
Abstract
Over the last few decades, population increases of sika deer (Cervus nippon) have become a major issue in various forest ecosystems across temperate regions; however, the influences of deer browsing on riparian forests are less known. In this chapter, we illustrate the herbivore–forest vegetation relationships over a long term from the past when deer was absent to the current when deer became overabundant in an old-growth riparian forests of Ooyamazawa, the Chichibu Mountains of central Japan. We revealed that (1) the browsing activity of deer has a negative influence on riparian forests, (2) the damage of these species is mainly induced by easiness to browsing by deer resulting from small tree size structure, and (3) the resistance to the deer browsing differs among tree species. Thus, small mature trees (i.e., shrub species) with low browsing resistance should be primarily protected for effective management of riparian forests.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
1 Introduction
Browsing by large herbivores can drastically modify the structure, environment, and tree-species composition of forests (Gill 1992a, b). Over the last few decades, sika deer (Cervus nippon) populations have increased due to the absence of predators and a gradual decline in hunting, and have become a major issue for forest ecosystems in several regions of Japan (Uno et al. 2007; Takatsuki 2009; Kaji et al. 2010). In places where deer density is too high, plant-species diversity has decreased due to a drastic decline in forest-floor vegetation (Suda et al. 2001; Suzuki et al. 2008) and the increased mortality of palatable plant species (Akashi and Nakashizuka 1999; Yokoyama et al. 2001) caused by heavy browsing and debarking activities of deer. Thus, the adverse effects of these activities on the regeneration of tree species in natural forests have become a major concern (Takatsuki and Gorai 1994; Akashi and Nakashizuka 1999; Suda et al. 2001; Yokoyama et al. 2001). Previous studies have shown that increasing deer populations can also have other impacts on forest ecosystems and resources. For example, they may reduce the quality of timber resources (Kaji et al. 2000; Oi and Suzuki 2001; Ueda et al. 2002; Akashi and Terazawa 2005; Suzuki et al. 2008), decrease the diversity of insect communities (Kanda et al. 2005; Ueda et al. 2009), alter the structure and function of soil microbial food webs (Niwa et al. 2011), and increase the risks of soil erosion and landslides in mountainous areas (Furusawa et al. 2003).
An understanding of herbivore–vegetation relationships is necessary for the assessment of potential risks and design of management plans. In Japan, deer consume forest-floor vegetation and tree leaves from spring to autumn and utilize tree bark and dead fallen leaves when facing food shortages, which usually occur in winter (Takahashi and Kaji 2001; Takatsuki 2009). Browsing damage from deer occurs intensively on certain plant species among a wide array of co-occurring species (Gill 1992a; Augustine and McNaughton 1998; Côté et al. 2004; Boulangera et al. 2009). Tree-species composition and food-resource availability in forests affect the dietary selections of deer (Gill 1992a; Moser et al. 2006; Jayasekara and Takatsuki 2000; Takahashi and Kaji 2001). Many researchers have proposed factors to explain differences in the plant-species preferences of deer, including the morphological and physiological traits of plants (Gill 1992a; Ando et al. 2003; Jiang et al. 2005; Moser et al. 2006; Sauvé and Côté 2006). For example, smaller trees are well known to be preferentially browsed by deer (Gill 1992a; Boulangera et al. 2009; Didion et al. 2009) because they have more leaves and branches at heights easily accessible to deer, as well as thin and soft bark. The trunk size of these trees is also suitable for antler fraying . Large-herbivore densities also affect the occurrence and intensity of browsing damage on plants (Gill 1992a). However, the temporal changes in dietary selection and browsing damage on plants associated with deer density changes have been poorly documented. In particular, information about the initial changes that occur when an area transitions from the absence to overabundance of deer is lacking.
Riparian forests are important and unique ecosystems that provide specialized habitats and resources for wildlife, support and maintain faunal and floral diversity, and prevent soil erosion in precipitous mountain terrain (Malanson 1993; Richards et al. 2002; Ward et al. 2002). The majority of riparian forests in Japan disappeared during the twentieth century due to increased industrial or agricultural land use and the construction of embankments for river management (Sakio 2008); several of the remaining forests are being subjected to heavy browsing pressure by deer. In an old-growth riparian forest dominated by Ulmus davidiana Planch. var. japonica (Rehder) Nakai in the district of Nikko in central Japan, heavy browsing and debarking by deer have resulted in the decline of forest-floor vegetation and dieback of mature trees (Nomiya et al. 2003). The deer population is expected to continue to increase and expand throughout the less-snowy regions of Japan. Hence, riparian forests are expected to be subject to increasingly serious damage due to deer browsing activity.
In this chapter, we illustrate long-term herbivore–forest vegetation relationships in an old-growth riparian forest of Ooyamazawa, in the Chichibu Mountains of central Japan, from the past, when deer were absent, to the present, when deer are overabundant. Using long-term tree-census data, we elucidated the temporal changes in the dietary selections of deer and species-specific differences in browsing damage. Finally, we discuss effective management plans for riparian forests.
2 Ooyamazawa Riparian Forest
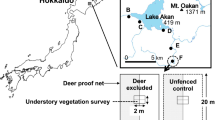
In Ooyamazawa (35°57′30″N, 138°46′32″E, 1200–1620 m a.s.l.), an old-growth natural forest lies along a mountain stream of the Arakawa River in part of Chichibu-Tama-Kai National Park, Saitama Prefecture, central Japan (Fig. 9.1). The annual mean temperature, annual precipitation, and maximum snow depth in the nearest settlement (Nakatsugawa; 700 m a.s.l. and 4.6 km from the study site) are 10.7 °C, 1100 mm, and 30 cm, respectively (Sakio 1997). The estimated annual mean temperature at the study site (1450 m a.s.l.) is 6.5 °C, based on a temperature lapse rate of 0.6 °C per 100-m increase in elevation (Sakio 1997). The site lies in the upper part of a cool-temperate, deciduous broad-leaf forest zone (Sakio 1997). The dominant species of the forest canopy are Flaxinus platypoda Oliv., Pterocarya rhoifolia Siebold et Zucc., and Cercidiphyllum japonicum Siebold et Zucc. ex Hoffm. et Schult., which are more than 30 m in height (Table 9.1). The subcanopy is dominated by Acer shirasawanum Koidz. and Acer pictum Thunb. The understory is composed primarily of Acer carpinifolium Siebold et Zucc. and Acer argutum Maxim (Table 9.1). Before deer became abundant, the forest floor had dense vegetation cover with greater plant-species richness, including Parasenecio tebakoensis (Makino) H.Koyama, Parasenecio delphiniifolius (Siebold et Zucc.) H.Koyama, Chrysosplenium macrostemon Maxim. var. macrostemon, Laportea bulbifera (Siebold et Zucc.) Wedd., Impatiens noli-tangere L., Dryopteris crassirhizoma Nakai, Dryopteris polylepis (Franch. et Sav.) C.Chr., and Polystichum tripteron (Kunze) C.Presl (Sakio et al. 2013).

Index and location maps of the study area
In the Ooyamazawa riparian forest, a permanent study plot (4.71 ha; Fig. 9.1) consisting of three subplots (upper, middle, and lower) was established, and tree censuses were conducted at each subplot at 5-year intervals from 1991 to 2008. The upper and middle subplots (3.2 ha) were established in 1991, and the lower subplot (1.5 ha) was established in 1998. All living trees with diameters at breast height (DBHs) > 4 cm were numbered and identified to the species level. Tree DBHs, alive/dead status, and extent of browsing damage by deer were recorded. An initial survey was conducted at the upper subplot in 1991 and at the middle subplot in 1992; 1982 standing trees of 42 species were observed and measured, and the total basal area was 36.3 m2 ha−1. A second survey was conducted in the upper, middle, and lower subplots from 1996 to 1998. The third and fourth surveys were conducted from 2001 to 2003 and 2006 to 2008, respectively. The second, third, and fourth surveys included 2396 standing trees of 46 species, 2335 standing trees of 45 species, and 2050 standing trees of 41 species, and the total basal areas were 34.2, 35.2, and 35.5 m2 ha−1, respectively. The entire permanent plot was designated as an associate site of the Japan Long-Term Ecological Research Network in 2006, and the middle subplot has been a core site of the nationwide Japanese Monitoring Sites 1000 program since 2008.
3 Increase in Deer Density Around Ooyamazawa
Deer density has been monitored using the block-count method for more than 30 years as part of density monitoring for a protected species, Japanese serow (Capricornis crispus) (Saitama Museum of Natural History 1983; Gunma, Saitama, Tokyo, Yamanashi and Nagano Prefectural Boards of Education 1988, 2010). During the 1980s and 1990s, the deer density around the study site was 0–6.3 heads/km2. This density began to increasing in the late 1990s, and had reached 20.9 heads/km2 in the 2000s (Saitama Museum of Natural History 1983; Gunma, Saitama, Tokyo, Yamanashi and Nagano Prefectural Boards of Education 1988, 2010). The deer censuses were performed using the block-count method on only single days every few years. The visibility of the census site is well known to affect the accuracy of density estimation when using this method (Maruyama and Furubayashi 1983). In Ooyamazawa and its surroundings, the slope is very steep and the geomorphology is complex. Therefore, the actual densities may be greater than estimated. The deer are permanent, year-round residents of the forest.
4 Temporal Changes in Browsing Damage
Grazing and browsing of overabundant large herbivores drastically change the structure and species composition of forest vegetation (Ripple and Beschta 2008; Didion et al. 2009; Salk et al. 2011). By the 2010s, most herbaceous plants and the seedlings and saplings of all tree species had disappeared due to heavy deer foraging pressure (Sakio et al. 2013). On the forest floor, only unpalatable and poisonous plants , such as Veratrum album subsp. oxysepalum (Turcz.) Hultén, Aconitum tonense Nakai ex H.Hara, and Scopolia japonica Maxim., remain in low abundance (Sakio et al. 2013).
Many studies have shown that heavy browsing and debarking activities of deer result in failed regeneration of tree species and reduced plant-species diversity (Takatsuki and Gorai 1994; Akashi and Nakashizuka 1999; Suda et al. 2001; Yokoyama et al. 2001; Suzuki et al. 2008). Similar phenomena were observed in the Ooyamazawa riparian forest. In the first survey, no trace of browsing damage was found on any standing tree in the upper or middle subplot (Table 9.2). In the second survey, evidence of debarking was found on three trees of U. laciniata and one tree of A. shirasawanum (Table 9.3). Thereafter, evidence of deer browsing increased rapidly; it was observed on 170 trees of eight species in the third survey. Among the damaged species, A. carpinifolium and A. argutum showed considerably increased browsing damage, which affected approximately 20 and 25%, respectively, of the total number of trees of these species. The three dominant canopy species, F. platypoda, C. japonicum, and P. rhoifolia, exhibited no evidence of browsing damage. By the fourth survey, browsing damage had increased to 536 trees of 19 species, including some trees of the dominant canopy species (Table 9.3). The likelihood ratio test indicated that browsing damage had increased significantly throughout the survey period (P < 0.001), and that the extent of damage varied among species (P < 0.001). However, the interaction between year and species was not significant (P = 0.96).
5 Species and Size Preferences, and Mortality
Among the browsed tree species, U. laciniata and A. argutum had significantly higher ratios of damaged trees among the total number of trees in the forest (P < 0.01), and higher Ivlev’s modified electivity indices (D values) in all surveys (Table 9.3). The index D i (Jacobs 1974) for species i was calculated as follows:
where r i is the ratio of the number of damaged trees of the ith species to the total number of damaged trees, and P i is the ratio of the total number of trees of the ith species to the total number of trees (Jacobs 1974). The index D is 1 for preferred species and −1 for non-preferred species. The D values for P. hispida and A. carpinifolium increased gradually throughout the survey period, reaching 0.93 and 0.68, respectively, by the fourth survey. By contrast, F. platypoda and A. pictum had lower proportions of damaged trees among the total number of trees (P < 0.01) and lower D values. Initially, browsing damage was found primarily on trees with smaller DBHs (4–20 cm); its distribution then expanded gradually to include larger trees (DBH > 20 cm; Fig. 9.2). The likelihood ratio test indicated that the DBH correlated negatively with browsing damage caused by the deer (P < 0.01). This correlation was particularly strong for U. laciniata, A. argutum, and A. carpinifolium (Fig. 9.3). Exceptionally, the DBH showed no obvious relationship to browsing damage to trees of P. rhoifolia and C. japonicum. During the tree census, we recorded browsing-damage levels and divided them into three categories: (1) feeding on leaves and stems; (2) debarking, including the fraying of bark; and (3) a combination of feeding and debarking damage (Fig. 9.4). Evidence of feeding on leaves and stems, debarking, and combined feeding and debarking damage was found on almost all dominant and abundant species in 2006–2008 (Table 9.3). Among the excessively browsed species, debarking damage occurred more frequently than feeding damage on U. laciniata and A. shirasawanum. By contrast, A. carpinifolium and P. hispida were subjected to heavier feeding on leaves and stems than on bark. Feeding and debarking damage were recorded in equal quantities on A. argutum. The dieback of damaged trees of excessively browsed species increased until the fourth survey (likelihood ratio test, P < 0.01; Fig. 9.3, Table 9.3). However, the mortality of damaged trees varied among species. For example, the mortality rates of U. laciniata, A. argutum, and P. hispida were high, whereas that of A. carpinifolium was relatively low (Table 9.3).

Temporal changes in tree-size preference by sika deer and DBH distributions of all trees in the survey area of the Ooyamazawa riparian forest from the second through fourth surveys (upper three panels). The lower three panels indicate the relationship between DBH and the presence/absence of browsing damage, and estimated probabilities derived using generalized linear mixed models

Temporal changes in browsing damage by sika deer and DBH distributions (number of standing trees in 4.71 ha) of the main 10 tree species in the Ooyamazawa riparian forest from the second through fourth surveys. Browsing damage included feeding damage to leaves and stems, bark stripping , and fraying damage to bark

Feeding on leaves and stems of P. hispida (left) and debarking of A. carpinifolium (right) by sika deer
These results suggest that A. argutum, U. laciniata, P. hispida, and A. carpinifolium were preferentially browsed by deer compared with F. platypoda, A. pictum, and A. shirasawanum (Table 9.3). The diet of deer is determined by the physical ease of feeding, and the nutritional and lignin contents of plants (Gill 1992a; Jiang et al. 2005; Kojima et al. 2006). The major damage to A. argutum, A. carpinifolium, and P. hispida may have been caused by the relatively small size of these three species in the forest (Table 9.1). In general, DBH and tree height variation differ among species in natural forests. These three species have relatively low tree heights and small maximum DBHs, in addition to producing many sprouts without scarring caused by disturbances. Smaller trees have more leaves and stems at lower heights, accessible to deer. In addition, bark becomes thicker, harder, rougher, and more suberizing with increasing tree diameter. Therefore, these three species may be physically easy to feed on by deer. By contrast, F. platypoda and A. pictum have greater tree heights and trunk diameters. The forest-floor vegetation, which includes the seedlings and saplings of these two species, has almost disappeared due to heavy browsing pressure of deer, except for poisonous herbs such as Veratrum album subsp. oxysepalum (Turcz.) Hultén Aconitum tonense Nakai ex H.Hara, and Scopolia japonica Maxim. Thus, the lower preference for these species by deer, despite their palatability, may be caused by the presence of fewer leaves and stems at lower heights, in addition to thicker bark relative to other species. These results suggest that deer browsing activity especially reduces tree-species diversity of certain functional types, such as shrub species (i.e., species with relatively low tree heights and small maximum DBHs). On the other hand, U. laciniata was subject to the most damage among the four preferred species. The greater preference for this species has also been confirmed in previous studies (Imagawa and Tanaka 1996; Kaji et al. 2010). This species may be targeted preferentially due to the low lignin content in its bark (Imagawa and Tanaka 1996; Kojima et al. 2006) and the physical ease of strip-peeling (Imagawa and Tanaka 1996).
6 Resistance of Trees to Deer Browsing
Heavy browsing, especially bark-stripping by large herbivorous mammals, induces the dieback of damaged trees (Gill 1992b; Akashi and Nakashizuka 1999). The increase in dieback of damaged trees among the four preferred species in our study (A. argutum, U. laciniata, P. hispida, and A. carpinifolium) during 2006–2008 suggests that deer browsing caused serious damage to tree species in this old-growth riparian forest (Fig. 9.3, Table 9.3). However, the mortality rates differed among the four species. Acer argutum and U. laciniate, which had high mortality rates, were subjected to more debarking than feeding damage. Conversely, A. carpinifolium had a lower mortality rate than did A. argutum and U. laciniata, even though 79 trees of A. carpinifolium were affected by debarking damage and 264 trees showed damage caused by feeding on stems and leaves (Table 9.3). These results indicate that these four species have different levels of resistance to deer browsing, with the highest being shown by A. carpinifolium. Similar findings were obtained in several previous studies based on simulated experiments of browsing and debarking (Noel 1970; Bergström and Danell 1987; Delvaux et al. 2010). The high resistance of A. carpinifolium is considered to be due to its high rates of adventitious bud production and regrowth of bark, including the cambium layer and phloem, after browsing and debarking.
7 Effective Management of Ooyamazawa
Previous studies have suggested that browsing by sika deer adversely affects the regeneration of tree species in natural forests (Takatsuki and Gorai 1994; Akashi and Nakashizuka 1999; Suda et al. 2001; Yokoyama et al. 2001). Our study reveals an increase in serious damage to tree species in an old-growth riparian forest due to 18 years of heavy browsing activity by deer. Riparian forests have important and unique ecosystem functions (Malanson 1993; Richards et al. 2002; Ward et al. 2002); however, the areas of these forests in mountain landscapes are smaller than those of other forests on mountain slopes, such as Fagus crenata forest. Therefore, deer browsing appears to negatively impact not only riparian forests, but also mountain forest ecosystems . If the deer population density is maintained at the current level or increases further, modification of the forest structure and composition will be induced by the failure of preferred species to regenerate and the dieback of trees with small diameters. To maintain the ecosystems of riparian forests, immediate preventive management is required before browsing damage by deer expands.
Deer-exclusion fences are an effective means of forest management and have been used widely throughout Japan. However, these fences can protect only limited areas of forest, as installation and maintenance costs are very high. The control of the deer population through hunting or culling is an alternative method, and the number of sika deer harvested has increased rapidly over the last few decades (Takatsuki 2009). However, population control by hunting and culling is expected to be insufficient because the number of hunters is gradually declining in Japan (Takatsuki 2009). Hence, temporal and immediate activities, such as the selective conservation of tree-seed sources, are needed until a viable management program becomes operational.
In the Ooyamazawa riparian forest, extensive browsing damage and increased tree mortality were found for U. laciniata, A. argutum, and P. hispida. Therefore, in other riparian forests of similar species composition and size structure, mature trees of these three species with small diameters (i.e., shrub species) should be primarily protected and managed through the use of metallic-mesh trunk protectors.
References
Akashi N, Nakashizuka T (1999) Effects of bark-stripping by sika deer (Cervus nippon) on population dynamics of a mixed forest in Japan. For Ecol Manage 113:75–82
Akashi N, Terazawa K (2005) Bark stripping damage to conifer plantations in relation to the abundance of sika deer in Hokkaido, Japan. For Ecol Manage 208:77–83
Ando M, Yokota H, Shibata E (2003) Bark stripping preference of sika deer, Cervus nippon, in terms of bark chemical content. For Ecol Manage 177:323–331
Augustine DJ, McNaughton SJ (1998) Ungulate effects on the functional species composition of plant communities: herbivore selectivity and plant tolerance. J Wildl Manage 62:1165–1183
Bergström R, Danell K (1987) Effects of simulated winter browsing by moose on morphology and biomass of two birch species. J Ecol 75:533–544
Boulangera V, Baltzingera C, Saïdc S, Ballona P, Picardb JF, Dupoueyb JL (2009) Ranking temperate woody species along a gradient of browsing by deer. For Ecol Manage 258:1397–1406
Côté SD, Rooney TP, Tremblay JP, Dussault C, Waller DM (2004) Ecological impacts of deer overabundance. Annu Rev Ecol Evol Syst 35:113–147
Delvaux C, Sinsin B, Damme PV (2010) Impact of season, stem diameter and intensity of debarking on survival and bark re-growth pattern of medical tree species, Benin, West Africa. Biol Conserv 143:2664–2671
Didion M, Kupferschmid AD, Bugmann H (2009) Long-term effects of ungulate browsing on forest composition and structure. For Ecol Manage 258S:S44–S55
Furusawa H, Miyanishi H, Kaneko S, Hino T (2003) Movement of soil and litter on the floor of a temperate mixed forest with an impoverished understory grazed by deer (Cervus nippon centralis temminck). J Jpn For Soc 85:318–325 (in Japanese with English summary)
Gill R (1992a) A review of damage by mammals in north temperate forests. 1. Deer. Forestry 65:145–169
Gill R (1992b) A review of damage by mammals in north temperate forests. 3. Impact on trees and forests. Forestry 65:363–388
Gunma, Saitama, Tokyo, Yamanashi and Nagano Prefectural Boards of Education (eds) (1988) Bulletin report of Japanese Serow (Capricornis crispus) protection area in Kanto mountains. Gunma, Saitama, Tokyo, Yamanashi and Nagano Prefectural Boards of Education, Tokyo (in Japanese)
Gunma, Saitama, Tokyo, Yamanashi and Nagano Prefectural Boards of Education (eds) (2010) Bulletin report of Japanese Serow (Capricornis crispus) protection area in Kanto mountains. Gunma, Saitama, Tokyo, Yamanashi and Nagano Prefectural Boards of Education, Tokyo (in Japanese)
Imagawa H, Tanaka K (1996) Damage of forest trees by sika deer. Possible reason for the palatability. For Tree Breed Hokkaido 39:10–13 (in Japanese)
Jacobs J (1974) Quantitative measurement of food selection. A modification of the forage ratio and Ivlev’s electivity index. Oecologia 14:413–417
Jayasekara P, Takatsuki S (2000) Seasonal food habits of a sika deer population in the warm temperate forest of the westernmost part of Honshu, Japan. Ecol Res 15:153–157
Jiang Z, Ueda H, Kitahara M, Imaki H (2005) Bark stripping by sika deer on veitch fir related to stand age, bark nutrition, and season in northern mount Fuji district, central Japan. J For Res 10:359–365
Kaji K, Miyaki M, Saitoh T, Ono S, Kaneko M (2000) Spatial distribution of an expanding sika deer population on Hokkaido Island, Japan. Wildl Soc Bull 28:699–707
Kaji K, Saitoh T, Uno H, Matsuda H, Yamamura K (2010) Adaptive management of sika deer populations in Hokkaido, Japan: theory and practice. Popul Ecol 52:373–387
Kanda N, Yokota T, Shibata E, Sato H (2005) Diversity of dung-beetle community in declining Japanese subalpine forest caused by an increasing sika deer population. Ecol Res 20:135–151
Kojima Y, Yasui Y, Orihashi K, Terazawa M, Kamoda S, Kasahara H, Takahashi Y (2006) Relationship between bark preferences of Cervus nippon yesoensis and inner bark components of small-diameter tree trunk. J Jpn For Soc 88:337–341 (in Japanese with English summary)
Malanson G (1993) Riparian landscapes. Cambridge University Press, Cambridge
Maruyama N, Furubayashi K (1983) Preliminary examination of block count method for estimating number of sika deer in Fudakake. J Mamm Soc J 9:274–278 (in English with Japanese summary)
Moser B, Schütz M, Hindenlang KE (2006) Importance of alternative food resources for browsing by roe deer on deciduous trees: the role of food availability and species quality. For Ecol Manage 226:248–255
Niwa S, Mariani L, Kaneko N, Okada H, Sakamoto K (2011) Early-stage impacts of sika deer on structure and function of the soil microbial food webs in a temperate forest: a large-scale experiment. For Ecol Manage 261:391–399
Noel A (1970) The girdled tree. Bot Rev 36:162–195
Nomiya H, Suzuki W, Kanazashi T, Shibata M, Tanaka H, Nakashizuka T (2003) The response of forest floor vegetation and tree regeneration to deer exclusion and disturbance in a riparian deciduous forest, central Japan. Plant Ecol 164:263–276
Oi T, Suzuki M (2001) Damage to sugi (Cryptomeria japonica) plantations by sika deer (Cervus nippon) in northern Honshu, Japan. Mammal Study 26:9–15
Richards K, Brasington J, Hughes F (2002) Geomorphic dynamics of floodplains: ecological implications and a potential modelling strategy. Freshwat Biol 47:559–579
Ripple WJ, Beschta RL (2008) Trophic cascades involving cougar, mule deer, and black oaks in Yosemite National Park. Biol Conserv 141:1249–1256
Saitama Museum of Natural History (eds) (1983) Bulletin report of special natural monument: density of Japanese Serow. Saitama Prefectural Boards of Education (in Japanese)
Sakio H (1997) Effects of natural disturbance on the regeneration of riparian forests in a Chichibu mountains, central Japan. Plant Ecol 132:181–195
Sakio H (2008) Features of riparian forests in Japan. In: Sakio H, Tamura T (eds) Ecology of riparian forests in Japan: disturbance life history and regeneration. Springer, Tokyo, pp 3–11
Sakio H, Kubo M, Kawanishi M, Higa M (2013) Effects of deer on forest floor vegetation in the Chichibu mountains, Japan, J Jpn Soc Reveg Technol 39:226–231 (in Japanese with English summary)
Salk TT, Frelich LE, Sugita S, Calcote R, Ferrari JB, Montgomery RA (2011) Poor recruitment is changing the structure and species composition of an old-growth hemlock-hardwood forest. For Ecol Manage 261:1998–2006
Sauvé DG, Côté SD (2006) Winter frage selection in white-tailed deer at high density: balsam fir is the best of a bad choice. J Wildl Manage 71:911–914
Suda K, Araki R, Maruyama N (2001) The effects of sika deer on the structure and composition of the forests on the Tsushima islands. Biosphere Conserv 4:13–22
Suzuki M, Miyashita T, Kabaya H, Ochiai K, Asada M, Tange T (2008) Deer density affects ground-layer vegetation differently in conifer plantations and hardwood forests on the Boso peninsula, Japan. Ecol Res 23:151–158
Takahashi H, Kaji K (2001) Fallen leaves and unpalatable plants as alternative foods for sika deer under food limitation. Ecol Res 16:257–262
Takatsuki S (2009) Effects of sika deer on vegetation in Japan: A review. Biol Conserv 142:1922–1929
Takatsuki S, Gorai T (1994) Effects of sika deer on the regeneration of a Fagus crenata forest on Kinkazan Island, northern Japan. Ecol Res 9:115–120
Ueda A, Hino T, Ito H (2009) Relationships between browsing on dwarf bamboo (Sasa nipponica) by sika deer and the structure of ground beetle (coleoptera: Carabidae) assemblage. J Jpn For Soc 91:111–119 (in Japanese with English summary)
Ueda H, Takatsuki S, Takahashi Y (2002) Bark stripping of hinoki cypress by sika deer in relation to snow cover and food availability on Mt Takahara, central Japan. Ecol Res 17:545–551
Uno H, Yokoyama M, Sakata H, the Working Group for Sika Deer Management Mammalogical Society of Japan (2007) Current status of and perspectives on conservation and management for sika deer populations in Japan. Mamm Sci 47:25–38 (in Japanese with English summary)
Ward JV, Tockner K, Arscott DB, Claret C (2002) Riverine landscape diversity. Freshwat Biol 47:517–539
Yokoyama S, Maeji I, Ueda T, Ando M, Shibata E (2001) Impact of bark stripping by sika deer, Cervus nippon, on subalpine coniferous forests in central Japan. For Ecol Manage 140:93–99
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2020 The Author(s)
About this chapter

Cite this chapter
Higa, M., Kawanishi, M., Kubo, M., Sakio, H. (2020). Temporal Changes in Browsing Damage by Sika Deer in a Natural Riparian Forest in Central Japan. In: Sakio, H. (eds) Long-Term Ecosystem Changes in Riparian Forests. Ecological Research Monographs. Springer, Singapore. https://doi.org/10.1007/978-981-15-3009-8_9
Download citation
DOI: https://doi.org/10.1007/978-981-15-3009-8_9
Published:
Publisher Name: Springer, Singapore
Print ISBN: 978-981-15-3008-1
Online ISBN: 978-981-15-3009-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)