
Abstract
Large-scale transportation infrastructures, such as high-speed railway (HSR) systems, cause changes in surrounding ecosystems, thus generating direct and indirect impacts on bird communities. Such impacts are rooted in the individual responses of birds to infrastructure components, such as habitat occupancy of railway proximities, the use of structural elements (e.g., perching or nesting sites), flights over the railway, and behavior towards approaching trains. In this chapter, we present the most important results of several studies that were carried out on bird communities, between 2011 and 2015 on a 22-km stretch of HSR built on an agrarian landscape in central Spain. Available data describe the abundance and spatial distribution of birds up to 1000 m from the railway, bird infrastructure use (e.g., embankments, catenaries), cross-flights of the railway obtained through focal sampling, and animal responses to approaching trains recorded from train cockpits. These data depict how bird species respond at various scales to the presence of the HSR, and show how the infrastructure impacts bird communities, due to both habitat changes and increases in mortality risk.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
- Agrarian steppe
- Avian mortality
- Community homogenization
- Disturbance
- Environmental impact assessment
- Flight initiation distance
- Habitat degradation
- Road-kill
Introduction
The development of large-scale transport infrastructure impacts surrounding ecosystems. Environmental impact assessment is the formal procedure set up to predict, evaluate, and plan for the mitigation or compensation of such impacts in advance of project development (Petts 1999). Large projects impact various kinds of species; those that affect animal populations, especially vertebrates, are prioritized due to societal biases in conservation (Wilson et al. 2007). Nonetheless, the existing knowledge about bird responses to the presence and operation of railways is extremely scarce, and the data are even more scarce with respect to high speed railways (HSRs), due to their limited and relatively recent presence (Dorsey et al. 2015). Given the expected expansion of these railways in the near future (Campos and de Rus 2009; Todorovich et al. 2011; Fu et al. 2012), it is urgent that the environmental effects of this new technology be assessed to enable the development of mitigation strategies.
There is now reliable information on the impacts of roads on wildlife and on birds in particular (Benítez-López et al. 2010). We know that the effects are numerous, beginning with habitat loss generated by the construction of the infrastructure. However, the most serious impacts occur during the operating phase of roads, such as direct mortality of individuals and indirect impacts derived from the degradation of habitats close to roads. Similarly, railways alter bird communities both by destruction and degradation of habitats, as well as by lethal collision (Spencer 1965; Mammen et al. 2002). Due to the structural characteristics of HSRs (larger embankments, catenaries, etc.), their structural effects on habitats (and thus on species) are expected to exceed those of roads, while the degradation of habitats due to light, noise, and pollution, as well as mortality, are expected to be smaller due to lower traffic intensity.
In any case, the effects of HSRs on birds will depend upon the individual responses of species to the new landscape conditions. On the one hand, species will respond to the presence of structures that alter the natural physiognomy of the area, including the development of new habitats (e.g., revegetated embankments), and structural elements (e.g., catenaries, poles, bridges) that provide new potential perching and nesting sites (Tryjanowski et al. 2013; Morelli et al. 2014). On the other hand, the risk of mortality is restricted to birds that enter the narrow corridor where trains circulate, where the habitat has undergone extreme modification, from the gravel ballast where the rails lie to the catenary wires. To explain the impacts of future projects, it is crucial to know how birds respond to the presence of the railway.
This chapter summarizes the results gathered during the monitoring of birds in an agricultural area of the Iberian Peninsula that is crossed by a high-speed railway. We present a sequential approach that addresses the presence, abundance, and activity patterns of birds in the surroundings and on an HSR, complemented with data addressing the response of birds facing approaching trains obtained through cockpit video recordings.
Study Area
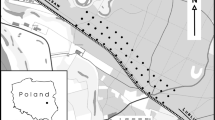
The study area is the HSR traversing Madrid-Castilla La Mancha-Valencia-Murcia, specifically on the high plateau (Mesa) of Ocaña. The selected section corresponds to km mileposts 80–102 of the HSR that runs through the municipalities of Noblejas, Villarrubia de Santiago, and Santa Cruz de la Zarza, in the northeast area of the Toledo province in central Spain (Fig. 8.1).

Location of study site along the Madrid-Albacete HSR. Inset image shows a magnified view of the study area (boxed-in main image)
The Mesa de Ocaña is a large flatland approximately 100 m above the Tagus River valley, with an average altitude between 750 m above sea level (a.s.l.) at the western end of the section, and 790 m a.s.l. on the eastern side. The climate is Mediterranean-continental, with an annual average temperature of 13.9 °C (average monthly minimum 7.4 °C, and maximum 20.5 °C), and an average annual rainfall of 437.8 mm (Ninyerola et al. 2005).
Of plain or slightly wavy relief, the area crossed by the railway is characterized by its open physiognomy, with a predominance of dry crops (mainly cereals, although legume crops like chickpeas, are frequently found), interspersed with olive crops and vineyards. In some areas north of the railway, the topography is somewhat more irregular, with ravines, hills, and hillocks where some patches of natural vegetation remain (oaks, pine trees, bushes, and Stipa tenacissima formations).
Part of the study area is included in or adjacent to natural protected areas under a conservation legal status. Notably, south of the railway, there are 10 polygons that constitute the NATURA2000 site “Áreas esteparias de la Mancha Norte,” which are especially important for the great bustard (Otis tarda) in the Castilla-La Mancha region. Approximately 60% of the birds in the region are found here.
Bird Occupation of Railway Surroundings
Spatial patterns of birds around the HSR give broad-scale information of infrastructure impacts on the animal community because the presence and abundance of bird species along the landscape mirrors their response to it. Thus, it is foreseeable that habitat destruction and degradation in the HSR vicinity will shape the way birds use the area (Benítez-López et al. 2010; Torres et al. 2011; Wiaçek et al. 2015). Comparisons of species abundance in close proximity to infrastructure locations with that in the wider landscape are useful for the assessment of infrastructure impacts (Torres et al. 2016). Ideally, to analyze human impacts, animal communities should be characterized before infrastructure construction as well as recurrently afterwards, during the operation phase, in a “before and after” control-impact design (Torres et al. 2011). However, this approach is frequently unfeasible and scientific quantification of changes in bird densities around roads has shown that most effects on birds extend up to 500 m from the infrastructure, with outlying areas largely free of impacts (Benítez-López et al. 2010).
In this case study, the characterization of bird populations potentially affected by the HSR line started before the operation phase, but after the line was already built. Thus, there is no information related to impacts associated with the construction phase. Nonetheless, control areas 500–1,000 m from the railway give a detailed picture of bird communities presumed to be negligibly affected by the presence of the project, and they are informative about local changes in bird communities by comparison with data obtained in close proximity to the HSR (Wiaçek et al. 2015). The first censuses were conducted in May of 2010, coinciding with the breeding season. Afterwards, the monitoring of bird populations took place during the first three years of operation (2011–2013), with censuses in both winter and spring. A final year-round monitoring with four seasonal samplings was conducted in 2014–2015. All of these censuses involved two complementary methodologies: (1) linear transects on foot, and (2) direct counts from a car along dirt paths.
The census of passerine birds and other small-sized species was performed by walking 1 km line-transects without a defined count bandwidth. Each transect was walked by a single experienced observer, and all birds identified–visually or acoustically–were recorded. This method is frequently used in extensive surveys to estimate abundance, distribution, and patterns of habitat selection by birds (Bibby et al. 2000). The transects were regularly distributed on both sides of the railway in two lines parallel to the HSR, one next to the fence to sample the bird community affected by the railway, and the second (control) at a distance of 500 m from it.
Direct counts of mid- and large-sized birds were conducted by car, as is common (Alonso et al. 2005; Morales et al. 2008). For these counts, the car was driven slowly along dirt tracks up to 1000 m from the railway, with a fixed detection band of 1000 m to each side of the vehicle. Regular stops were made to prospect the area thoroughly, including the surroundings of the railway and control sites on both sides. All morning, censuses were carried out within four hours of sunrise, and afternoon censuses were done in the 2 h before sunset. Censuses were done on days with good visibility and no rain, and hunting days were avoided.
In the landscape traversed by the HSR, the avian community was typical of that in the (Traba et al. 2013). Outstanding species typical of steppes include the great bustard, little bustard (Tetrax tetrax), and calandra lark (Melanocorypha calandra), together with others common in agricultural areas, including the spotless starling (Sturnus unicolor), rock dove (Columba livia), common magpie (Pica pica), sparrows (Passer spp., Petronia petronia), and several Fringilidae and Embericidae species.
In all, 80 bird species have been detected in the study area, reflecting the presence of a rich bird community, though 40–50% of the species that have been observed in the area were detected in fewer than 10 observations within each sampling period. Eleven species of high conservation interest were present in the area, including the black kite (Milvus milvus), which is nationally classified as “endangered” (Madroño et al. 2004). Species classified as “vulnerable” that were observed in the area include the great bustard, pin-tailed sandgrouse (Pterocles alchata), black-bellied sandgrouse (P. orientalis) and little bustard as well as Montagu’s harrier (Circus pygargus).
Regarding changes potentially introduced by the railway, it is noteworthy that total bird densities were reduced from approximately 550 birds/km2 in 2010–2011 to values in the range 360–390 birds/km2 range in the following years. This change occurred shortly after the trains began running, but there is no information from control sites remote enough to affirm that the decrease was directly associated with the railway rather than other regional-scale phenomena. Among the local bird species, some showed a constant growing trend in their densities over the 5-year study period, including the corn bunting (Emberiza calandra), house sparrow (Passer domesticus), rock dove, common magpie, and Eurasian skylark (Alauda arvernsis) (Table 8.1). Some species, such as the crested lark (Galerida cristata), showed consistently high average densities. Reductions in population density estimates over time were observed for the common quail (Coturnix coturnix), European serin (Serinus serinus), common linnet (Carduelis cannabina), meadow pipit (Anthus pratensis), calandra lark, Spanish sparrow (Passer hispaniolensis), and red-legged partridge (Alectoris rufa), among others.
Analysis of the spatial distribution of birds around the railway showed a differential use across the species (Table 8.1), though strong seasonal variability makes it difficult to detect the effects of HSR proximity. This relationship was negative for a number of species, including the calandra lark, crested lark, rock dove, and spotted starling. However, it was strongly positive for several species of finches (common linnet, European serin) that take advantage of the grassland abundance on HSR embankments. Others, such as the house sparrow, benefitted from their use of railway structures to nest. In parallel, a specific analysis of large- and mid-sized bird locations showed that both the great bustard and the little bustard significantly increased their distance (by 60–150 m) from the railway once trains started running. However, for most of the species, potential trends were obscured by seasonal and inter-annual variability.
In conclusion, the construction of the HSR resulted in a general decrease in bird density in the area, and modified the small-scale spatial patterns of the avian community. Thus, certain birds typical of these pseudo-steppe open landscapes, such as the little bustard, common quail, calandra lark, and red-legged partridge appeared to move away from the HSR location (Traba et al. 2013). On the other hand, the infrastructure has led to the presence and/or increase in the numbers of certain species, mostly anthropohiles, that use it as a substrate for nesting, resting, or feeding, such as, most notably, sparrows, starlings, rock doves, and magpies (Clavel et al. 2011). The many perching possibilities enabled by the HSR led to a wide use by corn buntings as well as by some raptors and corvids. On a narrower scale, some species were found to concentrate around the railway (common linnet, European serin, house sparrow), while others tended to avoid it (calandra lark, common chaffinch). It is noteworthy that there were some incongruous observations, such as species attracted to the railway or the embankments having decreased in numbers in the area (European serin, common linnet), while others that avoid the railway increased their abundance (Eurasian skylark, spotless starling, rock dove).
Bird Use of Railway Structures
Bird species behavior is an important factor in determining whether the HSR has a positive or negative impact on proximal population density. The mortality of birds depends on the extent to which they are exposed to the risk of being over-run while flying, or being electrocuted by the HSR poles or catenary. In contrast, the provision of new elements in the landscape (e.g., poles, bridges, embankments) may facilitate the presence and/or breeding of some species (Tryjanowski et al. 2013; Morelli et al. 2014). It is therefore interesting to elucidate how birds use track environs, because that use may be beneficial, such as for nesting, or detrimental, such as by producing a mortality risk. In a sense, the above-mentioned changes in the bird community may correspond to landscape scale impacts, al-though some impacts of the railway are associated with much more specific processes that take place on the infrastructure itself (Mainwaring 2015).
Data in this section were obtained from fixed observation stations and made seasonally between the autumn of 2014 and the summer of 2015. The sampling unit in this case was 10-min observations of all birds that make any use of a 120 m section of the HSR from the position of the observer. This distance corresponds to two spans between catenary poles, elements that aide in systematic data collection and define a distance that allows the identification of species of almost all observed individual birds. For each observed bird, the infrastructure elements used, and the time spent in them, were noted. Recorded elements were powerlines (catenary, power line and all other supporting wires), built structures (passages and bridges), ballast (stone and gravel track bed), catenary poles, fences, and embankments.
Sampling station locations were selected with the premise of concentrating on high embankments where birds have access to all the typical HSR elements and directly face the need to determine at what height to cross the railway (see below). In the Villarrubia de Santiago area, 26–28 sampling stations were distributed over approximately 3360 m. In the Santa Cruz de la Zarza area, 20 stations were distributed over 2400 m. Data from both sections were clumped together for the presentation of results. The infrastructure use by birds (nesting, rest, power, etc.) is expressed as: (1) the number of birds and (2) the time of use per kilometer of HSR during 1 h of observation (birds km−1 h−1).
In total, 936 birds, belonging to 30 species, were observed with an average of 81.8 birds and 404 min of use per kilometer of HSR observed for 1 h (Table 8.2). The most common species observed in the vicinity of the HSR were the spotless starling, common linnet, rock dove, house sparrow, rock sparrow, crested lark, and common magpie (all >5 birds km−1 h−1). No protected species was found using the sampled elements of the infrastructure.
The structures used most were embankments (24 species in total; 26.4 birds km−1 h−1) and powerlines (12 species; 25.2 birds km−1 h−1), although the numerical importance of the latter is due to frequent perching by the most abundant species—the spotless starling. In this species, 94.3% of the observations were on these elements. In order of importance, following embankments and powerlines, are fences (20 species; 11.3 birds km−1 h−1), built structures (5 species; 9.9 birds km−1 h−1), ballast (9 species; 5.9 birds km−1 h−1), and, finally, catenary poles (5 species; 3.0 birds km−1 h−1).
Table 8.3 summarizes HSR use by species observed more than 10 times. The embankments are used by most species as a feeding substrate and nesting place. Such nesting takes place both in bushes used for plant restoration as well as on the ground protected by them. For these species, embankments are islands with permanent and green vegetation year-round, contrary to the crop fields and fallows in the surrounding areas. The power lines are used as vantage and singing points by the corn bunting, in addition to the spotless starling. In the case of passages and bridges, the only note-worthy use was by the rock dove and the rock sparrow; both species rest and nest in holes within these structures. The ballast is only used by the crested lark, while the catenary poles are occupied as vantage points by the common magpie. It is noteworthy that during the monitoring presented in this chapter, it was discovered that these poles act as pitfall traps that cause the death of birds that fall inside (Malo et al. 2016). The spotless starling was the most affected species by this, but magpies and kestrels also suffered this fate. Finally, although the fences are used by many species, their use is only remarkable in the case of the house sparrow and European stonechat (Saxicola torquatus).
In conclusion, the HSR provides new structural elements for birds, which are used in accordance with the particular requirements of the species (Morelli et al. 2014). However, among the species that use these elements, some were found to present their maximum densities in close proximity to the railway (common linnet, house sparrow). More frequently, however, species were repelled by the railway (spotless starling, rock dove, crested lark). Therefore, the direct expectation that species that make an active use of the new opportunities provided by the HSR would increase their densities in proximity to the railway (Benítez-López et al. 2010), and in the area as a whole, was not fully met.
Bird Flight Over the Railway
In a sequential approach to the presentation of findings in the previous sections, to understand the potential magnitude of the HSR impact on birds, it is key to know to what extent flying birds face the risk of being overrun by circulating trains. Mortality from roadkill is one of the most serious direct impacts of operating transportation infrastructures (Loss et al. 2014). The potential relevance of the HSR is conditioned by the fact that the great speed of the trains, in most cases, precludes birds from avoiding train collisions (DeVault et al. 2015). If birds could fully avoid running trains by flying above their height, the railway would remain responsible only for effects on habitats and for increases in energy expenditure by individuals that cross it in flight.
The collision risk area associated with the HSR corresponds basically to the section framed by the catenary and the rails. In a simplified view, for twin tracks, it corresponds to a rectangle with an 8 m-wide base that is 5.3 m high (height of the power wire) where the trains run. Additionally, it is hazardous for birds to fly above the train collision risk area where they may come in contact with the catenary, suspenders, power wire, feeder, earth cable, and tensors (5.3–8.5 m above the ground). Although there is a constant risk of collision with these elements even when trains are not running, the risk of collision with the catenary may be increased by the passage of trains due to the potential for turbulence generated by the moving train to destabilize the normal flight of birds. Thus, it is reasonable to assume that birds that cross the railway by flying between or below the catenary wires face the mortality risk from train-kill.
In the HSR stretch analyzed here, surveys of birds flying over the infrastructure were carried out between 2010 and 2015, with a total of approximately 270 h of observation at nearly 1100 observation points. Sampling structure varied, depending on the objectives of particular studies, but in all cases observations were made from fixed stations for 10 min or 20 min. Bird flight was recorded from these points, with HSR crossing flight height estimated with respect to the rails. Therefore, for all crossing events over the railway (N = 3313 records and 10,776 individuals), it is known whether the birds crossed within the collision risk area (train path or catenary), allowing a risk index to be calculated for each species. Additionally, data obtained within 120 m of the observer have been used to estimate HSR crossing rates throughout the year (birds h−1 km−1). Furthermore, spatial and temporal sampling patterns are informative for elucidating interactions of various factors, such as whether track crossover frequency is independent of HSR geometry, astronomical season, or environmental conditions such as wind speed. Complementarily, data available on birds from surrounding areas, such as parallel sampling of flight behavior, can be used to determine whether birds change their usual flight trajectories in the presence of the HSR. To ensure consistency of the results, the data presented are restricted to species for which, in at least one sampling station, 10 crossing records were obtained.
Our analysis showed that birds cross the HSR frequently, with an average HSR crossing frequency of 246.7 birds km−1 h−1 (experiment range, 160.2–514.0 birds km−1 h−1). No consistent seasonal pattern was observed across the trials. However, our data analysis did reveal alterations in bird flight patterns in the vicinity of the HSR. An experiment conducted between June and July of 2011 at 237 observation stations along the HSR and 85 remote control sites showed that birds of any species fly, on average, almost 5 m higher near the HSR than in control sites (9.9 vs. 5.2 m over the ground). This increase in flight heights reflects some avoidance of the infrastructure, with an added energy cost. The average flight height of birds over the HSR was lower in areas where the railway runs along embankments relative to sections of railway that are level with the surrounding landscape. In fact, several experiments showed that birds reduced their crossing railway height between 1 m and 4 m for every 10 m of embankment height. However, these general patterns were found to vary greatly across different species, and all of them combine observations of birds both inside and outside the cross-section of collision risk.
Although the birds fly higher over the infrastructure than over an open field, the frequency with which they cross the railway within the risk area is high. For the 33 analyzed species, almost half of railway crossings (combined average, 46.5%) occurred less than 8.5 m above the ground (Table 8.4). More than two-thirds of these low crossings were under the catenary, and the remainder were through the catenary wires. In fact, birds of 21 species crossed within (rather than over) the risk area for at least 33% of their HSR crossings, and 14 species did so for at least 50%.
The partridge (100%), Eurasian tree sparrow (Passer montanus, 90%), hoopoe (Upupa epops, 88.5%), and northern wheatear (Oenanthe oenanthe, 83%) stand out as species with high-frequency risks of colliding with running trains. Conversely, the great bustard (0.0%), black kite (5.7%), common swift (Apus apus, 5.9%), wood pigeon (Columba palumbus, 8.7%), and great cormorant (Phalacrocorax carbo, 8.1%) showed low-risk flying behavior. However, the fact that 12 great bustard carcasses have been observed within the HSR study area indicates that great bustards must fly under or through the catenary at least occasionally (unpublished data, LIFE+ Impacto 0 2016). The collision risk was elevated on embankments, with the risk of collisions in one study being 14% greater on embankments over 5 m high than in nearby flat sections of the HSR. Birds from virtually all of the protected species present in the area crossed the risk area at least occasionally. Notably, 40.0% of little bustard crossings, 28.6% of lesser kestrel (Falco naumanni) crossings, and 24.3% of Montagu’s harrier crossings were observed to be below 8.5 m.
Finally, another important factor that determines the height of flight over the HSR is wind. Because birds tend to fly lower in the presence of strong winds, percentages of railway crossings through the risk area increase under windy conditions. The presence of fog would be expected to affect birds’ mortality risk in the vicinity of the HSR as well. However, foggy days were not sampled specifically. Generally, the potential influence of atmospheric conditions on bird behavior with respect to the HSR should be clarified to better understand the risk faced by birds crossing the railway.
In conclusion, although birds show some avoidance of the railway and catenary, many birds fly within the collision risk area. This risky behavior is also engaged in by protected species, although it is especially prevalent among species that make use of specific elements of the railway (e.g., common linnet, European serin, Eurasian skylark). There are also some species, such as the red-legged partridge and hoopoe, that tend to fly close to the ground naturally and thus face a high risk of collision with trains and the catenary.
Bird Behavior Towards Approaching Trains
A more detailed scale of bird interactions with the HSR is the behavior of individual birds facing an approaching train. Note that if birds are able to react before the arrival of a train, even high rates of crossing the risk area would be harmless. However, it is methodologically complex to analyze this issue. Data presented in the previous sections were in all cases collected with traditional direct observation methodologies, which involve time-space limitations (e.g., length of sections under scrutiny, difficulty or impossibility of accessing the infrastructure, and frequency of trains) that limit data quantity and quality (Wells et al. 1999). Such limitations are especially noticeable for relatively rare animals and exacerbated by the difficulty of observing the process in real time, given the velocity of HSR trains (Rodríguez et al. 2008).
To overcome these limitations, data in this section were obtained in the framework of a project currently under development (LIFE+ Impacto 0 2016) that uses high-speed video recording from the cockpits of running trains. This study is the first time that this method was applied in a railway study, previous studies used direct observations by train conductors (SCV 1996; SEO/BirdLife 1997; Wells et al. 1999), and video recordings have begun to be used to assess the interactions of wildlife with powerlines (Carlton and Harness 2001), wind turbines (Desholm et al. 2006; Cryan et al. 2014), airplanes (Doppler et al. 2015), and cars (Legagneux and Ducatez 2013; DeVault et al. 2015). Cockpit video data provide accurate information on location, time, conditions (i.e., speed of the train, type of stretch), and the behavior (i.e., distance of flight, type of reaction) of animals detected in front of the vehicle in events of crossing, infrastructure use, and collision. Also, observations can be made continuously along the train path for tens or even hundreds of kilometers, and include sections inaccessible to walking personnel due to viaducts and tunnels. Hence the cockpit video approach increases the number and types of observations that can be made.
Video recordings included in the present analysis were made with forward-facing, high-speed cameras (120–240 frames per second, GoPro Hero 3+ Black Edition) through the cockpit windshield. A technician located in the cabin operated the recording equipment together with GPS register systems and recorded all bird observations (perching, crossings, collisions) on standardized forms. A series of basic data were thus used to simplify later analysis (e.g., recording time, relative size or bird species, position in the image, type of observation). These forms served as a reference for the systematic review of recordings, focusing attention on direct observations. Technicians’ observations, species, numbers of individuals, and behaviors were confirmed in the videos. In parallel, data from the navigation system, such as train speed, location (geographic coordinates and mile mark), stretch configuration, and surrounding habitat were extracted. High-speed recording allowed repeated and frame-to-frame review of each event to facilitate the analysis of the observations.
The train trips with high speed recording were between the Madrid and Albacete stations (321.7 km) of the Madrid-Levante line, which includes the section in which all field experiments were developed. A total of 66 trip recordings were collected over different seasons (14–20 per season). There were five additional trips during which the technical equipment was set up and tested (data not included). In all, 59 h and 55 min of recording (average trip, 55.3 min) along 14,700 km of accumulated train motion (average trip, 226.2 km; range: 78.3–288.7 km) were analyzed.
These recordings yielded 1090 confirmed bird observations, including 39 collisions (3.6% of observations), yielding a mortality risk of 0.0026 killed birds per km (1 hit per 406.1 km). Direct observation from the cockpit provided a new perspective in the analysis of the reactions of birds facing an approaching train. Train speed during bird collisions (mean ± standard deviation, 265.8 ± 39.2 km/h; range 175–305 km/h; N = 20) was similar to that during under-catenary crossings (251.5 ± 58.9 km/h; range 0–305 km/h; N = 183). However, in all cases, bird collisions occurred with trains travelling at high speeds. Collision rates varied seasonally, consistent with prior observations (SEO/BirdLife 1997; Frías 1999; Carvalho and Mira 2011; Bishop and Brogan 2013; Loss et al. 2014). The roadkill rate varied between 0.0018 and 0.0032 birds/km, with a noteworthy constant percentage of under-catenary crossings in front of the train resulting in collisions (~12.2 ± 0.01%). These data suggest that train speed determines mortality risk independent of particular species’ characteristics, with the risk being a result of the fact that birds are not adapted to avoiding objects approaching at such high velocities (Martin 2011; DeVault et al. 2015; Lima et al. 2015). Previous studies have pointed out that birds have difficulty avoiding vehicles travelling over 80–90 km/h (Pallag 2000; DeVault et al. 2014, 2015). The lowest speeds of collision recorded in our study were ~180 km/h, similar to the findings of DeVault et al. (2015), who suggested that such speeds exceed birds’ abilities to escape successfully.
Given these data and the infeasibility of reducing train speeds, information obtained from recordings with respect to birds’ infrastructure use and behavior when facing a train, particularly in cases of collision, is of great interest. Of all recorded birds, 29.4% flew below the catenary (i.e. through the risk area), a datum similar to that registered in previous field studies (33.1, see above section). Cockpit recordings enabled us to determine that 37.7% of the birds that crossed the risk area were using some element of the infrastructure moments before their train encounters. That is, they commenced flight (most likely) due to the approach of the train, but crossed in front of it. Among the data obtained from the train, it is remarkable that 40.1% of the detected birds were initially observed in the infrastructure, and 28% of them crossed below the catenary. In addition, 5 of 29 collisions recorded from the cockpit (17.2%) involved birds resting on or in the infrastructure. Therefore, a high percentage of the birds that are finally exposed to the risk of collision were using the railway as a place to rest or feed. Hence, corrective measures, such as barriers designed to raise the flight of birds above the catenary (LIFE+ Impacto 0 2016), should also deter birds from HSR structures in order to reduce bird mortality risk in the HSR.
On the other hand, our analysis of the flight initiation distance of different groups of birds showed that they do not seem to respond to trains as if they were predators, but rather human-associated elements (Møller et al. 2013; Neumann et al. 2013). Thus, species that are more tolerant of people or even anthropophilic (e.g., passerines and pigeons) show flight initiation distances that are significantly shorter than those of crows and birds of prey, which are more predatory (Table 8.5). Some species that are frequently observed around railways (e.g., the crested lark, doves, and pigeons) do not take flight in response to incoming trains, putting themselves at risk of being hit or dragged by the turbulence of the passing train.
In short, it is evident that at least some of the birds living in the surroundings of the HSR do not treat trains as a natural threat, and exhibit some habituation to HSR train circulation. However, HSR train speed often proves fatal when birds enter the collision risk area. Both factors could explain the high mortality rates near the HSR relative to rates near other linear infrastructures, though further research on other HSR stretches should be done to determine if this pattern of findings is representative of broader HSR areas.
Conclusions
Our results provide a first picture of the avian response to the presence and operation of HSR trains. Although the observed trends are not particularly prominent and are diluted by seasonal and inter-annual variability, they are consistent with the notion that train circulation could be responsible for a decrease in bird abundance in the traversed area. Accordingly, it can be concluded that the presence of the HSR has led to a process of ruderalization characterized by the increased numbers of species taking advantage of the railway. Some bird species representative of open and agricultural areas were found to avoid the HSR, whereas species that use man-made structures were found to be attracted to it. In fact, we documented the active use of these new elements, mainly the embankments and the catenary, by birds. In addition, birds alter their flight patterns near the railway, including showing some avoidance of it. However, birds risk train collision because such avoidance is not complete. Moreover, the species that are most exposed to train collisions are those that respond favorably to the railway and use its associated elements. The cockpit recordings confirmed bird habituation to moving trains and that some portion of over-run specific birds were actively using the railway moments before they faced the approaching train. Nevertheless, the flight characteristics of each species are also a determinant, and species that fly close to the ground are exposed to a disproportionate collision risk even if they do not make active use of the railway. These findings led us to conclude that changes in the community of birds in the surroundings of the HSR are relevant and related to structural landscape changes and the responses of bird species to the new infrastructure. In addition, bird mortality may have a significant impact on the populations of some species, given the fact that birds cross the collision risk area frequently. The results may be contentious from an environmental impact perspective given the limited opportunities for mitigation (Rodríguez et al. 2008). Firstly, changes in the avian community due to the presence of the HSR and the use of HSR structures are probably difficult to avoid because birds use elements that are fundamental to the infrastructure, thus, only minor design changes can be implemented to minimize HSR impacts on birds. Secondly, even if the installation of some devices can reduce flight paths across the railway, their implementation will be feasible only in short stretches of high ornithological interest due to cost limitations. Finally, reducing train speed in poder to reduce the risk of collisions with birds would conflict with the core objective of a HSR, making such a strategy again unlikely along long stretches of railway.
References
Alonso, J. C., Palacín, C., & Martín, C. A. (Eds.). (2005). La avutarda común en la Península Ibérica: población actual y método de censo. Madrid: SEO/BirdLife.
Benítez-López, A., Alkemade, R., & Verweij, P. A. (2010). The impacts of roads and other infrastructure on mammal and bird populations: A meta-analysis. Biological Conservation, 143, 1307–1316.
Bibby, C. J., Burgess, N. D., & Hill, D. A. (2000). Bird census techniques (2nd ed.). London: Academic Press.
Bishop, C. A., & Brogan, J. M. (2013). Estimates of avian mortality attributed to vehicle collisions in Canada. Avian Conservation and Ecology, 8, 2.
Campos, J., & de Rus, G. (2009). Some stylized facts about high-speed rail: A review of HSR experiences around the world. Transport Policy, 16, 19–28.
Carlton, R. G., & Harness, R. E. (2001). Automated systems for monitoring avian interactions with utility structures and evaluating the effectiveness of mitigative measures. In: Power Engineering Society winter meeting (Vol. 351, pp. 359–361). IEEE.
Carvalho, F., & Mira, A. (2011). Comparing annual vertebrate road kills over two time periods, 9 years apart: A case study in Mediterranean farmland. European Journal of Wildlife Research, 57, 157–174.
Clavel, J., Julliard, R., & Devictor, V. (2011). Worldwide decline of specialist species: Toward a global functional homogenization? Frontiers in Ecology and the Environment, 9, 222–228.
Cryan, P. M., Gorresen, P. M., Hein, C. D., Schirmacher, M. R., Diehl, R. H., Huso, M. M., et al. (2014). Behavior of bats at wind turbines. Proceedings of the National Academy of Sciences, 111, 15126–15131.
Desholm, M., Fox, A. D., Beasley, P. D. L., & Kahlert, J. (2006). Remote techniques for counting and estimating the number of bird-wind turbine collisions at sea: A review. Ibis, 148, 76–89.
DeVault, T. L., Blackwell, B. F., Seamans, T. W., Lima, S. L., & Fernández-Juricic, E. (2014). Effects of vehicle speed on flight initiation by turkey vultures: Implications for bird-vehicle collisions. PLoS ONE, 9, e87944.
DeVault, T. L., Blackwell, B. F., Seamans, T. W., Lima, S. L., & Fernández-Juricic, E. (2015). Speed kills: Ineffective avian escape responses to oncoming vehicles. Proceedings of the Royal Society of London B, 282, 20142188.
Doppler, M. S., Blackwell, B. F., DeVault, T. L., & Fernández-Juricic, E. (2015). Cowbird responses to aircraft with lights tuned to their eyes: Implications for bird–aircraft collisions. The Condor, 117, 165–177.
Dorsey, B., Olsson, M., & Rew, L. J. (2015). Ecological effects of railways on wildlife. In R. van der Ree, D. J. Smith, & C. Grilo (Eds.), Handbook of road ecology (pp. 219–227). West Sussex: Wiley.
Frías, O. (1999). Estacionalidad de los atropellos de aves en el centro de España: número y edad de los individuos y riqueza y diversidad de especies. Ardeola, 46, 23–30.
Fu, X., Zhang, A., & Lei, Z. (2012). Will China’s airline industry survive the entry of high-speed rail? Research in Transportation Economics, 35, 13–25.
Legagneux, P., & Ducatez, S. (2013). European birds adjust their flight initiation distance to road speed limits. Biology Letters, 9, 20130417.
LIFE+ Impacto 0. (2016). Development and demonstration of an anti-bird strike tubular screen for High Speed Rail lines. LIFE+ Biodiversity project (LIFE12 BIO/ES/000660). http://www.lifeimpactocero.com. Accessed October 14, 2016.
Lima, S. L., Blackwell, B. F., DeVault, T. L., & Fernández-Juricic, E. (2015). Animal reactions to oncoming vehicles: A conceptual review. Biological Reviews, 90, 60–76.
Loss, S. R., Will, T., & Marra, P. P. (2014). Estimation of bird-vehicle collision mortality on U.S. roads. Journal of Wildlife Management, 78, 763–771.
Madroño, A., González, A., & Atienza, J. C. (2004). Libro Rojo de las aves de España. Dirección General para la Biodiversidad-SEO/BirdLife, Madrid.
Mainwaring, M. C. (2015). The use of man-made structures as nesting sites by birds: A review of the costs and benefits. Journal of Nature Conservation, 25, 17–22.
Malo, J. E., García de la Morena, E. L., Hervás, I., Mata, C., & Herranz, J. (2016). Uncapped tubular poles along high-speed railway lines act as pitfall traps for cavity nesting birds. European Journal of Wildlife Research, 62, 483–489.
Mammen, U., Klammer, G., & Mammen, K. (2002). Greifvogeltod an Eisenbahntrassen—ein unterschaetztes problem [Dead of raptors at railways—An underestimated problem]. Paper presented at the 5. Internationales Symposium “Populationsökologie von Greifvogel- und Eulenarten” [“Population Ecology of Raptors and Owls”], Meisdorf/Harz.
Martin, G. R. (2011). Through birds’ eyes: Insights into avian sensory ecology. Journal of Ornithology, 153, 23–48.
Møller, A. P., Grim, T., Ibáñez-Álamo, J. D., Markó, G., & Tryjanowski, P. (2013). Change in flight initiation distance between urban and rural habitats following a cold winter. Behavioral Ecology, 24, 1211–1217.
Morales, M. B., Traba, J., & Carriles, E. (2008). Sexual differences in microhabitat selection of breeding little bustards Tetrax tetrax: Ecological segregation based on vegetation structure. Acta Oecologica, 34, 345–353.
Morelli, F., Beim, M., Jerzak, L., Jones, D., & Tryjanowski, P. (2014). Can roads, railways and related structures have positive effects on birds?—A review. Transportation Research Part D Transport and Environment, 30, 21–31.
Neumann, W., Ericsson, G., Dettki, H., & Radeloff, V. C. (2013). Behavioural response to infrastructure of wildlife adapted to natural disturbances. Landscape Urban Plan, 114, 9–27.
Ninyerola, M., Pons, X., & Roure, J. M. (2005). Atlas Climático Digital de la Península Ibérica. Metodología y aplicaciones en bioclimatología y geobotánica. Bellaterra: Universidad Autónoma de Barcelona.
Pallag, O. (2000). COST 341. Habitat fragmentation due to transportation infrastructure. Hungarian State of the Art Report. Technical and Information Services on National Roads (ÁKMI), Budapest.
Petts, J. (Ed.). (1999). Handbook of environmental impact assessment (2 Vols.). Oxford: Blackwell Science.
Rodríguez, J. J., García de la Morena, E. L., & González, D. (2008). Estudio de las medidas correctoras para reducir las colisiones de aves con ferrocarriles de alta velocidad. Ministerio de Fomento, CEDEX, Madrid.
S. C. V. (1996). Mortalidad de vertebrados en líneas de ferrocarril. Documentos Técnicos de Conservación SCV. Sociedad Para la Conservación de los Vertebrados, Madrid.
SEO/BirdLife. (1997). Mortalidad de aves en un tramo de línea de ferrocarril. SEO/BirdLife, Grupo local SEO-Sierra de Guadarrama.
Spencer, K. J. (1965). Avian casualties on railways. Bird Study, 12, 257.
Todorovich, P., Schned, D., & Lane, R. (2011) High-speed rail. International Lessons for U.S. Policy Makers. Lincoln Institute of Land Policy, Cambridge, Massachusetts, USA.
Torres, A., Jaeger, J. A. G., & Alonso, J. C. (2016). Assessing large-scale wildlife responses to human infrastructure development. Proceedings of the National Academy of Sciences, 113, 8472–8477.
Torres, A., Palacín, J., Seoane, J., & Alonso, J. C. (2011). Assessing the effects of a highway on a threatened species using Before-During-After and Before-During-After-Control-Impact designs. Biological Conservation, 144, 2223–2232.
Traba, J., Sastre, P., & Morales, M. (2013). Factors determining species richness and composition of steppe bird communities in Peninsular Spain: Grass-steppe vs. shrub-steppe bird species. In M. Morales & J. Traba (Eds.), Steppe ecosystems: Biological diversity, management and restoration (pp. 29–45). New York: Nova Publishers.
Tryjanowski, P., Sparks, T. H., Jerzak, L., Rosin, Z. M., & Skórka, P. (2013). A paradox for conservation: Electricity pylons may benefit avian diversity in intensive farmland. Conservation Letters, 7, 34–40.
Wells, P., Woods, J. G., Bridgewater, G., & Morrison, H. (1999). Wildlife mortalities on railways: Monitoring methods and mitigation. In Proceedings of the Third International Conference on Wildlife Ecology and Transportation. ICOET, Missoula, Montana.
Wiaçek, J., Polak, M., Filipiuk, M., Kucharczyk, M., & Bohatkiewicz, J. (2015). Do birds avoid railroads as has been found for roads? Environmental Management, 56, 643–652.
Wilson, J. R. U., Proches, S., Braschler, B., Dixon, E. S., & Richardson, D. M. (2007). The (bio)diversity of science reflects the interests of society. Frontiers in Ecology and the Environment, 5, 409–414.
Acknowledgments
Jorge Hernández Justribó, Mª José Pérez Sobola, Diego Sánchez Serrano, and safety personnel from Centro de Estudios y Experimentación of the Ministry for Public Works (CEDEX) and Ineco helped with fieldwork. Staff from CEDEX and Adif provided kind help with project logistics. This study was carried out as part the LIFE+ Impacto 0 research project (LIFE 12 BIO/ES/000660) and a research agreement between UAM and CEDEX; both focused on the mitigation of High Speed Railway impacts on bird populations. The Comunidad de Madrid, together with the European Social Fund, supports the Terrestrial Ecology Group research group through the REMEDINAL-3 Research Network (S-2013/MAE-2719).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2017 The Author(s)
About this chapter
Cite this chapter
Malo, J.E., García de la Morena, E.L., Hervás, I., Mata, C., Herranz, J. (2017). Cross-scale Changes in Bird Behavior Around a High Speed Railway: From Landscape Occupation to Infrastructure Use and Collision Risk. In: Borda-de-Água, L., Barrientos, R., Beja, P., Pereira, H. (eds) Railway Ecology. Springer, Cham. https://doi.org/10.1007/978-3-319-57496-7_8
Download citation
DOI: https://doi.org/10.1007/978-3-319-57496-7_8
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-57495-0
Online ISBN: 978-3-319-57496-7
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)