
Abstract
The growing demands for improved seeds of food, vegetables, flowers, other horticultural species, feed, fibre, forage, and fuel crops are driving the global seed industry at a CAGR of 6.6%, which is expected to grow from a market size of USD 63 billion in 2021 to USD 86.8 billion by 2026. This will primarily depend on two key features, genetic enhancement of the crop variety and seed quality. The focus of varietal improvement will be not only on yield increase but also on tolerance against biotic and abiotic stresses to meet the challenges of climate change, better input use efficiency, and improvement of the nutritional value. Advanced molecular tools and techniques including gene editing are likely to be used for precision breeding. The scope of seed quality, on the other hand, will grow beyond the basic parameters of purity, germination, and health, to the ability to perform better under adverse growing conditions, supplement the nutrient deficiency in the soil, and withstand pests during seed germination and early growth stages. Seed enhancement technologies will complement the genetic enhancement, as the advancements in seed technology will become an integral part of future crop improvement programmes. This will call for more efficient and precise technologies for determining quality parameters and predicting seed longevity, which will need to be developed using advanced marker technologies, 3D and multispectral imaging analysis, digital phenotyping, and other non-destructive methods. Novel approaches of hybrid seed production, including apomixis and doubled haploidy, will be integrated further for their commercial application. Multi-dimensional approaches need to be taken for addressing the gaps in understanding the key physiological processes underlying the regulation of dormancy (and pre-harvest sprouting), germination, recalcitrance, loss of viability, and seed priming.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- AI
- Apomixis
- Doubled haploidy
- Non-destructive testing for viability
- Multispectral imaging
- Regulation of dormancy
- Seed longevity
- Molecular markers for seed quality
- EDVs
1 Introduction
Seed technology, the vital link between plant breeding and crop husbandry, has turned out to be a multi-disciplinary subject, relying primarily on the scientific advancements in the fields of plant biology, genetics, plant breeding, plant physiology, plant pathology, entomology, agronomy, and agricultural engineering, as all of them would be involved in quality seed production, testing of its genetic purity, seed germination, vigour and health, and proper processing and storage to retain high quality till used by the farmers. There is, however, no defining line between the science and technology of seeds. While seed science refers to the research and advancements of scientific knowledge of the processes that determine the quality of seed, seed technology relates to commercial application of this knowledge for superior crop performance and also increased profitability for the seed industry. Identifying and understanding the problems, and finding scientific explanations for these, is the first step towards developing a successful technology. With the rapid advancements in modern agriculture, be it the genetic improvement through selection, hybridization, molecular breeding, gene editing, or genetic modifications; novel approaches to hybrid seed production; development of precise seed enhancement technologies to withstand abiotic and biotic stresses; or automation and AI, scientific research has led to the technological advancements as is evidenced by the enormous progress in this field. While significant scientific advancements have been made almost in every field of seed biology, production, and storage (also conservation), there are critical gaps in our understanding of several aspects of seed genetics, physiology, production, quality assessment, and quality improvement, which calls for more deep delving into these functions. More effective technologies can be built only on further advancements in the scientific bases of these functions. Recent years have seen some significant advances in addressing emerging problems in this sector, some of which are discussed here.
2 Hybrid Seed Production
The discovery of cytoplasmic male sterility (CMS) facilitated the hybrid breeding in several crop species, e.g. rice, sorghum, pearl millet, brassicas, sunflower, etc., enabling the production of high-yielding hybrids. Whereas heterosis breeding has been more effectively translated into hybrid development in cross-pollinated species, there is still plenty of scope to utilize heterosis in other economically important crop species, where pure line are still dominating, and the productivity has nearly reached a plateau. Identifying new sources of CMS and their restorer genes, broadening the genetic background by utilizing the crop wild relatives (CWR), and molecular breeding are being pursued most vigorously by plant geneticists, breeders, and molecular biologists globally. While these initiatives will undoubtedly lead to further increase in crop productivity, making genetically pure and high-quality hybrid seeds available to the smallholding farmers at an affordable cost remains a challenge in the absence of stable and functional CMS systems in many agriculturally important species. A stable expression of male sterility is of utmost importance not only for cost-effective hybrid seed production but also for the maintenance of the seed parents and breeder and foundation seed production. Effects of erratic environmental conditions on the gene flow and breakdown of male sterility need to be studied more comprehensively, specially in high-value crops, to understand the specific roles of different factors and accordingly plan hybrid seed production under suitable growing conditions.
The potential of genic (or genetic) male sterility (GMS) has found limited use in commercial hybrid seed production in field crops. In this system, the recessive/wild-type (WT) heterozygote is used to produce both the GMS line and the maintainer line. The need for roguing out 50% male fertile plants in seed parent population, and difficulty in differentiating the male fertile plants from the male sterile ones at an early stage makes this a tedious and economically non-viable except in some cucurbits and ornamental marigold. Despite the widespread application of PTGMS in rice, the systems are easily affected by unpredictable environmental conditions, causing intrinsic problems for sterile and maintainer line seed propagation and hybrid seed production (Chang et al. 2016). The barnase-barstar system of GMS is successful in hybrid development and seed production in Indian mustard (Mariani et al. 1992). Wu et al. (2016) proposed a next-generation hybrid seed production technology (SPT) by adopting a three-step approach comprising (1) a WT full coding sequence (CDS) of a male fertility gene for fertility restoration, (2) a pollen-inactivating gene that disrupts transgenic pollen development, and (3) a seed colour marker gene for seed sorting. Several similar strategies are being tested in rice (Chang et al. 2016) and maize (Zhang et al. 2018). Qi et al. (2020) proposed a simple next-generation sortable hybrid seed production strategy based on CRISPR/Cas-9 technology that creates a manipulated GMS maintainer (MGM) system via a single transformation. The derived single-copy hemizygous MGM lines bore a mutated MS26 gene, leading to complete male sterility but normal vegetative growth and grain yield. Such approaches will need to be standardized and adopted in other important crops, e.g. cotton.
New strategies for cost-effective hybrid seed production, e.g. blending (Nie et al. 2021), apomixis (Lawit 2012), or doubled haploidy (Pazuki et al. 2018), are being explored for a long time, but have not reached the stage of commercial exploitation for hybrid seed production in many species. Recently, Nie et al. (2021) reported that blends of restorer and male sterile lines (AL-type cytoplasmic male sterile lines), mixed in different proportions, can result in a significant increase in hybrid seed production in wheat, though maintaining the desired genetic purity of the hybrid seed remains an issue to be resolved.
Apomixis, which occurs throughout the plant kingdom (Grossniklaus et al. 2001), may provide one such tool for cost-effective hybrid seed production. The ability of angiosperms to reproduce apomictically is a trait having far-reaching consequences in plant breeding and seed production. Among angiosperms, more than 300 plant species from more than 35 families have been described as apomictic, with a distribution pattern that indicates a polyphyletic origin (Khan et al. 2015). Apomictic plants establish genetically stable, seed-propagating clones, which can produce numerous sporophytic true-to-type generations. This reduces the time to develop a new variety and the cost of hybrid seed production. The stabilization of heterozygous genotypes via apomixis is expected to make breeding programmes faster and cheaper, and hence, it has attracted the attention of seed biologists, plant breeders, and commercial seed producers alike. Strategies for introgression of apomixis from wild relatives into cultivated plant species and transformation of sexual genotypes into apomictically reproducing genotypes have been pursued (Barcaccia and Albertini 2013) for decades, but with limited success. Though from the evolutionary perspective apomixis may be considered a consequence of sexual failure and not advancement (Silvertown 2008), the success in apomictic seed development in agricultural species can have a far-reaching impact, specially in non-grain crops. Hence, molecular regulation of apomixis and its use in hybrid seed production will certainly remain an area of much research interest in the coming years.
Since the early reports of microspore embryogenesis by Guha and Maheshwari (1964), and doubled haploidy in maize (Sarkar and Coe 1971), the potential of doubled haploidy (DH) in hybrid production has been explored in many crops. Recent successes in many crops (Chaikam et al. 2019; Kurimella et al. 2021) have widened its scope as both a tool in molecular mapping and development of hybrids and maintenance of the homozygous parental lines for hybrid development and seed production. Standardization of suitable protocols from induction of DH to the seed production of the derived parental lines has, therefore, attracted much attention of both the public and private research institutions and service providers. Further studies will be needed to make this technology cost-effective and applicable to species that are recalcitrant to such manipulations.
3 Prediction of Seed Longevity
The knowledge of the storage potential of the seed lots is of critical value to the farmers and the seedsmen alike in planning the storage of the seed stock for the next sowing or for managing the seed inventory in a commercial seed programme. Since the early reports on seed longevity of different species by Ewart (1908), many systematic studies have been undertaken on the longevity of seeds of a large number of species under different conditions of commercial stores as well as of the gene banks for short-, medium-, and long-term storage (Cromarty et al. 1982; Steiner and Ruckenbauer 1995; Singh et al. 2016; Lu et al. 2018; Solberg et al. 2020). The viability equations proposed and modified by EH Roberts, RH Ellis, and their colleagues (Roberts and Abdalla 1968; Roberts 1973; Ellis and Roberts 1980; Ellis et al. 1989; Ellis and Hong 2007) provided a simple, yet valuable, method of estimating seed viability periods of crop species, based on three factors, i.e. temperature, moisture, and a species constant, based on the interception value of the linear survival curve, which, in spite of some lacunae, are in use. However, predicting seed deterioration and longevity of different species requires long-term storage experiments under ambient conditions and identification of the most sensitive physiological parameter that can pinpoint the state of losing viability. As a seed lot represents a population, in which each seed may be at a different physiological state, single seed analyses are receiving greater attention in recent years (Klaus Mummenhoff 2022). This trend, coupled with ambient storage studies, is expected to present a more accurate way of predicting seed longevity during storage.
With more and more studies showing the influence of anoxia in extending the longevity of seeds held in low M.C., validity of accelerated ageing, which predicts longevity by subjecting seeds to high humidity and high temperature, needs to be revisited. Scientists in the University of Wageningen are exploring the possibility of storing seeds in anoxia (for extending longevity) and developing a rapid seed ageing test under low moisture conditions and elevated partial pressure of oxygen to mimic the ambient storage and as an alternative to accelerated ageing test (Groot et al. 2015; Buijs et al. 2020; https://www.wur.nl/en/Research-Results/Projects-and-programmes/Wageningen-Seed-Science-Centre).
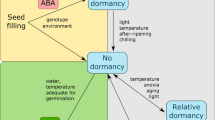
4 Seed Germination and Dormancy
The ability of desiccation-tolerant (orthodox) seeds to remain viable in the quiescent state, followed by resumption of metabolic activities culminating into germination upon hydration when other conditions are favourable, or the inability to do so by remaining dormant, and, finally, the loss of viability in the dry state, is still an enigma to the seed scientists.
Despite being an area widely investigated, critical gaps exist in our understanding of the regulation of seed dormancy and germination. In a recent review, Carrera-Castaño et al. (2020) pointed out the critical role of the seed coverings, and the need to study the genetic manipulation of specific cell wall components involving DOG1 and CWRE genes, and cell wall-related enzymes in the endosperm concerning dormancy and germination. Similarly, DOG1 and RDO5 genes appear to play key roles in dormancy regulation both via ABA accumulation and independent of ABA (Xiang et al. 2014; Fedak et al. 2016; Nonogaki 2019), which need to be elucidated. To understand the significance of RNA modifications such as N6 or N1, adenosine methylation and tissue-specific signalling mechanisms, such as pointed out by Carrera-Castaño et al. (2020), will probably need to be studied at the single-cell level.
A large volume of research point out the central role of the reactive oxygen species (ROS), which include free radicals, such as singlet oxygen (1O2), superoxide (O•−2) or hydroxyl radical (·OH), and non-radical hydrogen peroxide (H2O2), in the regulation of seed germination and dormancy (Baalbaki 2021). Bailly et al. (2008) introduced the concept of the ‘oxidative window for germination’ to explain the dual role of ROS in either signalling for release from dormancy or oxidative damage, which inhibits germination. However, the mechanism of actions of the ROS is far from fully understood. Of these, hydrogen peroxide, though not a free radical, is considered to be the main ROS involved in cellular signalling, as it is more stable and is capable of crossing biological membranes (Petrov and Van Breusegem 2012). Similarly, even though the involvement of plant hormones in regulating seed germination and dormancy is much investigated, their roles need to be enumerated to understand the modulation of gene expression in the presence of specific hormones (Miransari and Smith 2014). Though the involvement of ROS and hormones in seed germination appears quite evident, the underlying mechanisms of the cross-talk between the two and the regulation of these processes are far from clear (Bailly 2019). Thus, studying the genetic control of the production of hormones (ABA, GA, auxins, cytokinins, brassinosteroids, ethylene, etc.), spatiotemporal accumulation of ROS, and the roles of hormones and ROS in manipulating the processes imposing dormancy or triggering germination will remain an active area of seed research.
This is also expected to influence the breeding programmes both for introducing primary dormancy to control pre-harvest sprouting and for releasing physiological deep dormancy to promote germination.
5 Desiccation-Sensitive (Recalcitrant) and Intermediate Longevity Behaviour
In a recent article, Dickie John (2021) drew attention to the problems in ex situ conservation of wild species, many of which are desiccation sensitive and exhibit recalcitrance or intermediate longevity behaviour. This poses a challenge to the managers of the seed banks because unlike the cultivated plant species, where seed multiplication can be undertaken periodically, in the case of wild species, collecting seeds with high initial germination is aimed at, considering the problem of multiplication. Citing a paper by Ali et al. (2007), who observed an initial increase in desiccation tolerance in Anemone nemorosa seeds, followed by a decline, he inferred that continued growth and development of the embryo resulted in loss of desiccation tolerance. He also drew a parallel with the loss of desiccation tolerance in orthodox seeds upon radicle emergence, a benchmark in priming technology, where seed hydration is stopped short of radicle protrusion to arrest the metabolic advancement of seeds without losing the property of desiccation tolerance. This analogy also points out the possibility of learning more about the mechanism of recalcitrance and prolonging their longevity by manipulating the desiccation tolerance trait through investigations of the physiology of primed seeds. Means to protect the longevity of primed seeds might throw more light on the physiological basis of recalcitrance and help find means to extend the longevity of desiccation-sensitive seeds. Recent studies on the re-induction of desiccation tolerance, by either modulating the ROS (Peng et al. 2017) or altering the hormonal regulation (Marques et al. 2019), are promising in understanding the desiccation tolerance and its incorporation in recalcitrant and intermediate seeds.
6 Quantifiable, Universal Vigour Tests
Though good progress has been made in the last 50 years in elucidating the basic mechanisms and development of tests to assess seed vigour in a variety of species, including the large- and small-seeded field crops, vegetables, flowers, forage grasses, and green manuring species, we are still far from having a universal scale which can be used for quantifying seed vigour. Can we predict that wheat seeds recording 85–90% RE (radicle emergence) will certainly result in 85% or more field emergence under normal growing conditions at any location or that the seed lots which recorded 50, 60, and 70% germination after the AA test (or, for that matter, CD test) will result in ~50, ~60, and ~70% germination after ‘x’ months of storage under ambient conditions? Researchers have generated extensive data on both, vigour tests have been standardized based on the criteria of high r values between these parameters (Powell 2022; Marcos-Filho 2015; and Loeffler 2022), and yet these cannot be used for recommending Minimum Standards for Certification for seed vigour.
Since its constitution in 2001, the ISTA Vigour Committee has introduced several vigour tests (ISTA 2022), e.g. the accelerated ageing and conductivity tests for soybean (Glycine max); conductivity test for field bean (Phaseolus vulgaris L.), chickpea (Cicer arietinum L.), and radish (Raphanus sativus); controlled deterioration for Brassica spp.; radicle emergence for corn (Zea mays), radish (Raphanus sativus), and wheat (Triticum aestivum); and tetrazolium vigour test for Glycine max. The work to standardize many more is in progress. From indirect and lengthy tests of yesteryears, such as the glutamic acid decarboxylase activity (GADA) test (Grabe 1964), paper piercing test, and mean germination time (MGT), which required well-trained seed analysts to perform vigour tests, the focus has shifted to direct, rapid, and more robust tests at present. In some instances, a combination of more than one vigour test has been used for a more reliable vigour estimation (Mavi et al. 2016).
In industrialized countries, customers expect 100% seed germination and field emergence and are ready to pay a premium for that, specially in high-value seeds. Therefore, the seed industry focuses on achieving the highest seed performance and looks for precision in vigour assessment. The future vigour tests are expected to be simpler to perform, automated, and preferably non-destructive, based on measuring the ability of individual seeds in a lot to perform under a given set of storage and/or growing conditions. In a recent study, Ermis et al. (2022) found that RE counts between 30 and 34 h were highly correlated (P < 0.01) with normal germination percentage, Ki, and p50. Storing seeds at 75% relative humidity and 35 °C for over 100 days, they constructed seed survival curves based on normal germination using probit analysis and concluded that the RE test can be used to evaluate the seed storage potential of interspecific hybrid crosses of cucurbits. Recent studies using mathematical models for vigour estimation in selected crops based on electrical impedance spectroscopy (Feng et al. 2021), near-infrared spectroscopy (Al-Amery et al. 2018), quantification and profiling of the volatile organic compounds (Umarani et al. 2020; Zhang et al. 2022), and hyperspectral imaging (Yuan et al. 2022) have shown encouraging results, paving the way for further studies under a wide range of seed storage environments and by including more number crops and varieties. Application of mathematical models, combined with the physical, biochemical, and imaging techniques, is foreseen to provide better estimates of seed vigour than simple lab tests.
7 Non-destructive Rapid Assessment of Seed Viability/Germination
Rapid and reliable assessment of the seed viability is required for research purposes, germplasm conservation and regeneration, and management of commercial seed inventory. Research groups both in public research institutions and in the private seed and allied industry are pursuing innovative approaches to achieve this.
Respiration, being the key to all metabolic processes, is most vital in seed germination and seedling development. Thus, it is not surprising that many attempts have been made by the researchers for over a century (Hasegawa 1935; Grabe 1964; Lakon 1928) to identify the most stable and reliable parameter of respiration that can be used for the assessment of seed vigour and viability. While this led to the tetrazolium test for a quick assessment of seed viability (ISTA 1985), continuing efforts are being made to develop an automated, quick, and yet reliable indicator of seed viability and vigour, which can be used commercially for differentiating good seeds from the poor ones.
Oxygen consumption at the beginning of germination is considered an indicator of seed vigour (Reed et al. 2022). Determination of the respiration of individual seeds, thus, is a potential tool for seed quality assessment. A rapid and non-destructive method proposed by Tai-Gi Min and Woo Sik Kang (2011) for assessing the individual seed viability of Brassicaceae species using resazurin reagent (RR) attracted a lot of attention. Recently, a somewhat similar test ‘Seedalive™’ has been proposed for viability assessment. This rapid, non-destructive, and artificial intelligence (AI)-based test was reported to have been tested on seeds from 15 plant families including cultivated and wild species. Being non-destructive, and showing high correlation with germination rate, the test may have a potential application in germplasm conservation and regeneration planning (Klaus Mummenhoff 2022) besides its use in the commercial seed. ‘Fytagoras’, a research-based company, recommends seed respiration analyser (SRA), for a fast (10–72 h) and accurate measurement of individual seed respiration, which may be correlated to seed lot quality. They suggest that SRA, among other things, may be useful as a measuring tool to quickly determine seed viability, in the mapping of multiple germination and seed-related parameters, and in predicting the effect and feasibility of priming methods (https://www.fytagoras.com/en). Both the technologies show merit but need further investigation and appropriate validation.
8 Imaging Technologies for Quality Assessment
Potential of X-ray and other imaging technologies has been explored for a long time for various aspects of seed quality, from the morphological characterization of seed and other plant parts for variety characterization and DUS testing to detection of slight seed damages and insect invasion and visualization of embryonic structures and germinating seeds. Optical sensing and imaging technologies being rapid, non-destructive, and simple are also gaining importance for the quality assessment of single seeds, particularly in the management of seed banks (Hilhorst 2020).
ISTA rules already have provisions for using X-ray images for seed testing. AOSA developed a handbook for X-ray testing of seeds (AOSA 1979), and ISTA has included X-ray test for seed testing with the following objectives:
-
To provide a quick, non-destructive method of differentiating between filled, empty, insect-damaged, and physically damaged seed from the morphological characteristics evident on an X-radiograph.
-
To create a permanent photographic record of the proportions of filled, empty, insect-damaged, and physically damaged seeds in a sample.
Considering the growing importance of innovative technologies in seed programmes, ISTA organized a seed seminar on ‘Advancements and innovation in seed testing: from science to robust test’ during the Congress, 2022, at Cairo, Egypt, which deliberated on the possible uses of different types of imaging technologies for seed quality assessment. Incotec NL has introduced innovative seed sorting systems using X-ray imaging. This fully automated 2D X-ray upgrading technology uses fourth generation of analysis software (based on deep learning) and was found useful in upgrading seeds of tomato and pepper (based on. Among other similar technologies, infrared thermography seemed quite promising in identifying the differences in seed vigour in soybean seed lots (presentations in the ISTA Congress, Cairo, 2022).
Delayed luminescence (DL) is another imaging approach, which is based on the long-term decay of weak photon emissions from materials following exposure to light with a wavelength of 400–800 nm and provides a comprehensive method for measuring biological systems, including seeds (Vesetova et al. 1985). Adeboye and Börner (2020), based on the information available for over 40 years on DL as a possible tool for seed quality assessment, concluded that the diagnostic potentials of DL, based on its ability to penetrate and analyse molecular structures, could be explored for future use in digital varietal discrimination, determination of seed quality based on membrane integrity, and genetic composition of the seed.
Red-blue-green (RBG) and multispectral imaging showed the potential to differentiate seed quality based on the unique images of individual seeds at different wavelengths, which was found particularly useful in detecting seed-borne fungi in sugar beet seeds (Boelt et al. 2018). The imaging technologies combined with machine learning algorithms have made significant advances in seed science (Medeiros et al. 2020), which would support and facilitate the decision-making in the seed industry related to the marketing of seed lots.
Though non-invasive, these technologies are mostly single seed-based, require costly instrumentation and other infrastructure, and hence are costly for wider application. The advancements in the field of artificial intelligence (AI) are expected to reduce the cost and expand the scope.
Automatic imaging-based assessment of seeds and seedlings has shown some quantifiable advantages over visual assessment not only in terms of precision or speed but also in a range of parameters, including length, width, area of the seed or seedling, colours, shape, and other morphological parameters of each seed, which can be recorded simultaneously. This not only offers a more precise approach but also provides information for consistent documentation of the samples by storing the original images for comparison and future use (Jansen 2022, https://analytik.co.uk/webinar-artificial-intelligence-enables-new-ways-in-seed-quality-testing/ accessed 20/04/2022).
The industry prefers to follow a protocol, based on advance seed research, that is suitable for rapid and wider application (not crop- or condition-specific). While all the above-mentioned technologies showed promise, more work is required before these methods can be recommended for wide adoption. AOSA as well as ISTA with its Advanced Technologies Committee, Seed Science Advisory Group, and other specific technical committees would need to critically assess the scientific merits of the new technologies and validate their use in phenotyping and seed quality testing.
9 Application of Molecular Markers for Variety Identity and Seed Testing
Application of the protein and DNA markers in variety identification and hybrid purity testing has been long recognized. ISTA has standardized the protocols for protein markers in establishing variety identity in barley, Pisum and Lolium (SDS-PAGE); wheat (Acid–PAGE); wheat and Triticosecale (SDS-PAGE) and for hybrid purity testing in maize and sunflower (UTLIEF); and SSR markers (using a set of at least eight polymorphic markers) for variety identity in wheat and maize (ISTA Rules, 2022). Jamali et al. (2019), reviewing the applicability of DNA markers in plant variety protection, recognized their efficacy, particularly in case of establishing the status of essentially derived varieties (EDVs). However, the UPOV currently accepts the use of molecular markers only when they perfectly correlate with DUS traits. Knock (2021) questioned the extension of the protection to essentially derived varieties (EDVs) in the UPOV 1991 Convention and expressed concerns that while new breeding technologies (NBT) are the norm of variety development, the inclusion of molecular breeding-derived varieties as EDVs, instead of independent varieties (IV), put these at a serious disadvantage and suggested some modifications in the guidelines so that the innovative breeding is not curtailed.
Notwithstanding the wide application and precision of these markers, the need for using a large number of markers for arriving at a conclusive decision is seen as a deterrent. Hence, the next-generation marker technologies using SNPs (single nucleotide polymorphisms) are being promoted in breeding as well as seed technology (Zhang et al. 2020), which would have to emphasize the precision and stringency of the markers. With cost-effective advanced chip and array technologies consisting of hundreds of thousands of SNP markers, in combination with advanced computing technologies, the use of SNP markers for establishing variety identity and hybrid purity seems only a matter of time before arriving at an acceptable methodology. The advantages of using SNPs for assessing the genetic purity of maize hybrids and parental lines through the use of ddRADseq (double digest restriction-site associated DNA) as the sequencing protocol (Peterson et al. 2012) have been reported. The BMT group of UPOV, in collaboration with ISTA, and after necessary scientific deliberations, will be expected to come out with clear guidelines and protocols for using DNA markers for establishing variety identity, particularly for the essentially derived varieties (EDVs) and gene-edited varieties, which are expected to rise rapidly in the future. Standardization of precise molecular techniques for variety identification, specially for closely related, but independent, varieties, and genetic purity testing will need to be developed parallel to the advances in molecular plant breeding.
With the increasing issues on IPRs of the various varietal technologies and increasing cases of spurious seed in the market, the barcode system based on a set of crop-specific molecular markers needs to be developed for all crop species in commerce. ISTA has recommended protocols for establishing variety identity in wheat and maize using a set of at least eight SSR markers each (ISTA 2022). Making DNA fingerprinting an integral part of crop improvement programme as a regulatory requirement may be considered. A national library of DNA fingerprints needs to be established where a national variety register is maintained. The data of new varieties should be verified with the data available in the national library when a new variety is entered.
Accurate detection of seed-borne viruses, fungi, and bacteria using molecular markers is another important aspect of seed quality that has been pursued vigorously in the last two decades. PCR-based methods, e.g. multiplexed PCR (m-PCR), real-time PCR (RT-PCR), and bio-PCR, have been used effectively in the detection of seed-borne pathogens. However, the cost of equipment and infrastructure and the need for the technical expertise of seed analysts make these somewhat prohibitive. Technologies such as loop-mediated isothermal amplification (LAMP), developed by Notomi et al. (2000), which is a simple, cost-effective, and rapid method for the specific detection of genomic DNA and enables the synthesis of large amounts of DNA in a short time without the use of thermal cycler, may be further refined (see chapter “Molecular Techniques for Testing Genetic Purity and Seed Health” for details).
10 Next-Generation Seed Enhancement Technology
Seed enhancement technologies comprising physical, chemical, and biological treatments, priming, and coating/pelleting are now well recognized as means to improve the planting value of seeds in a wide range of conditions. Seed priming is identified as a potential tool that can provide synchronized, rapid, and on-demand germination, seedling establishment, and resilience against biotic and abiotic stresses improving plant survival in harsh and uncertain environments (Pedrini et al. 2020). Accurate and easy-to-apply seed priming and other enhancement technologies would offer substantial benefits to the rapidly growing business of seedlings (young plants), especially in the cultivation of high-value crops. Despite offering substantial advantages in field emergence under wide-ranging environments, priming is reported to adversely impact the vigour and longevity of seeds, if not sown immediately. Attempts have been made to extend the longevity of primed seeds by storing them in a partial vacuum (Chiu et al. 2003) or low-temperature conditions or induce longevity by exposing them to a hardening treatment (Bruggink et al. 1999). Recently, the advantage of priming for better seedling emergence and vigour has been reported in two desiccation-sensitive tropical rainforest species (Becerra-Vázquez et al. 2020), which not only has a high potential in restoration activities but may also broaden our understanding of the mechanism of recalcitrance.
Another area of growing interest relates to the development of precise seed enhancement technologies using biologicals, specially for organic cultivation, providing protection against pests and pathogens at seedling and early vegetative stages (Singh and Vaishnav 2022). Maintaining the viability and efficacy of fungal inoculum during storage, specially under warm and humid environment, will be a key research area. Nanotechnology is also emerging as a potential technology for supplementing micronutrients, pesticides, tolerants, and seed enhancers, though the standardization of technology and biosafety issues still remain to be addressed.
The future of seed research and technology development appears both exciting and challenging and calls for a closer linkage between the public research institutions, private seed industry, and technology developers cutting the boundaries of specialization. Platforms such as AOSA and ISTA would play important roles in validating their scientific merits and value for application of the new technologies. However, recognizing the fact that the way agriculture continues to be practised in the developing economies with the predominance of smallholders relying on low-cost inputs is quite different from that in the industrialized countries, affordability of quality-enhanced seeds of superior plant varieties will remain a major challenge.
References
Adeboye K, Börner A (2020) Delayed luminescence of seeds: are shining seeds viable? Seed Sci Technol 48:167. https://doi.org/10.15258/sst.2020.48.2.04
Al-Amery M, Geneve RL, Sanches MF, Armstrong PR, Maghirang EB, Lee C, Vieira RD, Hildebrand DF (2018) Near-infrared spectroscopy used to predict soybean seed germination and vigour. Seed Sci Res 28:245–252
Ali N, Probert RJ, Hay FR, Davies H (2007) Post-dispersal embryo growth and acquisition of desiccation tolerance in Anemone nemorosa L. seeds. Seed Sci Res 17(3):155–163
Association of Official Seed Analysts (1979) Radiographic analysis of agricultural and forest tree seed: handbook no. 31, by E Belcher and J Bozzo
Baalbaki R (2021) Editorial. Seed Sci Technol 49(3):321–330. https://doi.org/10.15258/sst.2021.49.3.11
Bailly C (2019) The signalling role of ROS in the regulation of seed germination and dormancy. Biochem J 476:3019–3032
Bailly C, El-Maarouf-Bouteau H, Corbineau F (2008) From intracellular signaling networks to cell death: the dual role of reactive oxygen species in seed physiology. C R Biol 31(10):806–814
Barcaccia G, Albertini E (2013) Apomixis in plant reproduction: a novel perspective on an old dilemma. Plant Reprod 3:159–179
Becerra-Vázquez ÁG, Coates R, Sánchez-Nieto S et al (2020) Effects of seed priming on germination and seedling growth of desiccation-sensitive seeds from Mexican tropical rainforest. J Plant Res 133:855–872
Boelt B, Shrestha S, Salimi Z, Jørgensen JR, Nicolaisen M, Carstensen JM (2018) Multispectral imaging – a new tool in seed quality assessment? Seed Sci Res 28:222–228
Bruggink G, Ooms J, Toorn PV (1999) Induction of longevity in primed seeds. Seed Sci Res 9:49–53
Buijs G, Willems LAJ, Kodde J, Groot SPC, Bentsink L (2020) Evaluating the EPPO method for seed longevity analyses in Arabidopsis. Plant Sci 301:110644
Carrera-Castaño G, Calleja-Cabrera J, Pernas M, Gómez L, Oñate-Sánchez L (2020) An updated overview on the regulation of seed germination. Plan Theory 9(6):703
Chaikam V, Molenaar W, Melchinger AE, Prasanna BM (2019) Doubled haploid technology for line development in maize: technical advances and prospects. Theor Appl Genet 132:3227–3243
Chang Z, Chen Z, Wang N, Xie G, Lu J, Yan W, Zhou J, Tang X, Deng XW (2016) Construction of a male sterility system for hybrid rice breeding and seed production using a nuclear male sterility gene. Proc Natl Acad Sci U S A 113(2016):14145–14150
Chiu KY, Chen CL, Sung JM (2003) Partial vacuum storage improves the longevity of primed sh-2 sweet corn seeds. Sci Hortic 98(2):99–111
Cromarty A, Ellis RH, Roberts EH (1982) The design of seed storage facilities for genetic conservation. In: Handbooks for genebanks, no 1, 100p. International Board for Plant Genetic Resources, Rome
Dickie JB (2021) Editorial. Seed Sci Technol 49(1):73–80
Ellis RH, Hong TD (2007) Quantitative response of the longevity of seed of twelve crops to temperature and moisture in hermetic storage. Seed Sci Technol 35:432–444
Ellis RH, Roberts EH (1980) Improved equations for the prediction of seed longevity. Ann Bot 45:13–30
Ellis RH, Hong TD, Roberts EH (1989) A comparison of low moisture content limit to the logarithmic relation between seed moisture and longevity in 12 species. Ann Bot 63:601–611
Ermis S, Oktem G, Mavi K, Hay F, Demir I (2022) The radicle emergence test and storage longevity of cucurbit rootstock seed lots. Seed Sci Technol 50(1):1–10
Ewart AJ (1908) On the longevity of seeds. Proc R Soc Victoria 21:1–120
Fedak H, Palusinska M, Krzyczmonik K, Brzezniak L, Yatusevich R, Pietras Z, Kaczanowski S, Swiezewski S (2016) Control of seed dormancy in Arabidopsis by a cis-acting noncoding antisense transcript. Proc Natl Acad Sci U S A 113:E7846–E7855
Feng L, Hou T, Wang B, Zhang B (2021) Assessment of rice seed vigour using selected frequencies of electrical impedance spectroscopy. Biosyst Eng 209:53–63
Grabe DF (1964) Glutamic acid decarboxylase activity as a measure of seedling vigor. Proc Assoc Off Seed Anal 54:100–109
Groot SPC, de Groot L, Kodde J, Treuren R (2015) Prolonging the longevity of ex situ conserved seeds by storage under anoxia. Plant Genet Resour Characterization Util 13(1):18–26
Grossniklaus U, Spillane C, Page DR, Kohler C (2001) Genomic imprinting and seed development: endosperm formation with and without sex. Curr Opin Plant Biol 4:21–27
Guha S, Maheshwari SC (1964) In vitro production of embryos from anthers of Datura. Nature 204:497
Hasegawa K (1935) On the determination of viability in seed by reagents. Proc Int Seed Testing Assoc 7:148–153
Hilhorst HWM (2020) Editorial, seed science and technology, vol 48, no 2, pp 315–323
International Rules for Seed Testing (2021), Full Issue I–19-8 (300);https://doi.org/10.15258/ista rules
ISTA (1985) Handbook of tetrazolium testing. Zurich, International Seed Testing Association, p 72
ISTA (2022) International rules for seed testing. International Seed Testing Association, Wallisellen, Switzerland. International Rules for Seed Testing (2021), Full Issue I–19-8 (300). https://doi.org/10.15258/istarules
Jamali SH, Cockram J, Lee T (2019) Insights on deployment of DNA markers in plant variety protection and registration. Theor Appl Genet 132(7):1911–1929
Jansen (2022) https://analytik.co.uk/webinar-artificial-intelligence-enables-new-ways-in-seed-quality-testing/. Accessed 20 Apr 2022
Khan YJ, Choudhary R, Tyagi H, Singh AK (2015) Apomixis: the molecular perspectives and its utilization in crop breeding. J AgriSearch 2(3):153–161
Klaus Mummenhoff (2022) Dead or alive: simple, nondestructive, and predictive monitoring of seedbanks. Presented in ISTA Seed Seminar, “Advancements and innovation in seed testing: from science to robust test”, 8 May 2022. ISTA Congress, Cairo
Kock Michael A (2021) Essentially derived varieties in view of new breeding technologies – plant breeders’ rights at a crossroads. GRUR Int 70(1):11–27
Kurimella RK, Bhatia R, Singh KP, Panwar S (2021) Production of haploids and doubled haploids in Marigold (Tagetes spp.) using anther culture. In: Segui-Simarro JM (ed) Doubled haploid technology. Methods in molecular biology, vol 2289. Humana, New York. https://doi.org/10.1007/978-1-0716-1331-3_18
Lakon G (1928) Ist die Bestimmung der Keimfahigkeit der Samen ohne Keimversuch moglich. Angewandte Botanik (Zeitschrift der Vereinigung fur angewandte Botanik) 10:470
Lawit SJ (2012) Self-reproducing hybrid plants. US20120266324A1 (Google Patents)
Loeffler T (2022) Editorial. Seed Sci Technol 50(1):163–174
Lu X, Chen X, Cui C (2018) Germination ability of seeds of 23 crop plant species after a decade of storage in the National Gene Bank of China. PGR Newslett 139:42–46
Marcos Filho JM (2015) Seed vigour testing: an overview of the past, present and future perspective. Sci Agric 72:363–374
Mariani C, Gossele V, De Beuckeleer M, De Block M, Goldberg RB, De Greef W, Leemans J (1992) A chimeric ribonuclease inhibitor gene restores fertility to male sterile plants. Nature 357:384–387
Marques A, Nijveen H, Somi C, Ligterink W, Hilhorst H (2019) Induction of desiccation tolerance in desiccation sensitive Citrus Limon seeds. J Integr Plant Biol 61:624–638
Mavi K, Powell AA, Matthews S (2016) Rate of radicle emergence and leakage of electrolytes provide quick predictions of percentage normal seedlings in standard germination tests of radish (Raphanus sativus). Seed Sci Technol 44:393–409
Medeiros AD, Silva LJD, Ribeiro JPO, Ferreira KC, Rosas JTF, Santos AA, Silva CBD (2020) Machine learning for seed quality classification: an advanced approach using merger data from FT-NIR spectroscopy and X-ray imaging. Sensors (Basel) 20(15):4319
Min T-G, Kang WS (2011) Simple, quick and nondestructive method for Brassicaceae seed viability measurement with single seed base using resazurin. Hortic Environ Biotechnol 52(3):240–245
Miransari M, Smith DL (2014) Plant hormones and seed germination. Environ Exp Bot 99:110–121
Nie, Kong, Cui, Sang, Mu, Xu, Tian (2021) Blend wheat AL-type hybrid and using SSRs to determine the purity of hybrid seeds. Seed Sci Technol 49(3):275–285
Nonogaki H (2019) Seed germination and dormancy: the classic story, new puzzles, and evolution. J Integr Plant Boil 61:541–563
Notomi T, Okayama H, Masubuchi H, Yonekawa T, Watanabe K, Amino N, Hase T (2000) Loop-mediated isothermal amplification of DNA. Nucleic Acids Res 28(12):E63
Pazuki A, Aflaki F, Gurel S, Ergul A, Ekrem G (2018) Production of doubled haploids in sugar beet (Beta vulgaris): an efficient method by a multivariate experiment. Plant Cell Tissue Org Cult 132:85–97
Pedrini S, Balestrazz A, Mdsen MD, Bhaising K, Hardegree SP, Dixon KW, Kildisheva OA (2020) Seed enhancement: getting seeds restoration-ready. Restor Ecol 28(3):266–275
Peng L, Lang S, Wang Y, Pritchard HW, Wang X (2017) Modulating role of ROS in re-establishing desiccation tolerance in germinating seeds of Caragana korshinskii Kom. J Exp Bot 68:3585–3601
Peterson BK, Weber JN, Kay EH, Fisher HS, Hoekstra HE (2012) Double digest RADseq: an inexpensive method for de novo SNP discovery and genotyping in model and non-model species. PLoS One 7(5):e37135
Petrov VD, Van Breusegem F (2012) Hydrogen peroxide-a central hub for information flow in plant cells. AoB Plants 2012:pls014. https://doi.org/10.1093/aobpla/pls014
Powell AA (2022) Seed vigour in the 21st century. Seed Sci Technol 50(1):45–73
Qi X, Zhang C, Zhu J et al (2020) Genome editing enables next-generation hybrid seed production technology. Mol Plant 13(9):1262–1269
Reed RC, Bradford KJ, Khanday I (2022) Seed germination and vigor: ensuring crop sustainability in a changing climate. Heredity, pp 1–10. https://doi.org/10.1038/s41437-022-00497-2
Roberts EH (1973) Predicting the storage life of seeds. Seed Sci Technol 1:499–514
Roberts EH, Abdalla FH (1968) The influence of temperature, moisture, and oxygen on period of seed viability in barley, broad beans and peas. Ann Bot 32:97–117
Sarkar KR, Coe EH (1971) Origin of parthenogenetic diploids in maize and its implications for the production of homozygous lines. Crop Sci 11(4):543–544
Silvertown J (2008) The evolutionary maintenance of sexual reproduction: evidence from the ecological distribution of asexual reproduction in clonal plants. Int J Plant Sci 169:157–168
Singh HB, Vaishnav A (eds) (2022) New and future developments in microbial biotechnology and bioengineering - sustainable agriculture: revitalization through organic products. Elsevier, Amsterdam, p 366
Singh N, Dash S, Khan YJ (2016) Survival of chickpea, sesame, niger, castor and safflower seeds stored at low and ultra-low moisture contents for 16-18 years. Seed Sci Technol 44:1–14
Solberg SØ, Yndgaard F, Andreasen C, von Bothmer R, Loskutov IG, Asdal Å (2020) Long-term storage and longevity of orthodox seeds: a systematic review. Front Plant Sci 11:1007
Steiner A, Ruckenbauer P (1995) Germination of 110-year-old cereal and weed seeds, the Vienna sample of 1877. Verification of effective ultra-dry storage at ambient temperature. Seed Sci Res 5:195–199
Umarani R, Bhaskaran M, Vanitha C, Tilak M (2020) Fingerprinting of volatile organic compounds for quick assessment of vigour status of seeds. Seed Sci Res 30(2):112–121
Vesetova TV, Veselovsky VA, Rubin AB, Bochvarov PZ (1985) Delayed luminescence of air-dry soybean seeds as a measure of their viability. Physiol Plant 65(4):493–497
Wu Y, Fox TW, Trimnell MR, Wang L, Xu RJ, Cigan AM, Huffman GA, Garnaat CW, Hershey H, Albertsen MC (2016) Development of a novel recessive genetic male sterility system for hybrid seed production in maize and other cross-pollinating crops. Plant Biotechnol J 14:1046–1054
Xiang Y, Nakabayashi K, Ding J, He F, Bentsink L, Soppe WJ (2014) Reduced Dormancy5 encodes a protein phosphatase 2C that is required for seed dormancy in Arabidopsis. Plant Cell 26:4362–4375
Yuan P, Pang L, Wang LM, Yan L (2022) Application of hyperspectral imaging to discriminate waxy corn seed vigour after ageing. Int Food Res J 29(2):397–405
Zhang D, Wu S, An X, Xie K, Dong Z, Zhou Y, Xu L, Fang W, Liu S, Liu S et al (2018) Construction of a multicontrol sterility system for a maize male-sterile line and hybrid seed production based on the ZmMs7 gene encoding a PHD-finger transcription factor. Plant Biotechnol J 16(2018):459–471
Zhang J, Yang J, Zhang L, Luo J, Zhao H, Zhang J, Wen C (2020) A new SNP genotyping technology target SNP-seq and its application in genetic analysis of cucumber varieties. Sci Rep 10(1):5623
Zhang T, Ayed C, Fisk ID, Pan T, Wang J, Yang N, Sun Q (2022) Evaluation of volatile metabolites as potential markers to predict naturally-aged seed vigour by coupling rapid analytical profiling techniques with chemometrics. Food Chem 367:130760
Acknowledgements
Sincere gratitude to Dr. H.S. Gupta, Former Director, ICAR-IARI, Dr. D. Vijay and Dr. S.K. Chakrabarty, Principal Scientists, Div. Seed Science & Technology, ICAR-IARI, and Dr. Partha R. Das Gupta, Emeritus Advisor, Syngenta Foundation, India, for their insightful observations and valuable inputs in preparing this chapter.
Author information
Authors and Affiliations
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2023 The Author(s)
About this chapter

Cite this chapter
Dadlani, M. (2023). Emerging Trends and Promising Technologies. In: Dadlani, M., Yadava, D.K. (eds) Seed Science and Technology. Springer, Singapore. https://doi.org/10.1007/978-981-19-5888-5_17
Download citation
DOI: https://doi.org/10.1007/978-981-19-5888-5_17
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-19-5887-8
Online ISBN: 978-981-19-5888-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)