shrubs, bare lands, etc.; and (2) artificial land cover is the land with human development, such as agricultural fields (irrigated or rainfed) and urban settlements (including irrigated and impervious surfaces). For instance, across agricultural regions of the western Unites States, recent land-use classification algorithms capable of mapping irrigated croplands have revealed widespread land fallow during drought events, especially in regions of secondary water rights (Deines et al. 2017; Norton et al. 2021). There is an intrinsic link between each different land-use type and its associated ET rate, which can be determined by statistical processing of the RS data, as discussed below. Land-use changes over time in any basin, and remote sensing can regularly update land-use images to re-calculate ET estimates.
Remote sensing to measure precipitation—Coupling remote sensing derived precipitation (Pradhan et al. 2022) with ground rainfall observations can generate accurate precipitation estimates with fine spatial resolution throughout an entire river basin or watershed. The approach adopts machine learning based statistical technology to quantify the relationship between precipitation and such influential factors as vegetation, topography, cloud, and other physical variables, and build the regression formulation: P = F(Veg., Terrain., Cloud, Others), and then use the high resolution variables as input to generate high resolution precipitation (Elnashar et al. 2020). This downscaling approach works well for annual precipitation estimates. For monthly or daily precipitation estimates, more sensitive variables with precipitation than vegetation should be integrated into downscaling models, such as the physical suitable variables of cloud that can overcome inaccuracies due to the possible lag in response of vegetation to precipitation (Duan and Bastiaanssen 2013).
Remote sensing to predict ETnatural—Natural ET can be calculated from specialized remote sensing-based algorithms such as ET measurements from ETWatch (Wu et al. 2020), land cover and land use data, and known environmental factors. ET algorithms can be mechanistic or based on machine learning statistical models (Javadian et al. 2020). Total ET for any plot of land (or pixel in the satellite image) is a function of environmental factors such as precipitation, vapor pressure deficiency, wind speed, radiation status, surface temperature, soil properties, terrain, etc. Remote sensing, environmental and meteorological data can be combined in advanced, cognitive computational models using data mining in a machine learning to generate the statistical link between the natural land cover types, environmental factors, and the estimated ET. Statistical models can be used to predict ET rates for any land-use type throughout a given basin. Figure 2.6 shows a schematic of the pixel-by-pixel land-use correlations with ET rates used in machine learning analytical models for natural ET. First, the land cover is reclassified into natural and artificial land covers, and then the environmental factors and ET are divided into natural and artificial components; Second, the machine learning method is used to explore the link between ET and environmental factors of natural land cover to build the natural ET prediction model; Third, the environmental factors of the artificial land cover are used as input to predict the natural ET of artificial land cover types; Finally, the natural ET of the artificial land cover are subtracted from the total ET to obtain the human management ET.

Analytical model for predicting natural ET. The land cover was divided into natural land cover, rainfed, mosaic, irrigated agriculture, and settlement. The environmental factors and ET were also separated into natural and artificial parts. The natural ET prediction model will be built for natural land cover using the machine learning, and then will be used to predict the natural ET for each artificial land cover types
The RS-determined values for land-use, precipitation, and ETnatural can then are used in the water balance equation, along with traditional measurements for external-basin inflows and outflows. The outcome determines the ACW—i.e., the sustainable limit on total human water consumption in the basin.
This new RS-ACW water accounting methodology provides significant benefits over the traditional water resources assessment (WRA) method. Satellite RS data allow regular, inexpensive re-estimation of water balances from contemporary land-uses in the basin on an annual basis (or monthly, with some modifications). The RS-ACW approach is also more accurate, better accounting for actual precipitation and ET levels for various land-use types in the basin. Also, the RS-ACW approach does not over-estimate available water—unlike the WRA method (Wu et al. 2018). Finally, the water required for sustainable natural environments and ecosystems is ‘set aside’ before calculating the water available for human consumption.
This approach has been successfully applied in three recent World Bank projects in China. Turpan is among the poorest, extreme drylands in western China. There exists an observed paradox in Turpan whereby modern, large-scale efforts to save water eventually led to total water consumption increase as in other arid regions around the world (Tan et al. 2018; The World Bank 2012; Grafton et al. 2018). In 2008, the World Bank launched the Xinjiang Turpan Water Conservation Project to investigate the issues and recommended innovative and workable solutions (The World Bank 2012). The project’s solution to this paradox is to define a sustainable limit on total water consumption at the basin scale and watersheds, set targets for reduction, and then apply irrigation and agronomic interventions carefully to reduce the actual water consumption (ET), monitor and supervise the actual water consumption of farmers, WUAs, townships, and counties using advanced remote-sensing, and reform water rights and water pricing incentives based on ET (Wu et al. 2021a).
The project set a sustainable cap on water use and enforced a strict cap on overall water consumption for Turpan Basin and 11 watersheds, which allocated to 3 counties and down to townships and villages by conducting water balance/budget analysis with multi-stakeholder decision-making to define a prioritized budget for water consumption in the basin. This basin-level balance accounts for all water supplies and all competing water demands (i.e., ecosystem, agricultural, industry, municipal demands, etc.).
The project recognized that only “the reduction of water consumption” can fundamentally solve the water resources problem in the basin. When each watershed within the basin has arrived close to the maximum water consumption limitation then more water consumption is limited, thus it can fundamentally avoid groundwater over-extraction. The project demonstrates that remote sensing provides tools to monitor ET and cropland as independent means of monitoring actual ET vs target of farmers, WUAs, townships, and counties at relatively low cost as compared to expensive water monitoring systems used in countries such as Australia (Grafton 2017).
3.4 Hydrological Impacts of Land-Use Change
Land use changes can dramatically modify water dynamics globally (Sterling et al. 2013; Bosmans et al. 2017) and in dryland ecosystems (Lv et al. 2018; Feng et al. 2016; Yin et al. 2017). Changing current land cover types and land management activities in river basin areas will change hydrological processes such as surface runoff, base flow, ET, soil water holding capacity, interception and groundwater recharge, leading the changes of the path from rainfall to runoff, thereby reflecting changes in water demand (Naha et al. 2021; Rogger et al. 2017; Lv et al. 2019; Chen et al. 2019c; Yang et al. 2019a). Therefore, a comprehensive understanding and assessment of the impact of land cover changes on hydrological processes is necessary for watershed management, environmental policy, ecological governance, and the choice of restoration measures.
Distributed hydrological models (e.g., SWAT, VIC, MIKESHE) are used in assessing coupled hydrological processes with analyzing effects of LULC changes on these processes, where the hydrological models are calibrated and validated using ground observational data based on baseline land use scenarios. The calibrated model is then run for various land cover scenarios, and then the changes in the simulation are compared (Naha et al. 2021; Li et al. 2018a). However, there are some uncertainties concerning the input, parameters, and structure of the chosen model (Chen et al. 2019c; Her et al. 2019). Given these uncertainties, it seems reasonable to doubt the reliability of the estimated hydrological response to land cover changes, particularly when the responses are mild or moderate (Yin et al. 2017). The uncertainty induced by the model parameters or structure has unpredictable bias on the impact on the assessment of land use changes (Chen et al. 2019c). Without parameters to be calibrated, remote sensing based approach might be more promising (Wu et al. 2018; Zeng et al. 2019).
Changes from natural land cover types to artificial land cover types will significantly alter the regional hydrological characteristics. Since the development of civilization, agriculture has taken land (and water) from natural ecosystems such as forests, savannas, and grasslands. In the process of agricultural land increase, water and biogeochemical cycles have been significantly changed (Bonan 2008; Davidson et al. 2012; Runyan and D'Odorico 2016). For instance, recent findings have indicated significantly increased ET across global croplands with exceeded water inputs, suggesting that recent increase in food production may be dependent on unsustainable water inputs (Javadian et al. 2020; Pascolini-Campbell et al. 2021). Rainfed farmland keeps lower ET rates with smaller leaf area, smoother surface roughness, shallower root depth, and higher albedo to reflect solar radiation reaching ground surface (Bonan 2008; Perugini et al. 2017). Influenced by more compacted cropland soils, the infiltration rates in cropland are also smaller than natural land cover types from intensive machinery operations and fallow activities. In those regions, higher surface runoff is expected after decreasing ET and surface infiltration (Runyan and D'Odorico 2016). On the contrary, in irrigated cropland, water usage for crop growing can easily dry surface water bodies (Jägermeyr et al. 2017).
Afforestation/revegetation is encouraged worldwide for ecological purposes, particularly in Loess plateau and Sahel. However, the influence of re-vegetation on water resources remains controversial in humid tropics (Lacombe et al. 2016), but decreases water availability in drylands (Wang et al. 2017a; Lian et al. 2021; Feng et al. 2016), which may lead to irreversible catastrophic consequences for dryland ecosystems (Li et al. 2021a). In drylands, the cascade reaction of land cover changes will cause important changes in water resources, including the spatiotemporal pattern of precipitation and ET deduced from regional microclimate conditions, leading to significant changes of human available water resources (Perugini et al. 2017; Wu et al. 2018). The loss of terrestrial water storage in Yellow River was mainly caused by ET increase, which was resulted from higher vegetation cover and more irrigation water use (Lv et al. 2019). The streamflow is temporally companied with agricultural land returning forest activities in Wei River of China (Wang et al. 2017a).
Researches warn that the revegetation on the semi-arid Loess Plateau has already reached the limit of soil water-carrying capacity for vegetation (Feng et al. 2016). The resulting widespread dried soil layer potentially threatened tree mortality (Huang 2019; Wang et al. 2018). Although at present efforts are taken to better understand vegetation productivity thresholds (Feng et al. 2016), equilibrium vegetation cover (Zhang et al. 2018), regional water resource development boundaries (Wang et al. 2018), and soil water-carrying capacity for vegetation (Huang 2019; Jia et al. 2019), it remains a challenge to keep in balance between vegetation productivity and water use in order to sustain a healthy ecosystem.
Another significant land cover change is ecological protection prospective, such as farmland returning to forest and afforestation activities (Li et al. 2021a). In dryland system, ecological restoration significantly influences local hydrological cycling patterns with different water consumption mechanisms. With growing fraction of vegetation cover, plants in revegetated areas are more active in photosynthesis and transpiration processes, thus raising the demand of water consumption (Ma et al. 2019), as the revegetation in China’s Loess Plateau demonstrated that the limit of regional water resources requires sustainable ecological restoration planning (Feng et al. 2016). Large land cover shifts in Europe (e.g., deforestation or afforestation, urbanization) from the 1950s are evaluated as the same degree of net impact on the amount and distribution of water resource availability (both ET and streamflow) as precipitation changes from climate changes (Teuling et al. 2019).
The Great Green Wall (GGW) project in Sahara is conducted as a pan-African program with a strong reforestation focus (Goffner et al. 2019). However, farming and afforestation in the African Sahel and Great Green Wall (GGW) regions are constrained by climate variability, water scarcity, and degradation of lands (Mirzabaev et al. 2021). Accordingly, 1,337,535 km2 (43.5%) and 729,576 km2 (25.6%) of the Sahel and the proposed GGW region, respectively, not feasible for sustainable planting for rainfed and natural vegetation growth conditions, require supplementary irrigation (Elagib et al. 2021). Certain land use purposes as afforestation and cropland cannot be achieved without sufficient water resources, which is more water consumptive with higher ET.
Conversion of forest or woodland to cropland over large regions (e.g., >100 km) is highly correlated with precipitation reduction (especially the rainfall frequency) and diurnal air temperature increase (Bonan 2008) The regional climate can be changed by LUCC with variant component amounts of the land surface energy balance and ground-atmosphere interaction, influencing near-surface temperature, boundary layer stability, and convective precipitation (Bonan 2008; Perugini et al. 2017). The rainfall regime can also be changed within the same region in which land cover change occurs to even influence adjacent ecosystems (Ray et al. 2006). Another impact of land cover change is the modification of cloud microphysics and cloud processes by altering the biological aerosols emission rate (Pöschl et al. 2010). Regional precipitation highly relying on the regional ET will largely decrease with lower air moisture and decreased ET (Yosef et al. 2018). Although land cover changes have potential impact on precipitation, rainfall is mostly influenced by climate changes and land cover is straightly connected with water consumption processes, i.e., ET (Lian et al. 2021). China and India are leading the greening phenomenon (Chen et al. 2019a). In India, cropland expansion from bare soil or desert grass is in charge of bulk of the ET increase and water consumed in this process can is equivalent to almost ten times the area of the degraded desert grass ecosystem recovery water amounts (Das et al. 2018). Stricter policy to limit cropland expansion is expected in sustainable ecosystem development planning (Das et al. 2018; Zeng et al. 2019). Considering the consequences of India polices, countries shouldn’t neglect the significant ET changes accompanying land cover transitions in implementing different future dryland development plans, such as China and Europe for ecological protection with restoration plans, India for food security with cropland expansion plans as well as social and economic development plans with urbanization (Teuling et al. 2019; Wu et al. 2014). ET is the priority element in scheduling the allocations of dryland water resources, which is recommended in China’s future water resources policies (Wu et al. 2021a).
4 Driving Forces of Dryland Changes
Climate change and human activities are widely considered as major driving factors of dryland dynamics and have intensified the risk of desertification and land degradation in drylands (Li et al. 2021a; Ruppert et al. 2015; Stringer et al. 2021). Climate change mainly refers to the intensifying interannual and interdecadal variability of precipitation and temperature in drylands under the impact of global warming, with wetter weather in the American drylands and drier weather in Eastern Hemisphere drylands as the modelling result of Huang et al. (2017). High variability of precipitation and temperature means more extreme droughts and more extreme rainfall in dryland regions, which further exacerbates water stress and water competition among dryland ecosystem components, reduces the stability and resilience of dryland ecosystem, thus elevates the risk of desertification and land degradation in dryland regions. Human activities mainly include agricultural development, overgrazing, urbanization, and ecological restoration. In the drylands of northern China, large-scale ecological conservation and restoration projects are the main drivers of local dryland dynamics. Such measures reflect the influence of government policies on the dynamic evolution of drylands, enhancing local greening and ecosystem services, but also imposing significant water stress in China’s drylands (Li et al. 2021a). Agricultural development and grazing are the most widely distributed human activities, which are more prone to lead land degradation in drylands owing to the low productivity nature of dryland ecosystem (Reynolds et al. 2007). Further, the extent of farmland and pastures are mainly driven by population and agricultural efficiency. Changed land tenure systems and consumption preferences play a key role in reducing the risk of land degradation and food insecurity as well (Stehfest et al. 2019). Understanding the driving mechanisms and extent of natural and human forces to dryland change at multiple spatial scales, and the dynamics of dryland ecosystems under their influence is critical to the sustainable management of dryland ecosystems (Liu et al. 2015a).
4.1 Climate Change
As the monitoring results show, global drylands have expanded in the last 60 years and would continue to expand in the twenty-first century (Huang et al. 2017, 2016; Koutroulis 2019; Spinoni et al. 2021). Long-term monitoring also indicates that global drylands are in a state of accelerated expansion (Huang et al. 2016). Evidences have shown that global dryland extent had increased by 4% between 1991 and 2005 (Feng and Fu 2013), and the modeling for future climate scenarios has shown that drylands will experience increase in aridity (Yuan et al. 2019), and more frequent and more severe extreme events, such as extreme droughts and extreme precipitation (Sloat et al. 2018; Gampe et al. 2021; Zhang et al. 2021; Berdugo et al. 2020). As a result, global dryland areas are expected to increase by 11–23% by the end of this century (Huang et al. 2016; Prăvălie 2016). Global drylands are changing in extent, structure, and function, and could expand by 23% by the end of the century under a pessimistic future climate scenario (RCP 8.5), coupling with the impact of aridity (Berdugo et al. 2020), implying that drylands will account for 56% of global area (Huang et al. 2016). However, it is also argued that current projection of dryland expansion is overestimated, which means global drylands will not expand as significantly under a warming climate (Berg and McColl 2021).
Dryland expansion will reduce soil water content, soil organic carbon and gross primary production, further intensifying regional warming, resulting in a warming trend in drylands that is twice that of wet areas (Huang et al. 2016). The warming could lead higher evaporative demand and less soil moisture, which further cause an increased sensible heat flux and declined latent heat flux, meaning a strong impact on temperature extremes (Seneviratne et al. 2014). Thus, the simulated warming trends in drylands is more significant than that in wet areas. Moreover, expanded dryland may result in a declined soil organic carbon storage, as studies show that soil organic carbon storage would decrease with increase temperature and decrease with decline soil moisture (Huang et al. 2016). Furthermore, the lost soil organic carbon would increase emit CO2 into atmosphere and the declined soil moisture would suppress the photosynthesis activities of plants and gross primary production accumulation, both of which may further reinforce the warming and form positive feedback (Huang et al. 2016).
Extreme climate events such as droughts and heat waves significantly increase the risk of negative changes in dryland ecosystem dynamics and reduce the resilience of dryland ecosystem (Zhang et al. 2021; Gampe et al. 2021; Yuan et al. 2019; Sloat et al. 2018). Aridification leads to systematic and abrupt changes in the ecosystem structure and function, including plant productivity, soil fertility, plant cover, and richness (Berdugo et al. 2020). Warming may reduce soil water availability (Schlaepfer et al. 2017), soil fertility, plant productivity (Berdugo et al. 2020), leaf abundance, and species diversity (Maestre et al. 2016) in dryland ecosystems. The most obvious soil drying occurred over transitional areas between dry and wet regions (Cheng and Huang 2016). Furthermore, the decrease in soil moisture and intense droughts would expand the major deserts in the world, including the Sahara, Arabian, Kalahari, Gobi, and Great Sandy Desert (Zeng and Yoon 2009).
However, desertification is not a global trend as vegetation cover and rainfall are increasing in many drylands. Evidence from time series analysis of satellite images shows some arid lands, such as the Sahel and the Mediterranean basin, as well as China Loess plateau (Li et al. 2021a; Chen et al. 2019a; Gonsamo et al. 2021), are currently greening up (Hellden and Tottrup 2008). This obvious inconsistency is due to different interpretations of aridity, whether in an atmospheric, agricultural, hydrologic, or ecological context. In the semi-arid regions of China, vegetation greening leads to an increase in net primary productivity (NPP), attributing to increased atmospheric CO2 concentrations (i.e., CO2 fertilization) (Gonsamo et al. 2021). However, CO2 fertilization greening increases ET and thus decreases soil moisture in this region (Feng et al. 2016), making existing ecosystem hardly sustainable because the declining soil moisture will limit primary productivity (Peng et al. 2013) and affect photosynthesis in plants that can absorb CO2 and store carbon, especially in C4 plants with high photosynthesis levels (Li et al. 2021a), as well as shape the patterns of plant species richness (Sun et al. 2021).
4.2 Agricultural Development
Agricultural development is another important factor contributing to the degradation of dryland ecosystems, which consists mainly of overgrazing or unsustainable agricultural practices that exceed the limits allowed by the fragile environment. Cropland expansion has serious adverse effects on biodiversity and carbon stocks through habitat loss and fragmentation (Molotoks et al. 2020, 2018), whereas it provides necessary food for the growing population. However, although unreasonable agricultural measures may temporarily obtain higher input returns, they are at the expanse of ecosystem stability and are not sustainable. The increased frequency of droughts poses a significant risk to agriculture as well, with different impacts on the water balance and crop cropping systems (Sivakumar et al. 2005). Further, overgrazing has been considered to be related to grassland degradation and shrub encroachment in many dryland regions (Gaitan et al. 2018; Fredrickson et al. 2006).
In dryland regions, one of the major unsustainable agricultural practices is the overdevelopment of irrigation infrastructures with the aims of increasing crop or livestock production (Geist 2017). These practices are often based on the misconception that livestock production or crop yield in dryland regions is limited by a lack of water for drinking or irrigation. However, the development of these irrigation systems without considering the vulnerability of dryland soil and water availability intensifies the use of land resource, which may further lead to desertification (Niamir-Fuller 1999; Zeng et al. 2019). Moreover, the construction of these infrastructures limits the mobility of herbs and crop cultivations, thereby reducing the restoration ability of land rotation schemes (Wang and D'Odorico 2008). Furthermore, the initial investments in these infrastructures reduce the short-term gains, which enhances the intensive use of the land. As a result, these investments may lead the entire ecosystem toward an unstable state.
In addition, other unsustainable agricultural practices include residue removal, continuous cropping with limited inputs, and cultivating soils that are marginal for crop production (Blanco-Canqui and Lal 2009), which further degrade the land quality in drylands. Crop residues are used as fuel or fodder, which would accelerate the loss of soil carbon and increase soil erosion. Erosion reduces soil fertility by removing nutrients and organic matter from the land, and degraded soils are not conducive to water infiltration, thus leading to a continued decline in crop productivity and soil quality.
Overgrazing and grazing abandonment also play an important role in the degradation of dryland ecosystems. In the Mediterranean region, overgrazing has led to land degradation in Spain, Greece, and Cyprus (Gaitan et al. 2018; Peco et al. 2017; Concostrina-Zubiri et al. 2017; Riva et al. 2017). In China, both drought and overgrazing are regarded as the major drivers of the decline in grassland biodiversity, ecosystem functions, and services in the northern arid and semi-arid zones (Li et al. 2021a), with overgrazing having a greater negative impact on plant species richness, plant productivity, and soil carbon content than drought (Peco et al. 2017). Some researches indicate that overgrazing is one of the main factors to the shrub encroachment in grassland in arid and semi-arid regions as well (van de Koppel et al. 2002; Fredrickson et al. 2006). Proper grazing speeds up the reproduction and regeneration of vegetation, thus has a positive impact to grassland ecosystems. However, on the one hand, overgrazing will increase the livestock load of grassland and reduce the structure and stability of grassland ecosystem. On the other hand, it tends to alter the community structure of the grassland ecosystem and increase the proportion of shrub by prefer to eat herbaceous and spread shrub seeds (Fredrickson et al. 2006). In contrast, some studies also point out overgrazing may not be the direct cause of shrub encroachment. It is the grazing abandonment after long term overgrazing who triggers shrub encroachment in grassland rather than overgrazing (Sanjuán et al. 2018), yet the mechanism behind it is still unclear. Moreover, the impact of grazing abandonment on dryland ecosystems might not be the expected positive. Studies have shown that although grazing abandonment may have a positive impact on the increased cover of soil crusts (Concostrina-Zubiri et al. 2017), it may also have a negative impact on soil fertility, carbon storage, soil multifunctionality, and soil microbial activity (Peco et al. 2017). In addition, studies have shown that wildfires in abandoned terraces could be the primary reason for severe soil degradation in the Mediterranean (Lucas-Borja et al. 2018) and that fires may be the main driver of the transition from Mediterranean oak woodlands to shrublands (Guiomar et al. 2015).
4.3 Urbanization
Monitoring of global drylands shows that the region is maintaining rapid urbanization despite the lack of water and other essential natural resources. The trend of rapid urbanization would be maintained until at least year 2040 as the modelling result shows, which may have a significant negative impact on food production (Chen et al. 2020a). Rapid urban expansion had significantly negative impacts on environmental sustainability, leading to a decline in the comprehensive index of environmental sustainability in northern China (Liu et al. 2019b). Moreover, large scale urban development tends to occupy the original farmland, forestland, and shrubland, which leads to the fragmentation of ecosystem landscapes, resulting in the loss of ecosystem service value (Chen et al. 2020b) and ecosystem carbon stock (Yan et al. 2016). As a result, assessing the environmental impacts of urban expansion is highly important for the sustainable development of drylands.
In the drylands of northern China, rapid urbanization has led to serious challenges to sustainable development due to its influence to ecology and the environment. The water demand in this region accelerated by 50.75% from the 1980s to 2010 (Liu et al. 2015b), while the per capita water resources in the region are very limited, at 68% of the national average. Moreover, the land area undergoing desertification increased to 2.636 million km2 in 2010 (Xu et al. 2014), while the NPP of grasslands is experiencing a steady decline (Tian and Qiao 2014). Although urbanization poses a threat to the ecosystem stability and water resources tension in some drylands, it also brings more employment opportunities for people living there. Land is no longer the only way for local people to invest in labor. For example, the poverty alleviation projection has greatly reduced the incidence of poverty in the arid areas of Northwest China. People have more opportunities for employment, study and medical treatment, and the quality of life has been improved. In addition, studies indicate that climate change will be the key factor affecting urban expansion in this region, and the area of urban land affected by climate change is expanding (Liu et al. 2019a). To improve the sustainability of cities, effective measures to mitigate and adapt to the impacts of climate change on urban expansion are required.
4.4 Population Increase and Poverty Alleviation
Drylands hold to more than 38% of the global population (Reynolds et al. 2007), and increasing population pressure means that the impact of human activities on dryland ecosystems will continue unabated. Population growth has a number of consequences, such as increased demand for food and the expansion and intensification of crop/livestock production systems (Liao et al. 2020). Crop and livestock production is shifting from a relatively extensive, low-input/output production mode to a more intensive, higher-input/output production mode (Powell et al. 2004), which will increase the burden on dryland ecosystem services. Once the limits of ecosystem services provided by dryland are exceeded, irreversible ecosystem collapse may occur. Some studies also indicate the increased demand for agricultural land and forest products from population growth may offset the increased vegetation cover driven by increased CO2 and precipitation in sub-Saharan Africa (Brandt et al. 2017).
By 2015, 700 million people still lived below the extreme poverty line (Steele et al. 2017). Eradicating extreme poverty and hunger are the first two of the sustainable development goals (SDGs) (Assembly 2015). Developing agriculture and increasing land productivity can directly increase the income of the poor and are essential to achieve the SDG2-zero hunger initiative (Mason-D’Croz et al. 2019). Studies have shown that increasing the crop productivity of land (Pingali 2012) and expanding arable land area are effective methods of reducing poverty rates. However, for fragile dryland ecosystems, the ecological problems caused by agricultural development cannot be ignored. On the one hand, the expansion of arable land leads to the reduction of woodlands and grasslands, crucial for soil and water conservation. On the other hand, the water resources consumed by agriculture further exacerbate the water stress in drylands and intensify the instability of dryland ecosystems. This will increase the cost of local ecological compensation (McLeman et al. 2014). Therefore, sustainable agricultural development is an effective way to address population growth and poverty reduction in dryland regions. In China, labor transfer and relocation have also proved to be effective means of poverty alleviation. By transferring the rural idle surplus labor, land is no longer the only way for people’s labor force investment in drylands, and the local ecological pressure has been alleviated. Relocation is one of the ways of land consolidation. Relocation not only increases the employment opportunities for the people in remote areas, but also improves the stability and resilience of dryland ecosystem by ecological restoration after relocation.
5 Prediction of Dryland Changes
In the future, the world will face multiple interrelated challenges, including climate change, global income levels and population increases, as well as an increase in food consumption. Therefore, the simulation of future land use will play a vital role in achieving sustainable development goals through more rational land use management and effective global policy and technology improvement (Popp et al. 2017). Modeling is the major way to predict potential land dynamics in the future, such as pattern formation and ecosystem responses to disturbances. Scenario-based land use simulation model could assess the overall situation of future land use based on various scenarios (Liao et al. 2020; Chen et al. 2020a), which is different estimation of future population growth, economic development, and other socioeconomic and environmental variables. However, the spatial distribution of various land use is hard to determine. In contrast, the spatially explicit assignment of predicted land use changes to specific grid cells involves a complex process (Li et al. 2017; Liang et al. 2021), during which the relationship between land use changes and their driving forces must be accurately determined. Existing literature has successfully applied cellular automata, Markov chains, and neural networks to the spatial prediction of land use change and designed a series of models, such as the future land use simulation (FLUS) (Li et al. 2017) and patch-generating land use simulation (PLUS) model (Liang et al. 2021), as well as surface modeling of land cover change (SMLC) and dynamic conversion of land use and its effects (Dyna-CLUE) models (Sun et al. 2012). Although several land use change simulation products have been proposed based on the existing scenario data or actual models (Li et al. 2017; Dong et al. 2018), few concentrates on the land cover projection in dryland regions considered to be the Earth’s largest biome.
Population growth, climate change, and economic development will certainly alter land use structure and pattern in future. Based on the IPCC SRES scenarios, Li et al. (2017) produced a high-resolution future land use simulation product with a human–environment interaction system. In this simulation, the farmland area is more related to population, which means increase farmland are more likely to emerge in general and greater increase would occur in an economic emphasizing future. Simulation result Liao et al. (2020) support the same view, in which the cropland in North China Plain is expected to expansion due to rapid population growth and demand for biomass energy. The increased farmland comes at the cost of natural landscapes (Li et al. 2017). Thus, both grassland and forest areas experience significant losses. Further, future changes in pasture in the drylands of western China are highly dependent on changes in the demand for animal products and the efficiency of livestock production systems (Liao et al. 2020). Future pasture shrinkage is most pronounced under scenarios of lower demand and higher efficiency of livestock production.
However, there are still some defects in the model used. Firstly, the description of relationship between driving factors and land use change is too simple. The response of land use to changes in social-economical-environmental conditions are view to be very complicated and follow multiple linear and non-linear patterns with abrupt and gradual shifts. But in the models, this complex correlation is simply descripted by nonlinear, multivariate regression or machine learning based statistical technology. Secondly, the description variable for driving factors is too less. For example, in the simulation of dryland ecosystems, existing models, such as FLUS (Li et al. 2017) and PLUS (Liang et al. 2021), often use indicators of accessibility, such as the distance to settlements and the density of the road network, to reflect the intensity of human activities in the study area. This is logical in practice in that the areas with poor accessibility generally have small populations, weak human activities, and thus low impacts on ecosystems. However, the simple use of accessibility as a factor to characterize the intensity of human activity hinders further improvement of model simulation accuracy, so it is necessary to design more other indicators or their combinations to describe the impacts from various driving factors.
Moreover, the coupling of land use simulation models with other models provides an effective way to study the changes in ecosystem structure, services, and processes in drylands under various scenarios in the future. Coupled PLUS and GMOP models were used to analyze land use changes and their resulting changes in ecosystem service values under different scenarios (Li et al. 2021b), while the trade-offs and synergistic relationships of ecosystem services under different scenarios were explored in Yili Valley based on the coupling of PLUS and InVEST models (Shi et al. 2021). However, the uncertainty of data source and model leads to uncertainty of simulation results. How to eliminate and evaluate this uncertainty becomes very important.
6 Perspective
Dryland land cover, as a geographic feature and physical land type, is the synthesis of observable natural and artificial objects within water limited regions of the land surface of the Earth. As one of the significant and special land cover in drylands, vegetation pattern can not only intercept and retain essential resources, indicate the health of local landscape (Ludwig et al. 2007), but also have the potential to indicate the tipping of ecosystem and improve the resilience of ecosystem (Rietkerk et al. 2021; Scheffer et al. 2009). As one of the most characteristic biological landscapes in dryland region, soil biocrusts are the pioneer species of desert vegetation succession and play an important role in promoting the evolution of desert vegetation (Qi et al. 2021; Zhao et al. 2021). The establishment of photovoltaic systems in dryland region is a growing impact of human activities on dryland ecosystems and its impact on local climate and ecosystem function must be more comprehensively determined with future research (Kruitwagen et al. 2021). However, all the issues mentioned above are subject to further study with more and finer resolution data, including field survey and remote sensing.
Dryland ecosystems are water-limited, which is obvious as dryland plants tend to maximize water usage during short rainfall windows and optimize the ratio of the plant growth rate to water consumption (Stringer et al. 2021). Remote sensing technology provides new insights into the calculation of the basin water balance equation and human available water for consumption after the separation of human induced and natural water consumption (Wu et al. 2018). Land cover changes exert a strong impact on the hydrological system, as they can change the amount of ET and precipitation, which significantly alters the water system input and consumption (Lian et al. 2021).
The dynamics of drylands and the response of drylands to environmental change are complex. As mentioned above, environmental changes and irrational land use interact and together lead to the degradation of dryland ecosystems. Aridification due to global warming (Yuan et al. 2019) would lead to the expansion of dryland areas and the degradation of dryland ecosystems (Berdugo et al. 2020). In contrast, global aridification associated with rapid warming since the late 1970s is largely attributed to anthropogenic increases in greenhouse gas emissions (Edenhofer et al. 2013). Mediated by the complex direct and indirect factors, the future trajectory of drylands dynamic remains unclear and is an active research field in the context of rapid climate change and intensifying human activities. Although a variety of models have been proposed and applied to simulate land use transformations, there is still a high uncertainty across models and scenarios (Molotoks et al. 2020). In addition, although models such as structural equations and multiple response-diffusion models have been used to explain the drivers of impact factors on dryland dynamics (von Hardenberg et al. 2001; Li et al. 2021a), the mechanisms by which each driver mediates changes in land use dynamics are still largely unknown. Understanding these complex and interacting drivers of dryland dynamics has a long way to go.
References
Abrams M, Kubiske M, Steiner K (1990) Drought adaptations and responses in five genotypes of fraxinus pennsylvanica marsh: photosynthesis, water relations and leaf morphology. Tree Physiol 6(3):305–315
Ahlström A, Raupach MR, Schurgers G et al (2015) The dominant role of semi-arid ecosystems in the trend and variability of the land CO2 sink. Science 348(6237):895–899
Amani M, Mahdavi S, Bullock T et al (2020) Automatic nighttime sea fog detection using goes-16 imagery. Atmos Res 238:104712. https://doi.org/10.1016/j.atmosres.2019.104712
Andersen GL (2006) How to detect desert trees using corona images: discovering historical ecological data. J Arid Environ 65(3):491–511. https://doi.org/10.1016/j.jaridenv.2005.07.010
Andrade JL (2003) Dew deposition on epiphytic bromeliad leaves: an important event in a mexican tropical dry deciduous forest. J Trop Ecol 19(5):479–488
Antoninka A, Faist A, Rodriguez-Caballero E et al (2020) Biological soil crusts in ecological restoration: emerging research and perspectives. Restor Ecol 28(S2):S3–S8. https://doi.org/10.1111/rec.13201
Arnds D, Böhner J, Bechtel B (2017) Spatio-temporal variance and meteorological drivers of the urban heat island in a European city. Theoret Appl Climatol 128(1):43–61
Assembly G (2015) Sustainable development goals. SDGs Transform Our World 2030
Bacelar EA, Santos DL, Moutinho-Pereira JM et al (2006) Immediate responses and adaptative strategies of three olive cultivars under contrasting water availability regimes: changes on structure and chemical composition of foliage and oxidative damage. Plant Sci 170(3):596–605
Barron-Gafford GA, Minor RL, Allen NA et al (2016) The photovoltaic heat island effect: larger solar power plants increase local temperatures. Sci Rep 6(1):35070
Bastin GN, Ludwig JA, Eager RW et al (2002) Indicators of landscape function: comparing patchiness metrics using remotely-sensed data from rangelands. Ecol Ind 1(4):247–260
Belnap J (2003) The world at your feet: desert biological soil crusts. Front Ecol Environ 1(4):181–189
Belnap J, Weber B, Büdel B (2016) Biological soil crusts as an organizing principle in drylands. Biological soil crusts: an organizing principle in drylands. Springer, Switzerland, pp 3–13
Benvenutto-Vargas VP, Ochoa-Hueso R (2020) Effects of nitrogen deposition on the spatial pattern of biocrusts and soil microbial activity in a semi-arid mediterranean shrubland. Funct Ecol 34(4):923–937
Berbel J, Esteban E (2019) Droughts as a catalyst for water policy change. Analysis of Spain, Australia (mdb), and California. Glob Environ Change 58:101969
Berdugo M, Delgadobaquerizo M, Soliveres S et al (2020) Global ecosystem thresholds driven by aridity. Science 367(6479):787–790
Berg A, McColl KA (2021) No projected global drylands expansion under greenhouse warming. Nat Clim Chang 11(4):331–337
Biederman JA, Scott RL, Bell TW et al (2017) Co2 exchange and evapotranspiration across dryland ecosystems of southwestern north America. Glob Change Biol 23(10):4204–4221
Blanco-Canqui H, Lal R (2009) Crop residue removal impacts on soil productivity and environmental quality. Crit Rev Plant Sci 28(3):139–163. https://doi.org/10.1080/07352680902776507
Bolton H Jr, Smith J, Link S (1993) Soil microbial biomass and activity of a disturbed and undisturbed shrub-steppe ecosystem. Soil Biol Biochem 25(5):545–552
Bonan GB (2008) Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science 320(5882):1444–1449
Bonell M, Williams J (1986) The two parameters of the philip infiltration equation: their properties and spatial and temporal heterogeneity in a red earth of tropical semi-arid Queensland. J Hydrol 87(1–2):9–31
Bosmans JHC, van Beek LPH, Sutanudjaja EH et al (2017) Hydrological impacts of global land cover change and human water use. Hydrol Earth Syst Sci 21(11):5603–5626. https://doi.org/10.5194/hess-21-5603-2017
Brandt M, Rasmussen K, Penuelas J et al (2017) Human population growth offsets climate-driven increase in woody vegetation in Sub-saharan Africa. Nat Ecol Evolut 1(4):0081. https://doi.org/10.1038/s41559-017-0081
Bräutigam A, Schlüter U, Eisenhut M et al (2017) On the evolutionary origin of cam photosynthesis. Plant Physiol 174(2):473–477. https://doi.org/10.1104/pp.17.00195
Burkle LA, Marlin JC, Knight TM (2013) Plant-pollinator interactions over 120 years: loss of species, co-occurrence, and function. Science 339(6127):1611–1615
Celebi SZ, Demir S, Celebi R, et al (2010) The effect of arbuscular mycorrhizal fungi (amf) applications on the silage maize (zea mays l.) yield in different irrigation regimes. Europ J Soil Biol 46(5):302–305
Chamizo S, Cantón Y, Miralles I et al (2012) Biological soil crust development affects physicochemical characteristics of soil surface in semiarid ecosystems. Soil Biol Biochem 49:96–105
Chen J, Yuan Zhang M, Wang L et al (2005) A new index for mapping lichen-dominated biological soil crusts in desert areas. Remote Sens Environ 96(2):165–175. https://doi.org/10.1016/j.rse.2005.02.011
Chen J, Chen J, Liao A et al (2015) Global land cover mapping at 30m resolution: a pok-based operational approach. ISPRS J Photogramm Remote Sens 103:7–27. https://doi.org/10.1016/j.isprsjprs.2014.09.002
Chen C, Park T, Wang X et al (2019a) China and India lead in greening of the world through land-use management. Nat Sustain 2(2):122–129
Chen Y, Li B, Fan Y et al (2019b) Hydrological and water cycle processes of inland river basins in the arid region of northwest China. J Arid Land 11(2):161–179. https://doi.org/10.1007/s40333-019-0050-5
Chen Y, Xu C-Y, Chen X et al (2019c) Uncertainty in simulation of land-use change impacts on catchment runoff with multi-timescales based on the comparison of the hspf and swat models. J Hydrol 573:486–500. https://doi.org/10.1016/j.jhydrol.2019.03.091
Chen G, Li X, Liu X et al (2020a) Global projections of future urban land expansion under shared socioeconomic pathways. Nat Commun 11(1):537. https://doi.org/10.1038/s41467-020-14386-x
Chen S, Feng Y, Tong X et al (2020b) Modeling esv losses caused by urban expansion using cellular automata and geographically weighted regression. Sci Total Environ 712:136509. https://doi.org/10.1016/j.scitotenv.2020.136509
Cheng S, Huang J (2016) Enhanced soil moisture drying in transitional regions under a warming climate. J Geophys Res Atmos 121(6):2542–2555
Chopping M, Su L, Rango A et al (2008) Remote sensing of woody shrub cover in desert grasslands using misr with a geometric-optical canopy reflectance model. Remote Sens Environ 112(1):19–34. https://doi.org/10.1016/j.rse.2006.04.023
Ci L, Yang X (2010) Desertification and its control in china. Springer, Heidelberg
Concostrina-Zubiri L, Molla I, Velizarova E et al (2017) Grazing or not grazing: implications for ecosystem services provided by biocrusts in mediterranean cork oak woodlands. Land Degrad Dev 28(4):1345–1353
Couradeau E, Giraldo-Silva A, De Martini F et al (2019) Spatial segregation of the biological soil crust microbiome around its foundational cyanobacterium, microcoleus vaginatus, and the formation of a nitrogen-fixing cyanosphere. Microbiome 7(1):55. https://doi.org/10.1186/s40168-019-0661-2
Couteron P, Lejeune O (2001) Periodic spotted patterns in semi-arid vegetation explained by a propagation-inhibition model. J Ecol 89(4):616–628
Dakos V, Kéfi S, Rietkerk M et al (2011) Slowing down in spatially patterned ecosystems at the brink of collapse. Am Nat 177(6):E153–E166
Dannenberg MP, Wise EK, Smith WK (2019) Reduced tree growth in the semiarid united states due to asymmetric responses to intensifying precipitation extremes. Sci Adv 5(10):eaaw0667
Darrouzet-Nardi A, Reed SC, Grote EE et al (2015) Observations of net soil exchange of co2 in a dryland show experimental warming increases carbon losses in biocrust soils. Biogeochemistry 126(3):363–378
Darrouzet-Nardi A, Reed SC, Grote EE et al (2018) Patterns of longer-term climate change effects on co2 efflux from biocrusted soils differ from those observed in the short term. Biogeosciences 15(14):4561–4573. https://doi.org/10.5194/bg-15-4561-2018
Das P, Behera MD, Patidar N et al (2018) Impact of lulc change on the runoff, base flow and evapotranspiration dynamics in eastern indian river basins during 1985–2005 using variable infiltration capacity approach. J Earth Syst Sci 127(2):1–19
Davidson EA, de Araújo AC, Artaxo P et al (2012) The amazon basin in transition. Nature 481(7381):321–328
Deines JM, Kendall AD, Hyndman DW (2017) Annual irrigation dynamics in the us northern high plains derived from landsat satellite data. Geophys Res Lett 44(18):9350–9360
Delgadobaquerizo M, Maestre FT, Gallardo A et al (2013) Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 502(7473):672–676
Dichio B, Xiloyannis C, Sofo A et al (2006) Osmotic regulation in leaves and roots of olive trees during a water deficit and rewatering. Tree Physiol 26(2):179–185
D’Odorico P, Bhattachan A (2012) Hydrologic variability in dryland regions: impacts on ecosystem dynamics and food security. Philos Trans R Soc B-Biol Sci 367(1606):3145–3157. https://doi.org/10.1098/rstb.2012.0016
D’Odorico P, Porporato A, Runyan CW (2006) Dryland ecohydrology, vol 9. Springer, Heidelberg
Döll P, Siebert S (2002) Global modeling of irrigation water requirements. Water Resourc Res 38(4):8–1–8–10
Dong N, You L, Cai W et al (2018) Land use projections in China under global socioeconomic and emission scenarios: utilizing a scenario-based land-use change assessment framework. Glob Environ Chang 50:164–177. https://doi.org/10.1016/j.gloenvcha.2018.04.001
Donohue RJ, Roderick ML, McVicar TR et al (2013) Impact of CO2 fertilization on maximum foliage cover across the globe’s warm, arid environments. Geophys Res Lett 40(12):3031–3035
Duan Z, Bastiaanssen WGM (2013) First results from version 7 trmm 3b43 precipitation product in combination with a new downscaling–calibration procedure. Remote Sens Environ 131:1–13. https://doi.org/10.1016/j.rse.2012.12.002
Edenhofer O, Seyboth K, Shogren J (2013) Intergovernmental panel on climate change (ipcc). In: Encyclopedia of energy, natural resource, and environmental economics (volume 1: Energy), vol 1. Elsevier, pp 48–56
Elagib NA, Khalifa M, Babker Z et al (2021) Demarcating the rainfed unproductive zones in the african sahel and great green wall regions. Land Degrad Dev 32(3):1400–1411
Elbert W, Weber B, Burrows S et al (2012) Contribution of cryptogamic covers to the global cycles of carbon and nitrogen. Nat Geosci 5(7):459–462. https://doi.org/10.1038/ngeo1486
Elnashar A, Zeng H, Wu B et al (2020) Downscaling trmm monthly precipitation using google earth engine and google cloud computing. Remote Sensing 12(23):3860
Evans J, Geerken R (2004) Discrimination between climate and human-induced dryland degradation. J Arid Environ 57(4):535–554
Ezcurra E (2006) Global deserts outlook. UNEP/Earthprint, Kenya
Fatchen TJ, Barker S (1979) Cyclic vegetation pattern in the southern simpson desert australia. Trans R Soc South Australia Incorporated 103:113–122
Feng S, Fu Q (2013) Expansion of global drylands under a warming climate. Atmos Chem Phys 13(19):10081–10094
Feng X, Fu B, Piao S et al (2016) Revegetation in china’s loess plateau is approaching sustainable water resource limits. Nat Clim Chang 6(11):1019–1022
Ferrenberg S, Tucker C, Reed SC (2017) Biological soil crusts: diminutive communities of potential global importance. Front Ecol Environ 15(3):160–167
Flores J, Jurado E (2003) Are nurse-protégé interactions more common among plants from arid environments? J Veg Sci 14(6):911
Fredrickson EL, Estell R, Laliberte A et al (2006) Mesquite recruitment in the chihuahuan desert: historic and prehistoric patterns with long-term impacts. J Arid Environ 65(2):285–295
Gaitan JJ, Bran DE, Oliva GE et al (2018) Aridity and overgrazing have convergent effects on ecosystem structure and functioning in patagonian rangelands. Land Degrad Dev 29(2):210–218. https://doi.org/10.1002/ldr.2694
Gampe D, Zscheischler J, Reichstein M et al (2021) Increasing impact of warm droughts on northern ecosystem productivity over recent decades. Nat Clim Chang 11(9):772–779
Gao Z, Shi W, Wang X et al (2020) Non-rainfall water contributions to dryland jujube plantation evapotranspiration in the hilly loess region of China. J Hydrol 583:124604. https://doi.org/10.1016/j.jhydrol.2020.124604
García-Palacios P, Gross N, Gaitán J et al (2018) Climate mediates the biodiversity–ecosystem stability relationship globally. Proc Natl Acad Sci 115(33):8400. https://doi.org/10.1073/pnas.1800425115
Geist H (2017) The causes and progression of desertification. Routledge, London
Giraldo-Silva A, Nelson C, Penfold C et al (2020) Effect of preconditioning to the soil environment on the performance of 20 cyanobacterial strains used as inoculum for biocrust restoration. Restor Ecol 28(S2):S187–S193. https://doi.org/10.1111/rec.13048
Goffner D, Sinare H, Gordon LJ (2019) The great green wall for the sahara and the sahel initiative as an opportunity to enhance resilience in sahelian landscapes and livelihoods. Reg Environ Change 19(5):1417–1428
Gonsamo A, Ciais P, Miralles DG et al (2021) Greening drylands despite warming consistent with carbon dioxide fertilization effect. Glob Change Biol 27(14):3336–3349
Grafton RQ (2017) Water reform and planning in the Murray–darling basin, Australia. Water Economics and Policy 3(03):1702001
Grafton RQ, Pittock J, Davis R et al (2013) Global insights into water resources, climate change and governance. Nat Clim Chang 3(4):315–321
Grafton RQ, Williams J, Perry C et al (2018) The paradox of irrigation efficiency. Science 361(6404):748–750
Gremer JR, Bradford JB, Munson SM et al (2015) Desert grassland responses to climate and soil moisture suggest divergent vulnerabilities across the southwestern united states. Glob Change Biol 21(11):4049–4062
Guiomar N, Godinho S, Fernandes PM et al (2015) Wildfire patterns and landscape changes in mediterranean oak woodlands. Sci Total Environ 536:338–352. https://doi.org/10.1016/j.scitotenv.2015.07.087
Hanasaki N, Kanae S, Oki T et al (2008) An integrated model for the assessment of global water resources—part 2: applications and assessments. Hydrol Earth Syst Sci 4(5):1027–1037
Hanasaki N, Fujimori S, Yamamoto T et al (2013) A global water scarcity assessment under shared socio-economic pathways–part 2: water availability and scarcity. Hydrol Earth Syst Sci 17(7):2393–2413
Hassell MP, Comins HN, Mayt RM (1991) Spatial structure and chaos in insect population dynamics. Nature 353(6341):255–258
He L, Chen JM, Pisek J et al (2012) Global clumping index map derived from the modis brdf product. Remote Sens Environ 119:118–130. https://doi.org/10.1016/j.rse.2011.12.008
Hellden U, Tottrup C (2008) Regional desertification: a global synthesis. Glob Planet Change 64(3–4):169–176. https://doi.org/10.1016/j.gloplacha.2008.10.006
Henschel JR, Seely MK (2008) Ecophysiology of atmospheric moisture in the Namib desert. Atmos Res 87(3):362–368. https://doi.org/10.1016/j.atmosres.2007.11.015
Her Y, Yoo S-H, Cho J et al (2019) Uncertainty in hydrological analysis of climate change: multi-parameter vs. multi-gcm ensemble predictions. Sci Rep 9(1):4974. https://doi.org/10.1038/s41598-019-41334-7
Hoover DL, Bestelmeyer B, Grimm NB et al (2020) Traversing the wasteland: a framework for assessing ecological threats to drylands. Bioscience 70(1):35–47. https://doi.org/10.1093/biosci/biz126
Huang L (2019) Advances and perspectives on soil water research in China’s Loess Plateau. Earth Sci Rev 199:102962
Huang J, Yu H, Guan X et al (2016) Accelerated dryland expansion under climate change. Nat Clim Chang 6(2):166–171
Huang J, Li Y, Fu C et al (2017) Dryland climate change: recent progress and challenges. Rev Geophys 55(3):719–778. https://doi.org/10.1002/2016rg000550
Jägermeyr J, Pastor A, Biemans H et al (2017) Reconciling irrigated food production with environmental flows for sustainable development goals implementation. Nat Commun 8(1):15900
Javadian M, Behrangi A, Smith WK et al (2020) Global trends in evapotranspiration dominated by increases across large cropland regions. Remote Sens 12(7):1221
Jeffries D, Klopatek J, Link S et al (1992) Acetylene reduction by cryptogamic crusts from a blackbrush community as related to resaturation and dehydration. Soil Biol Biochem 24(11):1101–1105
Jia R-l, Li X-r, Liu L-c et al (2014) Effects of sand burial on dew deposition on moss soil crust in a revegetated area of the tennger desert, northern China. J Hydrol 519:2341–2349. https://doi.org/10.1016/j.jhydrol.2014.10.031
Jia X, Shao M, Yu D et al (2019) Spatial variations in soil-water carrying capacity of three typical revegetation species on the loess plateau, china. Agr Ecosyst Environ 273:25–35
Juergens N (2013) The biological underpinnings of Namib desert fairy circles. Science 339(6127):1618–1621
Kabela ED, Hornbuckle BK, Cosh MH et al (2009) Dew frequency, duration, amount, and distribution in corn and soybean during SMEX05. Agric for Meteorol 149(1):11–24. https://doi.org/10.1016/j.agrformet.2008.07.002
Karnieli A (1997) Development and implementation of spectral crust index over dune sands. Int J Remote Sens 18(6):1207–1220
Karnieli A, Kidron GJ, Glaesser C et al (1999) Spectral characteristics of cyanobacteria soil crust in semiarid environments. Remote Sens Environ 69(1):67–75. https://doi.org/10.1016/S0034-4257(98)00110-2
Kaseke KF, Mills A, Esler KJ et al (2012) Spatial variation of “non-rainfall” water input and the effect of mechanical soil crusts on input and evaporation. Pure Appl Geophys 169(12):2217–2229
Kaseke KF, Wang L, Seely M (2017) Nonrainfall water origins and formation mechanisms. Sci Adv 3:e1603131. https://doi.org/10.1126/sciadv.1603131
Kefi S, Rietkerk M, Alados CL et al (2007) Spatial vegetation patterns and imminent desertification in mediterranean arid ecosystems. Nature 449(7159):213–217
Kilgore JS, Jacobsen AL, Telewski FW (2021) Hydraulics of pinus (subsection ponderosae) populations across an elevation gradient in the santa catalina mountains of southern arizona. Madroño 67(4):218–226
Klausmeier CA (1999) Regular and irregular patterns in semiarid vegetation. Science 284(5421):1826–1828
Klironomos JN (2003) Variation in plant response to native and exotic arbuscular mycorrhizal fungi. Ecology 84(9):2292–2301
Klopatek JM (1992) Cryptogamic crusts as potential indicators of disturbance in semi-arid landscapes. Ecological indicators. Springer, Boston, pp 773–786
Knauft RL, Arditti J (1969) Partial identification of dark 14co2 fixation products in leaves of cattleya (orchidaceae). New Phytol 68(3):657–661
Konings AG, Dekker SC, Rietkerk M et al (2011) Drought sensitivity of patterned vegetation determined by rainfall‐land surface feedbacks. J Geophys Res Biogeosci 116:G04008
Koutroulis AG (2019) Dryland changes under different levels of global warming. Sci Total Environ 655:482–511. https://doi.org/10.1016/j.scitotenv.2018.11.215
Kruitwagen L, Story KT, Friedrich J et al (2021) A global inventory of photovoltaic solar energy generating units. Nature 598(7882):604–610. https://doi.org/10.1038/s41586-021-03957-7
Lacombe G, Ribolzi O, de Rouw A et al (2016) Contradictory hydrological impacts of afforestation in the humid tropics evidenced by long-term field monitoring and simulation modelling. Hydrol Earth Syst Sci 20(7):2691–2704. https://doi.org/10.5194/hess-20-2691-2016
Lajtha K, Kolberg K, Getz J (1997) Ecophysiology of the saguaro cactus (carnegiea gigantea) in the saguaro national monument: relationship to symptoms of decline. J Arid Environ 36(4):579–590. https://doi.org/10.1006/jare.1996.0240
Lan S, Ouyang H, Wu L et al (2017) Biological soil crust community types differ in photosynthetic pigment composition, fluorescence and carbon fixation in Shapotou region of China. Appl Soil Ecol 111:9–16
Leblanc M, Tweed S, Van Dijk A et al (2012) A review of historic and future hydrological changes in the Murray-darling basin. Glob Planet Change 80–81:226–246. https://doi.org/10.1016/j.gloplacha.2011.10.012
Lehnert LW, Jung P, Obermeier WA et al (2018) Estimating net photosynthesis of biological soil crusts in the Atacama using hyperspectral remote sensing. Remote Sensing 10(6):891
Li P, Gao X (2021) The impact of photovoltaic power plants on ecological environment and climate: a literature review. Plateau Meteorol 40(3):702–710
Li B, Qin Y, Duan H et al (2011) Genome-wide characterization of new and drought stress responsive micrornas in Populus euphratica. J Exp Bot 62(11):3765–3779
Li X, Chen G, Liu X et al (2017) A new global land-use and land-cover change product at a 1-km resolution for 2010 to 2100 based on human–environment interactions. Ann Am Assoc Geogr 107(5):1040–1059. https://doi.org/10.1080/24694452.2017.1303357
Li Y, Chang J, Luo L et al (2018a) Spatiotemporal impacts of land use land cover changes on hydrology from the mechanism perspective using swat model with time-varying parameters. Hydrol Res 50(1):244–261. https://doi.org/10.2166/nh.2018.006
Li Y, Kalnay E, Motesharrei S et al (2018b) Climate model shows large-scale wind and solar farms in the sahara increase rain and vegetation. Science 361(6406):1019–1022
Li C, Fu B, Wang S et al (2021a) Drivers and impacts of changes in China’s drylands. Nat Rev Earth Environ 2(12):858–873. https://doi.org/10.1038/s43017-021-00226-z
Li C, Wu Y, Gao B et al (2021b) Multi-scenario simulation of ecosystem service value for optimization of land use in the Sichuan-yunnan ecological barrier, China. Ecol Ind 132:108328
Lian X, Piao S, Chen A et al (2021) Multifaceted characteristics of dryland aridity changes in a warming world. Nat Rev Earth Environ 2(4):232–250. https://doi.org/10.1038/s43017-021-00144-0
Liang X, Guan Q, Clarke KC et al (2021) Understanding the drivers of sustainable land expansion using a patch-generating land use simulation (plus) model: a case study in wuhan, China. Comput Environ Urban Syst 85:101569. https://doi.org/10.1016/j.compenvurbsys.2020.101569
Liao W, Liu X, Xu X et al (2020) Projections of land use changes under the plant functional type classification in different SSP-RCP scenarios in China. Sci Bull 65(22):1935–1947. https://doi.org/10.1016/j.scib.2020.07.014
Liu X, Ma K (2015) Plant functional traits-concepts, applications and future directions. Sci Sinica Vitae 45(4):325–339
Liu S, Wang T, Kang W et al (2015a) Several challenges in monitoring and assessing desertification. Environ Earth Sci 73(11):7561–7570. https://doi.org/10.1007/s12665-014-3926-x
Liu X, Shen Y, Guo Y et al (2015b) Modeling demand/supply of water resources in the arid region of northwestern China during the late 1980s to 2010. J Geog Sci 25(5):573–591
Liu Z, Yang Y, He C et al (2019a) Climate change will constrain the rapid urban expansion in drylands: a scenario analysis with the zoned land use scenario dynamics-urban model. Sci Total Environ 651:2772–2786
Liu ZF, Ding MH, He CY et al (2019b) The impairment of environmental sustainability due to rapid urbanization in the dryland region of northern China. Landsc Urban Plan 187:165–180. https://doi.org/10.1016/j.landurbplan.2018.10.020
Liu H, Gong P, Wang J et al (2020) Annual dynamics of global land cover and its long-term changes from 1982 to 2015. Earth Syst Sci Data 12(2):1217–1243. https://doi.org/10.5194/essd-12-1217-2020
Lu Z, Zhang Q, Miller PA, et al (2021) Impacts of large-scale sahara solar farms on global climate and vegetation cover. Geophys Res Lett 48(2):e2020GL090789. https://doi.org/10.1029/2020GL090789
Lucas-Borja ME, Calsamiglia A, Fortesa J et al (2018) The role of wildfire on soil quality in abandoned terraces of three mediterranean micro-catchments. CATENA 170:246–256. https://doi.org/10.1016/j.catena.2018.06.014
Ludwig JA, Eager RW, Bastin GN et al (2002) A leakiness index for assessing landscape function using remote sensing. Landscape Ecol 17(2):157–171
Ludwig JA, Bastin GN, Chewings VH et al (2007) Leakiness: a new index for monitoring the health of arid and semiarid landscapes using remotely sensed vegetation cover and elevation data. Ecol Ind 7(2):442–454
Lv M, Ma Z, Lv M (2018) Effects of climate/land surface changes on streamflow with consideration of precipitation intensity and catchment characteristics in the yellow river basin. J Geophys Res Atmos 123(4):1942–1958
Lv M, Ma Z, Li M et al (2019) Quantitative analysis of terrestrial water storage changes under the grain for green program in the yellow river basin. J Geophys Res Atmos 124(3):1336–1351. https://doi.org/10.1029/2018JD029113
Ma Z, Yan N, Wu B et al (2019) Variation in actual evapotranspiration following changes in climate and vegetation cover during an ecological restoration period (2000–2015) in the loess plateau, china. Sci Total Environ 689:534–545
Maestre FT, Quero JL, Gotelli NJ et al (2012) Plant species richness and ecosystem multifunctionality in global drylands. Science 335(6065):214–218
Maestre FT, Eldridge DJ, Soliveres S et al (2016) Structure and functioning of dryland ecosystems in a changing world. Annu Rev Ecol Evol Syst 47(1):215–237
Maharjan SK, Poorter L, Holmgren M et al (2011) Plant functional traits and the distribution of west african rain forest trees along the rainfall gradient. Biotropica 43(5):552–561
Malahayati M (2020) Achieving renewable energies utilization target in south-east Asia: progress, challenges, and recommendations. Electr J 33(5):106736
Malek E, McCurdy G, Giles B (1999) Dew contribution to the annual water balance in semi-arid desert valleys. J Arid Environ 42:71–80. https://doi.org/10.1006/jare.1999.0506
Mander L, Dekker SC, Li M et al (2017) A morphometric analysis of vegetation patterns in dryland ecosystems. R Soc Open Sci 4(2):160443. https://doi.org/10.1098/rsos.160443
Martin CE, von Willert DJ (2000) Leaf epidermal hydathodes and the ecophysiological consequences of foliar water uptake in species of crassula from the namib desert in southern Africa. Plant Biol 2(2):229–242. https://doi.org/10.1055/s-2000-9163
Mason-D’Croz D, Sulser TB, Wiebe K et al (2019) Agricultural investments and hunger in Africa modeling potential contributions to SDG2–zero hunger. World Dev 116:38–53
Masood M, Yeh PJF, Hanasaki N et al (2015) Model study of the impacts of future climate change on the hydrology of Ganges–Brahmaputra–Meghna basin. Hydrol Earth Syst Sci 19(2):747–770. https://doi.org/10.5194/hess-19-747-2015
McCabe GJ, Wolock DM (2007) Warming may create substantial water supply shortages in the colorado river basin. Geophys Res Lett 34(22)
McCollum DW, Tanaka JA, Morgan JA et al (2017) Climate change effects on rangelands and rangeland management: affirming the need for monitoring. Ecosyst Health Sustain 3(3):e01264. https://doi.org/10.1002/ehs2.1264
McHugh TA, Morrissey EM, Reed SC et al (2015) Water from air: an overlooked source of moisture in arid and semiarid regions. Sci Rep 5(1):1–6
McLeman RA, Dupre J, Ford LB et al (2014) What we learned from the dust bowl: lessons in science, policy, and adaptation. Popul Environ 35(4):417–440
Meng Y, Wen X (2016) Characteristics of dew events in an arid artificial oasis cropland and a sub-humid cropland in China. J Arid Land 8(3):399–408
Millstein D, Menon S (2011) Regional climate consequences of large-scale cool roof and photovoltaic array deployment. Environ Res Lett 6(3):034001
Mirzabaev A, Sacande M, Motlagh F, et al (2022) Economic efficiency and targeting of the african great green wall. Nat Sustain 5:17–25
Miskin CK, Li Y, Perna A et al (2019) Sustainable co-production of food and solar power to relax land-use constraints. Nat Sustain 2(10):972–980
Moat J, Orellana-Garcia A, Tovar C et al (2021) Seeing through the clouds—mapping desert fog oasis ecosystems using 20 years of Modis imagery over Peru and Chile. Int J Appl Earth Obs Geoinf 103:102468. https://doi.org/10.1016/j.jag.2021.102468
Mohsin M, Rasheed A, Sun H et al (2019) Developing low carbon economies: an aggregated composite index based on carbon emissions. Sustain Energy Technol Assess 35:365–374
Molotoks A, Stehfest E, Doelman J et al (2018) Global projections of future cropland expansion to 2050 and direct impacts on biodiversity and carbon storage. Glob Change Biol 24(12):5895–5908. https://doi.org/10.1111/gcb.14459
Molotoks A, Henry R, Stehfest E et al (2020) Comparing the impact of future cropland expansion on global biodiversity and carbon storage across models and scenarios. Philos Trans R Soc Lond B Biol Sci 375(1794):20190189. https://doi.org/10.1098/rstb.2019.0189
Munné-Bosch S, Alegre L (1999) Role of dew on the recovery of water-stressed Melissa officinalis l. Plants. J Plant Physiol 154(5–6):759–766
Naha S, Rico-Ramirez MA, Rosolem R (2021) Quantifying the impacts of land cover change on hydrological responses in the Mahanadi river basin in India. Hydrol Earth Syst Sci 25(12):6339–6357. https://doi.org/10.5194/hess-25-6339-2021
Ngosong C, Buse T, Ewald M et al (2020) Influence of management intensity and environmental conditions on microbiota in biological soil crust and crust-free soil habitats of temperate forests. Soil Biol Biochem 144:107761. https://doi.org/10.1016/j.soilbio.2020.107761
Niamir-Fuller M (1999) Managing mobility in African rangelands. In: Property rights, risk, and livestock development in Africa. International Food Policy Research Institute (IFPRI), pp 102–131
Norton CL, Dannenberg MP, Yan D et al (2021) Climate and socioeconomic factors drive irrigated agriculture dynamics in the lower colorado river basin. Remote Sensing 13(9):1659
Okin GS, Heras MM, Saco PM, et al (2015) Connectivity in dryland landscapes: shifting concepts of spatial interactions. Front Ecol Environ 13(1):20–27. https://doi.org/10.1890/140163
Pascolini-Campbell M, Reager JT, Chandanpurkar HA et al (2021) A 10 per cent increase in global land evapotranspiration from 2003 to 2019. Nature 593(7860):543–547. https://doi.org/10.1038/s41586-021-03503-5
Peco B, Navarro E, Carmona CP et al (2017) Effects of grazing abandonment on soil multifunctionality: the role of plant functional traits. Agr Ecosyst Environ 249:215–225. https://doi.org/10.1016/j.agee.2017.08.013
Peng S, Piao S, Ciais P et al (2013) Asymmetric effects of daytime and night-time warming on northern hemisphere vegetation. Nature 501(7465):88–92
Perugini L, Caporaso L, Marconi S et al (2017) Biophysical effects on temperature and precipitation due to land cover change. Environ Res Lett 12(5):053002
Pingali PL (2012) Green revolution: impacts, limits, and the path ahead. Proc Natl Acad Sci U S A 109(31):12302–12308. https://doi.org/10.1073/pnas.0912953109
Pisinaras V, Wei Y, Bärring L et al (2014) Conceptualizing and assessing the effects of installation and operation of photovoltaic power plants on major hydrologic budget constituents. Sci Total Environ 493:239–250
Pointing SB, Belnap J (2012) Microbial colonization and controls in dryland systems. Nat Rev Microbiol 10(8):551–562. https://doi.org/10.1038/nrmicro2831
Popp A, Calvin K, Fujimori S et al (2017) Land-use futures in the shared socio-economic pathways. Glob Environ Chang 42:331–345. https://doi.org/10.1016/j.gloenvcha.2016.10.002
Pöschl U, Martin S, Sinha B et al (2010) Rainforest aerosols as biogenic nuclei of clouds and precipitation in the amazon. Science 329(5998):1513–1516
Powell JM, Pearson RA, Hiernaux PH (2004) Crop-livestock interactions in the west African drylands. Agron J 96(2):469–483. https://doi.org/10.2134/agronj2004.0469
Pradhan RK, Markonis Y, Vargas Godoy MR et al (2022) Review of GPM Imerg performance: a global perspective. Remote Sens Environ 268:112754. https://doi.org/10.1016/j.rse.2021.112754
Prăvălie R (2016) Drylands extent and environmental issues. A global approach. Earth-Sci. Rev. 161:259–278. https://doi.org/10.1016/j.earscirev.2016.08.003
Pringle RM, Doak DF, Brody AK et al (2010) Spatial pattern enhances ecosystem functioning in an African savanna. PLoS Biol 8(5):e1000377. https://doi.org/10.1371/journal.pbio.1000377
Qi J, Liu Y, Wang Z et al (2021) Variations in microbial functional potential associated with phosphorus and sulfur cycling in biological soil crusts of different ages at the Tengger desert, china. Appl Soil Ecol 165:104022. https://doi.org/10.1016/j.apsoil.2021.104022
Ray DK, Nair US, Lawton RO et al (2006) Impact of land use on Costa Rican tropical montane cloud forests: Sensitivity of orographic cloud formation to deforestation in the plains. J Geophys Res Atmos 111:D02108
Reddy AR, Chaitanya KV, Vivekanandan M (2004) Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J Plant Physiol 161(11):1189–1202
Reed SC, Coe KK, Sparks JP et al (2012) Changes to dryland rainfall result in rapid moss mortality and altered soil fertility. Nat Clim Chang 2(10):752–755
Reich PB, Oleksyn J (2004) Global patterns of plant leaf n and p in relation to temperature and latitude. Proc Natl Acad Sci 101(30):11001–11006
Reid WV, Mooney HA, Cropper A, et al (2005) Ecosystems and human well-being-synthesis: a report of the millennium ecosystem assessment. Island Press
Reynolds JF, Smith DMS, Lambin EF et al (2007) Global desertification: building a science for dryland development. Science 316(5826):847–851
Rietkerk M, Boerlijst MC, van Langevelde F et al (2002) Self-organization of vegetation in arid ecosystems. Am Nat 160(4):524–530
Rietkerk M, Dekker SC, De Ruiter PC et al (2004) Self-organized patchiness and catastrophic shifts in ecosystems. Science 305(5692):1926–1929
Rietkerk M, Bastiaansen R, Banerjee S, et al (2021) Evasion of tipping in complex systems through spatial pattern formation. Science 374(6564):eabj0359. https://doi.org/10.1126/science.abj0359
Riva MJ, Daliakopoulos IN, Eckert S et al (2017) Assessment of land degradation in mediterranean forests and grazing lands using a landscape unit approach and the normalized difference vegetation index. Appl Geogr 86:8–21
Rodríguez-Caballero E, Escribano P, Olehowski C et al (2017) Transferability of multi-and hyperspectral optical biocrust indices. ISPRS J Photogramm Remote Sens 126:94–107
Rodriguez-Caballero E, Belnap J, Büdel B et al (2018) Dryland photoautotrophic soil surface communities endangered by global change. Nat Geosci 11(3):185–189
Rogger M, Agnoletti M, Alaoui A et al (2017) Land use change impacts on floods at the catchment scale: challenges and opportunities for future research. Water Resour Res 53(7):5209–5219. https://doi.org/10.1002/2017WR020723
Root HT, Brinda JC, Dodson EK (2017) Recovery of biological soil crust richness and cover 12–16 years after wildfires in Idaho, USA. Biogeosciences 14(17):3957–3969. https://doi.org/10.5194/bg-14-3957-2017
Runyan C, D’Odorico P (2016) Global deforestation. Cambridge University Press, Cambridge
Ruppert JC, Harmoney K, Henkin Z et al (2015) Quantifying drylands’ drought resistance and recovery: the importance of drought intensity, dominant life history and grazing regime. Glob Change Biol 21(3):1258–1270. https://doi.org/10.1111/gcb.12777
Rutherford WA, Painter TH, Ferrenberg S et al (2017) Albedo feedbacks to future climate via climate change impacts on dryland biocrusts. Sci Rep 7(1):44188. https://doi.org/10.1038/srep44188
Safriel U, Adeel Z (2008) Development paths of drylands: thresholds and sustainability. Sustain Sci 3(1):117–123. https://doi.org/10.1007/s11625-007-0038-5
Sanchez AC, Haq N, Assogbadjo A (2010) Variation in baobab (adansonia digitata l.) leaf morphology and its relation to drought tolerance. Genet Resourc Crop Evolut 57(1):17–25
Sanjuán Y, Arnáez J, Beguería S et al (2018) Woody plant encroachment following grazing abandonment in the subalpine belt: a case study in northern Spain. Reg Environ Change 18(4):1103–1115
Scheffer M (2020) Critical transitions in nature and society, vol 16. Princeton University Press, Princeton
Scheffer M, Bascompte J, Brock WA et al (2009) Early-warning signals for critical transitions. Nature 461(7260):53–59. https://doi.org/10.1038/nature08227
Schimel DS (2010) Drylands in the earth system. Science 327(5964):418–419
Schlaepfer DR, Bradford JB, Lauenroth WK et al (2017) Climate change reduces extent of temperate drylands and intensifies drought in deep soils. Nat Commun 8(1):14196. https://doi.org/10.1038/ncomms14196
Schneebeli M, Wolf S, Kunert N et al (2011) Relating the x-band opacity of a tropical tree canopy to sapflow, rain interception and dew formation. Remote Sens Environ 115(8):2116–2125
Seneviratne SI, Donat MG, Mueller B et al (2014) No pause in the increase of hot temperature extremes. Nat Clim Chang 4(3):161–163
Shi M, Wu H, Fan X et al (2021) Trade-offs and synergies of multiple ecosystem services for different land use scenarios in the Yili river valley, china. Sustainability 13(3):1577
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks involved in drought stress response and tolerance. J Exp Bot 58(2):221–227
Siebert S, Döll P (2010) Quantifying blue and green virtual water contents in global crop production as well as potential production losses without irrigation. J Hydrol 384(3–4):198–217
Sivakumar M, Das H, Brunini O (2005) Impacts of present and future climate variability and change on agriculture and forestry in the arid and semi-arid tropics. Increasing climate variability and change. Springer, Netherlands, pp 31–72
Sloat LL, Gerber JS, Samberg LH et al (2018) Increasing importance of precipitation variability on global livestock grazing lands. Nat Clim Chang 8(3):214–218
Smith MS, McAllister RR (2008) Managing arid zone natural resources in Australia for spatial and temporal variability–an approach from first principles. Rangeland J 30(1):15–27
Smith D, Morton SR (1990) A framework for the ecology of arid Australia. J Arid Environ 18(3):255–278
Smith WK, Dannenberg MP, Yan D et al (2019) Remote sensing of dryland ecosystem structure and function: progress, challenges, and opportunities. Remote Sens Environ 233:111401
Spinoni J, Barbosa P, Cherlet M et al (2021) How will the progressive global increase of arid areas affect population and land-use in the 21st century? Global Planet Change 205:103597. https://doi.org/10.1016/j.gloplacha.2021.103597
Steele JE, Sundsoy PR, Pezzulo C et al (2017) Mapping poverty using mobile phone and satellite data. J R Soc Interface 14(127):20160690. https://doi.org/10.1098/rsif.2016.0690
Stehfest E, Van Zeist WJ, Valin H et al (2019) Key determinants of global land-use projections. Nat Commun 10(1):2166
Sterling SM, Ducharne A, Polcher J (2013) The impact of global land-cover change on the terrestrial water cycle. Nat Clim Chang 3:385–390. https://doi.org/10.1038/NCLIMATE1690
Stringer LC, Mirzabaev A, Benjaminsen TA et al (2021) Climate change impacts on water security in global drylands. One Earth 4(6):851–864. https://doi.org/10.1016/j.oneear.2021.05.010
Sun X, Yue T, Fan Z (2012) Scenarios of changes in the spatial pattern of land use in china. Procedia Environ Sci 13:590–597
Sun Y, Sun Y, Yao S et al (2021) Impact of climate change on plant species richness across drylands in China: from past to present and into the future. Ecol Ind 132:108288. https://doi.org/10.1016/j.ecolind.2021.108288
Swenson TL, Karaoz U, Swenson JM et al (2018) Linking soil biology and chemistry in biological soil crust using isolate exometabolomics. Nat Commun 9(1):19
Tan S, Wu B, Yan N et al (2018) Satellite-based water consumption dynamics monitoring in an extremely arid area. Remote Sensing 10(9):1399
Tarnita CE, Bonachela JA, Sheffer E et al (2017) A theoretical foundation for multi-scale regular vegetation patterns. Nature 541(7637):398–401
Teuling AJ, De Badts EA, Jansen FA et al (2019) Climate change, reforestation/afforestation, and urbanization impacts on evapotranspiration and streamflow in europe. Hydrol Earth Syst Sci 23(9):3631–3652
The World Bank (2012) Design of water consumption based water rights administration system for Turpan Prefecture of Xinjiang China. Water partnership program, Washington DC. http://documents.worldbank.org/curated/en/588081468216268772/Design-of-water-consumption-based-water-rights-administration-system-for-Turpan-prefecture-of-Xinjiang-China
Thuiller W, Lavorel S, Midgley G et al (2004) Relating plant traits and species distributions along bioclimatic gradients for 88 leucadendron taxa. Ecology 85(6):1688–1699
Tian G, Qiao Z (2014) Assessing the impact of the urbanization process on net primary productivity in China in 1989–2000. Environ Pollut 184:320–326
Trenberth KE, Smith L, Qian T et al (2007) Estimates of the global water budget and its annual cycle using observational and model data. J Hydrometeorol 8(4):758–769
Tueller PT (1987) Remote sensing science applications in arid environments. Remote Sens Environ 23(2):143–154
Uclés O, Villagarcía L, Cantón Y et al (2013) Microlysimeter station for long term non-rainfall water input and evaporation studies. Agric for Meteorol 182:13–20
Ustin SL, Valko PG, Kefauver SC et al (2009) Remote sensing of biological soil crust under simulated climate change manipulations in the mojave desert. Remote Sens Environ 113(2):317–328. https://doi.org/10.1016/j.rse.2008.09.013
van de Koppel J, Rietkerk M, van Langevelde F et al (2002) Spatial heterogeneity and irreversible vegetation change in semiarid grazing systems. Am Nat 159(2):209–218
van der Molen MK, Dolman AJ, Ciais P et al (2011) Drought and ecosystem carbon cycling. Agric for Meteorol 151(7):765–773
Verrelst J, Schaepman ME, Koetz B et al (2008) Angular sensitivity analysis of vegetation indices derived from chris/proba data. Remote Sens Environ 112(5):2341–2353. https://doi.org/10.1016/j.rse.2007.11.001
Verstraete MM, Scholes RJ, Smith MS (2009) Climate and desertification: looking at an old problem through new lenses. Front Ecol Environ 7(8):421–428. https://doi.org/10.1890/080119
von Hardenberg J, Meron E, Shachak M et al (2001) Diversity of vegetation patterns and desertification. Phys Rev Lett 87(19):198101
Wang L, D’Odorico P (2008) The limits of water pumps. Science 321(5885):36–37. https://doi.org/10.1126/science.321.5885.36c
Wang H, Sun F, Xia J et al (2017a) Impact of lucc on streamflow based on the swat model over the wei river basin on the loess plateau in China. Hydrol Earth Syst Sci 21(4):1929–1945. https://doi.org/10.5194/hess-21-1929-2017
Wang H, Zhang G-h, Liu F et al (2017b) Temporal variations in infiltration properties of biological crusts covered soils on the loess plateau of china. CATENA 159:115–125
Wang L, Kaseke KF, Seely MK (2017c) Effects of non-rainfall water inputs on ecosystem functions. Wires Water 4(1):e1179. https://doi.org/10.1002/wat2.1179
Wang S, Fu B, Chen H et al (2018) Regional development boundary of china’s loess plateau: water limit and land shortage. Land Use Policy 74:130–136
Wang Y, Zhang X, Guo Z (2021) Estimation of tree height and aboveground biomass of coniferous forests in north china using stereo zy-3, multispectral sentinel-2, and dem data. Ecol Ind 126:107645. https://doi.org/10.1016/j.ecolind.2021.107645
Weber B, Olehowski C, Knerr T et al (2008) A new approach for mapping of biological soil crusts in semidesert areas with hyperspectral imagery. Remote Sens Environ 112(5):2187–2201
Weber B, Wu D, Tamm A et al (2015) Biological soil crusts accelerate the nitrogen cycle through large no and hono emissions in drylands. Proc Natl Acad Sci 112(50):15384–15389
Wild TB, Khan Z, Clarke L, et al (2021) Integrated energy-water-land nexus planning in the colorado river basin (argentina). Reg Environ Change 21(3). https://doi.org/10.1007/s10113-021-01775-1
Williams L, Loewen-Schneider K, Maier S, et al (2016) Cyanobacterial diversity of western european biological soil crusts along a latitudinal gradient. FEMS Microbiol Ecol 92(10). https://doi.org/10.1093/femsec/fiw157
Woodhouse CA, Pederson GT (2018) Investigating runoff efficiency in upper colorado river streamflow over past centuries. Water Resour Res 54(1):286–300
Wu X, Li S (2014) Automatic sea fog detection over Chinese adjacent oceans using terra/Modis data. Int J Remote Sens 35(21):7430–7457
Wu B, Qian J, Zeng Y (2017) Land cover atlas of the people’s republic of China (1: 1 000 000). China Map Publishing House, Beijing) (in Chinese)
Wu B, Zeng H, Yan N et al (2018) Approach for estimating available consumable water for human activities in a river basin. Water Resour Manag 32(7):2353–2368
Wu B, Zhu W, Yan N et al (2020) Regional actual evapotranspiration estimation with land and meteorological variables derived from multi-source satellite data. Remote Sensing 12(2):332
Wu B, Zeng H, Zhu W et al (2021a) Enhancing China’s three red lines strategy with water consumption limitations. Sci Bull 66(20):2057–2060. https://doi.org/10.1016/j.scib.2021.06.012
Wu BF, Zeng HW, Lu N et al (2021b) Essential dryland ecosystem variables. Curr Opin Environ Sustain 48:68–76. https://doi.org/10.1016/j.cosust.2020.10.007
Wu B, Jiang L, Yan N, et al (2014) Basin-wide evapotranspiration management: concept and practical application in Hai basin, China. Agric Water Manag 145. https://doi.org/10.1016/j.agwat.2013.09.021
Xu C, Holmgren M, Van Nes EH et al (2015) Can we infer plant facilitation from remote sensing? A test across global drylands. Ecol Appl 25(6):1456–1462. https://doi.org/10.1890/14-2358.1
Xu G, Kang M, Metzger M, et al (2014) Vulnerability of the human-environment system in arid regions: the case of xilingol grassland in northern china. Polish J Environ Stud 23(5):1773–1785
Yan Y, Zhang C, Hu YF et al (2016) Urban land-cover change and its impact on the ecosystem carbon storage in a dryland city. Remote Sensing 8(1):6. https://doi.org/10.3390/rs8010006
Yang W, Long D, Bai P (2019a) Impacts of future land cover and climate changes on runoff in the mostly afforested river basin in north China. J Hydrol 570:201–219
Yang X, Xu M, Zhao Y et al (2019b) Moss-dominated biological soil crusts improve stability of soil organic carbon on the loess plateau, china. Plant Soil Environ 65(2):104–109
Ye Z, Chen Y, Li W (2010) Ecological water demand of natural vegetation in the lower Tarim river. J Geog Sci 20(2):261–272
Yin J, He F, Xiong YJ et al (2017) Effects of land use/land cover and climate changes on surface runoff in a semi-humid and semi-arid transition zone in northwest china. Hydrol Earth Syst Sci 21(1):183–196
Yokoyama G, Yasutake D, Wang W et al (2021) Limiting factor of dew formation changes seasonally in a semiarid crop field of northwest china. Agric for Meteorol 311:108705. https://doi.org/10.1016/j.agrformet.2021.108705
Yosef G, Walko R, Avisar R et al (2018) Large-scale semi-arid afforestation can enhance precipitation and carbon sequestration potential. Sci Rep 8(1):1–10
Yu J, Wang Z, Majumdar A et al (2018) Deepsolar: a machine learning framework to efficiently construct a solar deployment database in the United States. Joule 2(12):2605–2617. https://doi.org/10.1016/j.joule.2018.11.021
Yuan W, Zheng Y, Piao S, et al (2019) Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci Adv 5(8):eaax1396
Zastrow M (2019) China’s tree-planting drive could falter in a warming world. Nature 573(7775):474
Zeng H, Wu B, Zhu W et al (2019) A trade-off method between environment restoration and human water consumption: a case study in Ebinur lake. J Clean Prod 217:732–741
Zeng N, Yoon J (2009) Expansion of the world’s deserts due to vegetation‐albedo feedback under global warming. Geophys Res Lett 36(17):L17401
Zhai P, Larsen P, Millstein D et al (2012) The potential for avoided emissions from photovoltaic electricity in the United States. Energy 47(1):443–450
Zhang Q, Wang S, Yang F-L et al (2015) Characteristics of dew formation and distribution, and its contribution to the surface water budget in a semi-arid region in china. Bound-Layer Meteorol 154(2):317–331. https://doi.org/10.1007/s10546-014-9971-x
Zhang S, Yang D, Yang Y et al (2018) Excessive afforestation and soil drying on china’s loess plateau. J Geophys Res Biogeosci 123(3):923–935
Zhang F, Biederman JA, Dannenberg MP, et al (2021) Five decades of observed daily precipitation reveal longer and more variable drought events across much of the western united states. Geophys Res Lett 48(7):e2020GL092293
Zhao LX, Xu C, Ge ZM et al (2019) The shaping role of self-organization: linking vegetation patterning, plant traits and ecosystem functioning. Proc R Soc B 286(1900):20182859. https://doi.org/10.1098/rspb.2018.2859
Zhao Y, Xu W, Wang N (2021) Effects of covering sand with different soil substrates on the formation and development of artificial biocrusts in a natural desert environment. Soil Tillage Res 213:105081. https://doi.org/10.1016/j.still.2021.105081
Zhou X, An X, De Philippis R et al (2019) The facilitative effects of shrub on induced biological soil crust development and soil properties. Appl Soil Ecol 137:129–138
Zhou H, Gao Y, Jia X et al (2020) Network analysis reveals the strengthening of microbial interaction in biological soil crust development in the mu us sandy land, northwestern China. Soil Biol Biochem 144:107782
Zhu Z, Lu L, Yao X, et al (2019) Southeast asia energy outlook 2019. International Energy Agency
Zhuang Y, Zhao W (2017) Dew formation and its variation in haloxylon ammodendron plantations at the edge of a desert oasis, northwestern China. Agric for Meteorol 247:541–550
Zipper SC, Jaramillo F, Wang‐Erlandsson L, et al (2020) Integrating the water planetary boundary with water management from local to global scales. Earth’s Fut 8(2):e2019EF001377
Acknowledgements
This research was financially supported by the International Partnership Program of Chinese Academy of Sciences (121311KYSB20170004-01/03) and National Natural Science Foundation of China (41991232). The authors thank Prof Bojie Fu for his comments and suggestions, and Prof. Fernando T. Maestre for his contribution to the relevant part of the Global-DEP science plan.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2024 The Author(s)
About this chapter

Cite this chapter
Wu, B., Smith, W.K., Zeng, H. (2024). Dryland Dynamics and Driving Forces. In: Fu, B., Stafford-Smith, M. (eds) Dryland Social-Ecological Systems in Changing Environments. Springer, Singapore. https://doi.org/10.1007/978-981-99-9375-8_2
Download citation
DOI: https://doi.org/10.1007/978-981-99-9375-8_2
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-99-9374-1
Online ISBN: 978-981-99-9375-8
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)