
Abstract
Small organisms provide the bulk of biodiversity. Here, we look at rewilding from their perspective. As an umbrella group for other terrestrial invertebrates, we focus on the diverse group of Lepidoptera. More specifically, we set out to explore their response to farmland abandonment. So far, studies have warned against farmland abandonment, which is for instance listed as one of the key threats to European butterfly diversity. Here, partly based on a case study within the Peneda mountain range, we argue (i) that the majority of Lepidoptera is to a greater or lesser extent forest-dependent, (ii) that effects on species composition should be considered at regional rather than smaller scales, and (iii) that habitat resource heterogeneity at multiple spatial scales is key. As such, we believe that rewilding does offer opportunities to Lepidoptera. However, we recommend rewilding not to be equalled to a hands-off approach, but rather to a goal-driven conservation management approach. It should monitor, and where necessary intervene to provide habitat heterogeneity at multiple spatial scales, in order to cater for the whole gradient of sedentary to mobile species. Given that sufficient levels of habitat heterogeneity are maintained, Lepidoptera are one of probably many taxa that are likely to benefit from rewilding processes on European marginal farmland. The resulting improved species composition will help achieve European species conservation targets. It may also lead to more viable populations of moths, butterflies and other invertebrates, which will foster more resilient food-webs and increased ecosystem functioning.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
1 Rewilding Small-Sized Biodiversity Too
So far, the debate on rewilding opportunities for biodiversity has been mainly centred upon popular and hence large-sized taxa, such as large mammals and birds (see Chaps. 1, 4, 5, and 8). As home range extent is typically mirrored by organismal size, the relatively high mobility and large spatial footprint of large-sized taxa are mainly situated at the extreme end of a whole gradient. The bulk of biodiversity is smaller, less mobile, and hence operates at smaller spatial scales . Because of the considerable dimension of the amount of European agricultural land that is already being abandoned, and that is set to be abandoned over the next couple of decades (see Chap. 1), it is obvious why the rewilding concept provides a most welcome opportunity for wide-ranging and cursorial species, like wolves for example, whose ecology has simply not been compatible with the typically small nature reserves and the intensified countryside of Western Europe. However, for rewilding to be adopted as a credible land-use option and conservation strategy, it will need to provide more than only a handful of iconic large animals. The successful uptake of the rewilding approach may depend upon three main points: (i) rewilding will need to make (socio-)economic sense (see Chap. 10), (ii) ample supply of ecosystem services will need to be guaranteed (see Chap. 3), and last but not least, (iii) rewilding will need to make sense from a biodiversity conservation viewpoint too. As such, it needs to cater for all kinds of biodiversity, i.e. for rare, range-restricted and ubiquitous species, for generalists and specialists, for currently threatened and least-concern species, for species operating at all kinds of spatial scales. With regard to this latter point, we here look at rewilding from the perspective of smaller-sized taxa. Although these taxa provide the bulk of biodiversity and ecosystem functioning, they are severely under-represented in conservation research (Clark and May 2002; Cardoso et al. 2011; Pereira et al. 2012). As an (incomplete) umbrella group for other terrestrial invertebrates (Thomas 2005), we focus on the ecologically diverse group of Lepidoptera .
2 European Lepidoptera: Numbers and Trends
Lepidoptera are scale-winged and almost exclusively phytophagous insects, representing a mega-diverse radiation, probably correlated with the great diversification of flowering plants since the Cretaceous (Menken et al. 2010). This major insect order has been divided traditionally into the day-flying butterflies and largely nocturnal moths . In Europe, the order currently contains close to 9900 recorded species (www.lepidoptera.pl), of which 482 species (i.e. ca. 5 %) are butterflies. About a third of these butterfly species have currently declining populations on a European scale, and 9 % are threatened (van Swaay et al. 2010). In some European regions these figures are far higher; in Flanders, for example, 19 out of 67 resident butterfly species (28 %) went extinct since the start of the twentieth century, whilst 25 species (37 %) are currently threatened (Maes et al. 2013). Such high proportions can be explained by cumulative effects of environmental pressures due to a long history of economic development (Dullinger et al. 2013). European-wide declines are especially worrisome since Lepidoptera provide many vital and economically important services within terrestrial ecosystems, such as nutrient cycling, prey resources and pollination. The European Red List for butterflies identifies the main drivers for these declines as habitat (connectivity) loss and degradation due to agricultural intensification and the invasion of shrubs and trees resulting from farmland abandonment (van Swaay et al. 2010).
Moth trends have not yet been evaluated at the European scale, although national trends are available for a handful of countries. For example, the very recent assessments in Britain and The Netherlands show a picture similar to butterflies. Across Britain, overall abundance of macro-moths declined by 28 % over a recent 40-year period, with total numbers having decreased by 40 % in the more populated southern half of Britain. Two-thirds of common and widespread macro-moth species for which national population trends were calculated, decreased in abundance, with 61 species having declined by 75 % or more over 40 years (Fox et al. 2013). The preliminary Red List for macro-moths of The Netherlands (841 species) shows that 70 species (8 %) went extinct since the nineteenth Century and that 300 species (35 %) are currently threatened (Ellis et al. 2013). The decreasing extent of habitat and the degradation of its quality, more specifically via agricultural intensification, changing woodland management , urbanisation, climate change and light pollution , are likely causes of the observed changes in moth biodiversity (Fox et al. 2013).
3 Lepidoptera: Diurnal and Nocturnal Life-Styles
Thus, whilst farmland abandonment is generally reported to be relevant with regard to butterfly declines, moth declines are rarely linked to farmland abandonment. This observation makes sense if we consider their contrasting life-histories. Butterflies are day-flying ectotherms that need direct sunlight in order to raise their flight muscle temperature to optimal levels, most often well above ambient temperatures. By contrast, most moths are nocturnal and endothermic, raising their body temperatures above ambient levels by generating internal heat via muscle activity. As such, most butterflies and (ectothermic) day-flying moths occur typically in open, sun-lit habitats, whereas a majority of moths are to a greater or smaller degree linked to wooded habitats. For example, even in a largely deforested region (woodland cover < 5 %) of Flanders, 58 % of in total 499 macro-moth species observed in 1980–2012 use shrubs or trees as foodplants (Sierens and Van de Kerckhove 2014). However, the situation is far from black and white as some butterflies are woodland specialists and many nocturnal moths require more open habitat conditions. Still, based on their contrasting thermoregulatory requirements it is clear why—in general—butterflies are perceived to be more susceptible than moths to farmland abandonment, which is typically accompanied by scrub and forest encroachment, shading out formerly sun-lit biotopes (van Swaay et al. 2010).
Although day-flying Lepidoptera are numerically the exception to the nocturnal norm, sound conservation strategies need to be inclusive of both. For example, in a context of temperate landscapes under intense human land-use, Merckx et al. (2012a) recommend zoned woodland management for the effective conservation of both life-history strategies. Their research showed that the late-successional deciduous woodland biotope is characterised by high numbers of both individuals and species of moths, being especially important for some scarce and specialist species of conservation concern (see also Baur et al. 2006), while coppicing and ride widening, which open up dense forest structures, are valuable woodland conservation tools for Lepidoptera species with an affinity for more open biotopes (see also Fartmann et al. 2013). The mechanism behind the pattern of increased lepidopteran species richness at the woodland-scale, due to such zoned management, involves an increased structural and micro-climatic diversity, and, more generally, increased habitat resource diversity (Merckx et al. 2012a). The importance of habitat heterogeneity at such larger scales is further highlighted by the results obtained from a recent study on butterfly richness of semi-natural meadows in Estonia; using a sample of 22 meadows with a total of 56 butterfly species, the research showed a positive correlation between butterfly diversity on local meadows and forest cover in the landscape directly surrounding (i.e. 250 m radius) these meadows, whilst meadow cover in the surrounding landscape at various spatial scales actually impacted the butterfly diversity of these meadows in a negative way (Ave Liivamägi pers. comm.). For the Mediterranean region too, Verdasca et al. (2012) showed that whilst regular management (i.e. removing understory vegetation) in oak stands has a positive effect on butterfly assemblages, undisturbed stands are nevertheless needed by some butterfly species. A study by Baur et al. (2006) on the abandonment of subalpine semi-natural grasslands in Transylvania, Romania, found that whilst vascular plants reached highest species richness in yearly-mown hay meadows, diurnal Lepidoptera were actually most species-rich in meadows abandoned for three years or more (see also Dover et al. 2011 and references therein), and nocturnal Lepidoptera and Gastropoda were most species-rich in young (20–50 years) and mature (50–100 years) forests, respectively. Because the complementarity of species composition increased with successional age in all four taxonomic groups, and because the proportion of red-listed nocturnal Lepidoptera increased with successional age too, their results indicate the high conservation value of all stages of grassland succession, and especially so the later seral stages up to mature woodland. Hence, all these studies highlight that although semi-natural open biotopes may locally reach high diversity levels (for some taxa), their partial abandonment at the landscape-scale has a role to play for biodiversity in general.
4 Conservation Objectives: Semi-Natural Biotopes Versus Rewilding
Conservation objectives are the subject of much debate for regions with a long history of human alteration, like in most of Europe (Merckx et al. 2013). Climax forests have been replaced, often millennia ago, by so-called semi-natural biotopes, which are essentially different versions of early- to mid-successional natural seral stages, arrested from developing towards mature woodland. Nowadays, only scattered fragments of ancient woodland remain, and these have suffered continuous but varying disturbance regimes by humans. For example, up to a century ago most European woodland was maintained as coppice or coppice-with-standards. Today much woodland plantation has a uniformly closed canopy, which is shadier than is found in ancient forests with little history of human disturbance. A consequence of these various ways by which the development towards climax forest has been arrested, is that the rarest types of arboreal habitats in west-European countries today are those associated with rotting wood on ancient trees. These saproxylic habitats are associated with high extinction rates (Hambler et al. 2011), and support numerous invertebrate specialists, especially beetles and flies, and some moths (Thomas et al. 1994). Although many species undoubtedly suffered great losses (and extinctions) over the centuries during the transition from natural forest to semi-natural habitats, others (e.g. species associated with early-successional stages) did benefit or managed to adapt successfully to these semi-natural biotopes (e.g. heaths, meadows, coppiced woodland) (Young 1997; Monbiot 2013).
Since the 1950s, agricultural intensification, forestry intensification, urbanization, and farmland abandonment have all severely decreased such semi-natural biotopes in quantity, quality, and connectivity, and with them their specialist fauna and Lepidoptera. As a result, most European ‘conservationists’ traditionally seek to sustain or restore semi-natural biotopes, and do so by maintaining very specific disturbance regimes, often simply by copying traditional agricultural practices, since most large wild herbivores were excluded centuries ago (New 2009). Nevertheless, popular management operations to influence vegetation structure, such as burning and grazing/mowing, need careful planning (mainly to ensure refugia) as they may destroy much of the existing invertebrate fauna if applied too intensively, too infrequently, on too large a scale or at unsuitable times of year (New 2009). Hence, although wrongly applied conservation management may have unintended negative consequences, management is seen as a good thing overall, whereas rewilding is often perceived as a threat (Merckx et al. 2013).
Abandonment of human disturbance does indeed pose a threat to many specialist species that have become adapted to certain semi-natural biotopes, especially when they have nowhere else to go because natural succession dynamics are currently too disturbed and suitable natural patches are too small and/or isolated. On the other hand, the recovery of native forest ecosystems due to farmland abandonment is likely to benefit a majority of Lepidoptera, since most moths (which make up 95 % of Lepidoptera) are reliant on woody foodplants or wooded biotopes. Forest recovery will obviously favour endangered specialist faunal groups too (e.g. closed-woodland Lepidoptera, Gastropoda, and saproxylic groups) .
5 Controlled Rewilding: Reconciling the Objectives
So, how do we reconcile things? Earlier, we reported on a woodland conservation management policy inclusive of both open- and closed-habitat species at the woodland patch scale (Merckx et al. 2012a). Here, we propose another win-win situation, but now within a rewilding context: a conservation management strategy that could reconcile the needs of semi-natural habitat specialists at the regional scale.
The outcome of a management strategy is likely to differ dependent on whether it is applied at the local, landscape or regional scale, and this because species diversity under conditions of low land-use intensity is strongly dependent on spatial scale (see Chap. 1, Fig. 1.2) . We here opt for the regional scale, because we believe that within a European context it is more important, and more feasible too, to safeguard a certain species somewhere within a certain region (i.e. regional scale), rather than to safeguard the precise locations of the local populations of that species (i.e. local scale). As such, we consider the relevant spatial scale at which to consider the effects on species composition of the proposed strategy to be the regional (i.e. ca. 100 × 100 km) scale.
Our proposed strategy is not intended to be applied to European regions with fertile agricultural land, but rather to regions characterised by agricultural marginal land. Merckx and Pereira (in press) already warned against an overly agro-centric view on conservation for marginal land, which instead does provide excellent rewilding opportunities. Under our strategy, which we call ‘Controlled Rewilding’—combining forest recovery with monitoring and management of semi-natural biotopes—many such regions could evolve towards mosaics including mature climax vegetation, semi-natural biotopes, and natural successional stages, such as river areas, wood gaps and high-altitude areas. Nevertheless, open habitats may be rarer than in pre-historic landscapes, owing to the absence of most former natural herbivores (Merckx et al. 2013), but sun-lit biotopes and grazing can be achieved via other means (see Chap. 8). Eventually, the recovered forests within the resulting mosaics will become more and more dynamic and heterogeneous as a result of natural disturbance regimes operating on a wide range of spatial scales, characteristic of natural forests, and which further enhance biodiversity (Lindenmayer et al. 2006; Lehnert et al. 2013).
We believe there is room within Europe for Controlled Rewilding. It entails both passive abandonment and active (temporary) management interventions to ecologically restore semi-natural biotopes within a rewilding context. As a prerequisite, this strategy needs to include the monitoring of habitat heterogeneity levels at multiple spatial scales . Its aim is to pinpoint where conservation management interventions are required, so as to provide sufficient levels of habitat heterogeneity for specialists of both open and closed biotopes, and at multiple spatial scales in order to cater for small-sized (less mobile) and large-sized taxa (see above). In our opinion, conservation focused on semi-natural biotopes and rewilding should be complementary, via a Controlled Rewilding strategy, and now is the time to designate regions within Europe where both approaches could be combined (see Chaps. 2 and 11).
6 What About Fertile Agricultural Regions?
Does Controlled Rewilding for marginal land mean that we should forget about the ecosystems within fertile agricultural regions? Although it makes sense within a Europe-wide land-sparing framework to intensively farm such fertile regions (Merckx and Pereira in press), we remark that a dominant land-use of intensive farming does not necessarily have to imply current destructive practices, but merely the provision of high yields. Moreover, the conservation value of remaining semi-natural patches can be increased by an ecological upgrade of the intervening ‘matrix’, which basically consists of farmland, but also of brownfield sites and urbanised areas (Dennis 2010). Tangible environmental benefits can be obtained on farmland via agri-environment schemes (AES) (Donald and Evans 2006; Scheper et al. 2013), where the aim should be to reconcile intensive agricultural practices with wider societal benefits, including biodiversity. Here, the basic questions are which landscape elements to restore, how, and at what spatial scale, in order to make farmland less hostile to a broad range of declining ‘wider countryside’ and rare, localized species (Merckx et al. 2010a). Brownfield and even urban sites provide opportunities for restoration of successional biotopes otherwise not strongly represented locally, with restoration plans best tailored to focal species and/or to improving biotopes by assuring a sufficient quantity, quality and spatio-temporal diversity of habitat resources (New 2009; Dennis 2010).
AES can reverse negative biodiversity trends by increasing resource heterogeneity and improving dispersal success (Dennis 2010). However, they must be made more efficient and cost-effective (Scheper et al. 2013). One way to achieve this is by implementing specific measures for high-priority species targeted at landscapes where such species occur. However, we argue that this species-specific approach must be complemented by a multi-species approach in order to more fully address the steep declines in farmland biodiversity. General AES that are focused on the restoration and implementation of vital landscape elements are key to this multi-species approach. Even simple AES management prescriptions applied to relatively small areas can benefit Lepidoptera populations. For example, the restoration and management of arable field margins has been shown to benefit a range of insect groups (Haaland et al. 2011). Sympathetic management of hedgerows has positive effects on vulnerable insects, such as the brown hairstreak Thecla betulae butterfly (Merckx and Berwaerts 2010) and the lackey Malacosoma neustria (T. Merckx pers. data), a macro-moth of which the larvae feed gregariously on blackthorn Prunus spinosa and hawthorn Crataegus sp. In addition, we have recently discovered that the protection of existing hedgerow trees, and the provision of new ones, is likely to be a highly beneficial conservation tool for populations of moths, and probably many other flying insects too, as hedgerow trees provide a sheltered microclimate and other key habitat resources (Merckx et al. 2012b). The implementation of hedgerow tree and field margin AES options is likely to provide even better results in areas where farmers are targeted to join AES across the landscape. This approach results in a landscape-scale joining-up of habitat resources. Such a higher connectivity between resources at the landscape-scale does benefit fairly mobile species, which use the farmland biotope at a landscape-scale (see Chap. 7). A fair amount of macro-moth species falls into this category (Merckx and Macdonald in press).
7 A Case Study: Farmland Abandonment in Peneda and Its Effects on Macro-moths
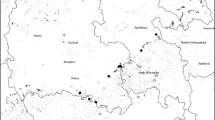
Coming back to the European marginal land context, where farmland abandonment is currently perceived as a major threat to the diversity of specialist butterflies from open biotopes and to other open-biotope invertebrates (Marini et al. 2009a; van Swaay et al. 2010), we recently carried out a two-year project (2011–2012) in Portugal to study the response of moths to farmland abandonment . There are a total of 2583 Lepidoptera species currently recognised from Portugal (Corley et al. 2013). We light-trapped macro-moths at 84 fixed sampling sites, each of which was repeatedly sampled six times. Sampling sites were part of a semi-nested sampling design (Proença and Pereira 2013) in three study landscapes that represented a farmland abandonment gradient within the Peneda mountain range (NW-Portugal; Fig. 6.1; elevation 750–1155 m). Here, an on-going rural exodus since the 1950s has led to farmland abandonment and regeneration of native woodland cover, although significant amounts of land, especially close to villages, are still being farmed (Rodrigues 2010). This situation is common to many other rural areas in Europe, and the Peneda area is considered to be representative of other traditional agricultural landscapes in mountainous areas of Southern Europe (Queiroz et al. submitted). We were able to analyse the species composition response to farmland abandonment and forest regeneration of both open-biotope and closed-biotope species across spatial scales, from local up to landscape scales. Nice add-ons to our research were the discovery of a micro-moth species new to science, namely Isotrias penedana, currently only known from the Peneda mountain range (Trematerra 2013; Fig. 6.2a), as well as the discovery of 12 macro-moth species new for Portugal (Corley et al. 2012, 2013; Fig. 6.2b).

Location of the study area within Portugal. Semi-nested sampling design: Each black dot represents the location of a fixed light-trap sampling site. Sampling occurred at four spatial scales (i.e. 20 × 20 m; 80 × 80 m; 320 × 320 m; 1280 × 1280 m) within each of three ‘landscapes’ that differed in terms of their dominant land-use cover: (i) ‘Forest-dominated’: the southernmost ‘landscape’ was mainly characterised by native, semi-natural forest, (ii) ‘Shrub-dominated’: the middle ‘landscape’ was mainly characterised by scrubland, and (iii) ‘Meadow-dominated’: the northernmost ‘landscape’ was mainly characterised by extensively managed meadows

a Notable moth discoveries Isotrias penedana, a tortricid (micro-moth) species new to science, discovered during 2012 within the Peneda mountain range (Trematerra 2013) (photo credit: Martin Corley). b Watsonarctia deserta, an arctiid (macro-moth) species added to the Portuguese list in 2012. In Portugal currently only known from four locations, all within the Peneda mountain range (Corley et al. 2013). (Photo credit: Eduardo Marabuto)
In a first analysis of the ca. 23.000 individuals and ca. 380 species rich dataset, we lumped the data from the six sampling rounds for each of the 84 local sites (20 × 20 m, as light traps have relatively small attraction radii: Merckx and Slade 2014). This analysis shows that overall macro-moth species richness is locally richest at woodland sites (mean ± SE: 77.1 ± 4.2), intermediate at meadows (63.2 ± 3.8) and tall shrub (61.7 ± 5.9), and poorest at low shrub sites (51.0 ± 4.3). Not only species richness, but functional diversity too was significantly higher within woodland than in meadows or shrubland for macro-moths (Queiroz et al. submitted). Although this overall pattern among the four main biotopes is basically present in the meadow-dominated, shrub-dominated and forest-dominated landscapes, species richness is locally, at all four biotope types, consistently lower within the shrub-dominated landscape than in meadow-dominated and forest-dominated landscapes (Fig. 6.3a). Possible confounding factors, such as altitude, slope and soil fertility, were not analysed. Although they may have played a role in the patterns observed, their role is likely to have been small given the large amount of sites sampled within the same region, and given the limited altitudinal differences between sites .

Macro-moth species richness (± SE)—Local sites (20 × 20 m; N = 84) were each sampled six times over 2 years (2011–2012). Data from the sampling rounds were aggregated for each site. a Overall richness at local sites for each of four biotopes (Meadow; Low Shrub; Tall Shrub; Woodland), separately for three landscape types (Meadow-dominated; Shrub-dominated; Forest-dominated) (see also Fig. 6.1). Note that in the forest-dominated landscape only one meadow site and two tall shrub sites were sampled, explaining the absent and very large error bars, respectively. b Overall richness at four spatial scales [20 × 20 m: local site (N = 84); 80 × 80 m: lumping four sites (N = 12); 320 × 320 m: lumping seven sites (N = 12); 1280 × 1280 m: lumping 28 sites (N = 3)], separately for three landscape types (Meadow-dominated; Shrub-dominated; Forest-dominated) (see also Fig. 6.1). The absence of error bars at the largest spatial scale is because these three landscape types were each represented by one landscape only. c Richness of closed-biotope (i.e. woodland) versus open-biotope species at five spatial scales (see Fig. 6.3b, but with an additional larger scale lumping all 84 sites). For contrast, a majority of species (N = 223) that occur in mixed or intermediate biotopes are not retained
These observations suggest that local abandonment of relatively species-rich, semi-natural meadows may reduce (−20 %) local richness levels of nocturnal macro-moths, when meadows turn into low shrub biotopes, but that richness levels should locally increase again when these abandoned fields gradually turn into native forest, eventually reaching diversity levels well above those associated with meadows (+ 22 %). The above results may also mean that when the resulting shrubby vegetation from farmland abandonment becomes the dominant land cover within a landscape (for instance due to simultaneous abandonment of large areas within the same landscape and/or due to the arrested development of the resulting low shrub by overgrazing/shepherding and fire), that this landscape effect lowers local species richness levels. This would happen within remaining meadows and woodlands due to fragmentation effects, and also within the scrubland itself due to lower species inputs from different habitats (i.e. reduced spillover effects because of lower heterogeneity). Eventually, such a process may lead to an impoverished species composition of mainly shrub specialists (e.g. the horse chestnut moth Pachycnemia hippocastanaria) and ubiquitous species, and this at both the local and landscape-scale.
Our data also show that this negative landscape-effect of a dominance of shrub vegetation within the surrounding landscape does not only reduce local species richness, affecting all biotopes, but translates to reduced species richness at larger scales too (Fig. 6.3b). By contrast, overall macro-moth richness was highest in the forest-dominated landscape and intermediate in the meadow-dominated landscape, at all four spatial scales tested (Fig. 6.3b). Moreover, the difference between the forest-dominated and meadow-dominated landscape in absolute species richness steadily increases with spatial scale, from an 11 species difference at the local scale, over a 24 and 32 species difference at intermediate spatial scales , to an excess of 41 species for the forest-dominated landscape at the largest scale tested (i.e. 1280 × 1280 m) (Fig. 6.3b) . We interpret these results as follows: extensively farmed landscapes do indeed provide high levels of moth diversity, both locally and at the landscape-scale. Still, more has to be gained from landscapes with a high amount of forest cover. Although the species richness difference is not that big at the local scale, the difference becomes larger and more notable with increasing scale, which points to larger beta-diversity levels for forest-dominated than for meadow-dominated landscapes/regions. Indeed, it is well known that semi-natural grasslands are able to reach high local species richness (i.e. alpha diversity) , at least for certain taxa (e.g. butterflies: van Swaay 2002; flowering plants: Wilson et al. 2012) . However, we show, for macro-moths, that those local diversity levels can be even higher within woodlands, and importantly, that beta-diversity levels may be consistently larger at larger spatial scales within landscapes mainly covered by forests compared to landscapes mainly covered by extensively managed agricultural land. So, although the effects on macro-moth diversity of woodlands and forest-dominated landscapes already compare positively with meadows and meadow-dominated landscapes at the local scale, forest-dominated landscapes outcompete meadow-dominated landscapes more strongly at larger spatial scales. We believe this is an important point since rewilding is to be applied at larger, regional scales, whereas biodiversity has traditionally been measured mainly at local scales alone.
A key result linked to this is the interaction we observed between spatial scale and species’ biotope characteristics, whereby the species richness of woodland moths shows a steeper increase with spatial scale than the richness of open-biotope moths (Fig. 6.3c). Whilst we found on average more open-biotope species than typical woodland species locally, this difference disappears at the field-scale, and reverses at the largest scales. Thus, it appears that closed-biotope species have higher beta-diversity than open-biotope species, and that the former are indeed responsible for the higher diversity levels within forest-dominated landscapes.
Finally, these results also demonstrate that although forest-dominated landscapes provide advantages over meadow-dominated landscapes, one needs to take good care not to get stuck into a landscape largely dominated by shrubs alone (Fig. 6.3a, b). Rather, we advise to monitor, and if needed to cater for spatial and temporal habitat heterogeneity within landscapes undergoing farmland abandonment, so as to combat these negative effects .
8 Habitat Resource Heterogeneity at Multiple Spatial Scales is Key
Let’s now zoom out from the specifics of the Peneda case study on macro-moths to a more general view on rewilding landscapes for multiple taxa. In order to do so, it is essential to consider dispersal, not only because it is a fundamental process that bridges across spatial scales (Chave 2013), but also because it allows us to understand the importance of habitat resource heterogeneity. Non-sessile organisms need to move in order to reach their essential habitat resources, needed for the completion of their life-cycle. These varied resources are often spread over spatial and temporal gradients, and are most often patchily distributed, even within continuous vegetation types (Dennis 2010) . Since dispersal is costly in terms of energy expenditure and predation risk, organisms hence generally benefit from resource configurations that limit dispersal needs (Vanreusel and Van Dyck 2007). Landscapes characterised by high habitat resource heterogeneity are more likely to provide such resource configurations (Dennis 2010; Vickery and Arlettaz 2012), and hence to provide increased species diversity (Verhulst et al. 2004). However, since there is a vast variety in terms of dispersal capacities and resource use among, and even within taxa, such heterogeneity needs to be provided at multiple spatial scales in order to cater for all. After all, each species/individual is adapted to exploit resources within a spatial range, some more specifically so than others, and this over a whole gradient from extreme widely to extreme narrowly spaced resources. Evidently, the more biotope types within a landscape, the wider the array of resources and thus the more organisms supported. Also, for a given species, large enough quantities of its essential resources need to be present, limiting the minimum patch size below which it will be absent. In general, mobile organisms need larger areas of habitat than less-mobile ones (Pereira et al. 2004). For instance, mobile woodland moth species were not found in small (< 5 ha) wood patches (Slade et al. 2013). As such, the high macro-moth diversity in the forest-dominated landscape of Peneda (see above) can be explained by a combination of (i) sufficiently large woodland patches to cater for the needs of mobile woodland specialists, and (ii) sufficient resource heterogeneity at multiple spatial scales . Indeed, the specific study landscape was not a forest ‘monoculture’ but consisted of a patchwork of meadows and scrubland of varying size embedded within a dominant forest matrix (Fig. 6.1). This patchwork allows a diverse composition of meadow, scrubland and woodland species.
Habitat heterogeneity is known to strongly influence the abundance and diversity of species within landscapes (Tjørve 2002). For instance, the change of the typically diverse habitat mosaic of extensive farmland towards the spatially and temporally—both at multiple scales—increasingly uniform intensive farmscapes, has been identified as the root cause of the decline in European farmland biodiversity, whether measured at a small or large scale (Benton et al. 2003). Likewise, we believe that rewilding exercises need to pay attention that sufficient heterogeneity remains during the whole rewilding process. For example, whilst bird species typical of extensive farmland disappear when landscapes become too open due to intensification, they may at the other extreme also disappear from areas where forest recovery removes all open areas (Vickery and Arlettaz 2012). Rewilding projects should hence monitor heterogeneity and intervene with conservation management when necessary (Controlled Rewilding: see above). Because the intense defaunation (especially of large mammals) since the Pleistocene has lead to a decreased environmental heterogeneity in remaining ecosystems (Corlett 2013), and since current rewilding exercises are not continent-wide but region-wide at best, it should be clear that hands-off restoration processes may not be sufficient to reach desirable biodiversity outcomes, and that Controlled Rewilding is more advisable.
9 Wrapping It Up and the Way Forward
Semi-natural meadows and other open biotopes are important contributors to agro-ecosystem biodiversity on marginal land (Knop et al. 2006). Whilst the negative effects of grassland intensification on biodiversity are relatively well understood (Marini et al. 2008), the impact of grassland abandonment—common to European mountainous areas—is much less understood, and hence controversial. In our view, the alleged negative effects result from two biases: species composition studies are often performed at relatively small scales, and narrowly focus on open-biotope species alone. These biases ignore that ecosystem dynamics occur across spatial scales, and that ecosystems consist of other functional groups too.
Nevertheless, evidence exists that cessation of management, and the resulting transitional vegetation types, can be important—even at the field-scale—for several taxonomic groups (Lepidoptera: Balmer and Erhardt 2000; Baur et al. 2006; Öckinger et al. 2006; Skórka et al. 2007—Apoidea: Kruess and Tscharntke 2002—Orthoptera: Marini et al. 2009a). Similarly, a recent review paper convincingly shows that reduced grazing generally positively affects arthropod diversity (van Klink et al. 2014).
We believe that studies on biodiversity effects should include landscape and/or regional scales. Ecological processes are not limited to the field-scale, as many organisms move at larger spatial scales , and respond differently to the surrounding landscape according to their size, mobility and functional traits (Steffan-Dewenter et al. 2002; Tscharntke et al. 2005; Merckx et al. 2009, 2010b; Slade et al. 2013; Queiroz et al. submitted). At these larger scales we expect positive effects from land abandonment. Indeed, reduced effects of habitat fragmentation, and high landscape heterogeneity, may lead to better functioning, more diverse, and hence more resilient ecosystems (Loreau and de Mazancourt 2013). For example, Marini et al. (2008, 2009b) show that a high proportion of woody vegetation at a landscape-scale positively affects Orthopteran species richness, whereas a high proportion of grasslands did so negatively. Other examples are birds of prey which obtain resources from both farm- and woodland. Their densities tend to increase with forest cover; Booted eagle (Aquila pennata) densities, for instance, peak at ca. 80 % forest cover (Sánchez-Zapata and Calvo 1999). Conservation of raptors, and many other mobile taxa, thus requires a regional approach towards the right landscape mosaics of forests and open biotopes (Vickery and Arlettaz 2012). A marked increase in native forest cover within Europe may locally also combat the increased frequency and intensity of fires due to climate change (Proença et al. 2010), which are considered threats to European butterfly diversity (van Swaay et al. 2010). Nevertheless, as the highest diversity of (threatened) butterflies is found in mountainous areas in Southern Europe, where numerous restricted-range species are encountered (van Swaay et al. 2010), and where a high degree of farmland abandonment is likely (see Chap. 1, Fig. 1.4), rewilding projects in such areas will need to make sure they retain all key habitat resources of such high-priority species at sufficient levels within (managed) open habitats (see Chap. 8).
Here, we reported on research within the Peneda mountain range in which we looked at the response of macro-moth species composition to farmland abandonment across multiple spatial scales. Based on our findings and on general ecological insights, we strongly recommend that a possible rewilding approach would not be equalled to a hands-off approach, but rather to a conservation management approach that monitors, and when necessary intervenes to provide habitat heterogeneity at multiple spatial scales , in order to cater for the whole gradient of sedentary to mobile species. Given that sufficient levels of habitat heterogeneity are maintained, Lepidoptera are one of probably many taxa that are likely to benefit from rewilding exercises on European marginal farmland. An important point to take on board for nocturnal Lepidoptera, and other nocturnal biodiversity, is the issue of light pollution , which may cause adverse effects on larval development, fitness and population dynamics (van Geffen et al. 2014), and may hence result in negative effects cascading through whole ecosystems (van Langevelde et al. 2011; Fox et al. 2013). Rewilding exercises should thus include measures to actively mitigate light pollution sources (see Chap. 2). The likely improved species composition as a result will not only help achieve European species conservation targets (i.e. halting and reversing declines in biodiversity), but stronger populations of moths , butterflies and other invertebrates will also result in increased resilience of food-webs and ecosystem functioning (Loreau and de Mazancourt 2013).
Since there is an urgent need to determine the effects of farmland abandonment and landscape context on communities over a wide range of spatial scales (Cozzi et al. 2008), we believe that our study on macro-moths in the Peneda range provides useful and timely scientific evidence on the benefits, requirements and limitations of the rewilding approach for marginal land, such as mountainous areas and High Nature Value farmland within Europe. We hope that the approach of Controlled Rewilding here proposed, may help design an effective conservation policy regarding the European farmland abandonment process. We here call for more research into how optimal configurations of rewilded land on abandoned farmland would look like. We also call to develop regional goals for biodiversity on land characterised by a high degree of farmland abandonment. Although it is clear that a universal approach to provide habitat heterogeneity at multiple spatial scales is the key to deliver and sustain the required biodiversity, we may need different types of heterogeneity in different places and regions.
Acknowledgements
I am immensely grateful to Martin Corley, who provided lots of help and advice with the identification of the numerous samples. I also thank Henrique M. Pereira, Laetitia M. Navarro, the Theoretical Ecology and Biodiversity Change Group, Eduardo Marabuto, Anabela Moedas and Pedro Alarcão for their help and support throughout. The Fundação para a Ciência e a Technologia provided financial support (postdoc grant SFRH/BPD/74393/2010), and both the Peneda-Gerês National Park and the Instituto da Conservação da Natureza e da Biodiversidade allowed carrying out the fieldwork.
References
Balmer, O., & Erhardt, A. (2000). Consequences of succession on extensively grazed grasslands for Central European butterfly communities: Rethinking conservation practices. Conservation Biology, 14, 746–757.
Baur, B., Cremene, C., Groza, G., Rakosy, L., Schileyko, A. A., Baur, A., Stoll, P., & Erhardt, A. (2006). Effects of abandonment of subalpine hay meadows on plant and invertebrate diversity in Transylvania, Romania. Biological Conservation, 132, 261–273.
Benton, T. G., Vickery, J. A., & Wilson, J. D. (2003). Farmland biodiversity: Is habitat heterogeneity the key? Trends in Ecology and Evolution, 18, 182–188.
Cardoso, P., Erwin, T. L., Borges, P. A., & New, T. R. (2011). The seven impediments in invertebrate conservation and how to overcome them. Biological Conservation, 144, 2647–2655.
Chave, J. (2013). The problem of pattern and scale in ecology: What have we learned in 20 years? Ecology Letters, 16, 4–16.
Clark, J. A., & May, R. M. (2002). Taxonomic bias in conservation research. Science, 297, 191–192.
Corlett, R. T. (2013). The shifted baseline: Prehistoric defaunation in the tropics and its consequences for biodiversity conservation. Biological Conservation, 163, 13–21.
Corley, M. F. V., Merckx, T., Cardoso, J. P., Dale, M. J., Marabuto, E., Maravalhas, E., & Pires, P. (2012). New and interesting Portuguese Lepidoptera records from 2011 (Insecta: Lepidoptera). SHILAP Revista de Lepidopterología, 40, 489–511.
Corley, M. F. V., Merckx, T., Marabuto, E., Arnscheid, W., & Maravalhas, E. (2013) New and interesting Portuguese Lepidoptera records from 2012 (Insecta: Lepidoptera). SHILAP Revista de Lepidopterología, 41, 449–477.
Cozzi, G., Müller, C. B., & Krauss, J. (2008). How do local habitat management and landscape structure at different spatial scales affect fritillary butterfly distribution on fragmented wetlands? Landscape Ecology, 23, 269–283.
Dennis, R. L. H. (2010). A resource-based habitat view for conservation: Butterflies in the British landscape. Chichester: Wiley-Blackwell.
Donald, P. F., & Evans, A. D. (2006). Habitat connectivity and matrix restoration: The wider implications of agri-environment schemes. Journal of Applied Ecology, 43, 209–218.
Dover, J. W., Spencer, S., Collins, S., Hadjigeorgiou, I., & Rescia, A. (2011). Grassland butterflies and low intensity farming in Europe. Journal of Insect Conservation, 15, 129–137.
Dullinger, S., Essl, F., Rabitsch, W., Erb, K. H., Gingrich, S., Haberl, H., Hülber, K., Jarosik, V., Krausmann, F., Kühn, I., Pergl, J., Pysek, P., & Hulme, P. E. (2013). Europe’s other debt crisis caused by the long legacy of future extinctions. Proceedings of the National Academy of Sciences, 110, 7342–7347.
Ellis, W., Groenendijk, D., Groenendijk, M., Huigens, T., Jansens, M., van der Meulen, J., van Nieukerken, E., & de Vos, R. (2013). Nachtvlinders Belicht—Dynamisch, Belangrijk, Bedreigd. De Vlinderstichting:The Netherlands.
Fartmann, T., Müller, C., & Poniatowski, D. (2013). Effects of coppicing on butterfly communities of woodlands. Biological Conservation, 159, 396–404.
Fox, R., Parsons, M. S., Chapman, J. W., Woiwod, I. P., Warren, M. S., & Brooks, D. R. (2013). The State of Britain’s Larger Moths 2013. Butterfly Conservation and Rothamsted Research, Wareham, Dorset, UK.
Haaland, C., Naisbit, R. E., & Bersier, L. F. (2011). Sown wildflower strips for insect conservation: a review. Insect Conservation and Diversity, 4, 60–80.
Hambler, C., Henderson, P. A., & Speight, M. R. (2011). Extinction rates, extinction-prone habitats, and indicator groups in Britain and at larger scales. Biological Conservation, 144, 713–721.
Knop, E., Kleijn, D., Herzog, F., & Schmid, B. (2006). Effectiveness of the Swiss agri-environment scheme in promoting biodiversity. Journal of Applied Ecology, 43, 120–127.
Kruess, A., & Tscharntke, T. (2002). Grazing intensity and the diversity of grasshoppers, butterflies and trap-nesting bees and wasps. Conservation Biology, 16, 1570–1580.
Lehnert, L. W., Bässler, C., Brandl, R., Burton, P. J., & Müller, J. (2013). Conservation value of forests attacked by bark beetles: Highest number of indicator species is found in early successional stages. Journal for Nature Conservation, 21, 97–104.
Lindenmayer, D. B., Franklin, J. F., & Fischer, J. (2006). General management principles and a checklist of strategies to guide forest biodiversity conservation. Biological Conservation, 131, 433–445.
Loreau, M., & de Mazancourt, C. (2013). Biodiversity and ecosystem stability: A synthesis of underlying mechanisms. Ecology Letters, 16, 106–115.
Maes, D., Vanreusel, W., & Van Dyck, H. (2013). Dagvlinders in Vlaanderen—Nieuwe Kennis voor Betere Actie (Butterflies in Flanders—New Knowledge for Better Action). Lannoo, Tielt, Belgium.
Marini, L., Fontana, P., Scotton, M., & Klimek, S. (2008). Vascular plant and Orthoptera diversity in relation to grassland management and landscape composition in the European Alps. Journal of Applied Ecology, 45, 361–370.
Marini, L., Fontana, P., Battisti, A., & Gaston, K. J. (2009a). Response of orthopteran diversity to abandonment of semi-natural meadows. Agriculture, Ecosystems and Environment, 132, 232–236.
Marini, L., Fontana, P., Battisti, A., & Gaston, K. J. (2009b). Agricultural management, vegetation traits and landscape drive orthopteran and butterfly diversity in a grassland-forest mosaic: A multi-scale approach. Insect Conservation and Diversity, 2, 213–220.
Menken, S. B. J., Boomsma, J. J., & van Nieukerken, E. J. (2010). Large-scale evolutionary patterns of host plant associations in the Lepidoptera. Evolution, 64, 1098–1119.
Merckx, T., & Berwaerts, K. (2010). What type of hedgerows do Brown hairstreak (Thecla betulae L.) butterflies prefer? Implications for European agricultural landscape conservation. Insect Conservation and Diversity, 3, 194–204.
Merckx, T., & Macdonald, D. W. (in press). Landscape-scale conservation of farmland moths. In D. W. Macdonald & R. E. Feber (Eds.), Wildlife conservation on farmland. Oxford University Press.
Merckx, T. & Pereira, H. M. (in press). Reshaping agri-environmental subsidies: From marginal farming to large-scale rewilding. Basic and Applied Ecology. doi: 10.1016/j.baae.2014.12.003
Merckx, T., & Slade, E. M. (2014). Macro-moth families differ in their attraction to light: Implications for light trap monitoring programs. Insect Conservation and Diversity, 7, 453–461.
Merckx, T., Feber, R. E., Dulieu, R. L., Townsend, M. C., Parsons, M. S., Bourn, N. A. D., Riordan, P., & Macdonald, D. W. (2009). Effect of field margins on moths depends on species mobility: Field-based evidence for landscape-scale conservation. Agriculture, Ecosystems and Environment, 129, 302–309.
Merckx, T., Feber, R. E., Parsons, M. S., Bourn, N. A. D., Townsend, M. C., Riordan, P., & Macdonald, D. W. (2010a). Habitat preference and mobility of Polia bombycina: Are non-tailored agri-environment schemes any good for a rare and localised species? Journal of Insect Conservation, 14, 499–510.
Merckx, T., Feber, R. E., Mclaughlan, C., Bourn, N. A. D., Parsons, M. S., Townsend, M. C., Riordan, P., & Macdonald, D. W. (2010b). Shelter benefits less mobile moth species: The field-scale effect of hedgerow trees. Agriculture, Ecosystems and Environment, 138, 147–151.
Merckx, T., Feber, R. E., Hoare, D. J., Parsons, M. S., Kelly, C. J., Bourn, N. A. D., & Macdonald, D. W. (2012a). Conserving threatened Lepidoptera: Towards an effective woodland management policy in landscapes under intense human land-use. Biological Conservation, 149, 32–39.
Merckx, T., Marini, L., Feber, R. E., & Macdonald, D. W. (2012b). Hedgerow trees and extended width field margins enhance macro-moth diversity: Implications for management. Journal of Applied Ecology, 49, 1396–1404.
Merckx, T., Huertas, B., Basset, Y., & Thomas, J. A. (2013). A global perspective on conserving butterflies and moths and their habitats. In D. W. Macdonald & K. J. Willis (Eds.), Key topics in conservation biology 2 (pp. 239–257). Chichester: Wiley-Blackwell.
Monbiot, G. (2013). Feral—Searching for enchantment on the frontiers of rewilding. London: Penguin Books.
New, T. R. (2009). Insect species conservation. Cambridge: Cambridge University Press.
Öckinger, E., Eriksson, A. K., & Smith, H. G. (2006). Effects of grassland abandonment, restoration and management on butterflies and vascular plants. Biological Conservation, 133, 291–300.
Pereira, H. M., Daily, G. C., & Roughgarden, J. (2004). A framework for assessing the relative vulnerability of species to land-use change. Ecological Applications, 14, 730–742.
Pereira, H. M., Navarro, L. M., & Martins, I. S. (2012). Global biodiversity change: The bad, the good, and the unknown. Annual Review of Environment and Resources, 37, 25–50.
Proenca, V., & Pereira, H. M. (2013). Species-area models to assess biodiversity change in multi-habitat landscapes: The importance of species habitat affinity. Basic and Applied Ecology, 14, 102–114.
Proença, V., Pereira, H. M., & Vicente, L. (2010). Resistance to wildfire and early regeneration in natural broadleaved forest and pine plantation. Acta Oecologica, 36, 626–633.
Queiroz, C., Proença, V., Merckx, T., Guilherme, J., Ceaușu, S., Pereira, H. M., & Lindborg, R. (submitted). Habitat, scale and species richness affect functional diversity of plants, moths and birds. Ecography.
Rodrigues, P. (2010) Landscape changes in Castro Laboreiro: From farmland abandonment to forest regeneration. MSc Thesis, Faculdade de Ciências da Universidade de Lisboa, Portugal.
Sánchez-Zapata, J. A., & Calvo, J. F. (1999). Raptor distribution in relation to landscape composition in semi-arid Mediterranean habitats. Journal of Applied Ecology, 36, 254–262.
Scheper, J., Holzschuh, A., Kuussaari, M., Potts, S. G., Rundlöf, M., Smith, H. G., & Kleijn, D. (2013). Environmental factors driving the effectiveness of European agri-environmental measures in mitigating pollinator loss—a meta-analysis. Ecology Letters, 16, 912–920.
Sierens, T., & Van de Kerckhove, O. (2014). 32 years of moth inventories between Bruges and Ghent [In Dutch]. Natuur.Focus, 13, 66–71.
Skórka, P., Settele, J., & Woyciechowski, M. (2007). Effects of management cessation on grassland butterflies in southern Poland. Agriculture, Ecosystems and Environment, 121, 319–324.
Slade, E. M., Merckx, T., Riutta, T., Bebber, D. P., Redhead, D., Riordan, P., & Macdonald, D. W. (2013). Life-history traits and landscape characteristics predict macro-moth responses to forest fragmentation. Ecology, 94, 1519–1530.
Steffan-Dewenter, I., Münzenberg, U., Bürger, C., Thies, C., & Tscharntke, T. (2002). Scale-dependent effects of landscape context on three pollinator guilds. Ecology, 83, 1421–1432.
Thomas, J. A. (2005). Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Philosophical Transactions of the Royal Society B: Biological Sciences, 360, 339–357.
Thomas, J. A., Morris, M. G., & Hambler, C. (1994). Patterns, mechanisms and rates of extinction among invertebrates in the United Kingdom [and discussion]. Philosophical Transactions of the Royal Society of London Series B: Biological Sciences, 344, 47–54.
Tjørve, E. (2002). Habitat size and number in multi-habitat landscapes: A model approach based on species-area curves. Ecography, 25, 17–24.
Trematerra, P. (2013). Isotrias penedana sp. n. a new species of Lepidoptera (Tortricidae: Chlidanotinae: Polyorthini) from Portugal. Journal of Entomological and Acarological Research, 45, e1.
Tscharntke, T., Kleijn, A. M., Kruess, A., Steffan-Dewenter, I., & Thies, C. (2005). Landscape perspectives on agricultural intensification and biodiversity: Ecosystem service management. Ecology Letters, 8, 857–874.
van Geffen, K. G., van Grunsven, R. H., van Ruijven, J., Berendse, F., & Veenendaal, E. M. (2014). Artificial light at night causes diapause inhibition and sex-specific life history changes in a moth. Ecology and Evolution, 4, 2082–2089.
van Klink, R., van der Plas, F., van Noordwijk, C. G. E., Wallis de Vries, M. F., & Olff, H. (2014). Effects of large herbivores on grassland arthropod diversity. Biological Reviews, DOI: 10.1111/brv.12113.
van Langevelde, F., Ettema, J. A., Donners, M., Wallis de Vries, M. F., & Groenendijk, D. (2011). Effect of spectral composition of artificial light on the attraction of moths. Biological Conservation, 144, 2274–2281.
Vanreusel, W., & Van Dyck, H. (2007). When functional habitat does not match vegetation types: A resource-based approach to map butterfly habitat. Biological Conservation, 135, 202–211.
van Swaay, C. A. M. (2002). The importance of calcareous grasslands for butterflies in Europe. Biological Conservation, 104, 315–318.
van Swaay, C., Cuttelod, A., Collins, S., Maes, D., López Munguira, M., Šašić, M., Settele, J., Verovnik, R., Verstrael, T., Warren, M., Wiemers, M., & Wynhof, I. (2010). European Red List of butterfies. IUCN and Butterfly Conservation Europe. Luxembourg: Publications Office of the European Union.
Verdasca, M. J., Leitão, A. S., Santana, J., Porto, M., Dias, S., & Beja, P. (2012). Forest fuel management as a conservation tool for early successional species under agricultural abandonment: The case of Mediterranean butterflies. Biological Conservation, 146, 14–23.
Verhulst, J., Báldi, A., & Kleijn, D. (2004). Relationship between land-use intensity and species richness and abundance of birds in Hungary. Agriculture, Ecosystems and Environment, 104, 465–473.
Vickery, J., & Arlettaz, R. (2012). The importance of habitat heterogeneity at multiple scales for birds in European agricultural landscapes. In R. J. Fuller (Ed.), Birds and habitat: Relationships in changing landscapes (pp. 177–204). Cambridge University Press:UK.
Wilson, J. B., Peet, R. K., Dengler, J., & Pärtel, M. (2012). Plant species richness: The world records. Journal of Vegetation Science, 23, 796–802.
Young, M. (1997). The natural history of moths. Poyser Natural History:UK.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2015 The Author(s)
About this chapter
Cite this chapter
Merckx, T. (2015). Rewilding: Pitfalls and Opportunities for Moths and Butterflies. In: Pereira, H., Navarro, L. (eds) Rewilding European Landscapes. Springer, Cham. https://doi.org/10.1007/978-3-319-12039-3_6
Download citation
DOI: https://doi.org/10.1007/978-3-319-12039-3_6
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-12038-6
Online ISBN: 978-3-319-12039-3
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)