
Abstract
Dam construction goes back in human history for more than 5000 years (e.g., Sadd el-Kafara dam in Egypt for flood protection), but most of the world’s existing dams have been built after the Second World War as consequence or basis of economic development. Today, there are about 6000 existing or planned large hydropower dams (>15 m height) worldwide (Zarfl et al. 2014) and an uncountable number of small dams. For example, with more than 5000 mostly small hydropower plants, Austria is one of the countries with the highest density of hydropower dams (about 6 dams per 100 km2, Wagner et al. 2015). Downstream flows are mainly altered by large dams, e.g., there are 654 reservoirs with storage capacities ≥0.5 km3 (Lehner and Döll 2004). Damming rivers currently stores the equivalent of 15% of global annual river runoff (Likens 2010). As a result, 48% of rivers (expressed as river volume) globally are moderately to severely impacted by either flow regulation, fragmentation, or both.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


1 Introduction
Dam construction goes back in human history for more than 5000 years (e.g., Sadd el-Kafara dam in Egypt for flood protection), but most of the world’s existing dams have been built after the Second World War as consequence or basis of economic development. Today, there are about 6000 existing or planned large hydropower dams (>15 m height) worldwide (Zarfl et al. 2014) and an uncountable number of small dams. For example, with more than 5000 mostly small hydropower plants, Austria is one of the countries with the highest density of hydropower dams (about 6 dams per 100 km2, Wagner et al. 2015). Downstream flows are mainly altered by large dams, e.g., there are 654 reservoirs with storage capacities ≥0.5 km3 (Lehner and Döll 2004). Damming rivers currently stores the equivalent of 15% of global annual river runoff (Likens 2010). As a result, 48% of rivers (expressed as river volume) globally are moderately to severely impacted by either flow regulation, fragmentation, or both.
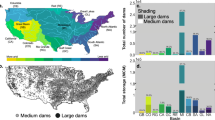
Besides flow, sediment transport is severely altered by dams. A total of approximately 25–30% of pre-disturbance sediment flux is sequestered by modern impoundments (Fig. 6.1).

Sediment trapping by large dams. GWSP Digital Water Atlas (Available online at http://atlas.gwsp.org)
Impacts might double should all planned dams be constructed by 2030 (Grill et al. 2015). This is especially so in areas with expanding economies and extensive unexploited river reaches, such as China, which currently is building 130 major dams in its Southwest (Lewis 2013) and has constructed more than half the new dams built since 1950 worldwide (Wang and Chen 2010).
Unfortunately, there is no generally accepted descriptive nomenclature of dams. The term “dam” is often applied to both the physical structure retaining the water and the water so retained. For the purposes of this chapter, dam will be used solely to describe the physical structure (e.g., weir), and the term “reservoir” will be used to denote the artificially created water body. This leads to the following definition: “A dam is a barrier to obstruct the flow of water and to create a reservoir.” Reservoirs are also called “impoundments.” Reservoirs are built for specific community needs:
-
Drinking, industrial, and cooling water supply
-
Hydropower generation
-
Agricultural irrigation
-
River regulation and flood control
-
Navigation
-
Recreation and fisheries
Dams are among the most damaging human activities in river basins, deeply modifying the physiography of watersheds. Reservoirs may look very much like natural lakes; however, the operating regime determined by the purpose for which the reservoirs were created may significantly alter their physicochemical character and biological responses. The peculiar form of a reservoir, its location, and mode of operation may cause considerable, actual variation of the basic limnological behavior. Reservoirs undergo great changes in water quality during the early stages of their formation until a new ecological balance becomes established (Straskraba et al. 1993). Reservoirs follow a succession of (1) physicochemical alteration, (2) modification in the structure and dynamics of primary producers, and (3) changes in the community of consumers, especially invertebrates and fish (Petts 1985). After that, reservoirs may pass over into a kind of stability, but occurrence of floods, dam operation, or other impacts may create new disturbances to the system.
Reservoirs not only affect the inundated river sections but also block upstream fish migration (see Chap. 9) and downstream flow and sediment transport. The magnitude of impact is strongly correlated with the location of the dam, size of reservoir (height of dam, volume of reservoir), and water residence time. The average length of time water remains within the boundaries of an aquatic system is one of the key parameters controlling the system’s biogeochemical behavior. This time scale, which is generally referred to as the water residence time, is fundamental for multiple and complex processes in reservoirs (Rueda et al. 2006). Furthermore, the dam operation mode determines the seasonal variation of stored water, water level fluctuations, sediment capture and release, as well as daily and seasonal downstream flow patterns.
The main impacts associated with reservoirs are as follows:
-
Interruption of river continuity (longitudinal and lateral, fish migration, sediment and nutrient transport)
-
Siltation of river bed and clogging of interstitial
-
Homogenization of habitats
-
Downstream river bed incision
-
Alteration of river/groundwater exchange
-
Downstream flow and water quality alteration
2 Transforming Rivers to Reservoirs
River damming is a process so drastic that it results in the creation of a completely new ecosystem (Baxter 1977). Therefore, the occurrence of environmental impacts is inherent with any impoundment due to fundamental change of the hydrology and morphology of the river. Flow represents the main force behind freshwater ecosystems, and it is responsible for geohydrological structure, matter and energy fluxes, system productivity, and distribution and function of biota (Poff et al. 1997). As a consequence, the alteration of natural flow regimes and morphodynamic patterns has far-reaching impacts (see Chaps. 3 and 4), including production, biodiversity, and changes in functions and services provided by aquatic ecosystems (Nilsson et al. 2005).
Reservoirs differ from natural lakes with respect to hydrological, limnological, and ecological dynamics. Depending on the size and shape of the reservoir, a longitudinal hydrological gradient may develop from the dam (lentic or lacustrine zone) to upstream reaches (riverine zone), showing intermediate characteristics in middle stretches (transition zone, with lentic and lotic features) (Kimmel and Groeger 1984). While fluvial characteristics are maintained to some extent in small reservoirs, e.g., run-of-the-river hydropower plants, lentic conditions prevail in large storage reservoirs (Fig. 6.2).

Schematic representation of longitudinal reservoir zones and associated key processes, (a) plan view and (b) longitudinal profile; DMM dissolved mineral matter, DOM dissolved organic matter, PMM particulate mineral matter, POM particulate organic matter, BA bacteria, FU fungi (adapted after Herzig 1984; Kimmel and Groeger 1984)
Coarse sediments settle within the riverine section of the reservoir, while fine sediments (sand, silt) and particulate organic matter (POM) are deposited in the lacustrine zone (Fig. 6.2, Table 6.1). Soon after the reservoir is filled, patterns of thermal/chemical stratification intensify progressively in the water column, and eutrophication may occur due to upstream matter input, decay, and nutrient release from the flooded organic matter (vegetation, litter, and soil) or from pollution. Reservoirs are much more susceptible to eutrophication that rivers due to the higher self-purification capacity of running waters. Consequently, water quality may deteriorate in reservoirs (e.g., thermal stress, low dissolved oxygen, acidification), especially close to the bottom (Agostinho et al. 2008). Depending on a reservoir’s characteristics, anoxic water or sediment layers may evolve as a consequence of stratification, deposition, and decomposition of organic material. Stratification (thermocline, light conditions, etc.) changes production (autotrophic, heterotrophic) and the entire food chain (Fig. 6.2).
Dams are often associated with lateral dams or levees, disconnecting the reservoir hydrologically from the floodplains. This results in limited or abandoned inundation and reduced interchange with the groundwater and lowers chances of recolonization, both lateral and longitudinal. As a result, the new ecosystem is colonized by those species that inhabited the original river and are able to adapt to the new conditions (Agostinho et al. 2008). Nonmigratory, eurytopic species dominate the lacustrine zone of reservoirs because they usually have less complex requirements with regard to life-cycle dynamics. Migratory and rheophilic species experience declines in the reservoir due to the lentic environment, spatial fragmentation imposed by the dam, and the loss of critical habitats (e.g., spawning habitats in free-flowing river stretches). Consequently, lentic fish replace lotic species and dominate the reservoir fish communities (Zhong and Power 1996). In terms of richness, fish species tend to remain in environments that preserve the original fluvial characteristics or in those with relatively high habitat heterogeneity, i.e., tributaries and lotic stretches upstream, and it is common to find a gradient of decreasing fish diversity toward dams. If coldwater streams are dammed, the warmer water in reservoirs might favor species adapter to warmer temperatures, resulting in a so-called potamalization effect, i.e., a shift from rhithral to potamal communities (Jungwirth et al. 2003).
Fine sediment deposition in the reservoir leads to clogging of the river bottom (see Chap. 8). This affects aquatic communities of the hyporheic interstitial (Ward et al. 1998), resulting in depauperated fauna dominated by few species (e.g., chironomids, Table 6.1). As a consequence, the ecological status of reservoirs, in particular within the lacustrine section, is often classified as poor or bad (sensu EU Water Framework Directive, Ofenböck et al. 2011, Fig. 6.3).

Response of macroinvertebrate communities to increased proportion of fine sediments (akal/psammal/pelal) in three impounded streams (Austria) expressed as multimetric index (1 = high ecological status, 0 = bad ecological status; adapted from Ofenböck et al. 2011)
While the energy and matter fluxes in the riverine section are based on allochthonous matter input, the energy fluxes of the lacustrine section are also triggered by photosynthesis and inner cycling. Allochthonous organic matter (DOM, dissolved organic matter; POM, particulate organic) is directly taken up by consumers or indirectly via detritus decomposed by bacteria and fungi. Besides water residence time, mineral components (DMM, dissolved mineral matter) and light conditions (PMM, particulate mineral matter; POM) regulate algal growth and overall productivity in the lacustrine section (see Fig. 6.2 and Herzig 1984).
In all three zones of the reservoir, biodiversity is highest in the littoral environment as a result of the greater availability and heterogeneity of feeding resources, shelter, and habitats (Agostinho et al. 2008). However, the littoral may be exposed to water level fluctuations, causing frequent stress events to fauna and flora. High magnitude and frequency of water level fluctuations creates “dead zones” along the reservoir shores.
Whenever rivers are turned into reservoirs, the former fluvial habitat is widely lost. The new lentic ecosystem resembles lake-type systems, but, depending on type and dam operation, reservoirs are disturbed by artificial water level fluctuations, drawdowns, and floods. Consequently, dammed rivers are hybrid systems that lose their lotic but gain only partly lacustrine functions.
Sediment trapping by reservoirs is well recognized, with the extent to which sediments are trapped dependent on the morphology of the impoundment, the characteristics (grain size) of the inflowing sediments, the hydrodynamics within the impoundment, and the operating regime of the dam. Smaller impoundments have lower rates as compared to the larger reservoirs. Sediment trapping is cumulative, but as coarser material is trapped in upstream impoundments, the actual trapping rates in downstream impoundments may decline, due to the finer nature of the influent sediment load. This preferential trapping can also lead to nutrients being trapped in different proportions as compared to sediments, due to nutrients’ affinity for the finer-grained sediments (Koehnken 2014).
As an example, the dam cascade at the Lancang River (upper Mekong) has the potential to trap most of the sediments (Kummu and Varis 2007, Fig. 6.4). Recent monitoring results suggest that suspended sediment loads in the lower Mekong basin (downstream of China) are now in the range of 44% compared with the historic values (~70 Mt/year compared with ~160 Mt/year; Koehnken 2014).

Average theoretical sediment trapping efficiency of reservoirs of the Lancang (Upper Mekong) Cascade in China (Kummu and Varis 2007) (reproduced from Geomorphology, 85/3–4, 2007, Matti Kummu, Olli Varis, Sediment-related impacts due to upstream reservoir trapping, the lower Mekong river, pp. 275–293, with permission from Elsevier. © 2007 Elsevier Inc. All rights reserved)
3 Downstream Effects
Although investigated less, downstream impacts are equally or even more damaging to aquatic fauna, given that impoundments affect primarily water flow dynamics, i.e., the main force working in fluvial ecosystems. Impoundments redistribute river discharge in space and time, affecting several hydrological attributes, e.g., flood period, intensity, amplitude, duration, frequency, and, consequently, the structure, dynamics, and functioning of ecosystems located downstream (see Chap. 4). In addition to flow regulation, other important alterations inevitably follow dam construction, such as blockage of migration routes for some fish species and the retention of sediments and nutrients upstream—a process that decreases turbidity as well as nutrient load and suspended material. This last phenomenon imposes limitations on biological productivity in areas downstream, reducing the fertility of wetlands, and affecting their carrying capacity. The loss of fertilizing services of the flood pulse has been documented in several systems worldwide (WCD 2000).
In case of downstream floodplains, negative effects of flow regulation are still more pronounced. The structure and functioning of such ecosystems rely on the alternation of extreme events, e.g., flood and drought (Junk et al. 1989), a dynamic pulsing that disappears with impoundment, because dams usually decrease maximum discharges (absence of seasonal flood pulses) and stabilize or increase minimum discharges. Consequently, hydrological connectivity among environments is considerably modified in space and time. The redistribution of the flooding regime has several direct and indirect effects on fish populations. The decrease in connectivity between the river and lateral floodplain affects riparian communities and reshapes other environments and interface zones that provide important habitats for fish, especially nurseries. In the absence of floods, even if adults successfully reproduce in tributaries, eggs, larvae, and young fish drifting downstream have limited access to lateral habitats, and population recruitment is negatively affected (Agostinho et al. 2008).
In 1960, the Volgograd hydropower dam, the last dam of the Volga–Kama cascade, was completed. After damming, annual maximum peak discharges have decreased, minimum discharges increased, but average discharges remained similar to pre-damming conditions (Fig. 6.5). Moreover, because of riverbed incision of over 1.5 m, a higher discharge is needed to reach bank-full level and to inundate floodplains, which is the largest floodplain in Europe (length 300 km, width 20 km). Commercial fish catches severely decreased after damming, both in the main channel and in the floodplain lakes (Fig. 6.6).

Alteration of discharge as a result of upstream damming in the Volga River (adapted from Górski et al. 2012)

Post-damming commercial fisheries catch decline in the Volga River (adapted from Górski et al. 2012)
If the continuity of sediment transport is interrupted by dams or removal of sediment from the channel by gravel mining, the flow may become sediment starved (hungry water) and prone to erode the channel bed and banks, producing channel incision (downcutting), coarsening of bed material, and loss of habitat for litophilic species (Kondolf 1997; Fig. 6.7). Riverbed incision reduces the connectivity to floodplain habitats. Together with reduced flood flows (e.g., due to storage) the dimension and quality of floodplain habitats is reduced, affecting the productivity of the entire river-floodplain system. Also, further downstream, lack of sediment may cause habitat degradation due to erosion of river deltas or coastal shores.

Riverbed incision of the Danube River downstream of the Austrian hydropower cascade in the years 1950–2003 expressed as low water level (m.a.sl. meter above sea level) at river 1894.7 km (gauging station Wildungsmauer, Reckendorfer et al. 2005) (© Reckendorfer, W. et al. 2005. The Integrated River Engineering Project for the free-flowing Danube in the Austrian Alluvial Zone National Park: contradictory goals and mutual solutions Archiv für Hydrobiologie, Supplementband “Large Rivers”, 155: 613–630, www.schweizerbart.de/series/archiv_Suppl, reproduced with permission from Schweizerbart’sche Verlagsbuchhandlung)
4 Other Downstream Impacts
Downstream segments are also subjected to other impacts related to dam operation and water quality of released water. The operation of hydroelectric impoundments tends to follow demands for electricity, creating variable flow regimes. Such irregular discharges called hydropeaking intensify erosive processes downstream and can caused drift and stranding of fish and macroinvertebrates (see Chap. 5).
Downstream release of poor-quality water by turbines and spillways also creates unfavorable conditions, e.g., anoxic hypolimnic water and altered water temperatures. For example, below the Xinanjiang and Danjiangkou dams, spawning of fish was delayed 20–60 days by lower water temperatures (Zhong and Power 1996). Spillflow at high dams may cause oversaturation of oxygen creating the so-called gas bubble disease in fish.
5 Mitigation Measures
Reservoirs impose system shifts on running waters, making restoration in the sense of reestablishing pre-damming conditions impossible. Hence, any attempt to improve the ecological condition can be regarded as a mitigation effort to reduce but not to remove the impacts. This is definitely true for the reservoir itself where mitigation measures have to take into account the new boundary conditions of the lacustrine environment. Therefore, mitigation measures mainly focus on the habitat improvements of the riverine section of reservoirs (head section) and on river sections located up- and downstream of the reservoir (Fig. 6.8). Focus is given to the reestablishment of longitudinal continuity by enabling fish migration through fish passes (see Chap. 9). Downstream mitigation measures involve environmental flow regulations (see Chaps. 4 and 5) and sediment transport by targeted sediment management (see Chap. 8).

Mitigation measures proposed for Danube reservoirs for improving connectivity and habitat (Jungwirth et al. 2005) (© Jungwirth, M et al. (2005). Leitbild-specific measures for the rehabilitation of the heavily modified Austrian Danube River. Archiv für Hydrobiologie, Supplementband “Large Rivers”, 155: 17–36, www.schweizerbart.de/series/archiv_Suppl, reproduced with permission from Schweizerbart’sche Verlagsbuchhandlung)
5.1 Reestablishing Longitudinal Continuity
A common restoration measure for dams is the implementation of fish passes to enable upstream fish passage (for more details on fish passage, see Chap. 9). While fish passes have proven to be effective to pass fish across dams when constructed according to the requirements of migrating species, their role in effectively maintaining populations in dam cascades is still unclear. Even highly efficient fish passes may not be able to pass enough fish upstream when fish have to negotiate multiple dams.
Some studies suggest that ladders are problematic in fish conservation as they lead fish into ecological traps (Pelicice and Agostinho 2008; Pelicice et al. 2015). Migratory fishes travel long distances during the reproductive season in search of habitats suitable for spawning and the development of young. The movement is mostly upstream and, in case of dams, often supported by fish passes. After spawning adults migrate back to their downstream habitats in main rivers. Eggs and/or larvae are then carried downstream by currents but are not further propagated downstream of reservoirs. This leads to lack of recruitment for riverine fish populations downstream of the dams.
Reservoirs themselves often provide unsuitable habitats for juvenile fish and finally represent ecological traps. Four conditions are required to characterize a fish passage/reservoir system as an ecological trap (Pelicice and Agostinho 2008): (1) attractive forces leading fish to ascend the passage; (2) unidirectional migratory movements (upstream); (3) the environment above the passage has poor conditions for fish recruitment, e.g., the absence of spawning grounds and nursery areas; and (4) the environment below the passage has a proper structure for recruitment. When these conditions exist, individuals move to poor-quality habitats, fitness is reduced, and populations are threatened. Based on current and proposed river regulation scenarios, it is concluded that conservation of migratory fish will be much more complicated than previously believed (Pelicice et al. 2015).
5.2 Sediment Management
The overarching goal of sediment management should be to make dams transparent to sediment transport as much as possible. Management options include sediment flushing, sediment bypass, and sediment augmentation downstream of reservoirs (Kondolf et al. 2014). Some general guidance relating the size, water inflows and sediment inflows, and applicable mitigation measures was developed by Basson and Rooseboom (1997), who identified a relationship between the capacity of reservoirs and the mean annual water and sediment inflows and appropriate mitigation measures (Fig. 6.9):

Reservoir management options depending on storage capacity, mean annual runoff, and mean annual sediment load (based on Basson and Rooseboom 1997 and adapted from Kimbrel et al. 2014). Kw is the ratio between reservoir storage capacity and mean annual runoff, and a ratio of >1 means that the reservoir can store more than the mean annual runoff. Kt is the ratio between reservoir storage and mean annual sediment yield. A ratio of 100 means that the mean annual sediment yield can be stored over 100 years until the reservoir is filled (without flushing)
Sediment sluicing: The aim of sediment sluicing is to maintain sediment in suspension and move it through the impoundment prior to deposition. Sediment sluicing typically involves a reduction in the water level in the impoundment by opening gates when sediment concentrations are elevated. Turbidity venting is similar to sediment sluicing but uses low level gates or deep sluices to enable sediment laden water to “flow” along the bottom of the reservoir to the toe of the dam.
For sediment flushing, reservoir levels are reduced to pre-impoundment levels, enabling the “river” to erode deposited sediments. At least twice the mean annual flow is required.
Bypass structures, whether they are tunnels, constructed canals, or existing river channels, can be used to pass high sediment-bearing water and bedload around an impoundment, thus decreasing the trapping of sediment. An advantage is that the seasonality of sediment delivery to the downstream river is maintained (e.g., Lake Miwa, Japan).
Reintroduction of dredged material: Approaches for reintroducing excavated or recovered material back into the downstream channel are implemented at Danube River downstream of Vienna, Austria, and Rhine River at Iffezheim, Germany.
Most reservoirs have a capacity mean annual flow ratio of between 0.2 and 3 and a life span of 50–2000 years when considering reservoir sedimentation. When the capacity mean annual flow ratio is less than 0.03, sediment sluicing or flushing should be carried out during floods and through large bottom outlets, preferably with free outflow conditions. Flushing is a sustainable operation and a long-term equilibrium storage capacity can be reached. Seasonal flushing for ca. 2 months per year could be used in regions where the hydrology is less variable with capacity mean annual flow ratio up to 0.2. When capacity mean annual flow ratios are, however, larger than 0.2, not enough excess water is available for flushing, and the typical operational model is storage operation. Density current venting can be practiced at these reservoirs as well as dredging to recover lost storage capacity (Basson 2004).
Reservoir flushing is an important mitigation measure for sediment remobilization and, thus, the restoration of natural sediment dynamics, including the formation of type-specific habitats. Nevertheless, reservoir flushing is also associated with immediate negative effects on physicochemical conditions, e.g., turbidity, oxygen deficiency, and hydropeaking, impacting fish directly, e.g., increased drift, gill, and skin injuries, stress, and fish kills, and indirectly, e.g., reduced food supply caused by increased drift and loss of benthic invertebrates, reduced growth, and lost habitats due to sedimentation (Henley et al. 2000; Crosa et al. 2009; Kemp et al. 2011; Jones et al. 2012).
Aquatic organisms have evolved with the dynamics of natural levels of total suspended solids (TSS). The natural level of TSS highly depends on the geo-hydromorphological conditions in the catchment and can be highly variable. The highest natural TSS peaks occur during floods and may affect fish. However, healthy ecosystems are usually able to compensate these effects in the long run. Reservoir flushing can cause TSS concentrations much higher than the natural background concentration and can result, depending on concentration level and duration, in stress or complete elimination of the fish stock.
Newcombe and Jensen (1996) classified the effects of reservoir flushing as (1) lethal effect (high-to-low mortality, high-to-medium habitat degradation), (2) lethal and para-lethal effects (high predatory pressure, prolonged hatching of larvae), (3) sublethal effects (reduction of growth, fitness and feeding, disturbed homing effect, physiologic stress, elevated breath frequency), and (4) behavioral effects (emigration, active/passive drift). While behavioral effects are mainly reversible and limited to the duration of exposure, physiologic changes have a more chronic character.
The intensity of impacts depends mainly on the concentration and duration of exposure, but also the size and texture of particles, water temperature, and chemical and physical conditions. Furthermore, toxic substances, acclimatization, and other stressors and their interaction are considered as relevant. For example, the release and decay of organic matter and resulting oxygen depletion may lead to suffocation of fish and benthic invertebrates. The “ranked effects model” is a tool for quantifying negative effects of suspended solids on fish (Newcombe and Jensen 1996). On the basis of duration (h) and concentration (mg/l) of exposure, a so-called severity of ill index (SEV) is calculated whereby several models are used depending on the species and age class. The resulting index ranges from 0 (no changes in behavior) to 14 (80–100% mortality).
5.3 Habitat Improvements in Reservoirs
Mitigation measures generally comprise “instream structures” such as gravel bars, islands, etc., “lateral widenings” of the cross profiles in riverine sections of impoundments, creating artificial habitats in lacustrine section, and “bypass systems” within the alluvial floodplains (Fig. 6.8, Jungwirth et al. 2005).
Due to raised water tables and sedimentation in the impoundments, the drastic loss of originally typical instream structures (gravel bars and islands) strongly affects reproduction and young-of-the-year habitats of the rheophilic fish community. Since the head sections of reservoirs still offer relatively high hydromorphological dynamics, river-type-specific rehabilitation can most likely be achieved here. The best way to recreate near-natural gravel bars and islands is to induce natural formation by lateral widenings of the cross profiles. Further rehabilitation measures comprise removing the embankments, reconnecting the former floodplains including abandoned arms, or forming new side arms (Fig. 6.8).
The lacustrine sections of reservoir sections are ecologically heavily degraded due to the loss of fluvial dynamics and intensive sedimentation of fine substrate. Establishing new gravel habitats would soon fail because of strong aggradation with fine sediments during high flow periods. Nevertheless, the construction of artificial, stabilized silt or sand islands at hydraulically appropriate zones along the embankments can provide valuable habitats for various species (Fig. 6.8, for more details for habitat improvements in reservoirs, see Chap. 24).
Bypass systems are designed to connect adjoining impoundments within alluvial floodplains and represent innovative solutions for enhancing longitudinal and lateral connectivity of the fragmented environment as well as for substituting lost fluvial habitat. Depending on the local situation and the ecological objectives, the bypass systems can be established by connecting existing floodplain water bodies or by constructing new artificial channels. The flow of the bypass system should mimic pre-damming flow hydrographs to enable dynamic hydromorphological processes (Fig. 6.10).

Example of a bypass system, a near-natural channel to circumvent the impoundment, implemented at hydropower station Ottensheim-Wilhering, Danube, Austria (15 km long, discharge 2.5–20 m3/s, adapted after http://www.life-netzwerk-donau.at, accessed 1.10.2016)
References
Agostinho AA, Pelicice FM, Gomes LC (2008) Dams and the fish fauna of the Neotropical region: impacts and management related to diversity and fisheries. Braz J Biol 68:1119–1132
Basson G (2004) Hydropower dams and fluvial morphological impacts – an African perspective. The 10th United Nations Symposium on Hydropower and Sustainable Development. Beijing, pp 1–16
Basson G, Rooseboom A (1997) Dealing with reservoir sedimentation. Water Research Commission, Pretoria
Baxter RM (1977) Environmental effects of dams and impoundments. Annu Rev Ecol Syst 8:255–283
Crosa G, Castelli E, Gentili G, Espa P (2009) Effects of suspended sediments from reservoir flushing on fish and macroinvertebrates in an alpine stream. Aquat Sci 72:85–95
Górski K, van den Bosch LV, van de Wolfshaar KE, Middelkoop H, Nagelkerke LAJ, Filippov OV, Zolotarev DV, Yakovlev SV, Minin AE, Winter HV, De Leeuw JJ, Buijse AD, Verreth JAJ (2012) Post-damming flow regime development in a large lowland river (Volga, Russian Federation): implications for floodplain inundation and fisheries. River Res Appl 28:1121–1134
Grill G, Lehner B, Lumsdon AE, MacDonald GK, Zarfl C, Reidy Liermann C (2015) An index-based framework for assessing patterns and trends in river fragmentation and flow regulation by global dams at multiple scales. Environ Res Lett 10:15001
Henley WF, Patterson MA, Neves RJ, Lemly AD (2000) Effects of sedimentation and turbidity on lotic food webs: a concise review for natural resource managers. Rev Fish Sci 8:125–139
Herzig A (1984) Zur Limnologie von Laufstauen alpiner Flüsse – Die Donau in Österreich. Osterreichische. Wasserwirtschaft 36:95–103
Jones JI, Murphy JFJ, Collins AL, Sear DA, Naden PS, Armitage PD (2012) The impact of fine sediment on Macro-Invertebrates. River Res Appl 28:1055–1071
Jungwirth M, Haidvogl G, Moog O, Muhar S, Schmutz S (2003) Angewandte Fischökologie an Fließgewässern. Facultas Universitätsverlag, Wien, p 547
Jungwirth M, Haidvogl G, Hohensinner S, Muahr S, Schmutz S, Waidbacher H (2005) Leitbild-specific measures for the rehabilitation of the heavily modified Austrian Danube River. Arch Hydrobiol 155:17–36
Jungwirth M, Moog O, Schmutz S (2006) Auswirkung der Stauregelung großer Flüsse auf die aquatische Tierwelt (Fische und Makro-zoobenthos). Limnologie Aktuell 12:79–98
Junk WJ, Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain systems. Can Spec Publ Fish Aquat Sci 106:110–127
Kemp P, Sear D, Collins A, Naden P, Jones I (2011) The impacts of fine sediment on riverine fish. Hydrol Process 25:1800–1821
Kimbrel S, Collins K, Randle T (2014) Formulating guidelines for reservoir sustainability plans. Denver
Kimmel BL, Groeger AW (1984) Factors controlling primary production in lakes and reservoirs: a perspective. Lake Reservoir Manage 1:277–281
Koehnken L (2014) Discharge Sediment Monitoring Project (DSMP) 2009–2013 summary & analysis of results. Vientiane
Kondolf GM (1997) Hungry water: effects of dams and gravel mining on river channels. Environ Manag 21:533–551
Kondolf GM, Gao Y, Annandale GW, Morris GL, Jiang E, Zhang J, Cao Y, Carling P, Fu K, Guo Q, Hotchkiss R, Peteuil C, Sumi T, Wang H-W, Wang Z, Wei Z, Wu B, Wu C, Yang CT (2014) Sustainable sediment management in reservoirs and regulated rivers: experiences from five continents. Earths Future 2:256–280
Kummu M, Varis O (2007) Sediment-related impacts due to upstream reservoir trapping, the Lower Mekong River. Geomorphology 85:275–293
Lehner B, Döll P (2004) Development and validation of a global database of lakes, reservoirs and wetlands. J Hydrol 296:1–22
Lewis C (2013) China’s great dam boom: a major assault on its rivers. http://e360.yale.edu/feature/chinas_great_dam_boom_an_assault_on_its_river_systems/2706/
Likens GE (2010) River ecosystem ecology: a global perspective. Academic Press, Amsterdam
Newcombe CC, Jensen JOTJ (1996) Channel suspended sediment and fisheries: a synthesis for quantitative assessment of risk and impact. N Am J Fish Manag 16:693–727
Nilsson C, Reidy CA, Dynesius M, Revenga C (2005) Fragmentation and flow regulation of the world’s large river systems. Science 308:405–408
Ofenböck T, Graf W, Hartmann A, Huber T, Leitner P, Stubauer I, Moog O (2011) Abschätzung des ökologischen Zustandes von Stauen auf Basis von Milieufaktoren. Vienna
Pelicice FM, Agostinho AA (2008) Fish-passage facilities as ecological traps in large neotropical rivers. Conserv Biol 22:180–188
Pelicice FM, Pompeu PS, Agostinho AA (2015) Large reservoirs as ecological barriers to downstream movements of Neotropical migratory fish. Fish Fish 16:697–715
Petts GE (1985) Impounded rivers – perspectives for ecological management. Wiley, Chichester
Poff NLR, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC (1997) The natural flow regime; a paradigm for river conservation and restoration. Bioscience 47(11):769–784
Reckendorfer W, Schmalfuss R, Baumgartner C, Habersack H, Hohensinner S, Jungwirth M, Schiemer F (2005) The integrated river engineering project for the free-flowing Danube in the Austrian Alluvial Zone National Park: contradictory goals and mutual solutions. Arch Hydrobiol 155:613–630
Rueda F, Moreno-Ostos E, Armengol J (2006) The residence time of river water in reservoirs. Ecol Model 191:260–274
Straskraba M, Tundisi JG, Duncan A (1993) State-of-the-art of reservoir limnology and water quality management. In: Comparative reservoir limnology and water quality management. Kluwer Academic Publisher, Dordrecht, pp 213–288
Wagner B, Hauer C, Schoder A, Habersack H (2015) A review of hydropower in Austria: past, present and future development. Renew Sust Energ Rev Elsevier 50:304–314
Wang Q, Chen Y (2010) Status and outlook of China’s free-carbon electricity. Renew Sust Energ Rev 14:1014–1025
Ward JV, Bretschko G, Brunke M, Danielopol D, Gibert J, Gonser T, Hildrew AG (1998) The boundaries of river systems: the metazoan perspective. Freshw Biol 40:531–569
WCD (2000) Dams and development. A new framework for decision making. Report. World Commission on Dams, London
Zarfl C, Lumsdon AE, Berlekamp J, Tydecks L, Tockner K (2014) A global boom in hydropower dam construction. Aquat Sci 77:161–170
Zhong Y, Power G (1996) Environmental impacts of hydroelectric projects on fish resources in China. Regul Rivers Res Manag 12:81–98
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made. The images or other third party material in this book are included in the book's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the book's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2018 The Author(s)
About this chapter

Cite this chapter
Schmutz, S., Moog, O. (2018). Dams: Ecological Impacts and Management. In: Schmutz, S., Sendzimir, J. (eds) Riverine Ecosystem Management. Aquatic Ecology Series, vol 8. Springer, Cham. https://doi.org/10.1007/978-3-319-73250-3_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-73250-3_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-73249-7
Online ISBN: 978-3-319-73250-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)