
Zusammenfassung
Schlüsselkonzepte
-
1.
Die somatosensorische Wahrnehmung basiert auf der Verarbeitung mechano-, thermo- und propriozeptiver Signale, die in parallelen Kanälen weitergeleitet und im Gehirn zusammengeführt werden.
-
2.
Schnell adaptierende Sensoren registrieren Informationen über Veränderungen eines Reizes, während langsam adaptierende Sensoren Reizstärke und -dauer repräsentieren.
-
3.
Somatosensorische Reize werden in peripheren Nervenendigungen in ein depolarisierendes Rezeptorpotenzial umgewandelt, dessen Amplitude die Stärke des Reizes codiert.
-
4.
Die sensorische Bedeutung einer Körperregion für einen Organismus korreliert unmittelbar mit der Größe der kortikalen Repräsentation dieses Areals.
-
5.
Nichtschmerzhafte Kälte- und Wärmereize werden von unterschiedlichen Rezeptoren detektiert und in getrennten Signalbahnen verarbeitet.
-
6.
Schmerzen sind subjektive Empfindungen, die im Gehirn erzeugt werden und ein wichtiges Warnsignal darstellen. Sie beruhen auf einer erhöhten Aktivität nozizeptiver Neurone.
-
1.
Die somatosensorische Wahrnehmung basiert auf der Verarbeitung mechano-, thermo- und propriozeptiver Signale, die in parallelen Kanälen weitergeleitet und im Gehirn zusammengeführt werden.
-
2.
Schnell adaptierende Sensoren registrieren Informationen über Veränderungen eines Reizes, während langsam adaptierende Sensoren Reizstärke und -dauer repräsentieren.
-
3.
Somatosensorische Reize werden in peripheren Nervenendigungen in ein depolarisierendes Rezeptorpotenzial umgewandelt, dessen Amplitude die Stärke des Reizes codiert.
-
4.
Die sensorische Bedeutung einer Körperregion für einen Organismus korreliert unmittelbar mit der Größe der kortikalen Repräsentation dieses Areals.
-
5.
Nichtschmerzhafte Kälte- und Wärmereize werden von unterschiedlichen Rezeptoren detektiert und in getrennten Signalbahnen verarbeitet.
-
6.
Schmerzen sind subjektive Empfindungen, die im Gehirn erzeugt werden und ein wichtiges Warnsignal darstellen. Sie beruhen auf einer erhöhten Aktivität nozizeptiver Neurone.
If you prick us do we not bleed? If you tickle us do we not laugh? If you poison us do we not die? And if you wrong us shall we not revenge?
William Shakespeare
Das somatosensorische System verarbeitet eine Vielzahl verschiedener Reize, die von unterschiedlichen Mechanosensoren in neuronale Signale umgewandelt und in zahlreiche kortikale Hirnregionen übertragen werden. Daher wird das somatosensorische System in mehrere Subsysteme unterteilt:
-
Die mechanische Oberflächensensibilität umfasst Informationen über Druck, Berührung, Spannung und Vibration. Sie wird durch Mechanosensoren in der Haut vermittelt. Die Oberflächensensibilität ermöglicht die Identifizierung und Handhabung von Objekten, ohne auf visuelle oder auditorische Signale angewiesen zu sein.
-
Die Thermosensibilität basiert auf der Aktivierung peripherer Warm- und Kaltsensoren, die auf Änderungen der Umgebungstemperatur reagieren.
-
Die Propriozeption ist Grundlage für die Wahrnehmung der Lage des Körpers im Raum, insbesondere der Position der Extremitäten. Neben dem Vestibularorgan (Abschn. 14.5) tragen Muskelspindeln und Sehnenorgane (Abschn. 12.3) in der Skelettmuskulatur zu dieser Empfindung bei.
-
Der Schmerzsinn (Nozizeption ) ist ein außerordentlich wichtiger Schutzmechanismus, der uns vor Gefahren in Form gewebeschädigender Reize warnt. Juckempfindungen – lange Zeit als eine unterschwellige Form des Schmerzes interpretiert – werden sehr wahrscheinlich durch spezifische, von der Schmerzbahn unabhängige Neurone vermittelt.
-
Die Viszerozeption registriert Signale aus den Eingeweiden.
Es ist eine interessante Erfahrung, sich einmal für einige Minuten die Augen zu verbinden und alltägliche Tätigkeiten wie Duschen oder Anziehen auszuführen. Dabei wird sofort deutlich, wie sehr wir uns normalerweise auf visuelle Informationen verlassen, um uns in unserer Umwelt zurechtzufinden. Dennoch können zahlreiche Handlungen auch ohne visuelle Unterstützung durchaus effizient ausgeführt werden: So sollte es keine Probleme bereiten, sich die Socken mit geschlossenen Augen anzuziehen oder ein Hemd zuzuknöpfen. Die Somatosensorik ist jedoch im Gegensatz zum visuellen und auditorischen System kein Fernsinn, sondern erfordert einen unmittelbaren Kontakt des Organismus mit dem jeweiligen Reiz. Dieser Kontakt setzt in der Regel eine aktive Handlung vonseiten des Organismus voraus.
1 Mechanische Sensibilität
Die SensorenFootnote 1 für die Transduktion mechanischer Reize befinden sich in der Haut, die beim Menschen mit einer Oberfläche von etwa 1,8 m2 das größte Sinnesorgan darstellt. Die Haut besteht im Wesentlichen aus drei aufeinanderfolgenden Schichten: (1) einem mehrschichtigen, zum Teil verhornten Plattenepithel, der Epidermis (Oberhaut), (2) der vorwiegend bindegewebigen Dermis (Lederhaut) und (3) der Subkutis (Unterhaut), die lockeres Bindegewebe sowie Fettgewebe enthält. Verschiedene Mechanosensoren befinden sich in allen drei Hautschichten – sie übertragen biologisch relevante Signale mithilfe afferenter Nerven in zentrale Regionen des Gehirns.
1.1 Mechanosensoren der Haut
In die Hautschichten eingebettet liegen vier verschiedene Typen von Mechanosensoren, die aufgrund ihrer strukturellen Eigenschaften unterschiedliche taktile Reize detektieren.Footnote 2 Es handelt sich hierbei um parallel verschaltete Sinneskanäle, die jeweils ihren spezifischen Beitrag zu einer als einheitlich empfundenen Sinneswahrnehmung leisten. Ein Stein, den wir in der Hand halten, besitzt eine charakteristische Oberflächenbeschaffenheit, Temperatur, Form und ein bestimmtes Gewicht – Eigenschaften, die gemeinsam zu einer zusammenhängenden taktilen Wahrnehmung des Steins beitragen. Wie in anderen sensorischen Systemen auch, werden komplexe Reize dieser Art durch periphere Sensoren in ihre elementaren Eigenschaften zerlegt, in parallelen Kanälen weiterverarbeitet und schließlich in höheren Hirnregionen, v. a. im cerebralen Cortex, auf bisher unbekannte Weise wieder zu einer kohärenten Wahrnehmung verknüpft.
Die funktionell unterschiedlichen Mechanosensoren werden durch drei grundlegende Eigenschaften beschrieben:
-
1.
Jeder Sensor antwortet am besten auf einen bestimmten Stimulus wie etwa Druck, Berührung oder Vibration. Die Spezifität ist jedoch nicht absolut – ein druckempfindlicher Sensor reagiert auch auf einen Vibrationsreiz, allerdings mit geringerer Empfindlichkeit.
-
2.
Die Sensoren werden nur dann aktiviert, wenn der Reiz innerhalb ihres rezeptiven Feldes auftritt. Hierunter verstehen wir das von jeweils einem Sensor versorgte Hautareal. Je höher die Sensordichte, desto kleiner ist das rezeptive Feld und desto höher die räumliche Auflösung der jeweiligen Sinnesqualität.
-
3.
Die Sensoren adaptieren mit unterschiedlicher Geschwindigkeit auf einen anhaltenden Stimulus. Schnell adaptierende Mechanosensoren (FA, Fast Adapting) reagieren nur auf den Beginn und das Ende eines Reizes – sie registrieren also vor allem Veränderungen. Langsam adaptierende Mechanosensoren (SA, Slow Adapting) bleiben während der gesamten Dauer des Stimulus aktiv, auch wenn die Aktionspotenzialfrequenz nach einem anfänglichen Maximum etwas zurückgeht. Dieser Sensortyp codiert die Reizgröße unabhängig von der Geschwindigkeit, mit der sie sich ändert.
Die im Folgenden beschriebenen Mechanosensoren bestehen aus einer kapselartigen Struktur, in die afferente Nervenendigungen eingebettet sind (Abb. 15.1). Die Kapsel dient in der Regel als eine Art Filter, der nur bestimmte Eigenschaften eines mechanischen Reizes auf die Nervenfaser überträgt. Die Signaltransduktion erfolgt in den Nervenendigungen, in deren Zellmembranen mechanosensitive Ionenkanäle auf einen Reiz hin öffnen und eine Depolarisation, das Rezeptorpotenzial, auslösen. Größe und Dauer des Rezeptorpotenzials bestimmen gemeinsam mit den Eigenschaften der afferenten Fasern die Aktionspotenzialfrequenz in den sensorischen Neuronen. Die sensorischen Afferenzen gehören zum Typ Aβ (Tab. Tab. 10.2).

Mechanosensoren in der Haut. Der schematische Querschnitt durch die haarlose menschliche Haut zeigt die Epidermis, die Dermis und den oberen Teil der Subkutis. Mechanosensoren befinden sich in allen drei Schichten
Merkel-Endigungen (Merkel-Zell-Axon-Komplexe) finden sich einzeln oder in Gruppen in der Epidermis und kommen an berührungsempfindlichen Stellen wie den Fingerspitzen besonders häufig vor. Sie bestehen aus 50 bis 70 umgewandelten Epithelzellen, den sogenannten Merkel-Zellen, die von einer afferenten Nervenendigung innerviert werden. Die Frage, wo die Signaltransduktion stattfindet, ist noch nicht abschließend geklärt. Merkel-Zellen sind zwar erregbare Zellen, die spannungsabhängige Ca\({}^{2+}\)-Kanäle und die für eine synaptische Freisetzung benötigten Proteine exprimieren (Abschn. 11.4.3). Möglicherweise findet jedoch die Signaltransduktion erst mithilfe der Mechanosensoren der afferenten Nervenfasern statt und die Aktivität der Merkel-Zellen dient lediglich der Modulation der postsynaptischen Aktionspotenzialfrequenz. Aktuelle Befunde deuten auf eine Arbeitsteilung innerhalb des Merkel-Zell-Axon-Komplexes hin: Die epithelialen Merkel-Zellen signalisieren statische Reize, wie einen anhaltenden Druck, während die afferenten Endigungen dynamische Stimuli – etwa einen sich über die Haut bewegenden Gegenstand – verarbeiten 9 ; 14 .
Merkel-Endigungen besitzen mit 0,5 mm die höchste räumliche Auflösung im Vergleich zu allen anderen in der Haut vorhandenen Mechanosensoren.Footnote 3 Sie werden am besten durch senkrecht auf die Haut einwirkende oder sich langsam bewegende taktile Reize aktiviert. So kann beispielsweise das raumzeitliche Codierungsmuster der Brailleschrift aufgrund der Aktivität von Merkel-Endigungen dechiffriert werden. Die afferenten Nervenfasern sind langsam adaptierend. Das Aktivierungsmuster der Merkel-Endigungen bildet die Oberflächenbeschaffenheit von Gegenständen mit hoher räumlicher Präzision ab.
Ruffini-Körperchen sind spindelförmige Strukturen in der Dermis, deren Längsachse annähernd parallel zur Hautoberfläche verläuft.Footnote 4 Sie bestehen aus einer zellulären Kapsel, in die kollagene Fasern eingelagert sind. Diese Faserbündel werden von einer sensorischen Nervenendigung umwickelt, die sich vermutlich bei einer mechanischen Beanspruchung verformt.
Funktionell handelt es sich bei Ruffini-Körperchen um langsam adaptierende Rezeptoren, die sich von den Merkel-Zell-Axon-Komplexen durch eine geringere Empfindlichkeit und eine noch langsamere Adaptation unterscheiden. Aufgrund ihrer Struktur und Lage werden sie vor allem durch eine tangentiale Dehnung der Haut aktiviert, wie sie beispielsweise beim Spreizen der Finger zum Ergreifen einer Kaffeetasse auftritt. Ruffini-Körperchen können anhand des Dehnungsmusters der Haut die Richtung von Objektbewegungen registrieren und sie vermitteln Informationen bezüglich der Position von Hand und Fingern.
Meißner-Körperchen kommen in den Papillarleisten der unbehaarten Haut vor.Footnote 5 Ihre Kapsel besteht aus mehreren lamellenartig angeordneten Schwann-Zellen (Abschn. 9.1.5) und ein bis zwei PerineuralzellenFootnote 6 , zwischen denen zwei bis sechs afferente Nervenfasern enden. Die Kapsel besitzt eine eher längliche Struktur, wobei ihre Längsachse weitgehend senkrecht zur Hautoberfläche orientiert ist. Dieser Typ von Mechanosensoren reagiert deutlich empfindlicher auf Druckänderungen als Merkel-Zellen; die im Vergleich größeren rezeptiven Felder sind jedoch für eine geringere räumliche Auflösung verantwortlich.
Meißner-Körperchen sprechen am besten auf Vibrationen im tieferen Frequenzbereich von 5 bis 50 Hz an, die charakteristisch für über die Hautoberfläche gleitende Objekte sind. Die Afferenzen adaptieren schnell und bilden die Geschwindigkeit der Bewegung mit einer entsprechenden Aktionspotenzialfrequenz ab. Wenn etwa der Kaffeebecher langsam aus der Hand zu rutschen droht, wird diese Objektbewegung relativ zur Haut von diesen Mechanosensoren registriert. Meißner-Körperchen reagieren demnach vor allem auf niederfrequente dynamische Deformationen der Haut und sind für die Regulation der Griffkraft von großer Bedeutung.
Pacini-Körperchen liegen tief im subkutanen Gewebe. Bei diesen Mechanosensoren wird eine sensorische Nervenfaser zwiebelartig durch zahlreiche Schichten von Schwann-Zellen und Perineuralzellen umhüllt. Bedingt durch diese kapselartige Struktur werden bevorzugt Vibrationen im Bereich von 50 bis 700 Hz registriert. Solche Vibrationen entstehen, wenn ein Objekt erstmalig die Hautoberfläche berührt, beispielsweise beim Landen einer Mücke auf dem Arm. Außerdem werden Vibrationen von Objekten, die in der Hand gehalten werden und andere Objekte berühren, übertragen – etwa beim Gebrauch von Werkzeugen und beim Schreiben.
Trotz ihrer Lage in den tieferen Gewebeschichten sind die Pacini-Körperchen die empfindlichsten Mechanosensoren der Haut. Sie können bereits auf eine mechanische Verformung von 10 nm mit einer Erhöhung ihrer Aktionspotenzialfrequenz reagieren. Ihre sensorischen Afferenzen sind schnell adaptierend und besitzen große rezeptive Felder, die eine hohe räumliche Auflösung ausschließen. Pacini-Körperchen vermitteln also vor allem höherfrequente Vibrationsreize, die von anderen Objekten auf die Haut übertragen werden.
In der behaarten Haut übernehmen Haarfollikelrezeptoren , die auf eine Auslenkung der Haare reagieren, die Funktion der dort fehlenden Meißner-Körperchen.
Tab. 15.1 fasst die wichtigsten Eigenschaften der Mechanosensoren der Haut zusammen.
1.2 Mechanismen der Signaltransduktion
Die Signaltransduktion erfolgt in den Endigungen sensorischer Nervenfasern. Hierbei handelt es sich um die peripheren Fortsätze pseudounipolarer Neurone, deren Zellkörper entweder in den Trigeminusganglien Footnote 7 im Hirnstamm oder in den parallel zum Rückenmark angeordneten Spinalganglien liegen. Die sensorischen Neurone der Trigeminusganglien versorgen den Kopfbereich, während die somatosensorische Innervation des übrigen Körpers von den Spinalganglien ausgeht.
In einer sensorischen Endigung befinden sich mechanosensitive Ionenkanäle, die als sogenannte Mechanotransducer einen mechanischen Reiz in den Einstrom von Kationen umwandeln. Na\({}^{+}\)- und Ca\({}^{2+}\)-Ionen diffundieren entlang ihres elektrochemischen Gradienten in die Nervenendigung und erzeugen dort eine schnelle Depolarisation, das Rezeptorpotenzial (Abb. 15.2). Bei einem Rezeptorpotenzial handelt es sich um eine vorübergehende Depolarisation der Membran, deren Amplitude innerhalb bestimmter Grenzen die Reizstärke codiert. Da die Entfernung von der sensorischen Endigung in der Körperperipherie bis zum zugehörigen Soma zu groß für eine passive Weiterleitung ist, werden Aktionspotenziale zur Signalübertragung erzeugt. Diese Aufgabe übernimmt eine unmittelbar auf die sensorische Endigung folgende Region, die sich durch eine hohe Dichte spannungsabhängiger Na\({}^{+}\)- und K\({}^{+}\)-Kanäle auszeichnet. Die Spannungsänderungen werden entlang myelinisierter Aβ-Fasern mit einer Geschwindigkeit von 30 bis 70 m s−1 in Richtung höherer Verarbeitungszentren weitergeleitet.

Aufbau und Eigenschaften sensorischer Nervenendigungen. Eine mechanische Kraft wird durch mechanosensitive Ionenkanäle in ein depolarisierendes Rezeptorpotenzial umgewandelt. In der aktionspotenzialerzeugenden Region des Axons befinden sich Na\({}^{+}\)- und K\({}^{+}\)-Kanäle in hoher Dichte, die das Rezeptorpotenzial in eine Sequenz von Aktionspotenzialen transformieren. Die Aktionspotenziale, deren Frequenz die Reizstärke codiert, werden als afferentes Signal entlang der myelinisierten Aβ-Faser ins Zentralnervensystem weitergeleitet
Mechanosensitive Ionenkanäle sind Transmembranproteine, die auf mechanische Reize wie Zug- oder Druckspannungen mit der Öffnung einer zentralen, ionenpermeablen Pore reagieren. Als Beispiele für eine mechanoelektrische Transduktion haben wir die Prozesse in den Haarzellen des Innenohrs und Vestibularorgans kennengelernt, wo Tip-Links die physikalische Kraft auf K\({}^{+}\)-Kanäle übertragen (Abschn. 14.3.2). Im Falle der Mechanosensoren der Haut wurden bisher weder die Ionenkanäle selbst zweifelsfrei identifiziert, noch sind die molekularen Mechanismen der Transduktion im Detail bekannt.Footnote 8 Sehr wahrscheinlich werden mechanische Kräfte mithilfe elastischer Strukturen, die mit Membranlipiden und/oder cytoskelettalen Elementen verbunden sind, auf einen Ionenkanal übertragen (Abb. 15.3).

Mechanismus der Mechanotransduktion. Eine mechanische Kraft wird durch ein elastisches Element auf einen Ionenkanal übertragen. a Liegt keine Krafteinwirkung vor, ist das elastische Element entspannt und der Ionenkanal geschlossen. b Eine mechanische Kraft spannt das elastische Element an, wodurch Domänen des
Ionenkanals auseinandergezogen werden, sodass der Kanal öffnet. c Bei andauernder Krafteinwirkung adaptiert der Kanal, indem er in eine geschlossene Konformation zurückkehrt. d Die Inaktivierung stellt eine Alternative zur Adaptation dar. In diesem Fall wird die Kanalpore durch ein Inaktivierungspartikel blockiert (nach 3 . Mit freundlicher Genehmigung von Nature Publishing Group.)
Bedingt durch die Art des jeweils exprimierten Mechanotransducers können biologisch relevante Parameter eines mechanischen Reizes codiert werden. Die in Abschn. 15.1.1 beschriebenen Mechanosensoren unterscheiden sich physiologisch in ihrer Adaptationsgeschwindigkeit: Merkel-Zellen beispielsweise sind langsam adaptierend, Meißner-Körperchen hingegen adaptieren schnell. Ein schnell adaptierender Mechanosensor reagiert auf eine abrupte Verformung mit einer Salve von Aktionspotenzialen, wohingegen eine langsame Verformung oder eine dauerhaft einwirkende Kraft keine oder nur wenige Aktionspotenziale auslöst. Im Gegensatz dazu führen beide Reizarten bei einem langsam adaptierenden Mechanosensor zu einer andauernden Aktivität (Abb. 15.4). Aufgrund ihrer Adaptationsrate fungieren schnell adaptierende Mechanosensoren als Geschwindigkeitsdetektoren, die dynamische Reizänderungen codieren, während langsam adaptierende Mechanosensoren eher statische Eigenschaften eines Reizes repräsentieren. Diese unterschiedlichen Antworteigenschaften lassen sich auf ein individuelles Expressionsmuster mechanosensitiver Ionenkanäle in den Nervenendigungen zurückführen.

Schnell und langsam adaptierende Mechanosensoren. a Schnell adaptierende Sensoren reagieren nur auf eine plötzliche Änderung eines Reizes, während eine langsame Steigerung der Reizstärke keine Aktionspotenziale in der afferenten Nervenfaser auslöst. b Langsam adaptierende Mechanosensoren reagieren auf beide Änderungen der Reizstärke mit einer Folge von Aktionspotenzialen. In den jeweils oberen Kästen ist die Änderung der Reizstärke dargestellt, darunter das entsprechende afferente Signal
1.3 Signalbahnen ins Gehirn
Sensorische Signale treten über die dorsalen Wurzeln der Spinalnerven ins Rückenmark ein und ziehen auf derselben Körperseite in den Hinterstrangbahnen bis zu den somatosensorischen Kerngebieten der Medulla oblongata. Die Verschaltung auf das zweite Neuron erfolgt also erst in den Hinterstrangkernen des Hirnstamms, die sich in der hinteren Wand der Medulla oblongata befinden. Der Nucleus cuneatus verarbeitet sensorische Signale aus dem Hand-, Arm- und Halsbereich, der Nucleus gracilis entsprechende Afferenzen aus dem Rumpf und den unteren Extremitäten.
Die Hinterstrangbahnen selbst weisen eine somatotope Gliederung auf (Abb. 15.5). Die Bahnen verlaufen von unten nach oben und neu hinzutretende Axone gliedern sich jeweils seitlich an bereits bestehende Bahnen an. Entsprechend finden sich die unteren Körperregionen im mittleren (medialen) Bereich der Hinterstrangbahnen, während die Fasern aus dem Halsbereich am weitesten seitlich (lateral) angelagert werden. Daher können sensorische Neurone, die die untere Körperhälfte innervieren, fast die gesamte Länge des Organismus durchspannen – etwa von der Fußunterseite bis ins verlängerte Mark des Hirnstamms.

Somatotope Gliederung der Hinterstrangbahn. Afferente Nervenfasern aus den unteren Körperregionen verlaufen im medialen Bereich der Hinterstrangbahn. Fasern aus dem Sakralbereich (S) und dem Lumbalbereich (L) liegen im Inneren der Hinterstrangbahn, an die sich lateral Fasern aus dem Thorakalbereich (Th) und ganz außen diejenigen aus dem Zervikalbereich (C) anschließen. Entsprechend ziehen die afferenten Fasern aus Sakral-, Lumbal- und unteren Thorakalregionen im Fasciculus gracilis zum Nucleus gracilis, während die restlichen Fasern als Fasciculus cuneatus zum Nucleus cuneatus projizieren. Die Vorderseitenstrangbahn übermittelt Signale von Thermosensoren und Nozizeptoren an das Gehirn
Eine Schädigung der Hinterstrangbahnen auf einer Seite führt zu einem Verlust des Berührungsempfindens auf derselben Seite; gleichzeitig sind das Vibrationsempfinden und die Propriozeption beeinträchtigt. Da sensorische Informationen über die Gelenkstellung und die Muskelspannung fehlen, weisen die betreffenden Patienten einen unsicheren Gang auf und können sich nur mit geöffneten Augen im Raum orientieren. Eine Störung der somatosensorischen Verarbeitung reicht also weit über das bloße Fehlen der Berührungsempfindlichkeit hinaus.
Die Fasern der zweiten Neurone in den Hinterstrangkernen kreuzen auf die Gegenseite und ziehen als Lemniscus medialis zum Thalamus im Zwischenhirn. Daher werden mechanische Reize auf der rechten Körperseite von der linken Gehirnhälfte verarbeitet und umgekehrt. Auch im Lemniscus medialis bleibt die somatotope Organisation der Projektion aus den verschiedenen Körperregionen erhalten.
Im Nucleus ventralis posterior des Thalamus erfolgt die Umschaltung auf das dritte Neuron. Vom Thalamus zieht ein Faserbündel, die Capsula interna , zum somatosensorischen Cortex (SI), der im Gyrus postcentralis des Scheitellappens lokalisiert ist. Hier und in weiteren somatosensorischen kortikalen Arealen findet die bewusste Wahrnehmung mechanischer Reize statt.
Sensorische Signale aus der Kopfregion werden zunächst in den Trigeminuskernen umgeschaltet. Die Fasern der zweiten Neurone lagern sich als Lemniscus trigeminalis dem Lemniscus medialis an und erreichen den Thalamus, wo sie von unterschiedlichen Regionen innerhalb des Nucleus ventralis posterior verarbeitet und an den somatosensorischen Cortex weitergeleitet werden.
Die Verarbeitung taktiler Reize im primären somatosensorischen Cortex SI beruht auf folgenden allgemeinen Eigenschaften:
-
SI besteht aus vier verschiedenen Regionen, von denen jede eine vollständige Repräsentation des Körpers enthält. Innerhalb dieser kortikalen Bereiche verarbeiten die Nervenzellen jeweils unterschiedliche mechanische Reize, wie beispielsweise eine Aktivierung der Mechanosensoren der Haut oder der Propriozeptoren der Skelettmuskulatur.
-
Bestimmte Körperregionen wie Hände (insbesondere Fingerspitzen) und Kopf (vor allem Lippen und Zunge) sind beim Menschen im SI deutlich überrepräsentiert. Die außerordentliche Komplexität von Fingerbewegungen sowie von Mimik und Sprache erfordert entsprechende Verschaltungen und ausgedehnte Verarbeitungsareale. Bei Ratten hingegen sind vor allem diejenigen kortikalen Bereiche, in denen Signale aus den Vibrissen des Gesichts verarbeitet werden, relativ zu den anderen Kopfregionen vergrößert. Die sensorische Bedeutung einer Körperregion korreliert also mit der Größe des kortikalen Areals, das Signale aus dieser Region verarbeitet.
-
Die kortikale Repräsentation der Hand liegt in unmittelbarer Nähe derjenigen Areale, die das Gesicht abbilden. Fehlt der sensorische Eingang aus der Hand etwa aufgrund einer Amputation, können sich die Gesichtsareale in die vormaligen Handbereiche ausdehnen. Da Informationen über diese Umstrukturierung kortikaler Karten nicht an höhere Hirnregionen weitergegeben werden, wird häufig eine Berührung im Gesicht als eine Reizung der fehlenden Hand wahrgenommen. Grundsätzlich führt die neuronale Plastizität kontinuierlich zu aktivitätsabhängigen Veränderungen der Verschaltung auch im erwachsenen Gehirn.
Plastische Phänomene können auch unterschiedliche Sinnessysteme einschließen. So führt das Verbinden der Augen bei freiwilligen Versuchspersonen innerhalb weniger Tage zu einer Verarbeitung somatosensorischer Informationen mithilfe des visuellen Cortex. Das Lesen der Brailleschrift wird also in das visuelle System, das keinen sensorischen Eingang über die Retina mehr bekommt, „ausgelagert“ 10 .
1.4 Zusammenfassung
Mechanosensoren wandeln mechanische Reize wie Druck, Spannungen und Vibrationen in elektrische Signale um. Jeder Sensor besitzt eine hohe, jedoch keine absolute Empfindlichkeit für einen bestimmten Stimulus. Die Größe des rezeptiven Feldes bestimmt die Genauigkeit der räumlichen Auflösung (Zwei-Punkt-Schwelle). Schnell adaptierende Mechanosensoren registrieren Veränderungen eines Signals und fungieren als Geschwindigkeitsdetektoren, während langsam adaptierende Sensoren die Reizintensität über den gesamten Zeitraum des Stimulus codieren. In der Regel besitzen Mechanosensoren eine kapselartige Struktur, die aus Proteinen und/oder Zellen des Bindegewebes besteht. Diese Kapsel fungiert wie ein Filter, der nur bestimmte Frequenzen eines Signals passieren lässt.
Merkel-Endigungen kommen vor allem an berührungsempfindlichen Stellen der Epidermis vor. Aufgrund ihrer hohen räumlichen Auflösung sind sie für eine Analyse der Oberflächenbeschaffenheit von Gegenständen besonders geeignet. Ruffini-Körperchen befinden sich in der Dermis der Haut, wo sie – parallel zur Hautoberfläche angeordnet – Dehnungsreize verarbeiten, die Informationen bezüglich der Position von Hand und Fingern beinhalten. Meißner-Körperchen in der unbehaarten Haut reagieren auf Vibrationen mit tiefen Frequenzen, wodurch eine präzise Regulation der Griffkraft ermöglicht wird. Die tief im subkutanen Gewebe liegenden Pacini-Körperchen werden durch höherfrequente Vibrationen aktiviert, die insbesondere über ein in der Hand gehaltenes Objekt übertragen werden.
Pseudounipolare Neurone in den Spinalganglien und in den Trigeminuskernen senden afferente Axone in die Peripherie, wo sie die Kapselstrukturen in den verschiedenen Hautschichten kontaktieren und gemeinsam Mechanosensoren bilden. In den Endigungen der afferenten Fasern befinden sich Ionenkanäle, die auf einen mechanischen Reiz hin öffnen und durch den Einstrom von Kationen eine Depolarisation des Membranpotenzials verursachen. Dieses Rezeptorpotenzial bildet in Amplitude und Dauer den auslösenden Reiz ab. Ist das Rezeptorpotenzial überschwellig, werden Aktionspotenziale erzeugt, die entlang der myelinisierten Aβ-Fasern in Richtung Zentralnervensystem weitergeleitet werden. Die Adaptationsgeschwindigkeit der Mechanosensoren selbst bestimmt, ob eine afferente Faser zur schnell oder langsam adaptierenden Kategorie gehört.
Die somatosensorische Signalbahn aus der Körperperipherie ins Gehirn besteht aus insgesamt drei hintereinander geschalteten Neuronen. Die zentralen Axone der somatosensorischen Neurone in den Spinalganglien ziehen in der Hinterstrangbahn des Rückenmarks ipsilateral bis in die Hinterstrangkerne der Medulla oblongata. Von dort kreuzen Fortsätze des zweiten Neurons auf die Gegenseite und bilden den Lemniscus medialis, der auf das dritte Neuron in den somatosensorischen Kerngebieten des Thalamus verschaltet. Axone thalamischer Neurone verlaufen in der Capsula interna zum primären somatosensorischen Cortex und weiter zu übergeordneten Arealen, in denen die bewusste Wahrnehmung taktiler Reize erfolgt.
Die Signalbahn zeigt insgesamt eine somatotope Repräsentation des Körpers, die sich im somatosensorischen Cortex in Form von vier vollständigen Karten der gesamten Körperoberfläche manifestiert. Mechanosensorisch wichtige Bereiche sind im Verhältnis zu ihrer tatsächlichen Größe im somatosensorischen Cortex überproportional repräsentiert. Die neuronale Plastizität führt zu einer ständigen Umstrukturierung der kortikalen Karten in Abhängigkeit von der Aktivität der sensorischen Kanäle.
2 Thermosensibilität
Die Temperatur spielt für biologische Prozesse eine außerordentlich wichtige Rolle (Abschn. 3.1) und die Fähigkeit, auf Änderungen der Umgebungstemperatur angemessen zu reagieren, ist für das Überleben eines Organismus essenziell. Daher stellt die Sensibilität für Temperaturreize eine der ältesten Sinnesleistungen dar, die von Bakterien bis hin zu Wirbeltieren in allen Lebewesen vorkommt. Bei Säugern verursachen Temperaturänderungen unbewusste physiologische Reaktionen und/oder bewusste Verhaltensanpassungen, die letztlich der Konstanthaltung der Körpertemperatur dienen.
2.1 Thermosensoren
Thermosensoren sind freie Nervenendigungen in der Epidermis der Haut; sie besitzen im Gegensatz zu den Mechanosensoren keine umhüllenden Strukturen (Abb. 15.1). Bei den zugehörigen Sinneszellen handelt es sich um pseudounipolare Neurone, deren Zellkörper in den Spinalganglien liegen und von dort zentrale Axone ins Rückenmark senden. Diese Bahnen leiten Signale über Temperaturänderungen im Bereich des Rumpfes ins Zentralnervensystem. Sie unterscheiden sich von der mechanosensorischen Signalbahn und werden zusammen mit der Schmerzbahn in Abschn. 15.3.3 besprochen. Der Versorgung des Kopfes dienen pseudounipolare Neurone im rechten und linken Trigeminalganglion, deren zentrale Axone in einem der Trigeminuskerne (Nucleus spinalis) auf das zweite Neuron umgeschaltet und von dort im Lemniscus medialis in Richtung Cortex weitergeleitet werden.
Bei der Verarbeitung temperaturrelevanter Signale unterscheiden wir insgesamt drei getrennte Bahnen, von denen jede eine wichtige physiologische Funktion vermittelt:
-
Rückziehreflex. Thermosensorische Neurone in den Spinalganglien verschalten auf Interneurone im Rückenmark, die wiederum Motorneurone im gleichen oder in benachbarten Rückenmarksegmenten aktivieren.Footnote 9 Die Motorneurone bewirken eine Muskelkontraktion, die zum Wegziehen des betroffenen Körperteils führt. Diese reflektorische Signalbahn wird bei schmerzhaft starken Kälte- und Wärmereizen aktiviert, um eine temperaturbedingte unmittelbare Gewebeschädigung zu vermeiden.
-
Wahrnehmung der Temperatur. Ausgehend von Spinal- und Trigeminalganglien werden die Signale über mehrere aufeinanderfolgende Neurone bis zum somatosensorischen Cortex weitergeleitet (Abschn. 15.1.3). Hier findet die bewusste Wahrnehmung der Temperatur statt, auf deren Grundlage willkürliche Verhaltensänderungen eingeleitet werden können.Footnote 10
-
Unbewusste thermoregulatorische Prozesse. Thermosensorische Informationen werden in die präoptische Region des Hypothalamus übertragen, die ihrerseits temperaturempfindliche Neurone enthält (Abschn. 3.4.1). Die Integration peripherer und zentraler Temperatursignale innerhalb des Hypothalamus löst thermoregulatorische Prozesse wie die Engstellung oder die Erweiterung peripherer Blutgefäße, Muskelzittern sowie die Thermogenese im braunen Fettgewebe aus (Abschn. 3.5).
Wärme- und Kälteempfindungen treten bei endothermen Organismen auf, wenn Abweichungen von der Indifferenztemperatur der Thermoneutralzone vorliegen (Abschn. 3.4.3). Beim Menschen reagieren Kaltsensoren maximal bei einer Temperatur von etwa \(25\,^{\circ}\mathrm{C}\), während Warmsensoren bei rund \(43\,^{\circ}\mathrm{C}\) ihre höchste Aktivität zeigen (Abb. 15.6). Temperaturen unterhalb von etwa \(20\,^{\circ}\mathrm{C}\) und oberhalb von ca. \(45\,^{\circ}\mathrm{C}\) aktivieren Nozizeptoren, deren Signale als thermische Schmerzreize ins Bewusstsein gelangen.

Aktivität von Kalt- und Warmsensoren. a Eine Erwärmung der Hautoberfläche führt zu einer Erhöhung der Aktionspotenzialfrequenz bei Warmsensoren, während die Entladungsrate der Kaltsensoren unverändert bleibt. Bei einer Abkühlung reagieren beide Sensortypen genau umgekehrt. Kalt- und Warmsensoren zeigen während einer Temperaturänderung die höchste Aktivität; eine konstant tiefe bzw. hohe Temperatur bewirkt eine geringfügige Adaptation. b Durchschnittliche Aktivitätsrate von Kalt- und Warmsensoren bei unterschiedlichen Hauttemperaturen. Die Aktivierung von Kaltrezeptoren bei Temperaturen über \(45\,^{\circ}\mathrm{C}\) verursacht eine paradoxe Kälteempfindung
Die Temperaturempfindungen „warm“ bzw. „kalt“ werden unabhängig voneinander in getrennten Kanälen verarbeitet. Kaltsensoren leiten ihre Signale vorwiegend über myelinisierte Aδ-Fasern, Warmsensoren hingegen bestehen hauptsächlich aus nichtmyelinisierten C-Fasern mit geringerer Leitungsgeschwindigkeit (Tab. Tab. 10.2). Warm- und Kaltsensoren besitzen rezeptive Felder auf der Hautoberfläche, deren Größe durch die Innervationsdichte mit freien Nervenendigungen bestimmt wird. Sie treten im Gesicht und hier insbesondere im Mundbereich am häufigsten auf, wohingegen der Rumpf eine relativ geringe Dichte an Thermosensoren aufweist. Grundsätzlich kommen mehr Kaltsensoren als Warmsensoren vor.
Das Temperaturempfinden zeigt eine ausgeprägte Adaptation , die auf einfache Weise durch den Drei-Schalen-Versuch von Weber verdeutlicht wird. Wenn wir die linke Hand in warmes Wasser von \(38\,^{\circ}\mathrm{C}\) und die rechte Hand in kaltes Wasser von \(26\,^{\circ}\mathrm{C}\) tauchen, nehmen wir zunächst einen deutlichen Temperaturunterschied wahr, der jedoch nach einigen Minuten verschwindet. Die Thermosensoren adaptieren an die jeweilige Wassertemperatur, sodass die zentrale Verrechnung keinen oder nur noch einen geringfügigen Unterschied zwischen der rechten und linken Hand anzeigt. Werden anschließend jedoch beide Hände gleichzeitig in \(33\,^{\circ}\mathrm{C}\) warmes Wasser – entsprechend der normalen Hauttemperatur – getaucht, fühlt sich die linke Hand zunächst kühler an als die rechte. Diese Befunde zeigen, dass Thermosensoren vor allem auf steigende oder fallende Temperaturen reagieren, während eine konstante Temperatur aufgrund von Adaptationsprozessen wenig oder keine Aktivitätsänderung hervorruft.
2.2 Thermorezeptoren und Signaltransduktion
Die Thermosensibilität basiert auf den biophysikalischen Eigenschaften von Ionenkanälen, die durch Änderungen der Temperatur geöffnet oder geschlossen werden. Grundsätzlich ist die Funktion aller Proteine von der Temperatur abhängig, was im Falle von Stoffwechselraten quantitativ durch den Q 10-Wert beschrieben wird (Abschn. 3.1.3). Auch für die temperaturabhängige Permeabilität von spannungsabhängigen oder ligandengesteuerten Ionenkanälen lässt sich der Q 10-Wert anwenden. Er beträgt normalerweise 1,2 bis 1,5, was auf einen relativ geringen Einfluss der Temperatur auf das Gating der Kanäle hindeutet. Bei Ionenkanälen hingegen, die spezifisch durch eine Erhöhung der Temperatur geöffnet werden, liegt der Q 10-Wert oberhalb von 5; Kanäle, die durch eine Verringerung der Temperatur aktiviert werden, besitzen einen Q 10-Wert, der kleiner als 0,2 ist. Die Temperaturabhängigkeit ist demnach eine spezifische Eigenschaft thermosensitiver Ionenkanäle und nicht eine allgemeine Folge temperaturinduzierter molekularer Umlagerungsprozesse.
Die Identität der an der Thermosensibilität beteiligten Ionenkanäle ist zu diesem Zeitpunkt noch nicht abschließend geklärt, aber offensichtlich spielen Vertreter der TRP-Kanäle (Transient Receptor Potential) als „molekulare Thermometer“ eine entscheidende Rolle bei der Signaltransduktion. TRP-Kanäle bestehen aus vier Untereinheiten, von denen jede sechs Transmembrandomänen besitzt. Sie ähneln damit sowohl in ihrer Quartärstruktur als auch in ihrer Membrantopologie den spannungsgesteuerten Ionenkanälen (Abb. 15.7a; Abschn. 9.2.1).

Struktur und Temperaturabhängigkeit thermosensitiver Ionenkanäle. a Alle TRP-Kanäle besitzen sechs Transmembransegmente, eine Porenschleife und einen intrazellulären N- und C-Terminus. b Abhängig von ihrem Typ reagieren TRP-Kanäle auf unterschiedliche Temperaturen mit einem Öffnen ihrer Kanalpore, sodass Kationen in die Zelle hineindiffundieren. Der Ionenstrom ist qualitativ aufgetragen
TRP-Kanäle sind Kationenkanäle, denen jedoch der Selektivitätsfilter der spannungsgesteuerten Ionenkanäle fehlt. Daher können sie nicht zwischen Na\({}^{+}\)-, K\({}^{+}\)- und Ca\({}^{2+}\)-Ionen unterscheiden. Das Öffnen der TRP-Kanäle führt aufgrund des Einstroms von Na\({}^{+}\) und Ca\({}^{2+}\) zur Depolarisation der Zellmembran.
Erstmalig wurde TRPV1 als Rezeptor für Capsaicin, einer Substanz, die Chilis ihre Schärfe verleiht, beschrieben. TRPV1 reagiert jedoch nicht nur auf Capsaicin, sondern vor allem auf Temperaturen von mehr als \(43\,^{\circ}\mathrm{C}\) und stellt damit einen wärmespezifischen Schmerzrezeptor (Nozizeptor) dar. Eine schmerzhafte Wärmeempfindung und der durch Chilis hervorgerufene brennende Schmerz werden also durch denselben Rezeptortyp vermittelt. TRPM3 wird wie TRPV1 ebenfalls von Neuronen in den Spinalganglien exprimiert und signalisiert noch höhere Temperaturen (Abb. 15.7b). TRPV1 und TRPM3 sind allerdings Nozizeptoren, die durch Wärmereize unterhalb der Schmerzschwelle nicht aktiviert werden. Die eindeutige Identifizierung nichtnozizeptiver Wärmerezeptoren steht noch aus.Footnote 11
Eine nichtschmerzhafte Kälteempfindung wird sehr wahrscheinlich durch den Ionenkanal TRPM8 vermittelt (Abb. 15.7b). Interessanterweise reagiert TRPM8 auch auf Substanzen, wie beispielsweise Menthol oder Eukalyptusöl, die eine subjektive Kälteempfindung hervorrufen. TRPA1 schließlich zeigt ein im Vergleich zu TRPM8 nach links verschobenes Aktivierungsprofil und ist damit möglicherweise an der schmerzhaften Kälteempfindung beteiligt.Footnote 12
Zahlreiche TRP-Kanäle besitzen neben ihrer thermosensitiven Funktion Eigenschaften ligandengesteuerter Ionenkanäle, indem chemische Substanzen wie Capsaicin oder Menthol mit einer Bindungsstelle auf dem Kanalprotein in Wechselwirkung treten. Capsaicin vermittelt ein brennend scharfes Empfinden, während Menthol als kühlend wahrgenommen wird. Obwohl keine Temperaturänderung vorliegt, aktiviert das Öffnen dieser TRP-Kanäle eine Signalbahn, die letztlich zu einer subjektiven Wärme- bzw. Kälteempfindung führt. Dies wiederum bedeutet, dass die Aktivierung bestimmter sensorischer Bahnen und ihrer Projektionsgebiete im Cortex einen spezifischen, invarianten Sinneseindruck hervorruft.
Tab. 15.2 fasst die Eigenschaften der wichtigsten aktuell bekannten thermosensitiven TRP-Kanäle zusammen. Sehr wahrscheinlich sind neben den bisher beschriebenen TRP-Kanälen noch andere Ionenkanäle, die entweder auch zur TRP-Familie oder aber zu anderen Familien von Ionenkanälen gehören, an der Transduktion thermischer Reize beteiligt.
Eine bislang ungeklärte Frage betrifft den oder die Mechanismen, die einem temperaturabhängigen Gating von Ionenkanälen zugrunde liegen. Gegenwärtig werden die folgenden Möglichkeiten diskutiert:
-
Intrinsische Temperaturabhängigkeit. Der Ionenkanal selbst reagiert auf Temperaturänderungen mit einer Konformationsänderung, wodurch seine Leitfähigkeit erhöht oder verringert wird (Abb. 15.8a). TRPM8 und TRPV1 werden wahrscheinlich mithilfe eines derartigen Mechanismus aktiviert.
Abb. 15.8 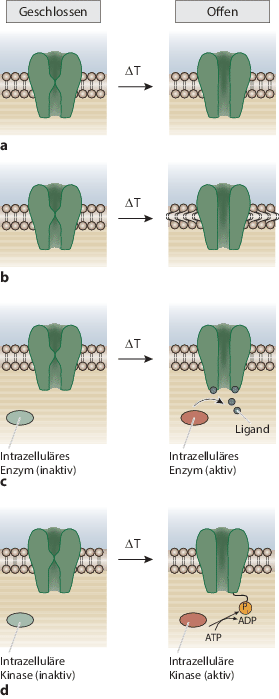
Mögliche Mechanismen der Regulation thermosensitiver Ionenkanäle. a Änderungen der Temperatur (\(\Updelta T\)) führen unmittelbar zu einer Konformationsänderung und damit zu einem Öffnen des Kanals. b Die Temperatur beeinflusst die Eigenschaften der Zellmembran und der Kanal reagiert auf Veränderungen der Dicke, der Spannung oder der Krümmung der Membran. c Ein inaktives intrazelluläres Enzym wird durch eine Temperaturänderung aktiviert und stellt einen Liganden her, dessen Bindung an den Kanal die Kanalpore öffnet. d Eine Änderung der Temperatur aktiviert eine intrazelluläre Kinase, die den Kanal durch eine Phosphorylierungsreaktion öffnet (nach 13 . Mit freundlicher Genehmigung von Nature Publishing Group.)
-
Änderungen der Membraneigenschaften. Lipidmembranen reagieren hochgradig empfindlich auf Temperaturerhöhungen, indem sie einen Phasenübergang von einem quasikristallinen in einen gelartigen Zustand vollziehen (Abschn. 3.1). Durch diesen Prozess können theoretisch mechanische Kräfte auf ein Transmembranprotein übertragen und so ein zentraler Ionenkanal geöffnet werden (Abb. 15.8b). In diesem Modell ist Thermosensitivität eine indirekte Form der Mechanosensitivität des Kanals.
-
Aktivierung eines intrazellulären Liganden. Die katalytische Aktivität eines Enzyms ist meist außerordentlich temperaturabhängig. So kann eine Erhöhung der Temperatur ein Enzym aktivieren, das die Synthese einer intrazellulären Substanz katalysiert, die ihrerseits als Ligand für einen thermosensitiven Ionenkanal fungiert. Die Erhöhung der intrazellulären Konzentration dieses Liganden führt zu einer höheren Offenwahrscheinlichkeit des Ionenkanals. Dieses Modell basiert auf den ligandenbindenden Eigenschaften einiger thermosensitiver Kanäle (Abb. 15.8c).
-
Phosphorylierung. Eine Temperaturänderung kann die katalytische Aktivität einer Kinase wie etwa Proteinkinase A oder Proteinkinase C verändern, was wiederum den Phosphorylierungszustand eines thermosensitiven Ionenkanals beeinflusst. Phosphorylierung und Dephosphorylierung sind bekannte Mechanismen zur Regulation der Permeabilität bei zahlreichen Ionenkanälen (Abb. 15.8d).
-
Rekrutierung von Untereinheiten. Eine Temperaturänderung kann die Konformation und/oder Lokalisation von Untereinheiten beeinflussen, die zum Öffnen des Ionenkanals erforderlich sind.

Der erste Mechanismus, die intrinsische Temperaturabhängigkeit des Ionenkanals, basiert auf der Annahme einer thermosensitiven Region innerhalb des Proteins. Diese Region wird von einer konservierten Aminosäuresequenz gebildet, die mit dem Spannungssensor spannungsabhängiger Ionenkanäle vergleichbar ist (Abschn. 9.2.1). Offensichtlich befinden sich diese thermosensitiven Bereiche jedoch in unterschiedlichen, relativ weit voneinander entfernten Regionen des Proteins, sodass wahrscheinlich zahlreiche submolekulare Konformationsänderungen letztlich für die Temperaturabhängigkeit des Kanals verantwortlich sind.
2.3 Zusammenfassung
Verlässliche Informationen über die Umgebungstemperatur sind für das Überleben eines Organismus von zentraler Bedeutung. Änderungen der Temperatur oder Abweichungen von der Thermoneutralzone werden von Thermosensoren registriert, die als freie Nervenendigung in der Epidermis der Haut liegen. Drei unterschiedliche Signalbahnen vermitteln wichtige physiologische Funktionen der Thermosensitivität: (1) Rückziehreflex zur Vermeidung kälte- oder wärmebedingter Gewebeschädigungen, (2) bewusste Wahrnehmung der Temperatur im somatosensorischen Cortex zur Einleitung von Verhaltensänderungen, (3) unbewusste thermoregulatorische Prozesse im Hypothalamus zur Konstanthaltung der Körpertemperatur.
Kaltsensoren zeigen eine maximale Reaktion bei einer Temperatur von etwa \(25\,^{\circ}\mathrm{C}\), Warmsensoren hingegen bei \(43\,^{\circ}\mathrm{C}\). Unterhalb von \(20\,^{\circ}\mathrm{C}\) und oberhalb von \(45\,^{\circ}\mathrm{C}\) werden zusätzlich temperaturabhängige Schmerzrezeptoren aktiviert. Kalt- und Warmsensoren zeichnen sich durch eine schnelle Adaptation aus: Temperaturänderungen führen zu einer Verstärkung der Aktionspotenzialfrequenz in der afferenten Nervenfaser, während eine konstante Temperatur keine Aktivitätssteigerung auslöst. Die thermosensitiven Nozizeptoren hingegen zeigen eine deutlich geringere Adaptation.
Die Umwandlung thermischer Reize in zelluläre elektrische Signale erfolgt in den freien Nervenendigungen mithilfe spezifischer Ionenkanäle der TRP-Familie. Für die Transduktion schmerzhafter Wärmereize werden TRPV1 und TRPM3 diskutiert, die bei entsprechend hohen Temperaturen einen Kationenkanal öffnen, was zur Depolarisation der Endigung und zur Erhöhung der Aktionspotenzialfrequenz in den afferenten Nervenfasern führt. TRPV1 wird ebenfalls durch Capsaicin aktiviert, wodurch die brennende Schärfe von Chilis hervorgerufen wird.
TRPM8 vermittelt nichtschmerzhafte Kälteempfindungen, während möglicherweise TRPA1 durch nozizeptive Kältereize aktiviert wird. Das temperaturabhängige Gating der Ionenkanäle ist gegenwärtig nicht geklärt. Mögliche Mechanismen reichen von einer intrinsischen Temperaturabhängigkeit der Kanäle über temperaturbedingte Änderungen der Membranfluidität bis hin zu einer Aktivierung intrazellulärer Enzyme, deren katalytische Aktivität ihrerseits von der Temperatur abhängt.
3 Nozizeption
Die meist unangenehme subjektive Wahrnehmung von Schmerzen, die sogenannte Nozizeption , stellt ein außerordentlich wichtiges Warnsignal dar, das den Organismus vor körperlichen Schäden schützt. Die eigentliche Schmerzempfindung entsteht im Gehirn, das die elektrische Aktivität in eng umgrenzten kortikalen Arealen als Schmerz in einer bestimmten Körperregion interpretiert. Nozizeption wird nicht durch eine übermäßige Aktivierung der bereits besprochenen Mechano- und Thermosensoren verursacht, sondern beruht auf getrennten Sensoren (Nozizeptoren) und spezifischen zentralnervösen Bahnen. Eine seltene Mutation in einem spannungsabhängigen Na\({}^{+}\)-Kanal, der vor allem von Nozizeptoren exprimiert wird, verursacht eine gestörte Signalerzeugung und -weiterleitung. Daher gelangen normalerweise schmerzhafte Reize aus der Peripherie nicht ins Gehirn, was den Verlust der Schmerzwahrnehmung beim Menschen zur Folge hat. Die Verarbeitung nichtschmerzhafter thermischer und mechanischer Reize bleibt hingegen unbeeinträchtigt 4 ; 5 . Da die betroffenen Patienten keine Schmerzen spüren, fehlen entscheidende Rückmeldungen über schädigende Prozesse – mit der Folge häufiger Verletzungen und einer signifikant reduzierten Lebenserwartung.
3.1 Nozizeptoren
Nozizeptoren sind spezifische Sensoren in der Haut und in den inneren Organen, die potenziell schädigende thermische, mechanische oder chemische Reize (noxische Reize) in ein graduiertes Rezeptorpotenzial und anschließend in eine Folge von Aktionspotenzialen in den afferenten Nervenfasern umwandeln. Sie liegen als unspezialisierte freie Nervenendigungen in der Epidermis der Haut und in den Organen. Die eigentlichen Sinneszellen sind pseudounipolare Neurone, deren Zellkörper sich entweder in den Spinalganglien oder in den Trigeminusganglien befinden. Meist werden Nozizeptoren hinsichtlich der Leitungsgeschwindigkeit ihrer Axone unterschieden, die entweder zu den myelinisierten Aδ- oder den nichtmyelinisierten C-Fasern gehören (Abb. 15.9).

Erster und zweiter Schmerz. Der als stechend scharf empfundene erste Schmerz wird von myelinisierten Aδ-Fasern weitergeleitet, während nichtmyelinisierte, langsamere C-Fasern den zweiten Schmerz vermitteln. Die Verästelungen der Nervenfasern repräsentieren freie Nervenendigungen in den Geweben des Körpers
-
Hochschwellige Nozizeptoren reagieren auf starke mechanische Reize. Ihre Signale werden schnell mithilfe von Aδ-Fasern ins Zentralnervensystem geleitet, wo sie zur Wahrnehmung eines kurzen, stechend scharfen Schmerzes führen, dem sogenannten ersten Schmerz. Ein derartiger Schmerz tritt beispielsweise bei einem Schnitt in den Finger auf. Die Aktivierung dieser Nozizeptoren löst einen Rückziehreflex als unmittelbare Schutzreaktion aus, der zeitlich vor der bewussten Schmerzwahrnehmung abläuft.
-
Polymodale Nozizeptoren leiten ihre Signale vor allem mittels langsamer C-Fasern weiter. Polymodal bedeutet, dass diese Schmerzsensoren sowohl durch übermäßige Hitze und Kälte als auch durch mechanische und chemische Einflüsse aktiviert werden können. Sie stellen die zahlenmäßig größte Gruppe nozizeptiver Afferenzen und sind für ein lang anhaltendes, dumpfes Schmerzempfinden, den sogenannten zweiten Schmerz, verantwortlich. Dieser Schmerz hat einen brennenden und bohrenden Charakter und tritt meist etwas zeitverzögert nach Verletzungen auf.
-
Schlafende Nozizeptoren sind im gesunden Gewebe inaktiv, können aber durch Entzündungsprozesse aktiviert werden. Dann reagieren sie bereits auf schwache mechanische Reize, die normalerweise kein Schmerzempfinden auslösen. Die Senkung der Reizschwelle führt zu einer Schmerzüberempfindlichkeit, die als primäre Hyperalgesie bezeichnet wird.
Innerhalb der Aδ-Fasern wird noch zwischen zwei Klassen von Sensoren unterschieden: (1) Sensoren, die vor allem auf mechanische und chemische Reize, aber weniger auf schmerzhafte Wärme oder Kälte reagieren, und (2) Sensoren mit einer niedrigen Schwelle für Wärme und einer hohen Schwelle für mechanische/chemische Reize.
3.2 Signaltransduktion
Ionenkanäle der TRP-Familie sind für die Umwandlung schmerzhaft kalter und warmer Reize in ein Rezeptorpotenzial verantwortlich (Abschn. 15.2.2) und sie sind sehr wahrscheinlich auch an der Signaltransduktion schmerzhafter mechanischer und chemischer Reize beteiligt. So wird zum Beispiel TRPA1 durch zahlreiche Chemikalien aktiviert, die alle eine brennende oder prickelnde Schmerzempfindung auslösen. TRPA1 dient möglicherweise auch als Rezeptor für mechanische Reize und kann daher verschiedene noxische Stimuli integrieren.
Wenn bei einer Verletzung Zellen zerstört werden, treten ATP und K\({}^{+}\)-Ionen aus dem geschädigten Bereich in das Interstitium des umliegenden Gewebes aus. Hinzu kommen Entzündungsmediatoren wie Bradykinin Footnote 13 und Prostaglandine , wobei das Prostaglandin PGE2 eine zentrale Rolle spielt. Prostaglandine werden aus Arachidonsäure mithilfe des Enzyms Cyclooxygenase synthetisiert. Die schmerzstillende Wirkung leichter Schmerzmittel wie Acetylsalicylsäure – dem Wirkstoff in Aspirin – und Ibuprofen beruht auf der Hemmung der Cyclooxygenase, wodurch die Synthese von PGE2 verhindert wird.Footnote 14
Die genannten Entzündungsmediatoren üben eine indirekte Wirkung auf TRP-Kanäle aus, indem sie über G-Proteine und intrazelluläre Signalkaskaden die Permeabilität der Ionenkanäle verändern. Bradykinin und PGE2 vermitteln zwei grundlegende intrazelluläre Mechanismen (Abb. 15.10).

Signaltransduktion in einer nozizeptiven Nervenendigung. Entzündungsmediatoren wie Bradykinin und Prostaglandin PGE2 binden an membranständige G-protein-gekoppelte Rezeptoren, wodurch eine intrazelluläre Signalkaskade ausgelöst wird. Proteinkinase A (PKA) und Proteinkinase C (PKC) phosphorylieren einen TRPV1-Ionenkanal, der auf diese Weise geöffnet wird und den Einstrom von Kationen in die Nervenendigung ermöglicht. Das überschwellige Rezeptorpotenzial löst die Erzeugung von Aktionspotenzialen aus, die entlang der Nervenfaser ins Zentralnervensystem weitergeleitet werden
-
Bradykinin bindet an Bradykininrezeptoren in der Membran der freien Nervenendigung, was zu einer G-protein-vermittelten Aktivierung von Phospholipase C (PLC) führt. PLC katalysiert die Bildung von Diacylglycerol, das wiederum Proteinkinase C (PKC) aktiviert. Die Phosphorylierung von TRPV1-Kanälen durch PKC führt zum Einstrom von Kationen (Na\({}^{+}\) und Ca\({}^{2+}\)) und zur Depolarisation des Nozizeptors.
-
Durch die Bindung von PGE2 an einen membranständigen Prostaglandinrezeptor wird das Enzym Adenylatcyclase aktiviert. Adenylatcyclase erhöht die intrazelluläre Konzentration von cAMP, wodurch die Proteinkinase A aktiviert wird, die ihrerseits den TRPV1-Kanal phosphoryliert und auf diese Weise öffnet.
Neben den genannten Substanzen sind noch andere inflammatorisch wirksame Stoffe, darunter zahlreiche Zytokine, an der Aktivierung der Nozizeptoren beteiligt. Sie können Nozizeptoren sensibilisieren und somit zu der erhöhten Schmerzempfindlichkeit beitragen, die häufig entzündliche Prozesse begleitet. Diese periphere Sensibilisierung ist die Grundlage für die primäre Hyperalgesie. In diesem Fall stellt bereits die normale Körpertemperatur einen schmerzhaften Wärmereiz dar, der durch Kühlen der entzündeten Geweberegion reduziert werden kann. Bei der primären Hyperalgesie wird der Schwellenwert eines Nozizeptors wie TRPV1 so weit gesenkt, dass bereits nichtnoxische Temperaturen eine Schmerzempfindung erzeugen.
Bedingt durch die Plastizität synaptischer Verbindungen kann eine kontinuierliche niederfrequente Aktivität von Nozizeptoren in den zweiten Neuronen des Rückenmarks eine überschießende Erregung auslösen, die als Schmerz wahrgenommen wird. Hierbei handelt es sich um eine zentrale Sensibilisierung , die für die sekundäre Hyperalgesie verantwortlich ist. Diese Form der Schmerzüberempfindlichkeit beruht auf zentralnervösen, meist spinalen Mechanismen und reicht in ihrer räumlichen Ausdehnung über den pathologisch veränderten Bereich hinaus. Aufgrund der zentralen Sensibilisierung kommt es bei der sekundären Hyperalgesie auch ohne sensorische Reize zu einer Schmerzempfindung.
In der Regel gehen die Entzündungsprozesse und damit auch die Hyperalgesie nach einer gewissen Zeit zurück und das normale Schmerzempfinden stellt sich wieder ein. Eine dauerhafte Schädigung zentraler Schmerzbahnen, wie sie bei Herpesinfektionen, Diabetes, multipler Sklerose, Schlaganfällen und anderen Erkrankungen auftreten kann, führt hingegen zu chronischen neuropathischen Schmerzen, die außerordentlich schwer zu behandeln sind.
3.3 Aufsteigende und absteigende Bahnen
Die zentralen Fasern der pseudounipolaren Nozizeptoren treten wie alle anderen sensorischen Fasern auch über die dorsalen Wurzeln ins Rückenmark ein. Dort verzweigen sie sich t-förmig und verlaufen über die Länge weniger Rückenmarksegmente nach oben und nach unten, bevor sie auf ein spinales Interneuron (2. Neuron) im dorsalen Horn des Rückenmarks umgeschaltet werden. Die Axone des 2. Neurons kreuzen auf die Gegenseite, wo sie im Vorderseitenstrangsystem in Richtung Gehirn verlaufen (Abb. 15.5). Signale über Temperaturänderungen werden ebenfalls im Vorderseitenstrangsystem weitergeleitet.
Die Umschaltung auf die kontralaterale Seite des Körpers erfolgt also bereits auf der Höhe des Rückenmarks und nicht – wie bei der mechanosensorischen Bahn – erst in der Medulla oblongata (Abschn. 15.1.3 ). Dieser anatomische Unterschied hat wichtige Konsequenzen für die Lokalisation einer unvollständigen Läsion des Rückenmarks, die mechanosensorische und nozizeptive Bahnen durchtrennt. Wird beispielsweise bei einer Halbseitenläsion das Rückenmark nur auf der rechten Seite beschädigt, fallen Mechanosensibilität und Propriozeption unterhalb der Läsionsstelle auf derselben Seite aus, während die Schmerz- und Temperaturempfindung auf der gegenüberliegenden Seite gestört sind. Diese dissoziierte Empfindungsstörung wird auch als Brown-Séquard-Syndrom bezeichnet.
Die Hauptbahn des Vorderseitenstrangs (Tractus spinothalamicus ) schließt sich dem Lemniscus medialis an und zieht zu verschiedenen Kernen innerhalb des Thalamus und anschließend in den somatosensorischen Cortex, wo die bewusste Bewertung des Schmerzes hinsichtlich seiner Lokalisation, Intensität und Qualität erfolgt. Ein kleinerer Teil des Vorderseitenstrangs, der Tractus spinoreticularis , projiziert in die Formatio reticularis des Hirnstamms und wird dort auf andere Thalamuskerne sowie den Hypothalamus verschaltet. Dieses Projektionssystem löst vegetative und emotionale Reaktionen auf einen Schmerzreiz aus. Emotionale Komponenten der Schmerzwahrnehmung umfassen die unangenehmen und beängstigenden Empfindungen, die meist mit Schmerzen verbunden sind. Projektionen in die Formatio reticularis des Hirnstamms und vor allem in limbische Areale sind demnach für die bewusste affektive Bewertung von Schmerzreizen verantwortlich.Footnote 15
Zahlreiche Beobachtungen deuten darauf hin, dass die subjektive Schmerzempfindung hochgradig variabel ist und sehr stark von den jeweiligen Umständen abhängt. So berichteten etwa während des Zweiten Weltkriegs auf dem Schlachtfeld schwer verletzte Soldaten, dass sie kaum oder überhaupt keine Schmerzen empfanden. Gleichzeitig unterliegt die Schmerzwahrnehmung einem gut dokumentierten Placeboeffekt Footnote 16 : Beispielsweise konnte in einer Studie bei 75 % der Patienten postoperativer Schmerz durch Injektion einer Kochsalzlösung zufriedenstellend behandelt werden. Diese Befunde legen nahe, dass die subjektive Schmerzwahrnehmung durch Signale aus dem Gehirn, die über absteigende Bahnen ins Rückenmark gelangen, nachhaltig verändert werden kann.
Diese absteigenden Bahnen entspringen in unterschiedlichen Hirnregionen und beeinflussen letztlich die Projektionsneurone im dorsalen Horn des Rückenmarks – die 2. Neurone der Schmerzbahn. Neben inhibitorischen Interneuronen im Rückenmark, die Glycin als Neurotransmitter verwenden, existieren lokale Interneurone, die endogene Opioide synthetisieren und an den Synapsen zwischen nozizeptiven Afferenzen und den Projektionsneuronen freisetzen (Abb. 15.11). Bedingt durch den hemmenden Einfluss der Opioide werden die Projektionsneurone der Schmerzbahn weniger stark oder gar nicht aktiviert, sodass die bewusste Schmerzempfindung entsprechend schwächer ausgeprägt ist.

Modulation der Schmerzbahn durch opioidfreisetzende Interneurone. Der Kasten zeigt die synaptische Verschaltung im dorsalen Horn des Rückenmarks. C-Fasern leiten nozizeptive Signale über die dorsalen Wurzeln ins Rückenmark, wo sie exzitatorische Synapsen mit Projektionsneuronen ausbilden. Die Axone der Projektionsneurone verlaufen in den Hinterstrangbahnen zu den somatosensorischen Kerngebieten in der Medulla oblongata. Absteigende Bahnen aus Hirnstammkernen aktivieren Enkephalin enthaltende Interneurone, die ihrerseits mit inhibitorischen Synapsen auf die Axonterminalien der C-Fasern verschalten. Die Übertragung nozizeptiver Signale von den C-Fasern auf die Projektionsneurone wird aufgrund dieser synaptischen Hemmung abgeschwächt. \(\oplus\) bezeichnet exzitatorische Synapsen, \(\ominus\) inhibitorische Synapsen
Derivate des Opiums, wie beispielsweise Morphium, werden therapeutisch als starkes Schmerzmittel (Analgetikum) eingesetzt. Morphium kann seine analgetische Wirkung nur entfalten, wenn es an einen passenden Rezeptor bindet. Da es jedoch höchst unwahrscheinlich ist, dass sich ein solcher Rezeptor evolutionär aufgrund einer externen Verabreichung von Opiaten entwickelt hat, liegt die Vermutung nahe, dass es körpereigene Substanzen geben muss, die an die entsprechenden Rezeptoren binden. Inzwischen sind mit den Enkephalinen , Endorphinen und Dynorphinen drei unterschiedliche Gruppen endogener Opioide identifiziert worden. Bei diesen Opioiden handelt es sich um kurze Neuropeptide, die an G-protein-gekoppelte Opioidrezeptoren binden und intrazellulär eine Hemmung der neuronalen Aktivität in den postsynaptischen Neuronen der Schmerzbahn verursachen.
3.4 Natriumkanäle und Schmerz
Das in den freien Nervenendigungen erzeugte Rezeptorpotenzial wird – wenn es überschwellig ist – in eine Folge von Aktionspotenzialen umgewandelt, deren Frequenz die Stärke des Schmerzreizes codiert. Aktionspotenziale werden von spannungsabhängigen Na\({}^{+}\)-Kanälen generiert, von denen jeweils bestimmte Isoformen in den nozizeptiven Endigungen vorkommen.Footnote 17
Mehrere Punktmutationen im Gen SCN9A, das die \(\mathrm{Na_{v}}1.7\)-Isoform codiert, verursachen das Krankheitsbild der erblichen Erythromelalgie (EEM) . Symptome der Erkrankung umfassen starke brennende Schmerzen in Händen und Füßen, verbunden mit einer Schwellung, Rötung und Überhitzung. Andere Punktmutationen in demselben Gen sind für die extreme paroxysmale Schmerzstörung (Paroxysmal Extreme Pain Disorder, PEPD) verantwortlich.Footnote 18 Im Gegensatz zu diesen funktionssteigernden Mutationen führt der vollständige Funktionsverlust von \(\mathrm{Na_{v}}1.7\)-Kanälen bedingt durch eine homozygote Nonsense-Mutation zu einer Form der Schmerzunempfindlichkeit, die als kongenitale Schmerzinsensitivität bezeichnet wird 4 . Andere sensorische Leistungen – insbesondere mechano- und thermosensorische Wahrnehmungen – sind mit einer Ausnahme nicht beeinträchtigt: Da \(\mathrm{Na_{v}}1.7\) auch an der olfaktorischen Signalleitung maßgeblich beteiligt ist, verursacht ein Funktionsverlust dieses Kanals Störungen bei der Geruchswahrnehmung bis hin zur Anosmie. Insgesamt deuten diese Befunde auf eine zentrale Rolle von \(\mathrm{Na_{v}}1.7\) in der Weiterleitung schmerzrelevanter Signale hin.
Erythromelalgie und PEPD werden durch Mutationen hervorgerufen, die zu einer Funktionssteigerung eines spannungsabhängigen Na\({}^{+}\)-Kanals führen (Gain-of-function-Mutationen). Eine Verschiebung der Strom-Spannungs-Kennlinie nach links in Richtung hyperpolarisierter Potenziale aktiviert die mutierten Kanäle bereits bei schwachen Depolarisationen, von denen Wildtypkanäle normalerweise nicht geöffnet werden (Abb. 15.12a). Darüber hinaus beeinträchtigen die Gain-of-function-Mutationen die Inaktivierung der \(\mathrm{Na_{v}}1.7\)-Kanäle, sodass sie im Vergleich zum Wildtyp erst bei positiveren Spannungen bzw. zu einem geringen Teil überhaupt nicht inaktivieren (Abb. 15.12b).

Punktmutationen verändern die biophysikalischen Eigenschaften von \(\mathrm{Na_{v}}1.7\)-Kanälen. a Die Strom-Spannungs-Kennlinie zeigt die Größe des Stroms in Abhängigkeit von der Spannung. Zur besseren Vergleichbarkeit wurde der Strom auf die maximale Stromamplitude normiert. Der unveränderte Wildtyp-(WT-)Kanal öffnet bei etwa −50 mV und erreicht einen maximalen Einwärtsstrom bei −10 mV. Eine Erythromelalgie verursachende Punktmutation (P1308L) im Gen des \(\mathrm{Na_{v}}1.7\)-Kanals führt zu einer Verschiebung der Strom-Spannungs-Kennlinie nach links, sodass der Kanal bereits bei Spannungen im Bereich des Ruhemembranpotenzials öffnet. b Die schnelle Inaktivierung schließt spannungsabhängige Na\({}^{+}\)-Kanäle und verhindert so vorübergehend die Erzeugung von Aktionspotenzialen. Symptome der extremen paroxysmalen Schmerzstörung werden durch Punktmutationen im Gen des \(\mathrm{Na_{v}}1.7\)-Kanals hervorgerufen, die die Inaktivierung der Kanäle beeinflussen. Die Mutation G1607R verschiebt die Inaktivierung um etwa 30 mV in die positive Richtung, sodass weniger Kanäle inaktivieren, wodurch eine Übererregbarkeit in der Schmerzbahn hervorgerufen wird (nach 4 )
Alle Gain-of-function-Mutationen erweitern das Spannungsfenster, in dem der mutierte Ionenkanal für Na \({}^{+}\) permeabel ist. Infolgedessen werden Aktionspotenziale bereits bei normalerweise unterschwelligen Depolarisationen ausgelöst (Abb. 15.13a) und sie weisen im Vergleich zu den nichtmutierten Kanälen eine höhere Frequenz auf (Abb. 15.13b).

Übererregbarkeit von \(\mathrm{Na_{v}}1.7\)-Kanälen in Spinalganglien. Auslösung von Aktionspotenzialen in Neuronen von Spinalganglien, die unveränderte \(\mathrm{Na_{v}}1.7\)-Kanäle (Wildtyp, links) oder die Punktmutation F1449V exprimieren (rechts). Die Mutation F1449V tritt bei Erythromelalgie auf. Aktionspotenziale werden durch Injektion eines positiven Stroms erzeugt. a Bei der mutanten Form des Kanals wird bereits bei Stromamplituden ein Aktionspotenzial ausgelöst, die bei der Wildtypform des Kanals nur eine unterschwellige Depolarisation verursachen. Die gestrichelte Linie markiert den Schwellenwert zur Auslösung eines Aktionspotenzials. b Eine überschwellige Depolarisation löst im Wildtyp nur relativ wenige Aktionspotenziale aus, während eine gleich starke Depolarisation die Aktionspotenzialfrequenz bei der Punktmutation deutlich erhöht (nach 4 )
\(\mathrm{Na_{v}}1.9\) ist ein weiterer spannungsgesteuerter Na\({}^{+}\)-Kanal, der von nozizeptiven Afferenzen in den Spinalganglien und den Trigeminusganglien exprimiert wird. Dieser Kanal zeigt eine Reihe pharmakologischer und elektrophysiologischer Besonderheiten:
-
1.
Resistenz gegenüber Tetrodotoxin (Abschn. 10.4.4),
-
2.
Aktivierung bei negativeren Potenzialen,
-
3.
Inaktivierung bei positiveren Potenzialen,
-
4.
extrem langsame Inaktivierung.
Die Eigenschaften 2 und 3 führen zu dauerhaft offenen Na\({}^{+}\)-Kanälen in einem Spannungsbereich zwischen −70 mV und −40 mV und daher zu einem kontinuierlichen Einstrom von Na\({}^{+}\)-Ionen beim Ruhemembranpotenzial (Abb. 15.14).

Biophysikalische Eigenschaften von \(\mathrm{Na_{v}}1.9\)-Kanälen. Aktivierungs- und Inaktivierungskurven für \(\mathrm{Na_{v}}1.9\)-Kanäle sowie für TTX-sensitive Na\({}^{+}\)-Kanäle sind in einem einzigen Diagramm dargestellt. Im Vergleich zu den TTX-sensitiven Na\({}^{+}\)-Kanälen überlappen Aktivierungs- und Inaktivierungskurven der \(\mathrm{Na_{v}}1.9\)-Kanäle in einem breiteren Spannungsfenster (schattierte Bereiche). In diesem Spannungsbereich sind die Kanäle konstitutiv geöffnet und bewirken so eine leichtere Erregbarkeit der Zellen (nach 5 )
\(\mathrm{Na_{v}}1.9\) trägt mit seiner Na\({}^{+}\)-Leitfähigkeit zu einem vergleichsweise depolarisierten Ruhemembranpotenzial bei und verlängert das durch unterschwellige Reize erzeugte Rezeptorpotenzial. Ähnlich wie bei den mutierten \(\mathrm{Na_{v}}1.7\)-Kanälen wird bei \(\mathrm{Na_{v}}1.9\) der Schwellenwert für die Auslösung von Aktionspotenzialen gesenkt und die Aktionspotenzialfrequenz bei einer bestimmten Reizstärke erhöht. Der Einstrom von Na \({}^{+}\) -Ionen durch \(\mathrm{Na_{v}1.9}\) -Kanäle ist weniger für die Amplitude der in diesen Nozizeptoren erzeugten Aktionspotenziale verantwortlich, sondern bestimmt vielmehr, ob und mit welcher Frequenz Aktionspotenziale generiert werden. Diese Eigenschaften prädestinieren \(\mathrm{Na_{v}}1.9\) für eine Rolle im Rahmen der Signalleitung in der Schmerzbahn.
Gain-of-function-Mutationen im Gen SCN11A, das den \(\mathrm{Na_{v}}1.9\)-Kanal codiert, rufen eine Übererregbarkeit sensorischer Neurone in den Spinalganglien hervor, die wiederum für periphere Neuropathien verantwortlich ist.Footnote 19 Diese Übererregbarkeit äußert sich in Form eines depolarisierten Ruhepotenzials, einer höheren Zahl von Neuronen mit Spontanaktivität sowie einer erhöhten Aktionspotenzialfrequenz bei einem schwachen depolarisierenden Stimulus.
Neben der Signaltransduktion in der Membran der freien Nervenendigungen spielen die Erzeugung und Weiterleitung von Aktionspotenzialen in den peripheren Nerven eine entscheidende Rolle für die Schmerzwahrnehmung. Durch einen einzigen Aminosäureaustausch kann sich die spannungsabhängige Aktivierung von Na\({}^{+}\)-Kanalisoformen, die selektiv in der Schmerzbahn exprimiert werden, zu etwas positiveren Potenzialen verschieben, wodurch Übererregbarkeit und chronische Schmerzen ausgelöst werden. Umgekehrt bewirkt ein Funktionsverlust dieser Na\({}^{+}\)-Kanäle eine vollständige Unempfindlichkeit gegenüber Schmerzen, ohne dass andere sensorische Leistungen eingeschränkt werden. Aufgrund ihrer spezifischen Expression in den peripheren Schmerzbahnen sind diese Na\({}^{+}\)-Kanalisoformen vielversprechende Zielstrukturen für eine effektive therapeutische Schmerzbehandlung, die andere Na\({}^{+}\)-kanalvermittelte Funktionen unbeeinträchtigt lässt.
3.5 Zusammenfassung
Schmerz ist ein außerordentlich wichtiges Warnsignal, das den Körper vor schädigenden mechanischen, thermischen und chemischen Einflüssen schützt. Spezifische Schmerzfasern, die als freie Nervenendigungen in der Epidermis der Haut und den inneren Organen liegen, wandeln noxische Reize in elektrische Signale um, die vom Gehirn als Schmerz interpretiert werden und zu angepassten Verhaltensänderungen führen. Hochschwellige Nozizeptoren, die vor allem auf starke mechanische Stimuli reagieren, senden Signale über myelinisierte Aδ-Fasern ins Rückenmark und vermitteln einen kurzen, scharfen ersten Schmerz. Polymodale Nozizeptoren leiten thermische und chemische Reize mittels nichtmyelinisierter C-Fasern weiter und rufen einen länger andauernden, brennenden zweiten Schmerz hervor. Schlafende Nozizeptoren werden erst durch Entzündungsprozesse aktiviert und verursachen eine Senkung der Schmerzschwelle (primäre Hyperalgesie).
Noxische thermische, aber möglicherweise auch mechanische und chemische Reize werden von unspezifischen Kationenkanälen der TRP-Familie in ein Rezeptorpotenzial umgewandelt. Bei Verletzungen spielt das Entzündungsmilieu, das aus ATP, K\({}^{+}\)-Ionen, Bradykinin, Prostaglandinen, verschiedenen Zytokinen und zahlreichen anderen Substanzen besteht, eine wesentliche Rolle für die Schmerzentstehung. Bradykinin und Prostaglandin E2 binden an spezifische Rezeptoren in der Membran der freien Nervenendigung und aktivieren intrazelluläre Signaltransduktionswege, an deren Ende Phosphorylierungsprozesse die Leitfähigkeit von TRP-Kanälen erhöhen.
Die zentrale Sensibilisierung (sekundäre Hyperalgesie) wird durch plastische Veränderungen der Synapsen zwischen Nozizeptoren und den 2. Neuronen im Rückenmark aufgrund andauernder unterschwelliger Aktivität hervorgerufen. Sie kann zu schwer therapierbaren chronischen neuropathischen Schmerzen führen.
Alle nozizeptiven Bahnen, aber auch Fasern, die nichtnozizeptive thermische Signale weiterleiten, kreuzen etwa auf der Höhe ihres Eintritts ins Rückenmark auf die kontralaterale Seite und steigen anschließend im Vorderseitenstrang gebündelt zum Gehirn auf. Eine Halbseitenläsion des Rückenmarks führt daher zu einer differenziellen Empfindungsstörung, dem Brown-Séquard-Syndrom.
Die sensorisch diskriminative Bewertung eines Schmerzreizes – Lokalisation, Intensität und Art des Schmerzes – erfolgt im somatosensorischen Cortex. Emotionale Aspekte werden hingegen in anderen Hirnregionen verarbeitet, wobei das limbische System mit Amygdala und cingulärem Cortex eine zentrale Bedeutung besitzt.
Die subjektive Schmerzwahrnehmung wird durch absteigende Bahnen, die endogene Opioide als Neurotransmitter verwenden, maßgeblich beeinflusst. Auf diese Weise können die Aktivität von Projektionsneuronen der Schmerzbahn und damit das Schmerzempfinden situationsbedingt angepasst werden.
Die zelltypspezifische Expression der Na\({}^{+}\)-Kanalisoformen \(\mathrm{Na_{v}}1.7-1.9\) in den peripheren Nervenendigungen sensorischer Neurone ist für die Erzeugung von Aktionspotenzialen zur Weiterleitung schmerzrelevanter Signale von großer Bedeutung. Gain-of-function-Mutationen in diesen Kanälen führen zu Übererregbarkeit und gesteigertem Schmerzempfinden, während ein Funktionsverlust eine vollständige Schmerzunempfindlichkeit zur Folge hat.
Notes
- 1.
Wir sprechen im Folgenden von Sensoren (Mechanosensoren, Thermosensoren etc.), wenn Sinneszellen bezeichnet werden sollen. Der Begriff Rezeptor wird für molekulare Strukturen, wie beispielsweise mechano- oder thermosensitive Ionenkanäle, verwendet (s. a. (Abschn. 10.4.4), Fußnote 17).
- 2.
Unter taktilen Reizen verstehen wir Druck, Spannungen und Vibrationen der Haut, die mithilfe des Tastsinns registriert werden.
- 3.
Eine räumliche Auflösung von 0,5 mm bedeutet, dass Oberflächenstrukturen, die einen halben Millimeter auseinander liegen, als zwei unterschiedliche Punkte wahrgenommen werden (Zwei-Punkt-Schwelle).
- 4.
Ruffini-Körperchen kommen auch in Sehnen und im Zahnhalteapparat vor.
- 5.
Papillarleisten bilden als charakteristische Hautlinien die strukturelle Grundlage für Fingerabdrücke.
- 6.
Perineuralzellen bilden die zellulären Bestandteile des Perineuriums, das die Axone peripherer Nerven bündelt. Perineuralzellen sind flache Epithelzellen, die den Fibroblasten des Bindegewebes ähneln.
- 7.
Der Nervus trigeminus ist der V. Hirnnerv. Er tritt auf der rechten und linken Körperseite jeweils lateral von der Brücke aus, wo er ein sensibles Ganglion, das Ganglion trigeminale bildet.
- 8.
Die Kationenkanäle Piezo1 und Piezo2 gelten als vielversprechende Kandidaten für die Mechanotransduktion in der Haut 1 .
- 9.
Zum Verschaltungsprinzip s. Abb. Abb. 12.18.
- 10.
Eine Wahrnehmung der absoluten Temperatur gibt es nicht. Es können lediglich Temperaturunterschiede zwischen verschiedenen Objekten oder Abweichungen von der Thermoneutralzone wahrgenommen werden.
- 11.
Mögliche Kandidaten umfassen Ionenkanäle, die nicht zur TRP-Familie gehören, beispielsweise temperaturabhängige K\({}^{+}\)-Kanäle, die auch für das Ruhemembranpotenzial verantwortlich sind 13 .
- 12.
Die Rolle von TRPA1 als Nozizeptor für Kältereize wird aufgrund z. T. widersprüchlicher Befunde kontrovers diskutiert.
- 13.
Bradykinin gehört zu den Kininen und kommt vor allem im Blutplasma vor, wo es eine gefäßerweiternde Wirkung besitzt.
- 14.
Schmerzmittel mit dieser Wirkungsweise werden als nichtsteroidale Entzündungshemmer (Antiphlogistika) bezeichnet. Sie unterscheiden sich von den steroidalen Antiphlogistika, bei denen es sich um Glucocorticoide handelt, die die Phospholipase A2 hemmen und somit die Bildung von Arachidonsäure aus Phospholipiden verhindern.
- 15.
Das limbische System besteht aus einer Reihe subkortikaler Kerngebiete, wie der Amygdala, sowie kortikaler Regionen, beispielsweise dem cingulären Cortex. Das limbische System spielt eine wesentliche Rolle bei der Integration emotionalen Verhaltens.
- 16.
Unter einem Placeboeffekt verstehen wir eine physiologische Reaktion auf eine pharmakologisch unwirksame Substanz wie Kochsalz oder Glucose.
- 17.
Bei Säugern wurden bisher neun Isoformen spannungsgesteuerter Na\({}^{+}\)-Kanäle nachgewiesen, von denen drei – \(\mathrm{Na_{v}}1.7\), \(\mathrm{Na_{v}}1.8\) und \(\mathrm{Na_{v}}1.9\) – in nozizeptiven Neuronen der Spinalganglien exprimiert werden und an der Signalleitung in den Schmerzbahnen beteiligt sind.
- 18.
Die Symptomatik der PEPD ist ähnlich der EEM, allerdings sind die Schmerzattacken auf rektale, okuläre und mandibuläre Regionen beschränkt.
- 19.
Neuropathische Schmerzen (Neuralgien) werden nicht durch eine Verletzung von Gewebe verursacht, sondern entstehen aufgrund von Schädigungen oder Funktionsstörungen in den signalleitenden Nerven. Bei einer peripheren Neuropathie sind die Axone der pseudounipolaren Neurone in den Spinalganglien betroffen.
Literatur
Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, Dubin AE, Patapoutian A (2010) Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 330:55–60
Costigan M, Scholz J, Woolf CJ (2009) Neuropathic pain: A maladaptive response of the nervous system to damage. Annu Rev Neurosci 32:1–32
Delmas P, Hao J, Rodat-Despoix L (2011) Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat Rev Neurosci 12:139–153
Dib-Hajj SD, Yang Y, Black JA, Waxman SG (2013) The Nav1.7 sodium channel: From molecule to man. Nat Rev Neurosci 14:49–62
Dib-Hajj SD, Black JA, Waxman SG (2015) Nav1.9: A sodium channel linked to human pain. Nat Rev Neurosci 16:511–519
Fields H (2004) State-dependent opioid control of pain. Nat Rev Neurosci 5:565–575
Flor H, Nikolajsen L, Jensen TS (2006) Phantom limb pain: A case of maladaptive CNS plasticity? Nat Rev Neurosci 7:873–881
Lumpkin EA, Caterina MJ (2007) Mechanisms of sensory transduction in the skin. Nature 445:858–865
Maksimovic S, Nakatani M, Baba Y, Nelson AM, Marshall KL, Wellnitz SA, Firozi P, Woo SH, Ranade S, Patapoutian A, Lumpkin EA (2014) Epidermal Merkel cells are mechanosensory cells that tune mammalian touch receptors. Nature 509:617–621
Merabat LB, Pascual-Leone A (2010) Neural reorganization following sensory loss: The opportunity of change. Nat Rev Neurosci 11:44–52
Patapoutian A, Peier AM, Story GM, Viswanath V (2003) ThermoTRP channels and beyond: Mechanisms of temperature sensation. Nat Rev Neurosci 4:529–539
Purves D, Augustine GJ, Fitzpatrick D, Hall WC (2012) Neuroscience, 5. Aufl. Sinauer, Sunderland
Vriens J, Nilius B, Voets T (2014) Peripheral thermosensation in mammals. Nat Rev Neurosci 15:573–589
Woo SH, Ranade S, Weyer AD, Dubin AE, Baba Y, Qiu Z, Petrus M, Miyamoto T, Reddy K, Lumpkin EA, Stucky CL, Patapoutian A (2014) Piezo2 is required for Merkel-cell mechanotransduction. Nature 509:622–626
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Copyright information
© 2017 Springer-Verlag GmbH Deutschland
About this chapter

Cite this chapter
Feigenspan, A. (2017). Somatosensorik. In: Prinzipien der Physiologie. Springer Spektrum, Berlin, Heidelberg. https://doi.org/10.1007/978-3-662-54117-3_15
Download citation
DOI: https://doi.org/10.1007/978-3-662-54117-3_15
Published:
Publisher Name: Springer Spektrum, Berlin, Heidelberg
Print ISBN: 978-3-662-54116-6
Online ISBN: 978-3-662-54117-3
eBook Packages: Life Science and Basic Disciplines (German Language)