
Abstract
Tropical peatlands have increasingly been targeted for logging or conversion to plantations in recent years. Tropical peatlands are unique ecosystems rich in biodiversity, but they have not attracted as much researcher attention as tropical forests, for example. There is still limited understanding of the ecological significance of peatland disturbance, or of the ecological resilience of peatland ecosystems. This study focuses on birds as indicator species in peatland ecologies. It compares bird communities in peatlands and non-peat lowlands in terms of: (1) species richness; (2) feeding guilds; and (3) responses to disturbance. Our research team analyzed bird communities in peatlands under several different land uses in Riau in comparison to those living in non-peat lowlands in Sumatra Island. We found that species richness in natural forests was lower in peatlands than in non-peat lowlands. The Jackknife estimator of species richness was 77.2 in natural forests on peatland, whereas on non-peat lowland, it was 114.8 and 241. Compared to non-peat lowland forests, the number of terrestrial insectivore and woodpeckers was lower in peatlands. Non-metric multidimensional scaling (NMDS) analysis showed that the bird community composition in peatland forests is unique compared to non-peat lowland forests, as they showed completely different lines of avifauna. Nevertheless, avifauna in disturbed sites on peatland were close to avifauna in disturbed non-peat lowland sites, which indicates that the disturbance of peatland would lead to homogenization of avifauna and loss of uniqueness, which in turn, leads to loss of biodiversity. Bird community composition in peatlands was very sensitive to land use change. Shifts in the community composition along the disturbance, as measured by Euclidean distances in the NMDS plot between each disturbed habitat and natural forest, were greater in peatland than in non-peat lowland. Although our knowledge and data of peatland ecologies are limited, it seems likely that certain peatland avifauna can only survive in natural peat swamp forest.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Birds are known to contribute to numerous ecosystem services, including pollination, pest control, and seed dispersal, which benefit both human welfare and economy at local and global scales (Millennium Ecosystem Assessment 2005; Şekercioğlu et al. 2016). Birds therefore serve as good indicators of overall ecosystem status (O’Connell et al. 2000). Sumatra Island has the largest and richest bird diversity of all the Indonesian Islands, with 580 known bird species, 21 of which are endemic species, and 120 migrant species (Whitten et al. 2000). Representative vegetation in the island consists mainly of lowland mixed dipterocarp forests, tropical montane forests, mangroves, and peat swamp forests. The original area of Sumatran peatland is estimated as 7.3–9.7 million ha, accounting for one-quarter of the world’s tropical peat swamp forests (Whitten et al. 2000), creating an important refuge for the birds of Sumatra (Yule 2010).
Lowland dipterocarp forests on the island have decreased dramatically in the last several decades due to development (Whitten et al. 2000; Laumonier et al. 2010), while a large area of natural forest remained in montane regions or peatlands. More recently peatland has become the target of commercial logging and conversion to plantations, especially for production of oil palm and acacia (Uryu et al. 2008; Corlett 2009; Koh et al. 2011). Peatland forests have attracted several basic studies on tree diversity (Anderson 1961; Bruenig 1990; Posa et al. 2011) and animal diversity (Whitemore 1984; Gaither Jr 1994; Philips 1998; Cheyne et al. 2010; Cheyne and Macdonald 2011; Posa 2011), but the impact of land use change on peatland ecosystems has been largely overlooked.
Posa (2011) reported that avian species diversity in selectively logged peatland forests in Central Kalimantan was quite similar to that found in intact forests. Avian diversity decreases by half in highly logged forest or recently burned brushland, however. Forest conversion, which usually replaces native trees with commercial trees or crops, generally has an even more significant effect on biodiversity (Barlow et al. 2007). For example, in Riau, Sumatra, Fujita et al. (2016) reported that the conversion of peat swamp forest into planted acacia forest and planted rubber forest had significant effects on bird diversity.
How resilient are peatland forests to forest degradation and conversion? What is their capacity recover desirable ecosystem states along with associated ecosystem services? Biodiversity is said to enhance ecosystem resilience (Elmqvist et al. 2003). In comparison to non-peat lowland forest, peat swamp forest supports low animal density and diversity (Janzen 1974; Whitten et al. 2000; Posa et al. 2011) due to its low nutrient content and low plant productivity (Bruenig and Droste 1995). Gaither Jr (1994) reported that although bird populations in peat swamp forest had low species diversity and density overall, some species have a higher density than in lowland dipterocarp forests. The uniqueness of the peatland ecosystem is attributed to a swampy ecosystem that is formed on peat soil, which contains high levels of organic carbon from undecomposed deadwood. Peatland soils are usually waterlogged and therefore have high acidity (pH 3–4.5) and low nutrient content.
Ecosystem or community resilience is evaluated by comparing pre-and post-disturbance variables and compositions (Norden et al. 2009; O’Dea and Whittaker 2007; Moretti et al. 2006). Our project used similar methods to compare the characteristics of bird communities on peatland and non-peat lowland by focusing on the following: (1) species richness; (2) feeding guilds; and (3) response of avifauna to disturbance. We analyzed how bird populations responded to rapidly changing patterns of peatland land use. We compared data of bird populations inhabiting several different peatland landscapes in Riau, with those inhabiting non-peat lowland landscapes in Sumatra Island. We then discussed the vulnerability and potential resilience of the peat swamp ecosystem as indicated by avian population response to disturbance.
2 Methods
2.1 Datasets in Peatland
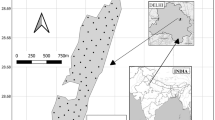
We used bird population datasets published by Fujita et al. (2016). The study site was located in the Bukit Batu area of the Giam Siak Kecil-Bukit Batu Biosphere Reserve, Riau Province, Indonesia (Fig. 3.1). Observations took place in the following sites: (1) natural peat swamp forest in the Bukit Batu Wildlife Reserve (hereafter referred to as BNF), which is managed by the Forestry Department of Riau; (2) plantation forest of Acacia crassicarpa, which is managed by the companies PT. Bukit Batu Hutani Alam and PT. Sakato Pratama Makmur (referred to as BAF);Footnote 1 (3) planted rubber (Hevea brasiliensis) forests, which are managed by local people (referred to as BRF); and (4) the residential village area in the Bukit Batu area (referred to as BRV; Fig. 3.2). BNF is dominated by Palaquium sumatranum, Eugenia paludosa, Diospyros hermaphroditica, Calophyllum lowii, and Shorea teysmanniana, with a peat depth of up to 6 m (Gunawan et al. 2012). According to Fujita et al. (2016), a bird survey was conducted in selectively logged forests which maintain a complex and multistory forest structure. Planted acacia forests are pure plantations of Acacia crassicarpa that are established on peatland where canals were dug in grids for drainage and transport purposes. Acacia trees are harvested within 5–6 years. The Acacia forest monocrop structure is simple—Acacia crassicarpa comprises the canopy layer, with only some ferns on the forest floor. Planted rubber forests, meanwhile, often include dense understories including shrubs, small trees, and saplings of native forest species. Although planted rubber forests range from highly-managed plantations with less understory to extensive forests with more understory trees, they are usually more diversified than industrial plantations. Land use in the residential village area consists of home gardens, rubber plantations, paddy fields, oil palm plantations, secondary forests (mainly Macaranga trees), coconut plantations, and burnt brush land.

Study sites. Abbreviations of study sites on the peatland of Fujita et al. (2016) are as follows: BNF natural peat swamp forest, BAF planted acacia forest, BRF planted rubber forest, BRV residential village. Study sites on non-peat lowland of Thiollay (1995) are TNF natural forest, TRF planted rubber forest, of Fujita et al. (2014) are FNF natural forest; FAF planted acacia forest, of Danielsen and Heegaard (1995) are DNF natural forest, DRF planted rubber forest, DOP oil palm plantation

Typical land use in peatland in Bukit Batu, Riau: (a) planted acacia forest (BAF); (b) residential village area (BRV); and (c) planted rubber forest (BRF) (Photos taken by Fujita)
Bird surveys were conducted using fixed-radius point count methods in twelve 1 km lines. Three surveys were conducted in each of the four land use categories. In each survey, four points were established at 250 m intervals, and every bird species that was seen or heard within a 25 m radius was recorded during the 20-min observation period. Observations were conducted in the morning and evening in March, May, and October 2011. In total, 34 censuses were made in the natural peat swamp forest (BNF), and 48 censuses were made in other sites (BAF, BRF, and BRV).
2.2 Datasets in Non-peat Lowland
We selected three papers that surveyed the bird community in non-peat lowland in Sumatra (Table 3.1). The criteria for the paper selection were that the datasets must be from direct observations of birds using either the line-transect or point-count method, both of which are useful in analysis of bird community composition. Thiollay (1995) investigated bird fauna in lowland dipterocarp forests (hereafter referred to as TNF) and planted rubber forests (hereafter referred to as TRF) in the Jambi and West Sumatra provinces in May–June 1991 and July 1992, respectively (Fig. 3.1). The emergent trees (44–55 m) in TNF were mostly Dipterocarpus sp., Shorea sp., and Cesalpiniaceae, with canopy trees (35–45 m), including numerous Dipterocarpaceae, Myristicaceae, Sapotaceae, and fig trees (Ficus sp.). TRF consists mainly of para-rubber trees, Hevea brasiliensis, which are lower (closed canopy at 20–30 m) and have dense undergrowth. An average stand includes 750 trees per hectare, 65% of which are rubber trees mixed with 12–20 other species of forest and fruit trees. A line-transect survey was performed, where a sample was defined as the first 50 individual birds that were recorded within 20–30 m on either side of the transect. Twenty eight-samples were collected in both TNF and TRF. Since the accessible tracts of the primary forest were limited, all the data from the three different areas were pooled into a single 28-sample set that was assumed to be representative of the western Sumatra natural forest. Counts were made from dawn to 11:00 h and from 15:00 h to sunset outside rainy periods.
Danielsen and Heegaard (1995) investigated bird fauna in both primary and logged lowland dipterocarp forests (hereafter referred to as DNF), planted rubber forests (hereafter referred to as DRF), and oil palm plantations (hereafter referred to as DOP) in Bukit Tigapuluh, Riau, and Jambi province in July–September 1991 (Fig. 3.1). The most prevalent tree families in DNF were Euphorbiaceae, Lauraceae, Burseraceae, and Dipteroearpaceae, which comprise a multistory forest structure, with a 26–38 m canopy layer, 18–26 m middle layer, and 10–21 m lower layer. The height of the canopy in DRF was 15–20 m, with emergents reaching 25 m, and 36% of the trees over 10 cm diameter reaching breast height (DBH) were estimated to be para-rubber tree (Hevea brasiliensis). Undergrowth consisted of 1–2 m tall perennials and thickets of bamboo Dendrocalamus sp.
The DOP plantation was 10,000 ha and consisted of mature oil palms, Elaeis guineensis. The average height of the palms was 11 m, and the density was 162 trees per ha. Within 1.5 m of the palms, an area was covered by dead palm leaves and ferns or no vegetation. The authors used a variable-distance line-transect method supplemented by systematic mist-netting. A line-transect survey route of 2 km was cut in a straight line in each habitat every 250 m. The route was walked by one person with a speed of 250 m per 15 min, including brief stopovers. The route was surveyed 4 times per day. In DNF and DRF, the route was surveyed for 40 h, while in DOP, the route was only surveyed for 20 h. Mist netting was carried out with 15–20 nets, which were mainly set at a height of 0.5–3.0 m for three to four consecutive days. The effort was 10,200–15,500 net-meter hours in DNF and DRF and only 3300 net-meter hours in DOP.
The third paper used to establish the present dataset, by Fujita et al. (2014), examined bird fauna in a lowland Acacia mangium plantation (FAF) and remnant logged natural forest (FNF). The height of the acacia canopy in FAF for the 4-year-old planted acacia forest was 10–20 m with less understory (1–10 m) vegetation, and the forest floor was occasionally covered by ferns or sparse scrub. The forest floor of the 1-year-old planted acacia forest had no vegetation other than 3–5 m of planted A. mangium. Most of the points in FNF consisted of trees that were 20–30 m tall and scattered with multistory vegetation. The other two sites of conserved logged forests (CF-07, CF-08) were burnt during a forest fire, and no living tall trees could be seen; instead, all sites were covered by dense bush.
Fixed-radius point counts were conducted in October–December 2007 and July–August 2008. Twenty survey points for FAF in the 1- and 4-year-old planted acacia forests and 12 survey points for FNF were selected. At each point, the bird species that were observed within a 25 m radius were recorded in the 10-min census. The number of censuses totaled 303 in FAF and 197 in FNF.
2.3 Feeding Guild
To compare and evaluate bird communities and forest ecosystem in terms of ecological function, feeding guilds of birds were used in the following analysis. A feeding guild is defined as the groups of species that have similar feeding habits. The following 17 feeding guild categories were used for the analysis: A-F, arboreal frugivore; AFG-I, arboreal foliage gleaning insectivore; AFG-IF, arboreal foliage gleaning insectivore–frugivore; A-FP, arboreal frugivore–predator; A-I, aerial insectivore; BG-I, bark gleaning insectivore; M-IP, miscellaneous insectivore–piscivore; NF, nectarivore-frugivore; NI, nectarivore-insectivore; NIF, nectarivore-insectivore-frugivore; R, raptor; S-I, sallying insectivore; SSG-I, sallying substrate gleaning insectivore; T-F, terrestrial frugivore; T-G, terrestrial granivore; T-I, terrestrial insectivore; and T-IF, terrestrial insectivore–frugivore. Each species was categorized in one of the feeding guilds, with reference to Lambert (1992), Styring et al. (2011), and Smythies (1999). Species previously unreferred to a feeding guild were categorized according to the feeding guild of the nearest species in the same genus.
2.4 Statistical Analysis
To compare the species richness at each site, all datasets were merged, and abundance data was converted to presence-absence data. Water birds, night birds, and swifts (excluding tree swifts) were omitted from the list. Scientific names followed the nomenclature of Sibley and Monroe (1990). Since observation efforts differ among sites, a direct comparison of species richness or number must be treated with caution. Therefore, we used the nonparametric Jackknife estimator of species richness to compare bird diversity among the sites. Jackknife 1 was calculated for the data of Bukit Batu and Fujita et al. (2014) using the software EstimateS ver 9.1 (Colwell 2013). Jackknife 1 was used since it has been widely used for years, including by Thiollay (1995). In this manner it was possible to compare data from at least three sites, with the exception of the dataset of Danielsen and Heegaard (1995).
To compare the species composition among the study sites, the ordination of sites and habitat types was performed against the presence-absence data. This was achieved using nonmetric multidimensional scaling (NMDS) with the package “vegan” (Oksanen et al. 2015) in the statistical software R version 3.2.2 (R Core Team 2015). NMDS was performed with Bray–Curtis as a dissimilarity index, and a maximum 100 times of iteration was conducted to obtain better ordination scores. Additionally, the species score against each axis was calculated, although only abundant species appeared with a label when several species appeared in the small area.
3 Results and Discussion
3.1 Bird Species Diversity on Peatland and Non-peat Lowland
The overall species number in the forests on peatland was lower than that of forests on non-peat lowland. The total number of species in peatland in the Bukit Batu area was 44 in the natural peat swamp forest (BNF), 20 in the planted acacia forest (BAF), 38 in the planted rubber forest (BRF), and 51 in the residential village area (BRV). In the forests in the non-peat lowland of Thiollay (1995), the number of species was 180 in natural forest (TNF) and 104 in the planted rubber forest (TRF). In the study by Danielsen and Heegaard (1995), there were 147 in the natural forest (DNF), 90 in the planted rubber forest (DRF), and 24 in the oil palm plantation (DOP). In the study by Fujita et al. (2014), there were 82 in the natural forest (FNF) and 63 in the planted acacia forest (FAF).
Out of 283 bird species in total, 101 species were recorded in forests on peatland in Bukit Batu, whereas in forests on non-peat lowland, 267 species were reported, and 85 species were seen in both peatland and non-peat lowland. Out of the 267 species that were seen in the forests on non-peat lowland, 182 species were not recorded in the forests on peatland. However, out of the 101 species on peatland, 16 species were seen only on peatland, of which four species (Prionochilus thoracicus, Ficedula zanthopygia, Malacocincla abbotti, and Nectarinia calcosthetha) were specifically seen only in natural forests on peatland (BNF). Of these, P. thoracicus and N. calcosthetha favor swamps or coastal forests as their habitat. Additionally, we observed several species that prefer swamps, including Malacopteron albogulare and Setornis criniger (Fujita et al. 2012), although they are not listed in the point-count survey by Fujita et al. (2016). Gaither Jr (1994) reported that several bird species, including M. albogulare, were found in higher density in peat swamp forests compared to non-peat lowland forests. Although the species richness is low compared to forests on non-peat lowland, forests on peatland could be home to several bird species that favor a swampy environment or species that avoid the highly competitive pressure in forests on non-peat lowland.
Jaccard’s coefficient of community (CC) index, which represents the community similarity between two sites, indicates that the peatland bird community has a low similarity (0.167–0.248) with other non-peat lowland bird communities, whereas the non-peat lowland bird community has a higher similarity (0.242–0.493) among sites (Table 3.2).
Since the efforts and methods of observation differ among sites, it is not possible to compare species richness directly. The Jackknife estimator of species richness, the Jack 1 mean, indicates the true species richness of natural forest in Bukit Batu (BNF) as 77.2, which is 33 species greater than the observed species richness (Table 3.3). Compared to natural forests on non-peat lowland, of which the estimated species richness is 241 species in TNF and 114.8 in FNF, natural forests on peatland have a lower species richness.
These measurements indicate poor species diversity in peat swamp forests, which follows the hypothesis that peat swamp forests support depauperate animal communities due to the cascading influence of poor soils (Janzen 1974). Nevertheless, the peat swamp forest is a unique ecosystem supporting several bird species that are not present in other ecosystems, including forests on non-peat lowland.
These direct comparisons of landcover and bird populations must be considered with caution for several reasons. First, observation efforts differ significantly in the different studies—up to 10 times in observation hours (Table 3.3). The lower observation time in the natural forest on peatland is due to limited accessibility, time, and labor power, all of which may be linked to the lower number of species observed. Comparisons of estimators of species richness, such as Jackknife, have solved this problem to some extent. Second, the observation methods differed among sites and included line transect, point count, and mist netting. Even the line-transect method differed slightly among sites, as Thiollay (1995) counted 50 individuals from the beginning, with no regard to the length of the line, whereas Danielsen and Heegaard (1995) walked the 250 m line and counted all birds observed. Third, habitat complexity differed among sites, as Thiollay (1995) merged all data from several patches from different regions and treated it as a single dataset for the primary forest. The regions include hill forests, which resulted in the presence of montane species and increased the overall species richness. Moreover, regarding Bukit Batu, all study points of natural peat swamp forest (BNF) were selectively logged a few years before the survey, perhaps affecting the study results. In short, such discrepancies in datasets indicates that the hypothesis that peatlands support poor bird species diversity still requires further intensive research.
3.2 Feeding Guild Change in Disturbed Sites on Peatland
In the disturbed sites of peatland in Bukit Batu, namely planted acacia forest (BAF), planted rubber forest (BRF), and residential village (BRV), a decline in the sallying insectivores (S-I and SSG-I) was observed (Table 3.4). This decline was also observed in non-peat lowland forest. BRV was characterized by more granivores (T-G) and terrestrial doves (T-IF), which reflect the abundance of open spaces and grass seeds as their food (Fig. 3.2b). Hornbills (A-FP) appeared more frequently in the residential village than expected. This could be because they were more easily detectable in open spaces than in the forest, since their sound was heard more often in the forested sites than BNF (although not every species was recorded when their call was beyond our observation range of 25 m).
Compared to natural forests on non-peat lowland (TNF, DNF, and FNF), natural forest in Bukit Batu (BNF) was characterized by fewer terrestrial insectivores (T-I), such as Trichastoma sp., Enicurus sp., Pitta sp., and Napothera sp., and fewer woodpeckers (BG-I) (Table 3.4). Gaither Jr (1994) reported similar results in a mist-net survey of understory birds in peat swamp forests in West Kalimantan. He found litter-gleaning insectivores (T-I in our category), tree foliage-gleaning insectivores (AFG-I in our category), and bark-gleaning insectivores in significantly lower numbers in the peat swamp forest than in the dipterocarp forest. Posa (2011) had partly contradicting results in the peat swamp forest in Central Kalimantan, as six species (8%) of woodpeckers (BG-I) were recorded in peat swamp forests, although litter-gleaning insectivores (T-I in our category) only comprised three species (4%). The abundance of woodpeckers was associated with the number of dead trees in the degraded peat swamp forest due to recent drought stress (Posa 2011). The decline in TI and BGI in Bukit Batu might be the result of logging activity in the natural forest. As recently as 2009, illegal logging has occurred in most of the accessible areas, including our study plots.
The low number of arboreal frugivores (A-F) could be explained by the seasonal change. Gaither Jr (1994) reported that many frugivore species were observed in June in the peat swamp forest, but not in other months.
3.3 Shift in Bird Community Composition along the Disturbance
Many fewer species were found in disturbed sites than in the natural peatland forests in Bukit Batu. Out of the 44 species found in natural forests in Bukit Batu (BNF), only eight species in planted acacia forest (BAF), 12 species in the planted rubber forest (BRF), and nine species in residential village (BRV) were shared, which indicated that more than 70% of species were lost in the disturbed sites. However, in non-peat lowland, 88 species in the planted rubber forest (TRF) out of 180 species in the natural forest (TNF), 66 species in the planted rubber forest (DRF) out of 147 species in natural forest (DNF), and 42 species in the planted acacia forest (FAF) out of 82 species in natural forest (FNF) were found, which account for approximately 50% of the species loss in the disturbed sites. The oil palm plantation showed a drastic species loss of up to 94%, as only eight species were found in DOP, in comparison to 147 species in the natural forest (DNF).
The NMDS analysis showed that the bird community composition in forests on peatland is unique compared to forests on non-peat lowland (Fig. 3.3a). Points of non-peat lowland (TNF, TRF, DNF, DRF, DOP, FNF, and FAF) showed a higher NMDS2 value than points of peatland, except for BRV, which is BNF, BAF, and BRF. This indicates that peatland has different avifauna compared to the forests on non-peat lowland. BRV showed a similar NMDS2 value as that of non-peat lowland; this might have resulted from either the high modification to the village or the presence of non-peat soil in the villages, which are located on the coastline. The NMDS2 axis was negatively correlated with the presence of Nectarinia calcosthetha, which is restricted to peat swamp forests (Fig. 3.3b).

Bird community similarities using an NMDS ordination of study points (a) and species (b). (a) Each point corresponds to a study point. Abbreviations in the legends are as follows: BNF natural peat swamp forest in Bukit Batu (closed triangle), BAF planted acacia forest in Bukit Batu (closed circle), BRF planted rubber forest in Bukit Batu (closed square), BRV residential village in Bukit Batu (star), TNF natural forest of Thiollay 1995 (pale gray triangle), TRF planted rubber of Thiollay 1995 (pale gray square), FNF natural forest of Fujita et al. 2014 (open triangle), FAF planted acacia forest of Fujita et al. 2014 (open circle), DNF natural forest of Danielsen and Heegaard 1995 (dark gray triangle), DRF planted rubber forest of Danielsen and Heegaard 1995 (dark gray square), DOP oil palm plantation of Danielsen and Heegaard 1995 (cross in open circle). The arrows indicate the directions from the natural forest to the disturbed habitat at each site. The values near the arrows indicate the Euclidean distances calculated from the NMDS 1 and 2 values. (b) In the species plot, only abundant species are labeled where several species are clustered. Abbreviations for the species names are as follows: Amauphoe Amaurornis phoenicurus, Bucevigi Buceros vigil, Celebrac Celeus brachyurus, Copssaul Copsychus saularis, Corvmacr Corvus macrorhynchos, Cotuchin Cotumix chinensis, Dicrpara Dicrurus paradiseus, Macrgula Macronous gularis, Malamagn Malacopteron magnum, Muscdauu Muscicapa dauurica, Nectcalc Nectarinia calcostetha, Orthseri Orthotomus sericeus, Pandhali Pandion haliaetus, Phylbore Phylloscopus borealis, Pycnbrun Pycnonotus brunneus, Rollroul Rollulus rouloul, Spilchee Spilornis cheela, Strechin Streptopelia chinensis, Trercurv Treron curvirostra, Trervern Treron vernans, Tricbico Trichastoma bicolor, Turnsusc Turnix suscitator, Zootinte Zoothera interpres
While the NMDS2 axis divided forests on non-peat lowland and peatland, NMDS1 axes showed the degree of disturbance. On both peatland and non-peat lowland, the natural forest points (BNF, TNF, DNF, and FNF) showed lower NMDS1 values than disturbed points, such as planted acacia forest, planted rubber forest, oil palm plantation, and residential village (Fig. 3.3a, direction of disturbance shown in the arrows for each site). Species that had a positive correlation with the NMDS1 axis include Cotumix chinensis and Turnix suscitator, both of which are open space species, and species that had a negative correlation include Calyptomena viridis and Trichastoma bicolor, both of which are species inhabiting less disturbed natural forests (Fig. 3.3b).
Disturbed points on peatland in Bukit Batu (BAF, BRF) showed a higher NMDS1 value and NMDS2 value compared to natural forests (BNF). This indicates that when natural forests are converted to planted acacia and rubber forests bird communities change accordingly, shifting toward the avifauna of non-peat lowlands. For example, Nectarinia calcostetha, which is found in natural peatland forests, has not been recorded in other habitat types. Disturbance of peatland therefore will likely lead to the homogenization of avifauna and loss of uniqueness, which would then lead to a loss in overall biodiversity.
Peatland-based bird community composition responded sensitively to land use change. Euclidean distances of the NMDS values of community composition between natural forest and planted rubber forest in non-peat lowland were 0.52 in TNF-TRF, 0.47 in DNF-DRF, and 0.93 in BNF-BRF in peatland, indicating a 1.7–1.9-times longer distance in peatland (Fig. 3.3a). As distances on NMDS values indicate differences in community composition, a longer Euclidean distance indicates a larger difference in community composition. Euclidean distances between natural forest and planted acacia forest in non-peat lowland were 0.37 in FNF-FAF and 1.41 in BNF-BAF in peatland, which shows a 3.8-times longer distance in peatland (Fig. 3.3a). Although avifauna clearly responded more strongly to disturbance in peatland, the reason for this vulnerability is unclear. It may be attributed low biodiversity and the idiosyncratic characteristics of the peat swamp ecosystem. To verify the resilience in the peat swamp forest, repetitive data from long-term studies on community composition and analyses from the perspective of landscape ecology are necessary. For example, differences in landscape structure and scale in peatlands might have significant impacts on bird communities, since plantations are often developed on a large scale due to the flat topography and lower utilization by local people.
3.4 Conclusion: Peatland Ecosystem for Birds
Our survey and analysis lead us to conclude the following: (1) although the peatland ecosystem has a comparably poorer species diversity than non-peat lowland forest, peatlands contains unique and idiosyncratic avifauna; (2) peat swamp forest avifauna lack terrestrial insectivore and woodpeckers; and (3) avifauna in peatland are sensitive to forest conversion to planted tree forest, and can experience a loss of more than 70% of species. In other words, unique and idiosyncratic avifauna on peatland are only maintained by natural peat swamp forest. Our first two findings support those of Janzen (1974), Gaither Jr (1994), and Posa (2011). The third finding, which has the most stimulating implication, suggests that the peat swamp ecosystem has a higher vulnerability to disturbance compared to non-peat lowland. It is still in the stage of hypothesis. Peat swamp vulnerability may be due to poor biodiversity and the idiosyncratic characteristic of the peat swamp ecosystem. Long-term studies on bird community and landscape ecology are required to evaluate this hypothesis. Nevertheless, our study found several important implications for peatland management, considering that there are limited studies on the effect of peatland disturbance on biodiversity.
Samejima et al. (2016) listed several other bird species detected by camera trap. Lophura erythrophthalma and Melanoperdix niger were both vulnerable species, and they were both restricted to natural peat swamp forest and not observed in the planted acacia forest. This indicates that disturbance has negative effects on these forest-dependent species, and the implication is consistent with our results. To conserve the unique peat swamp avifauna and maintain the ecosystem services that are provided by biodiversity, peat swamp forest should not be converted to plantations and agricultural lands, as peatland ecosystems have low resilience to disturbance.
Notes
- 1.
“PT.” is the abbreviation for Perseroan Terbatas, and it means Limited Liability Company.
References
Anderson J (1961) The ecology and forest type of the peat swamp forests of Sarawak and Brunei in relation to their silviculture. Forest Department Sarawak, Kuching
Barlow J, Gardner TA, Araujo IS et al (2007) Quantifying the biodiversity value of tropical primary, secondary, and plantation forests. PNAS 104:18555–18560. https://doi.org/10.1073/pnas.0703333104
Bruenig E (1990) Oligotrophic forested wetlands in Borneo. In: Lugo AE, Brinson M, Brown S et al (eds) Ecosystems of the world, Forested wetlands, vol 15. Elsevier, Amsterdam, pp 299–334
Bruenig E, Droste H (1995) Structure, dynamics and management of rainforests on nutrient-deficient soils in Sarawak. In: Primack RB, Lovejoy TE (eds) Ecology, conservation and management of Southeast Asian rainforests. Yale University Press, New Haven, pp 41–53
Cheyne SM, Macdonald DW (2011) Wild felid diversity and activity patterns in Sabangau peat-swamp forest, Indonesian Borneo. Oryx 45:119–124
Cheyne SM, Husson SJ, Chadwick RJ et al (2010) Diversity and activity of small carnivores of the Sabangau Peat-swamp Forest, Indonesian Borneo. Small Carniv Conser 43:1–7
Colwell RK (2013) EstimateS: statistical estimation of species richness and shared species from samples. Version 9. User’s guide and application. http://purl.oclc.org/estimates. Accessed 8 Aug 2016
Corlett RT (2009) The ecology of tropical East Asia. Oxford University Press, Oxford
Danielsen F, Heegaard M (1995) The birds of Bukit Tigapuluh, Southern Riau, Sumatra. KUKILA 7:99–120
Elmqvist T, Folke C, Nystrom M et al (2003) Response diversity, ecosystem change, and resilience. Front Ecol Environ 1:488–494. https://doi.org/10.1890/1540-9295(2003)001[0488:RDECAR]2.0.CO;2
Fujita MS, Irham M, Fitriana YS et al (2012) Mammals and birds in Bukit Batu area of Giam Siak Kecil–Bukit Batu Biosphere Reserve, Riau, Indonesia. Kyoto Working Papers on Area Studies No. 128 (G-COE Series 126), pp 1–70. Center for Southeast Asian Studies, Kyoto
Fujita MS, Prawiradilaga D, Yoshimura T (2014) Roles of fragmented and logged forests for bird communities in industrial Acacia mangium plantations in Indonesia. Ecol Res 29:741–755. https://doi.org/10.1007/s11284-014-1166-x
Fujita MS, Samejima H, Haryadi DS et al (2016) Low conservation value of converted habitat for avifauna in tropical peatland on Sumatra, Indonesia. Ecol Res 31:275–285. https://doi.org/10.1007/s11284-016-1334-2
Gaither JC Jr (1994) Understory avifauna of a Bornean peat swamp forest: is it depauperate? Wilson Bull 106:381–390
Gunawan H, Kobayashi S, Mizuno K et al (2012) Peat swamp forest types and their regeneration in Giam Siak Kecil-Bukit Batu Biosphere Reserve, Riau, East Sumatra, Indonesia. Mires Peat 10:1–17
Janzen DH (1974) Tropical blackwater rivers, animals, and mast fruiting by Dipterocarpaceae. Biotropica 6:69–103. https://doi.org/10.2307/2989823
Koh LP, Miettinen J, Liew SC et al (2011) Remotely sensed evidence of tropical peatland conversion to oil palm. PNAS 108:5127–5132. https://doi.org/10.1073/pnas.1018776108
Lambert FR (1992) The consequences of selective logging for Bornean lowland forest birds. Philos Trans R Soc B 335:443–457. https://doi.org/10.1098/rstb.1992.0036
Laumonier Y, Uryu Y, Stüwe M et al (2010) Eco-floristic sectors and deforestation threats in Sumatra: identifying new conservation area network priorities for ecosystem-based land use planning. Biodivers Conserv 19:1153–1174. https://doi.org/10.1007/s10531-010-9784-2
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: synthesis. Island Press, Washington, DC
Moretti M, Duelli P, Obrist MK (2006) Biodiversity and resilience of arthropod communities after fire disturbance in temperate forests. Oecologia 149:312–327. https://doi.org/10.1007/s00442-006-0450-z
Norden N, Chazdon RL, Chao A et al (2009) Resilience of tropical rain forests: tree community reassembly in secondary forests. Ecol Lett 12:385–394. https://doi.org/10.1111/j.1461-0248.2009.01292.x
O’Connell TJ, Jackson LE, Brooks RP (2000) Bird guilds as indicators of ecological condition in the Central Appalachians. Ecol Appl 10:1706–1721. https://doi.org/10.1890/1051-0761(2000)010[1706:BGAIOE]2.0.CO;2
O’Dea N, Whittaker RJ (2007) How resilient are Andean montane forest bird communities to habitat degradation? Biodivers Conserv 16:1131–1159. https://doi.org/10.1007/s10531-006-9095-9
Oksanen J, Blanchet FG, Kindt R et al (2015) Vegan: community ecology package. R package version 23-0 http://CRANR-projectorg/package=vegan. Accessed 24 Jun 2015
Philips VD (1998) Peatswamp ecology and sustainable development in Borneo. Biodivers Conserv 7:651–671. https://doi.org/10.1023/A:1008808519096
Posa MRC (2011) Peat swamp forest avifauna of Central Kalimantan, Indonesia: effects of habitat loss and degradation. Biol Conserv 144:2548–2556. https://doi.org/10.1016/j.biocon.2011.07.015
Posa MRC, Wijedasa LS, Corlett RT (2011) Biodiversity and conservation of tropical peat swamp forests. Bioscience 61:49–57. https://doi.org/10.1525/bio.2011.61.1.10
R Core Team (2015) R: a language and environment for statistical computing. R foundation for statistical computing, Vienna. https://www.R-project.org/. Accessed 10 July 2015
Samejima H, Fujita MS, Muhammad A (2016) Biodiversity in peat swamp forest and plantations. In: Mizuno K, Fujita MS, Kawai S (eds) Catastrophe and regeneration in Indonesia’s peatlands: ecology, economy and society, Kyoto CSEAS series on Asian studies, vol 15. NUS Press/Kyoto University Press, Singapore/Kyoto, pp 353–379
Şekercioğlu CH, Wenny DG, Whelan CJ (2016) Why birds matter: avian ecological function and ecosystem services. University of Chicago Press, Chicago and London
Sibley CG, Monroe BL (1990) Distribution and taxonomy of birds of the world. Yale University Press, New Haven
Smythies BE (1999) The birds of Borneo, 4th edn. Natural History Publications (Borneo), Kota Kinabalu
Styring AR, Ragai R, Unggang J et al (2011) Bird community assembly in Bornean industrial tree plantations: effects of forest age and structure. For Ecol Manag 261:531–544. https://doi.org/10.1016/j.foreco.2010.11.003
Thiollay JM (1995) The role of traditional agroforests in the conservation of rain forest bird diversity in Sumatra. Conserv Biol 9:335–353
Uryu Y, Mott C, Foead N et al (2008) Deforestation, forest degradation, biodiversity loss and CO2 emissions in Riau, Sumatra, Indonesia: one Indonesian province's forest and peat soil carbon loss over a quarter century and its plans for the future. WWF Indonesia technical report, WWF Indonesia, Jakarta
Whitemore TC (1984) Tropical rain forest of the Far East. Clarendon Press, Oxford
Whitten T, Damanik SJ, Anwar J et al (2000) The ecology of Sumatra. Periplus, Hong Kong
Yule CM (2010) Loss of biodiversity and ecosystem functioning in Indo-Malayan peat swamp forests. Biodivers Conserv 19:393–409
Acknowledgments
This study was partly supported by the JSPS Global COE Program “In Search of Sustainable Humanosphere in Asia.”
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2023 The Author(s)
About this chapter

Cite this chapter
Fujita, M.S., Samejima, H., Haryadi, D.S., Muhammad, A. (2023). Characteristics of Bird Community Response to Land Use Change in Tropical Peatland in Riau, Indonesia. In: Mizuno, K., Kozan, O., Gunawan, H. (eds) Vulnerability and Transformation of Indonesian Peatlands. Global Environmental Studies. Springer, Singapore. https://doi.org/10.1007/978-981-99-0906-3_3
Download citation
DOI: https://doi.org/10.1007/978-981-99-0906-3_3
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-99-0905-6
Online ISBN: 978-981-99-0906-3
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)