
Abstract
In this chapter, we summarize the ecology and conservation issues affecting greater (Centrocercus urophasianus) and Gunnison (C. minimus) sage-grouse, iconic and obligate species of rangelands in the sagebrush (Artemisia spp.) biome in western North America. Greater sage-grouse are noted for their ability to migrate, whereas Gunnison sage-grouse localize near leks year-round. Seasonal habitats include breeding habitat where males display at communal leks, nesting habitat composed of dense sagebrush and herbaceous plants to conceal nests, mesic summer habitats where broods are reared, and winter habitat, characterized by access to sagebrush for cover and forage. While two-thirds of sage-grouse habitat occurs on public lands, private land conservation is the focus of national groups including the USDA-NRCS Sage-Grouse Initiative. Sage-grouse are a species of great conservation concern due to population declines associated with loss and fragmentation of more than half of the sagebrush biome. Wildlife and land management agencies have been increasingly proactive in monitoring trends in sage-grouse populations (e.g., lek count index), adapting regulations to reduce harvest on declining populations, and in designing and implementing conservation policies such as core areas to conserve sage-grouse habitats and populations. Much of the remaining sagebrush habitat is threatened by altered fire regimes, invasive annual grasses and noxious weeds, encroaching piñon (Pinus edulis and monophylla)-juniper (Juniperus spp.) woodlands, sagebrush conversion, anthropogenic development, and climate change. Several diseases affect sage-grouse, but to date, disease has not been a widespread cause of declines. Proper livestock grazing and limited hunting appear to be sustainable with sage-grouse, whereas improper grazing, increasing free-roaming equid populations, and sagebrush conversion are primary concerns for future conservation. Research has identified additional concerns for sage-grouse including effects from fence collisions, predation from common ravens (Corvus corax), and reduced habitat effectiveness resulting from grouse avoidance of anthropogenic infrastructure. There is a need for future research evaluating sage-grouse habitat restoration practices following improper rangeland management, habitat alteration from invasive species and fire, effects on small and isolated populations, and effects from diseases.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Centrocercus urophasianus
- Centrocercus minimus
- Ecosystem threats
- Greater sage-grouse
- Gunnison sage-grouse
- Private and public land conservation
- Rangeland management
- Sagebrush
1 General Life History and Population Dynamics
Greater (Centrocercus urophasianus) and Gunnison (C. minimus) sage-grouse are icons of the sagebrush (Artemisia spp.) biome in western North America. Sage-grouse are world renowned for their spectacular lek breeding system where males congregate at traditional locations to display for and breed with females (Fig. 10.1). Competition among males at leks is intense and relatively few males breed with most of the females. Females are ground-nesting Galliformes that produce a maximum of 1 successful clutch with 7–9 chicks per year after a 4-week incubation period, with no help from the males (Schroeder et al. 2020; Young et al. 2020). Because nest success, chick survival, and rates of renesting are generally low, the relatively high survival of breeding-aged birds helps to maintain their populations (Crawford et al. 2004; Connelly et al. 2011a; Taylor et al. 2012; Blomberg et al. 2013c; Davis et al. 2014b; Dahlgren et al. 2016). Sage-grouse are long-lived relative to other game birds, with an observed maximum survival for greater sage-grouse of 9 years in females and 7 years in males (Zablan et al. 2003), though maximum longevity is likely higher.

Greater (Photo a) and Gunnison (Photo b) sage-grouse distributions in the western United States and Canada based on Schroeder et al. (2004), Environment Canada (2014), and USFWS (2015). Ecoregions represent EPA Level III ecoregions across North America (Wilken et al. 2011). Single hatched area represents current distribution of greater (black) and Gunnison (dark orange) sage-grouse, and cross-hatched area represents additional historical distribution of greater (grey) and Gunnison (light orange) sage-grouse
Although greater sage-grouse often migrate seasonally, migrations are typically within the same general regions and ecosystems (Connelly et al. 1988; Fischer et al. 1996; Fedy et al. 2012). However, they stand out among upland gamebirds in their capability for long-distance migrations—the longest migration by a greater sage-grouse was documented at 240 km for a female between southern Saskatchewan and north-central Montana (Newton et al. 2017). Telemetry studies indicate most Gunnison sage-grouse spend their annual life cycles within 5 km of their lek of capture (Aldridge et al. 2012; Young et al. 2020). Greater sage-grouse populations are often partially migratory, with some individuals migrating and others remaining within the same areas year-round (Fedy et al. 2012; Pratt et al. 2017, 2019), which can complicate conservation efforts (Dinkins et al. 2017; Pratt et al. 2019). Greater sage-grouse in many populations migrate altitudinally from xeric lower elevation Wyoming big sagebrush (A. tridentata wyomingensis) in spring to more mesic higher elevation mountain big sagebrush (A. t. vaseyana) during summer, and then to lower elevations for winter and the following spring (Beck et al. 2006; Pratt et al. 2017, 2019). In some areas, greater sage-grouse may move to irrigated agricultural fields, in many cases alfalfa (Medicago sativa), in response to summer desiccation of native forbs (Fischer et al. 1996).
Sage-grouse select specific structural aspects of sagebrush as well as herbaceous understory components throughout life stages including dense sagebrush and grass cover for successful nesting, increased forb cover at early (through 2 weeks; Thompson et al. 2006), and late brood-rearing/summer (> 2 weeks; Smith et al. 2018c), and dense sagebrush for winter habitat (Fig. 10.2). Sage-grouse depend on sagebrush as a primary food source, particularly during the late autumn, winter, and early spring (Schroeder et al. 2020). They digest the leaves of sagebrush, while also tolerating high levels of monoterpenoids and other plant secondary metabolites, toxins most species cannot consume (Sauls 2006; Kohl et al. 2016; Oh et al. 2019). Sage-grouse also depend on the cover provided by sagebrush during nesting and brood-rearing and during all times of year for protection from potential predators (Schroeder et al. 2020; Young et al. 2020). Although sage-grouse are adept at snow burrowing to escape severe winter conditions (Back et al. 1987), winter landscapes that support sage-grouse are characterized by south and west aspects and patches of taller sagebrush that sage-grouse can access following deep snow accumulations (Hupp and Braun 1989).

Life stages of sage-grouse within seasonal habitats. Successful nest and early brood photographs from N. Paothang. Nesting, early brood-rearing, and late brood-rearing/summer photos from T. Christiansen. Winter photograph from J. Lautenbach
The importance of sagebrush within sage-grouse habitat is clear, but additional vegetation components of these habitats provide necessary function as well (Connelly et al. 2011b; Dumroese et al. 2015; Pennington et al. 2016). Herbaceous cover (grasses and forbs) provides essential concealment of nests, often explaining differences in nest site selection (Hagen et al. 2007). However, only one study has reported a weak effect of grass cover and height positively influencing nest success in greater sage-grouse (Holloran et al. 2005). A rangewide meta-analysis indicated common fine-scale herbaceous and shrub structural characteristics do not consistently influence nest success in greater sage-grouse (Smith et al. 2020). This study also found shrub characteristics such as sagebrush cover had moderate, yet context‐dependent, effects, and herbaceous vegetation characteristics had weak effects, on sage-grouse nest site selection (Smith et al. 2020). Herbaceous cover, especially forbs, provides critical food during the late spring to early autumn period (Drut et al. 1994; Huwer et al. 2008), and provides habitat for a variety of invertebrates. Invertebrates such as ants, beetles, and grasshoppers are consumed by all sage-grouse (Klebenow and Gray 1968; Peterson 1970) and are essential for survival of young chicks (Johnson and Boyce 1990).
2 Species and Population Status
2.1 Historical Versus Current Distributions, Conservation Status
Sage-grouse were historically found throughout most of the vast sagebrush-dominated landscape in western North America (Bendire 1892; McClanahan 1940; Aldrich and Duvall 1955; Aldrich 1963; Zwickel and Schroeder 2003; Schroeder et al. 2004). Although the original distribution of both species (Fig. 10.1) was established by comparing historical observations (Aldrich and Duvall 1955) with the distribution of potentially suitable habitat (Küchler 1985), precise observations of distribution and abundance during the period prior to settlement by people of European descent were not possible (Schroeder et al. 2004). Examination of bones from the late Pleistocene and early Holocene shows that sage-grouse were present in western North America (Braun and Williams 2015; Wolfe and Broughton 2016), but delineation of a distribution from those data is not possible, especially because many of the bones were found outside the documented historical distribution (Braun and Williams 2015). Even with inevitable uncertainty, the association between sage-grouse and sagebrush-dominated habitat is undeniable (Schroeder et al. 2020; Young et al. 2020).
Gunnison sage-grouse were historically found in a relatively small area, including southern Colorado, northern New Mexico, northeastern Arizona, and southeastern Utah (Beck et al. 2003; Schroeder et al. 2004; Braun et al. 2014; Braun and Williams 2015). Greater sage-grouse were distributed more widely and historically found in portions of Arizona, Alberta, British Columbia, California, Colorado, Idaho, Montana, Nebraska, Nevada, North Dakota, Oregon, Saskatchewan, South Dakota, Utah, Washington, and Wyoming (Schroeder et al. 2004; Fig. 10.1). Delineations of historical and current distributions of both species is complicated by the translocations of grouse throughout the range, sometimes even including the translocation of greater sage-grouse into the historical range of Gunnison sage-grouse (Reese and Connelly 1997).
Long-term changes in the distribution of sage-grouse have followed trends in the distribution and quality of sagebrush habitat, typically dominated by big sagebrush (A. tridentata). However, in northeastern portions of the range including Alberta, eastern Montana, Saskatchewan, and North and South Dakota, silver sagebrush (A. cana) is extremely important or the sole source of sagebrush (Aldridge and Brigham 2002; Connelly et al. 2004; Carpenter et al. 2010). Because of declines in habitat quantity and quality, greater sage-grouse no longer occur in British Columbia, Arizona, and Nebraska and Gunnison sage-grouse no longer occur in Arizona and New Mexico (Schroeder et al. 2004; Fig. 10.1). Populations of greater sage-grouse are also dramatically reduced in most states and provinces, but especially Alberta, California, North and South Dakota, Saskatchewan, and Washington (Garton et al. 2011, 2015) where hunting is no longer permitted (Dinkins et al. 2021a). Research using rangewide greater sage-grouse data estimated > 50% extirpation probability for 45.7, 60.1, and 78.0% of leks based on 19, 38, and 56-year projections of population growth from 2019, respectively (Coates et al. 2021b: 3). Most extirpated leks were predicted to be on the periphery of greater sage-grouse range. This study also predicted > 50% extirpation probability for 12.3, 19.2, and 29.6% of populations, defined as neighboring clustered leks, over the same time frames (Coates et al. 2021b: 3).
Gunnison sage-grouse have not been legally hunted since 1999 (Dinkins et al. 2021a). Gunnison sage-grouse are almost extirpated from Utah and greatly reduced in Colorado; their populations are currently so small they have been federally listed as a Threatened species (USFWS 2014). In Canada, greater sage-grouse are federally listed as Endangered under the Alberta Wildlife Act, Saskatchewan Wildlife Act, and under the Canada’s Species at Risk Act (Government of Canada 2021). Greater sage-grouse are not federally listed in the United States (USFWS 2015). Hunting is currently permitted in Colorado, Idaho, Montana, Nevada, Oregon, Utah, and Wyoming (Dinkins et al. 2021a). Sage-grouse have often been viewed as an umbrella species, protecting habitat for up to 350 vertebrates through conservation of their sagebrush habitats (Rowland et al. 2006; Gamo et al. 2013). However, research designed to address this question indicates sage-grouse likely do not serve well in a surrogate role due to mismatches between temporal and spatial scales of seasonal distributions for other species (Carlisle et al. 2018).
The historical and current distribution of greater sage-grouse falls within 9 EPA Level III ecoregions (Wilken et al. 2011; Fig. 10.1). Gunnison sage-grouse were historically found only in the Colorado Plateau and Rocky Mountains ecoregions, with most of their current distribution lying within the Rocky Mountains ecoregion within Utah and Colorado (Fig. 10.1). Greater sage-grouse occur within 720,141 km2 across 10 western states and 2 Canadian provinces and Gunnison sage-grouse occur within 10,036 km2 in Colorado and Utah (Table 10.1). The current distribution of greater sage-grouse has been estimated at 56% of the historical distribution and the current distribution for Gunnison sage-grouse at 10% of its historical distribution (Schroeder et al. 2004).
2.2 Monitoring
The most common and widespread index for greater sage-grouse is based on annual counts of males attending leks (Connelly et al. 2003a, b). Seasonal and daily variation in lek attendance is well-documented (Emmons and Braun 1984; Walsh et al. 2004; Fremgen et al. 2016; Wann et al. 2019) and therefore using the maximum of repeated counts within a standardized period is recommended for more reliable inferences (Connelly et al. 2003a, b; Monroe et al. 2016). However, multiple factors can influence detection including weather (Baumgardt et al. 2017; Fremgen et al. 2019), vegetation and topography (Fremgen et al. 2016), males not attending leks (Blomberg et al. 2013b; Gibson et al. 2014), and age of grouse (Jenni and Hartzler 1978; Walsh et al. 2004; Wann et al. 2019), leading to concerns over the relevance of lek counts to the true population status. Any given count is likely an underestimate of the true population associated with a lek and does not indicate age and sex ratios or males attending unknown leks (Shyvers et al. 2018). Monitoring through mark-resight of leg bands or telemetry can estimate the attendance process (Walsh et al. 2004; Gibson et al. 2014; Fremgen et al. 2016; Wann et al. 2019), but also are more intensive. More recently, researchers have examined use of infrared imagery from aerial surveys to estimate sightability of sage-grouse, given their presence at the lek (Coates et al. 2019). Alternatively, researchers may take advantage of information from repeated counts to estimate the detection process using N-mixture models (McCaffery et al. 2016; Monroe et al. 2019). Due to the open nature of sage-grouse lek attendance, inferences from this modeling approach can be extended at most to the number of males attending a lek at least once in a season (Nichols et al. 2009). Indices may still offer useful inferences of population trends when detectability is constant or random over time (Johnson 2008; Monroe et al. 2016), but fail if detection covaries with population trends (Monroe et al. 2019; Blomberg and Hagen 2020). Additionally, the area used by sage-grouse attending leks is unknown and can vary, and not all active leks in a landscape are known to observers. A dual-frame approach may help jointly estimate both the number of active leks and the number of males attending leks, providing a more accurate estimate of the number of males across a landscape (Shyvers et al. 2018). Genetic data from non-invasive sources can be used to estimate winter (pre-breeding) population size with a mark-recapture approach (Shyvers et al. 2019).
Due to the cryptic nature of this species, use of radio telemetry and global positioning systems (GPS) to monitor habitat use and survival is common (e.g., Aldridge and Boyce 2007; Smith et al. 2018a). However, this type of monitoring can incur substantial costs to researchers and management agencies, both financially and in potential bias from monitoring units themselves (Fremgen et al. 2017; Severson et al. 2019). Indeed, stage-specific data on demographic parameters for this species are relatively limited, particularly for chick and juvenile survival (Taylor et al. 2012), and other data sources may be needed. For example, hunter-harvest data could be used to estimate survival rates, sex ratios, and recruitment metrics (Braun and Schroeder 2015; Hagen et al. 2018; Wann et al. 2020), and annual counts of sage-grouse broods may provide an index of productivity (Connelly et al. 2003a, b). Integrated population models (IPMs) which combine demographic information with lek count data may improve estimates of sage-grouse population size and other demographic parameters (Davis et al. 2014a; McCaffery and Lukacs 2016; Coates et al. 2018).
3 Habitat Associations
3.1 Historical/Evolutionary
Natural variation in the historical distribution of sage-grouse has occurred because of climate fluctuations, specifically in relation to elevational gradients. For example, warmer and drier sagebrush habitats at lower elevations tend to have a smaller component of native herbaceous cover when compared with higher elevation habitats. As a result, some of these lower elevations have reduced capacity to support sage-grouse. This can be illustrated by the decline in sage-grouse in the Bonneville Basin in Utah during a 4000-year period as the climate warmed and dried (Wolfe and Broughton 2016). Furthermore, declines in sage-grouse abundance (Connelly et al. 2004; Garton et al. 2011) and occupancy (Schroeder et al. 2004) increases fragmentation and isolation of populations. This loss of connectivity has dramatic demographic and genetic consequences that can reduce population viability (Oyler-McCance et al. 2005; Oyler-McCance and Quinn 2011), thus further decreasing populations and distribution.
3.2 Contemporary
Declines in sage-grouse populations indicate that identification of important habitats, especially breeding habitats, is critical for long-term conservation of sage-grouse, as most year-round activity for greater sage-grouse populations occurs within 8 km of active leks (Fedy et al. 2012; Coates et al. 2013). Doherty et al. (2016) delineated breeding habitat for greater sage-grouse and Doherty et al. (2018) delineated breeding habitat for Gunnison sage-grouse. For both species of sage-grouse, breeding habitat probabilities ≥ 65% provided a threshold to predict areas where leks occur (predicted breeding habitat; Doherty et al. 2016, 2018; Fig. 10.3). Predicted breeding habitat overlapped more (range = 15.6–24.7%) with federal lands than on other ownerships including private, state, and tribal in the U. S. within Western Association of Fish and Wildlife Agencies (WAFWA) Management Zones (MZs) II (Wyoming Basin), III (Southern Great Basin), IV (Snake River Plain), and V (Northern Great Basin; Doherty et al. 2016; Table 10.2). Predicted breeding habitat for greater sage-grouse overlapped more with private compared to public or tribal lands in MZs I (Northern Great Plains), VI (Columbia Basin), and VII (Colorado Plateau; Table 10.2). Only 14.8% (8.4% federal, 5.9% private, and 0.5% of state) of MZ VII was predicted breeding habitat for Gunnison sage-grouse in MZ VII (Table 10.2), with the majority located in the Gunnison Basin of Colorado (Doherty et al. 2018; Fig. 10.3).

Greater and Gunnison sage-grouse breeding habitat probabilities in the western United States. Inset map represents a detail of breeding habitat probabilities for distribution of Gunnison sage-grouse. Areas with > 65% breeding habitat probability indicate predicted breeding habitat suitable to support lek formation. The models that generated these predictions were originally created from separate analyses focused on greater (Doherty et al. 2016) and Gunnison (Doherty et al. 2018) sage-grouse. Models used data from 2010–2014 for greater and 2015 for Gunnison sage-grouse
4 Rangeland Management
We do not provide specific prescriptions for practitioners to manage or restore rangelands for sage-grouse. However, we do synthesize important issues related to Rangeland Management (Sect. 10.4; livestock grazing and grass height, improper grazing, mesic resources, fencing, habitat alteration treatments, feral equids, and ravens) and Ecosystem Threats (Sect. 10.6; fire, invasion from exotic annual grasses, conifer encroachment, sagebrush conversion and seeding introduced grasses, exurban development, energy development, and climate change). Topics listed under both sections highlight issues, describe concepts, and provide insights from relevant literature to assist in providing knowledge applicable to managing and restoring sage-grouse habitats at appropriate local scales. We encourage practitioners to access publications or other resources specifically developed to guide efforts to restore sage-grouse habitats such as Pyke et al. (2015a, b, 2017).
4.1 Livestock Grazing and Grass Height
Potential responses from sage-grouse to livestock grazing management have been hypothesized, because positive correlations between sage-grouse nest site selection and survival and grass height have been documented (Holloran et al. 2005; Doherty et al. 2014; Gibson et al. 2016a). Indeed, the 2014 Threatened listing decision for Gunnison sage-grouse suspected that failure of multiple allotments meeting relevant land health standards might have negatively affected the species (USFWS 2014). One such habitat metric is grass height, which is incorporated in habitat management guidelines (Connelly et al. 2000; Stiver et al. 2015), but the relevance of this parameter is increasingly questioned. The first criticism is methodological; due to phenology and vegetation growth during a season, researchers measuring grass height at nesting outcome (fail or fledge) inherently induce a bias in their measurements, where grass height is often lower for failed nests (Gibson et al. 2016b; Smith et al. 2018b). Correcting for this discrepancy resulted in more modest, and variable, relationships between sage-grouse nest survival and grass height (Gibson et al. 2016b; Smith et al. 2018b). Secondly, habitat associations are often context-dependent, and there is risk in extrapolating results from individual, localized studies to other parts of the sage-grouse range (Smith et al. 2020). Finally, while attention has been focused on potential effects of grazing on sage-grouse nest habitat and survival, livestock may influence other components of sage-grouse life history such as brood-rearing habitat. For example, habitat selected for nesting may correlate with subsequent brood survival (Gibson et al. 2016a), and a diverse diet of abundant plants and invertebrates may benefit sage-grouse productivity (Blomberg et al. 2013a; Smith et al. 2019). However, there is a lack of empirical support for the hypothesis that livestock grazing manipulates food availability for sage-grouse, and, in turn influences sage-grouse vital rates. This is a compelling hypothesis, though, as research from central Montana found greater insect diversity and activity-density of arthropods eaten by sage-grouse in sagebrush grazed by livestock under rest-rotation as compared to lands ungrazed for over a decade (Goosey et al. 2019).
Studies examining responses of sage-grouse populations to grazing are also still limited, likely because of the large areas sage-grouse use for their life history (Fedy et al. 2014). In one of the few grazing studies, based on public lands records from across Wyoming, Monroe et al. (2017) estimated a negative response to higher levels of reported grazing early in the growing season among male sage-grouse attending leks and a positive response when grazing occurred later. This relationship was apparent in areas where vegetation productivity was low, but not in high productivity areas. A field experiment study based on research conducted on private ranches in central Montana indicated a positive trend in daily survival rates for greater sage-grouse nests on ranches that implemented rotational livestock grazing, yet support for this effect was weak (Smith et al. 2018a). This study did not find grazing rest of 1 year or greater increased daily survival rates for nests and rotational grazing and rest compared to other grazing strategies in the area had negligible effects on herbaceous vegetation height and cover. These authors concluded that grazing strategies played a minor role in sage-grouse nest success relative to other factors such as climate and predators in the northern Great Plains (Smith et al. 2018a).
4.2 Improper Grazing
Improper livestock grazing is generally heavy, repeated grazing, particularly in the spring, of sagebrush communities and can be detrimental to large perennial bunchgrasses and favor exotic annual grasses, particularly in sites with lower resilience and resistance (Stewart and Hull 1949; Daubenmire 1970; Mack 1981; Knapp 1996). Improper grazing can negatively affect sagebrush communities by altering vegetation composition and structure. Some of the most significant effects of improper livestock grazing are related to its interaction with other factors. For example, improper grazing often depletes the perennial herbaceous component and increases sagebrush cover, but these communities may not convert to exotic annual grasslands until a wildfire removes the sagebrush and creates a pulse in resource availability (Davies et al. 2016a). In contrast to repeated heavy grazing, moderate levels of grazing with periods of growing season deferment and rest may not negatively impact sagebrush communities (West et al. 1984; Davies et al. 2018; Copeland et al. 2021) and may even decrease their risk of converting to exotic annual grasslands after burning by reducing fire severity (Davies et al. 2009, 2015a, 2016b).
4.3 Mesic Resources
Mesic resources include moist areas near springs, creeks, ponds, reservoirs, and wet meadows, which promote herbaceous production near water, attracting livestock and sage-grouse during summer as upland sites dry and herbaceous plants senesce (Connelly et al. 2011b; Swanson et al. 2015). Late brood-rearing habitat is more limited than early brood-rearing habitat, composing an estimated 5% of sage-grouse habitat in a study area in southern Alberta (Aldridge and Boyce 2007), and 2.4% of sage-grouse habitat in California, northwest Nevada, and Oregon (Donnelly et al. 2016). Furthermore, late brood-rearing habitat where broods survived composed only 2.8% of a study area in central Nevada (Atamian et al. 2010). Private lands are particularly important in providing mesic resources to sage-grouse, because 60% of sage-grouse habitat occurs on public lands, yet 68% of mesic sites available to sage-grouse occur on private lands (Donnelly et al. 2016, 2018). Sage-grouse population productivity, based on lek distribution and attendance data, increased with proximity to mesic sites for greater sage-grouse in California, northwest Nevada, and Oregon (Donnelly et al. 2016). Anecdotal evidence from earlier Nevada studies indicated moderate cattle grazing in mesic meadows induced use by sage-grouse because cattle herbivory exposed preferred forbs (Neel 1980; Evans 1986). Recent research from Idaho indicates use of early-season, high-intensity cattle grazing increases cover and biomass of high-value forbs used by sage-grouse in mesic meadows (Randall et al. 2022). Practices to restore meadows and riparian areas within the sagebrush biome include Zeedyk structures, beaver dam analogs, and grazing management—these practices may increase productivity of vegetation in sage-grouse brood-rearing habitats by 25% (Silverman et al. 2019). Conservation and management of mesic resources is imperative to maintain sage-grouse populations in the face of climate-driven variability in vegetation conditions (Donnelly et al. 2018).
4.4 Fencing
Structures associated with livestock production can be detrimental to sage-grouse. For example, fencing located near leks may increase collisions (Stevens et al. 2012b; Van Lanen et al. 2017), particularly in relatively flat topography (Stevens et al. 2012a; Fig. 10.4). Use of fence markers and wooden fence posts spaced < 4 m (13.1 feet) apart may reduce collisions (Stevens et al. 2012a; Van Lanen et al. 2017). Another option to consider is to move or remove fences in high-risk areas (Stevens et al. 2012b). Managers may also consider marking fences in sage-grouse breeding habitats when fence densities exceed 1 km/km2 (0.62 mile/mile2) within 2 km (1.2 miles) of active leks in areas with flat to gently rolling terrain (Stevens et al. 2012a). Marked compared to unmarked fences in Idaho reduced risk of fence collision by greater sage-grouse approximately 83% (Stevens et al. 2012b), and, in Wyoming, marking fences, regardless of marker type, reduced collisions approximately 57% (Van Lanen et al. 2017).

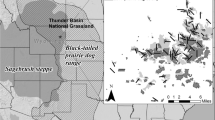
Illustration depicting major threats to sage-grouse and their habitat. Multiple threats that may constrain sage-grouse populations are indicated by arrows pointing at an adult male, adult female, and chick sage-grouse in the center of the illustration. Land use and development threats can occur at any elevation, may occur solely, or in combination and are not ubiquitous across the range of sage-grouse. Land use and development threats include effects from renewable energy (e.g., wind energy), non-renewable energy (e.g., oil and gas development), linear features (e.g., fences, roads, and transmission lines), exurban residential development, increasing predators (e.g., common ravens that utilize developed areas to a higher degree), and feral horses. Persistent ecosystem threats related to exotic annual grasses, conifer encroachment and fire occur rangewide. Resistance to invasive annual grasses and resilience to disturbance increases with elevation owing to increased precipitation and cooler soil temperatures (Chambers et al. 2014, 2017). Wyoming big sagebrush dominates lower elevations where invasion by exotic annual grasses create continuous fuel beds, leading to increased fire frequency and reducing Wyoming big sagebrush. Encroaching conifers at higher elevations outcompete mountain big sagebrush and herbaceous plants (see Davies et al. 2011). Figure created by Emilene Ostlind, University of Wyoming
4.5 Habitat Alteration Treatments
Improper vegetation treatments are also a concern in sagebrush communities, especially brush management in lower elevation sagebrush communities, which are hotter and drier than those at higher elevations are. Attempts to improve sage-grouse and other wildlife habitat by reducing sagebrush cover in Wyoming big sagebrush communities has not achieved desired results of enhancing habitat conditions that bolster populations (Beck et al. 2012). Reducing sagebrush in these communities also often substantially increases exotic annual grass and forb abundance and cover (Davies et al. 2012; Davies and Bates 2014). Thus, sagebrush control in Wyoming big sagebrush communities may facilitate conversion to an exotic annual grassland. Manipulating sagebrush to benefit sage-grouse populations is largely untested (but see Dahlgren et al. 2006, 2015; Smith and Beck 2018; Smith et al. 2023), and cumulative effects of treatments can be detrimental to populations (Dahlgren et al. 2015). Effects likely vary by sagebrush treatment type and scale, with negative responses for mechanical and prescribed fire but neutral-to-positive long-term responses to chemical reduction, where many structural components of sagebrush are retained (Smith and Beck 2018).
4.6 Feral Equids
Negative ecological effects, mainly from increasing feral horse (Equus ferus caballus), and, in limited areas, burro (E. asinus) populations, on federal public rangelands are a growing concern for sage-grouse habitats (see Chap. 21: Feral Equids; U. S. Fish and Wildlife Service 2013; Beever et al. 2018; Scasta et al. 2018). Rangewide, about 12% of sage-grouse habitat is also managed for feral equids. However, the amount of overlap varies among states, reaching a high of > 99% overlap where feral horses occur on Bureau of Land Management (BLM) lands in Wyoming (Beever and Aldridge 2011). Multiple direct and indirect effects have been hypothesized regarding how feral equids may influence sage-grouse life stages and habitat characteristics (Beever and Aldridge 2011). Field studies on feral horses have reported modification of habitat quality through increased bare ground and reduced grass height (Hennig et al. 2021), propagating cheatgrass seeds via feces (King et al. 2019), and disruption of male sage-grouse attending leks (Muñoz et al. 2021). Managing feral horses at or below appropriate management levels set by BLM is consistent with maintaining sage-grouse populations at levels where sage-grouse do not overlap with feral horses (Coates et al. 2021a). However, when horse numbers are 2, 2.5, and ≥ 3 times over maximum appropriate management levels, probability of sage-grouse population decline relative to controls is 76%, 97%, and > 99%, respectively (Coates et al. 2021a). These predictions indicate properly managing feral horse numbers that do not exceed appropriate management levels is most harmonious with maintaining sage-grouse populations where the two species share habitat.
4.7 Ravens
Common ravens (Corvus corax ‘hereafter, ravens’) are a major predator of sage-grouse nests (Dinkins et al. 2016a; Conover and Roberts 2017). During the last half century, ravens have expanded their distribution and increased in abundance in central and western North America (Dinkins et al. 2021b; Harju et al. 2021). Abundance of ravens from 1995 to 2014 was highest in western and southeastern WAFWA MZs (III, IV, V, VI, and VII), and ravens were expanding into and increasing in MZs I and II in the northeast (Dinkins et al. 2021b). High abundance of ravens in MZ VII indicates the Threatened Gunnison sage-grouse has been exposed to high raven numbers for a few decades (Dinkins et al. 2021b; Harju et al. 2021). Occurrence of ravens increases with presence of livestock and associated infrastructure, such as buildings and water sources (Coates et al. 2016a; Fig. 10.4). Increases in raven populations are related to their ability to exploit anthropogenic resource subsidies such as food, perches, and nesting structure in rangelands, uncoupling them from the availability of local indigenous resources (Boarman 2003). Adult female sage-grouse avoid nesting in habitat with higher densities of ravens (Dinkins et al. 2012). Predictive modeling across rangewide sage-grouse habitat indicates higher growth rates for ravens in landscapes with greater transmission line density. Dinkins et al. (2021b) found carrying capacity for ravens was higher with increasing proportion of urban land cover within 25 km and burned area within 3 km, and negatively correlated with greater distance from landfills and proportion of forest cover within 15 km. Management actions to reduce effects of ravens on sage-grouse populations include removing nesting structures and food sources, eliminating or covering landfills, and restoring burned sagebrush (Dinkins et al. 2021b).
5 Effects of Disease
5.1 General Concerns for Populations
Sage-grouse host a variety of potentially pathogenic organisms including macroparasitic arthropods (e.g., lice, ticks), helminths (e.g., nematodes, cestodes, trematodes), and microparasites (protozoa, bacteria, fungi and viruses; Christiansen and Tate 2011; Sinai et al. 2017). Various non-parasitic diseases or disease-like conditions can also affect sage-grouse (Christiansen and Tate 2011). Most macro- and microparasites, and infectious diseases documented in sage-grouse have not resulted in widespread population level effects to sage-grouse (Christiansen and Tate 2011). However, researchers have conducted few systematic surveys for parasites or pathogens in sage-grouse.
West Nile virus (WNV; Flaviviridae, Flavivirus) has emerged as a threat to sage-grouse (Walker and Naugle 2011). In addition to WNV, avian infectious bronchitis virus and other avian coronaviruses, avian retroviruses, Mycoplasma spp., and the Eimeria coccidians and associated enteric bacteria may be subject to amplification by climate change or anthropogenic disturbance or have a history of impacting sage-grouse. These historic and emerging risks indicate further monitoring and research of diseases and parasites in sage-grouse is warranted as suggested by Peterson (2004) for prairie grouse.
5.2 Diseases as Associated with Livestock
There are few direct disease relationships known to occur between rangeland livestock and sage-grouse. However, surface water, an important component of rangeland management, plays a role in the exposure of sage-grouse to WNV. Coccidiosis, tapeworms, and toxicosis are also discussed due to their local significance in the past and potential, though unlikely, risks to sage-grouse populations in the future.
West Nile Virus. West Nile virus, a mosquito-borne flavivirus, is recognized as an important source of mortality to sage-grouse in elevations below 1500 m, which is approximately 40% of current sage-grouse range (Walker and Naugle 2011). Few live sage-grouse have tested seropositive for WNV antibodies indicating sage-grouse rarely survive infection (Walker and Naugle 2011; Dusek et al. 2014). Population viability analyses indicate that local populations may be vulnerable to extirpation from even a single stressor, such as WNV (Taylor et al. 2013). Additionally, West Nile virus can have cascading effects as evidenced by a quadrupling of lek inactivity when populations already impacted by energy development are exposed to a West Nile virus outbreak (Taylor et al. 2013).
Livestock reservoirs, even water-filled hoof prints, can serve as breeding habitat for vector mosquitoes (Doherty 2007). The mesic areas created by the reservoirs attract sage-grouse in mid to late summer (Connelly et al. 2011b), during the peak of WNV transmission. Eliminating mosquito-breeding habitat or controlling mosquito larval populations in anthropogenic water sources can reduce effects of WNV on sage-grouse populations (Zou et al. 2006; Walker and Naugle 2011; Watchorn et al. 2018).
Summer temperatures may affect WNV viremia (Naugle et al. 2005; Walker and Naugle 2011) and vector mosquito autogeny (Brust 1991) indicating increasing temperatures associated with climate change may increase WNV risk to sage-grouse. Mammals, particularly humans and horses, can become infected through mosquito bites and represent dead-end hosts of WNV infection (Ahlers and Goodman 2018).
Coccidiosis. Prior to the emergence of West Nile virus, coccidiosis (Eimeria spp.) was the most important known disease of sage-grouse. Losses of young sage-grouse were documented in several states from 1932 to 1953, typically in areas where up to 2000 (estimated) birds congregated, resulting in fecal contamination of soil and water (Honess and Post 1968). Sporadic occurrence of coccidiosis-associated morbidity and mortality in individual birds is reported; however, notable mortality events attributed to coccidiosis in sage-grouse have not been documented since the early 1960s. This change in disease dynamic may be the result of decreased sage-grouse densities. Livestock are also susceptible to coccidiosis, but the infectious species of Eimeria are host specific (López-Osorio et al. 2020).
Tapeworms. The most visually apparent parasites of sage-grouse are tapeworms (Order: cestoda), which are commonly reported by hunters and field personnel. Sage-grouse show no apparent clinical signs of infection and may reflect an almost perfect adjustment between the host and its parasite (Honess 1982). However, “heavy” burdens of tapeworms could have direct and/or indirect adverse effects on individual birds, such as intestinal occlusion, reduction in vigor, and increased susceptibility to other parasites (Cole and Friend 1999). Livestock are also susceptible to tapeworms, but not the species that infect sage-grouse.
Toxicosis. During 1949–50, 1.7 million ha of Wyoming rangeland were aerially treated with Toxaphene and Chlordane bran bait to control grasshoppers (Family: Acrididae; Post 1951). Post (1951) reported game bird mortality and toxemia on treated areas. The scale and toxicity of grasshopper control efforts during this time indicate widespread but unquantified negative effects to sage-grouse populations. Neither Chlordane nor Toxaphene have been registered for grasshopper control since the early 1980s. Modern pesticides applied via Reduced Agent and Area Treatments approach (Lockwood et al. 2002) likely minimize effects.
Ivermectin, a broad-spectrum antiparasitic, is routinely and globally used to control parasitic worms of ruminant animals. Field studies have demonstrated the dung of animals treated with ivermectin influences abundance and ecology of invertebrates (Martinez et al. 2017; Finch et al. 2020). The direct or indirect effects of ivermectin (or similar compounds) to sage-grouse is unknown, but research in central Montana showed dramatically lower dung beetle activity-density on lands with managed grazing than on idled land, which were hypothesized to be due to anti-parasitic drugs like ivermectin (Goosey et al. 2019). However, sage-grouse food arthropods were still collected in higher numbers, overall, on managed than on idled land (Goosey et al. 2019).
6 Ecosystem Threats
Sagebrush ecosystems are experiencing numerous landscape-scale threats that decrease the quantity and quality of sagebrush communities (Davies et al. 2011). Historically, the sagebrush ecosystem occupied over 62 million hectares in western North America. By the early 2000s, it was estimated that sagebrush occupied less than 60% of its historical range and many remaining sagebrush communities were fragmented and degraded (Knick et al. 2003; Schroeder et al. 2004). The loss, degradation, and fragmentation of sagebrush occupied rangeland has continued with altered fire regimes, anthropogenic development, conifer encroachment, exotic annual grass invasion, climate change, and conversion to croplands and introduced grasslands. Many of these stressors are interrelated, with one threat exacerbating another, creating feedback loops. Habitat loss is viewed as the primary reason for declines in sage-grouse populations. For example, greater sage-grouse in Canada now only inhabit 7% of their historical distribution, primarily due to habitat loss from agricultural development and placement of anthropogenic features such as oil and gas wells in remaining habitats (Government of Canada 2021).
6.1 Altered Fire Regimes
Sagebrush ecosystems are experiencing widespread alteration to historical fire regimes. Fire frequency has increased in many hotter and drier sagebrush communities, driven mainly by exotic annual grass invasion (Balch et al. 2013) as well as increased human-caused ignitions (Bradley et al. 2018). Frequent fire prevents reestablishment of sagebrush and is detrimental to many native perennial species. In contrast, fire frequency has decreased in cooler and wetter sagebrush communities substantially because of fire suppression and historical heavy grazing (Miller and Wigand 1994; Miller and Rose 1999). Decreased fire frequency has allowed conifer woodlands to establish across many sagebrush communities, which further decreases fire frequency due to reduced fine fuel loads in woodland understories. Periodic fire in these cooler and wetter sagebrush communities is also important for promoting spatial and temporal heterogeneity of habitats (Davies and Bates 2020).
6.2 Invasion from Exotic Annual Grasses
Sagebrush plant communities are experiencing undesirable shifts in vegetation composition and structure from invasive species. Exotic annual grasses are the primary threat in hotter and drier sagebrush communities (Davies et al. 2011; Chambers et al. 2014) and native conifers are encroaching into cooler and wetter sagebrush communities (Miller and Wigand 1994; Miller et al. 2005). These two undesirable vegetation shifts largely occur in different sagebrush community types, but conifer encroachment and exotic annual grass invasion appear to be overlapping more frequently in recent decades (Davies et al. 2011) as annual grasses spread to higher elevations and north-facing aspects consistent with predictions of climate warming (Smith et al. 2022).
Exotic annual grasses, primarily cheatgrass (Bromus tectorum) and medusahead (Taeniatherum caput-medusae) have invaded tens of millions of hectares of the sagebrush ecosystem, particularly in the western portion of sage-grouse range (Meinke et al. 2009; Bradley et al. 2018). Cheatgrass, alone is estimated to be present in high abundance across almost a third (21 million ha) of the Great Basin (Bradley et al. 2018), primarily in sagebrush communities. Exotic annual grass invasion often triggers an exotic annual grass-fire cycle, where abundant exotic annual grass fuel promotes frequent fire, further decreasing native perennial vegetation (D’Antonio and Vitousek 1992; Balch et al. 2013). This cycle results in exponential declines in native plants and biodiversity as exotic annual grasses increase in abundance (Davies 2011). Currently there are no cost-effective treatments to control exotic annual grasses across the vast area of invasion (Stohlgren and Schnase 2006).
6.3 Conifer Encroachment
Conifers, primarily juniper (J. osteosperma, J. occidentalis, J. scopulorum) and piñon pine, in the sagebrush ecosystem were historically confined to fire-safe sites (i.e., rocky slopes and ridges with insufficient understory to carry surface fires) or occurred in open savannah-like stands (Romme et al. 2009). Decreased fire frequency has allowed conifers to expand into more productive sagebrush communities and increase in density, particularly in the western and southern regions of the sagebrush biome (Miller and Wigand 1994; Miller et al. 2005). Juniper and piñon woodlands currently occupy ~ 19 million ha in the Intermountain West; up to 90% of this area was historically sagebrush communities (Tausch et al. 1981; Miller et al. 2008). Conifer encroachment eliminates sagebrush and decreases the herbaceous understory, which can result in substantial erosion risk (Miller et al. 2000; Pierson et al. 2007). Conifer encroachment into sagebrush communities is especially detrimental to sage-grouse populations for several reasons. Greater sage-grouse will avoid sagebrush communities with as little as 4% tree cover (Baruch-Mordo et al. 2013; Severson et al. 2016). Conifer cover as low as 1.5% negatively influences survival for adult female greater sage-grouse (Coates et al. 2017) and increases risk of daily mortality, especially in juvenile and yearling birds, when navigating conifer-invaded sagebrush habitats (Prochazka et al. 2017). Population growth rates for greater sage-grouse are lower in conifer-invaded areas than adjacent areas where conifers have been removed (Olsen et al. 2021). In addition, simulated removal of conifer coverage up to 30% within a 0.56-km2 scale was predicted to increase high-quality Gunnison sage-grouse breeding habitat fourfold (Doherty et al. 2018). Despite unprecedented efforts in the sagebrush biome, conifer removal is barely keeping pace with its rate of expansion (Reinhardt et al. 2020).
6.4 Sagebrush Conversion and Seeding Introduced Grasses
The most arable sagebrush communities in the United States have been converted to cropland, thus the continued cultivation of intact landscapes produces marginal yields at high costs to wildlife. Crested wheatgrass (Agropyron cristatum and A. desertorum) has been seeded on 6–11 million hectares of rangelands in western North America, much of which was originally native sagebrush communities (Lesica and DeLuca 1996; Hansen and Wilson 2006). Crested wheatgrass is often seeded after wildfire in former sagebrush communities largely because of its ability to effectively compete with exotic annual grasses (Arredondo et al. 1998; Davies et al. 2010), but also because it is less expensive and often establishes more successfully than native bunchgrasses in hotter and drier sagebrush communities (Asay et al. 2003; James et al. 2012; Davies et al. 2015b). However, due to its ability to outcompete other species, crested wheatgrass can form monotypical plant communities, resulting in less value as wildlife habitat and lower overall biodiversity (Christian and Wilson 1999; Heidinga and Wilson 2002; Hamerlynck and Davies 2019).
Currently there exists a management conundrum that introduced grasslands are less desirable than native sagebrush rangelands, but native seedings using conventional seeding techniques frequently fail within sagebrush communities that have low resilience and resistance to exotic annual grass invasion (Knutson et al. 2014). Furthermore, exotic annual grasslands have lower habitat quality than introduced grasslands and the development of exotic annual grasslands increases the probability that surrounding areas will convert to exotic annual grasslands. For these reasons, introduced grasses are likely to continue to be seeded after fires in sagebrush communities with substantial risk of exotic annual grass dominance until native species establishment is improved. However, introduced grasses should not be seeded in more resilient and resistant sagebrush communities where seeded native species can successfully establish and persist (e.g. Davies et al. 2019; Urza et al. 2019).
6.5 Exurban Development
Exurban development is the process of dividing large parcels of mostly undeveloped lands into residential lots and is the fastest growing form of land conversion across the United States (Brown et al. 2005). As human population growth has continued in the western states and provinces, large expanses of the sagebrush ecosystem have seen negative effects from human development and subsequent habitat fragmentation. Exurban development directly converts native habitat, fragments habitat, and degrades remaining habitat for native species. This results in declines in native plant and animal diversity, increases in exotic species, and greatly limits the use of ecosystem management tools to achieve landscape level effects (Knight et al. 1995; Maestas et al. 2003; Hansen et al. 2005). Larger human populations are also tied to increased fire frequency in Mediterranean-climate ecosystems (Syphard et al. 2009), thus exurban development of sagebrush communities will likely increase fire frequency in some areas, further promoting exotic annual grass invasion and dominance.
6.6 Energy Development
Energy development has fragmented and degraded sagebrush communities in many western states and provinces (Bergquist et al. 2007; Lyon and Anderson, 2003; Naugle et al. 2011). This threat to the sagebrush ecosystem has increased during the twenty-first century with greater demand for renewable energy sources like wind and solar energy (Kiesecker and Naugle 2017). Influences of energy development are expected to rise as the United States continues to increase its domestic energy production (Doherty et al. 2010). For example, the land occupied by well pads, roads, and other facilities from recent (2000–2012) expansion of oil and gas extraction in North America is estimated at 3 million ha (Allred et al. 2015). This is not exclusive to sagebrush communities but highlights the threat of further energy development to these communities.
Energy extraction and development can cause high levels of fragmentation of sagebrush landscapes. In areas of northeastern Wyoming, every 1 km2 was bisected by a powerline and bounded by a road where energy development and agricultural production occurred (Naugle et al. 2011). Infrastructure for energy extraction and development including roads, pipelines, earthen dams, and well pads create substantial surface disturbance and are vectors for exotic plant invasions within sagebrush communities, contributing to further degradation (Gelbard and Belnap 2003; Bergquist et al. 2007). Sage-grouse typically respond to fragmented landscapes by avoiding human infrastructure (e.g., Aldridge and Boyce 2007; Dinkins et al. 2014; Kirol et al. 2015). For example, translocated female sage-grouse in Alberta were more likely to select habitat with increasing distance from infrastructure, up to 2.5 km from roads, 3 km from trees and gas wells, 10 km from buildings, 15 km from settlements, and at least 23 km from power lines (Balderson 2017).
6.7 Climate Change
Climate change is expected to negatively affect the sagebrush ecosystem. Reduced snowpack and earlier snowmelt (Klos et al. 2014; Harte et al. 2015) and longer, more arid summer conditions leading to drier soils (Palmquist et al. 2016) are factors predicted to lead to future reduction in coverage of sagebrush and associated plant species, altering sage-grouse habitat (Homer et al. 2015). Cultivation of sagebrush habitats in the eastern range of sage-grouse has completely altered carbon storage (Sanderson et al. 2020). Retaining and restoring sagebrush habitats represents the single largest natural opportunity to maintain carbon storage in rangelands (Fargione et al. 2018). Farther west, most models of climate change predict warmer winters and altered precipitation patterns, as well as an earlier onset of fire season and more wildfires, all conditions that perpetuate exotic annual grasses (Abatzoglou and Kolden 2011; Creutzburg et al. 2015). When these models are applied to the landscape, cheatgrass cover is predicted to remain stable or increase in much of the Great Basin for the next 50 years (Boyte et al. 2016). In addition, increasing atmospheric CO2 concentrations may increase exotic annual grass productivity and litter retention, leading to increased fuel loads, which may increase fire intensity and frequency (Ziska et al. 2005). Ongoing increases in cheatgrass and other exotic annual grasses and associated increased fire frequency will likely further reduce the area occupied by sagebrush. Furthermore, climate change predictions indicate sagebrush may not be viable to its entire historical range and may experience a distribution shift (Schlaepfer et al. 2015). Future climate scenarios indicate loss of sagebrush, and ultimately sage-grouse habitat, in some areas (Schlaepfer et al. 2012; Homer et al. 2015; Palmquist et al. 2016; Renne et al. 2019), as was historically documented with a warmer, drier climate in the Bonneville Basin of Utah (Wolfe and Broughton 2016). Increased drought conditions will likely also be detrimental to sage-grouse demographics (Blomberg et al. 2014).
7 Conservation and Management Actions
7.1 Private Lands
Approximately one-third of western rangelands are privately owned and managed, encompassing some of the most productive sage-grouse habitats (Donnelly et al. 2016). Grazing by domestic livestock is the common thread that maintains open spaces of intact rangelands at ecosystem scales, hereafter referred to as working rangelands. Working rangelands connect a checkerboard of public lands that together provide ecological footprints large enough to sustain sage-grouse populations and rural communities. In the 1930s the father of modern conservation, Aldo Leopold, said that ‘conservation will ultimately boil down to rewarding the private landowner who conserves the public interest (Flader et al. 1992: 202),’ a prognosis still true today. Partnerships are the cornerstone of private lands conservation, with many like-minded, landowner-led groups coalescing into umbrella organizations to offer their shared vision of wildlife conservation through sustainable ranching. State, federal, and non-governmental partners commonly provide human and financial resources to implement beneficial conservation practices on ranchers’ private operations.
The largest partnership is the Sage Grouse Initiative (SGI), which launched by the USDA Natural Resources Conservation Service (NRCS) in 2010, assists agricultural producers who volunteer to reduce threats facing sage-grouse on working rangelands. In 2012, the SGI served as the flagship for the establishment of Working Lands for Wildlife (WLFW), an effort to conserve other at-risk ecosystems and associated species. The NRCS employs the $60 billion conservation title of the federal Farm Bill legislation (title II of the Agriculture Improvement Act of 2018, FY2019–FY2023) to help landowners voluntarily implement conservation practices on private farms, ranches, and forestlands. Quantifying the outcomes of resulting sage-grouse conservation (Naugle et al. 2019; NRCS 2021) and the iterative use of emerging science to improve delivery are integral components of this effort (Naugle et al. 2020). Over the last decade, the SGI has become the primary catalyst for science-driven sagebrush conservation by using Farm Bill resources to restore or enhance more than 29,300 km2 of sage-grouse habitat on more than 1850 ranches, while supporting sustainable agricultural productivity on these working lands. Private lands conservation was featured prominently in the most recent listing decision for greater sage-grouse, placing voluntary conservation on par with regulatory mechanisms on public lands.
With WLFW codified nationally in the 2018 Farm Bill, NRCS continues to contribute to conservation of working rangelands as part of the Sagebrush Conservation Strategy administered by WAFWA. Ongoing contributions strategically target removal of expanding conifer, restoration of riparian areas and wet meadows, and reduction of cultivation, exurban sprawl, and cheatgrass invasion. Recent removal of expanding conifer in > 3300 square kilometers of priority sage-grouse habitats in places like southern Oregon have increased the population growth rate of sage-grouse by + 12% (Olsen et al. 2021) and doubled the abundance of sagebrush songbirds (Holmes et al. 2017). Riparian and wet meadow restorations on working lands using Zeedyk structures, beaver dam analogs, and grazing management have increased by 25% the productivity of sage-grouse brood habitats (Silverman et al. 2019). The strategic placement of conservation easements to alleviate in perpetuity the threat of cultivation and subdivision in places like northcentral Montana also have conserved the longest known migration corridors of sage-grouse and pronghorn (Antilocapra americana) in North America (Tack et al. 2019). Targeting tools created from the new NRCS-sponsored Rangelands Analysis Platform (https://rangelands.app) provide an integrated approach for reducing effects of cheatgrass (Western Governors’ Association 2020).
7.2 Public Lands
Approximately two-thirds of sage-grouse habitat in the United States occurs on public lands, including lands managed by the BLM (51%), the U.S. Forest Service (USFS; 8%), and States (5%; Knick 2011). The majority of land inhabited by sage-grouse in Alberta and Saskatchewan is provincially owned and leased for grazing or contained within Grasslands National Park. Both active leks (as of 2021) in Saskatchewan lie within Grasslands National Park and of the 3 active leks in Alberta, 2 occur on provincial and 1 on private land (J. T. Nicholson, written communication, Alberta Environment and Parks, 2022). The Western Association of Fish and Wildlife Agencies delineated seven Management Zones (MZs) across the sage-grouse range (Stiver et al. 2006; Tables 10.1 and 10.2; Fig. 10.3), and these are distinguished as floristic provinces based on climatic, elevational, topographic, and edaphic characteristics (Connelly et al. 2004). Within each Management Zone in the U.S., areas are prioritized for sage-grouse conservation through coordination of local working groups and by multiple agencies including at the federal and state levels with habitat issues being primarily managed by the BLM and USFS on federal lands and each state on state-owned lands. Federal lands in the United States provide the majority of habitat for greater sage-grouse in MZs II (Wyoming Basin), III (Southern Great Basin), IV (Snake River Plain), and V (Northern Great Basin), whereas private lands provide the majority of habitat in MZs I (Northern Great Plains), VI (Columbia Basin), and VII (Colorado Plateau; Table 10.1). Gunnison sage-grouse only inhabit portions of MZ VII, where land ownership for this species is nearly evenly split between public (federal [47.2%] and state ([2.1%]) and private (50.7%; Table 10.1).
State and provincial wildlife management agencies have the primary role of managing sage-grouse populations, inclusive of providing sustainable hunting opportunities. To implement and prioritize coordinated conservation efforts, numerous conservation plans have been drafted by federal, state, provincial, and local working groups that detail specific threats and necessary conservation practices, which typically address disturbance (sagebrush conversion, energy and mining development, fire), habitat (conifer encroachment, invasive plants, grazing), and predation. State and local plans generally delineated priority areas based on breeding habitat surrounding sage-grouse leks, and these delineations were used to inform national Priority Areas for Conservation (PACs) and Priority Habitat Management Areas in the U.S (PHMAs; USFWS 2013, 2015). Examples of state priority areas and associated conservation policies include Core Areas in Oregon, Montana, and Wyoming, state Management Zones and Conservation Areas in Idaho, and Sage-Grouse Management Areas in Utah. Because federal and state designations of priority habitat were both informed by state assessments, their shape and extent tend to be similar (USFWS 2013), and they generally aim to maintain stable and reverse declining sage-grouse populations by minimizing threats and restoring degraded habitat. In Wyoming, the rate of energy development was restricted in core areas (Gamo and Beck 2017), and sage-grouse populations tend to perform better within core areas (Spence et al. 2017; Dinkins and Beck 2019; Heinrichs et al. 2019). However, priority areas tend to conserve breeding habitat over winter habitat (Smith et al. 2016; Dinkins et al. 2017), some plans for priority areas do not explicitly consider livestock management (Dinkins et al. 2016b), and high levels of human development adjacent to priority areas may still negatively affect sage-grouse populations within these priority areas (Spence et al. 2017; Heinrichs et al. 2019). Development activities within core areas, as allowed by specific policies, may negatively affect sage-grouse populations if adjacent to large leks or high-quality sage-grouse habitats.
With the 2015 sage-grouse listing decision by USFWS, the BLM and USFS committed to monitoring and reviewing grazing authorizations (permits and leases) in sagebrush focal areas (SFAs; USFWS 2015), lands deemed highest priority for conserving sage-grouse within PHMAs (USFWS 2014). Monitoring and evaluation based on Land Health Standards would be prioritized in these areas to determine whether habitat objectives for greater sage-grouse are met, whether modifications and management are needed, and to ensure compliance (USFWS 2015). Additionally, a conceptual framework was recently formalized for understanding the ability of sagebrush ecosystems to recover following stressors and disturbance such as drought and fire (resilience) and retain their original state such as by resisting invasion by annual grasses (Chambers et al. 2014). Soil temperature and moisture can largely determine resilience and resistance of sagebrush ecosystems (for example, faster recovery under cool and moist conditions), and resilience and resistance can be predicted across the landscape using maps of soils data (Maestas et al. 2016). Federal agencies currently use resilience and resistance concepts to evaluate threats and risk to sagebrush ecosystems and prioritize resources for sage-grouse conservation (Chambers et al. 2017; Crist et al. 2019). At finer scales, federal agencies are committed to using the Habitat Assessment Framework (HAF; Stiver et al. 2015) to determine management objectives for sage-grouse habitat on public lands (USFWS 2015). The BLM, USGS, and WAFWA are currently working to refine habitat objectives at multiple scales using rangewide datasets of sage-grouse lek locations, movement, and remote sensing products (C. L. Aldridge, U.S. Geological Survey, written communication, 2021).
Hunting regulations for sage-grouse were instituted among states starting around the turn of the twentieth century. Restrictions to hunting regulations, such as season length, bag limits, and hunting season closures, have generally been implemented in response to declines in sage-grouse numbers (Wambolt et al. 2002; Dinkins et al. 2021a). Although hunting can negatively impact sage-grouse populations (Connelly et al. 2003a, b; Blomberg 2015; Caudill et al. 2017), it is generally unclear that current harvest limits are detrimental (Sedinger et al. 2010; Taylor et al. 2012), and states and provinces have adjusted harvest regulations to reduce hunting exposure to declining populations or in response to management challenges such as extreme fire events (Dinkins et al. 2021a).
8 Research/Management Needs
As iconic species dependent on sagebrush-dominated rangelands across western North America, we have summarized the myriad conservation challenges facing greater and Gunnison sage-grouse, yet gaps remain in both our knowledge base and potential solutions. Livestock production is one of the oldest and most widespread land use types overlapping sage-grouse range; however, experiments directly evaluating effects of livestock on sage-grouse demography and population trends are limited (Beck and Mitchell 2000) but have been performed in Montana (Smith et al. 2018a) and are currently being conducted in Idaho (Conway et al. 2021). Despite uncertainty over the importance of grass height for sage-grouse nest success, herbaceous cover, particularly forbs and associated arthropods, may still be important for brood-rearing sage-grouse, yet little research exists for livestock management in brood-rearing habitat (but see Street 2020). Given variability in environmental conditions over space and time (such as precipitation; Blomberg et al. 2012; Guttery et al. 2013), replicating grazing experiments and observational studies over long temporal periods (> 2 years) is needed. Further, greater sage-grouse respond and use habitat at multiple scales, including for nest sites, brood-rearing, and wintering habitat, so examining how rangeland management affects these different components at relevant scales is warranted (for example, see Smith et al. 2023). Broad-scale studies are needed to anticipate how policies and land use alternatives may affect sage-grouse populations. Records of livestock on public lands may indicate levels of use across these vast landscapes (Veblen et al. 2014; Monroe et al. 2017), but these data are often coarse (reported annually at the scale of the allotment). An evaluation of how these records relate to forage consumption, and therefore to the structure and composition of vegetation, is warranted. Federal agencies have committed to implementing management standards for sage-grouse in priority areas (USFWS 2015), but consistent, long-term monitoring will be critical to detect deviations from desired management outcomes (Veblen et al. 2014). Effects of increasing feral equid populations on habitat alteration and sage-grouse populations also merit further study.
Studying each of the ecosystem threats listed above in the context of other threats will be necessary, given the potential for interactions among multiple stressors. Sustainable management of livestock is compatible with sage-grouse (Beck and Mitchell 2000; Smith et al. 2018a), but levels of forage production that define sustainable grazing could change under future climate conditions (Reeves et al. 2014). While resilience and resistance concepts can be used to anticipate effects of climate change (Bradford et al. 2019; Crist et al. 2019), current consideration for this stressor by federal agencies may be limited (Brice et al. 2020). Given the effects of increasing wildfire and exotic annual grass spread on sage-grouse populations (Coates et al. 2016b; Smith et al. 2022), additional research and management directed at controlling exotic annual grasses and restoring degraded habitat will benefit sage-grouse. Finally, greater study of effects to small and isolated populations including constrained movements, predation, genetic issues, and emerging infectious diseases could identify interactions with other environmental and anthropogenic threats, potentially revealing additional mechanisms driving sage-grouse population trends.
References
Abatzoglou JT, Kolden CA (2011) Climate change in western US deserts: potential for increased wildfire and invasive annual grasses. Rangeland Ecol Manag 64:471–478. https://doi.org/10.20111/REM-D-09-00151.1
Ahlers LRH, Goodman AG (2018) The immune responses of the animal hosts of West Nile virus: a comparison of insects, birds, and mammals. Front Cell Infect Mi 8:96. https://doi.org/10.3389/fcimb.2018.00096
Aldrich JW (1963) Geographic orientation of American Tetraonidae. J Wildl Manage 27:528–545. https://doi.org/10.2307/3798463
Aldrich JW, Duvall AJ (1955) Distribution of American gallinaceous game birds. United States Fish and Wildlife Service, Circular 34, Washington, DC
Aldridge CA, Boyce MS (2007) Linking occurrence and fitness to persistence: habitat‐based approach for endangered greater sage‐grouse. Ecol Appl 17:508–526. https://www-jstor-org.libproxy.uwyo.edu/stable/40061874
Aldridge CL, Brigham RM (2002) Sage-grouse nesting and brood habitat use in Southern Canada. J Wildl Manage 66:433–444. https://doi.org/10.2307/3803176
Aldridge CL, Saher DJ, Childers TM, Stahlnecker KE, Bowen ZH (2012) Crucial nesting habitat for Gunnison Sage-Grouse: a spatially explicit hierarchical approach. J Wildl Manage 76:391–406. https://doi.org/10.1002/jwmg.268
Allred BW, Smith WK, Twidwell D, Haggerty JH, Running SW, Naugle DE, Fuhlendorf SD (2015) Ecosystem services lost to oil and gas in North America. Science 348:401–402. https://doi.org/10.1126/science.aaa4785
Arredondo JT, Jones TA, Johnson DA (1998) Seedling growth of intermountain perennial and weedy annual grasses. J Range Manage 51:584–589. https://doi.org/10.2307/4003380
Asay KH, Chatterton NJ, Jensen KB, Jones TA, Waldron BL, Horton WH (2003) Breeding improved grasses for semiarid rangelands. Arid Land Res Manag 17:469–478. https://doi.org/10.1080/713936115
Atamian MT, Sedinger JS, Heaton JS, Blomberg EJ (2010) Landscape-level assessment of brood rearing habitat for greater sage-grouse in Nevada. J Wildl Manage 74:1533–1543. https://doi.org/10.2193/2009-226
Back GN, Barrington MR, McAdoo JK (1987) Sage grouse use of snow burrows in northeastern Nevada. Wilson Bull 99:488–490. https://www.jstor.org/stable/4162435
Balch JK, Bradley BA, D’Antonio CM, Gómez-Dans J (2013) Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Glob Change Biol 19:173–183. https://doi.org/10.1111/gcb.12046
Balderson KL (2017) Habitat selection and nesting ecology of translocated greater sage-grouse. University of Regina, Regina, Saskatchewan, Thesis
Baruch-Mordo S, Evans JS, Severson JP, Naugle DE, Maestas JD, Kiesecker JM, Falkowski MJ, Hagen CA, Reese KP (2013) Saving sage-grouse from the trees: a proactive solution to reducing a key threat to a candidate species. Biol Conserv 167:233–241. https://doi.org/10.1016/j.biocon.2013.08.017
Baumgardt JA, Reese KP, Connelly NW, Garton EO (2017) Visibility bias for sage-grouse lek counts. Wildl Soc Bull 41:461–470. https://doi.org/10.1002/wsb.800
Beck JL, Mitchell DL (2000) Influences of livestock grazing on sage grouse habitat. Wildlife Soc B 28:993–1002. https://www.jstor.org/stable/3783858
Beck JL, Mitchell DL, Maxfield BD (2003) Changes in the distribution and status of sage-grouse in Utah. West N Am Nat 63:203–214. https://www.jstor.org/stable/41717283
Beck JL, Reese KP, Connelly JW, Lucia MB (2006) Movements and survival of juvenile greater sage-grouse in southeastern Idaho. Wildlife Soc B 34:1070–1078. https://doi.org/10.2193/0091-7648(2006)34[1070:MASOJG]2.0.CO;2
Beck JL, Connelly JW, Wambolt CL (2012) Consequences of treating Wyoming big sagebrush to enhance wildlife habitats. Rangeland Ecol Manag 65:444–455. https://doi.org/10.2111/REM-D-10-00123.1
Beever EA, Aldridge CL (2011) Influences of free-roaming equids on sagebrush ecosystems, with a focus on greater sage-grouse. In: Knick ST, Connelly JW (eds) Greater sage-grouse: ecology and conservation of a landscape species and its habitats. Studies in Avian Biology 38, University of California Press, Berkeley, California, pp 273–290. https://doi.org/10.1525/California/9780520267114.003.0015
Beever EA, Huntsinger L, Petersen SL (2018) Conservation challenges emerging from free-roaming horse management: a vexing social-ecological mismatch. Biol Conserv 226:321–328. https://doi.org/10.1016/j.biocon.2018.07.015
Bergquist E, Evangelista P, Stohlgren TJ, Alley N (2007) Invasive species and coal bed methane development in the Powder River Basin, Wyoming. Environ Monit Assess 128:381–394. https://doi.org/10.1007/s10661-006-9321-7
Bendire CE (1892) Life histories of North American birds. United States National Museum, Special Bulletin 1, Washington, DC
Blomberg EJ (2015) The influence of harvest timing on greater sage-grouse survival: a cautionary perspective. J Wildl Manage 79:695–703. https://doi.org/10.1002/jwmg.887
Blomberg EJ, Hagen CA (2020) How many leks does it take? Minimum samples sizes for measuring local-scale conservation outcomes in greater sage-grouse. Avian Conserv Ecol 15:9. https://doi.org/10.5751/ACE-01517-150109
Blomberg EJ, Sedinger JS, Atamian MT, Nonne DV (2012) Characteristics of climate and landscape disturbance influence the dynamics of greater sage-grouse populations. Ecosphere 3:55. https://doi.org/10.1890/ES11-00304.1
Blomberg EJ, Poulson SR, Sedinger JS, Gibson D (2013a) Prefledging diet is correlated with individual growth in greater sage-grouse (Centrocercus urophasianus). Auk 130:715–724. https://doi.org/10.1525/auk.2013.12188
Blomberg EJ, Sedinger JS, Nonne DV, Atamian MT (2013b) Annual male lek attendance influences count-based population indices of greater sage-grouse. J Wildl Manage 77:1583–1592. https://doi.org/10.1002/jwmg.615
Blomberg EJ, Sedinger JS, Nonne DV, Atamian MT (2013c) Seasonal reproductive costs contribute to reduced survival of female greater sage-grouse. J Avian Biol 44:149–158. https://doi.org/10.1111/j.1600-048X.2012.00013.x
Blomberg EJ, Sedinger JS, Gibson D, Coates PS, Casazza ML (2014) Carryover effects and climatic conditions influence postfledging survival of greater sage-grouse. Ecol Evol 4:4488–4499. https://doi.org/10.1002/ece3.1139
Boarman WI (2003) Managing a subsidized predator population: reducing common raven predation on desert tortoises. Environ Manage 32:205–217. https://doi.org/10.1007/s00267-003-2982-x
Boyte SP, Wylie BK, Major DJ (2016) Cheatgrass percent cover change: comparing recent estimates to climate change—driven predictions in the northern Great Basin. Rangeland Ecol Manag 69:265–279. https://doi.org/10.1016/j.rama.2016.03.002
Bradford JB, Schlaepfer DR, Lauenroth WK, Palmquist KA, Chambers JC, Maestas JD, Campbell SB (2019) Climate-driven shifts in soil temperature and moisture regimes suggest opportunities to enhance assessments of dryland resilience and resistance. Front Ecol Evol 7:358. https://doi.org/10.3389/fevo.2019.00358
Bradley BA, Curtis CA, Fusco EJ, Abatzoglou JT, Balch JK, Dadashi S, Tuanmu M-N (2018) Cheatgrass (Bromus tectorum) distribution in the intermountain Western United States and its relationship to fire frequency, seasonality, and ignitions. Biol Invasions 20:1493–1506. https://doi.org/10.1007/s10530-017-1641-8
Braun CE, Oyler-McCance SJ, Nehring JA, Commons ML, Young JR, Potter KM (2014) The historical distribution of Gunnison sage-grouse in Colorado. Wilson J Ornithol 126:207–217. https://doi.org/10.1676/13-184.1
Braun CE, Schroeder MA (2015) Age and sex identification from wings of sage-grouse. Wildl Soc Bull 39:182–187. https://doi.org/10.1002/wsb.517
Braun CE, Williams SO III (2015) History of sage-grouse (Centrocercus spp.) in New Mexico. Southwest Nat 60:207–213. https://doi.org/10.1894/MCG-141
Brice EM, Miller BA, Zhang H, Goldstein K, Zimmer SN, Grosklos GJ, Belmon P, Flint CG, Givens JE, Adler PB, Brunson MW, Smith JW (2020) Impacts of climate change on multiple use management of Bureau of Land Management land in the Intermountain West, USA. Ecosphere 11:e03286. https://doi.org/10.1002/ecs2.3286
Brown DG, Johnson KM, Loveland TR, Theobald DM (2005) Rural land-use trends in the conterminous United States, 1950–2000. Ecol Appl 15:1851–1863. https://doi.org/10.1890/03-5220
Brust RA (1991) Environmental regulation of autogeny in Culex tarsalis (Diptera: Culicidae) from Manitoba, Canada. J Med Entomol 28:847–853. https://doi.org/10.1093/jmedent/28.6.847
Carlisle JD, Keinath DA, Albeke SE, Chalfoun AD (2018) Identifying holes in the greater sage-grouse conservation umbrella. J Wildl Manage 82:948–957. https://doi.org/10.1002/jwmg.21460
Carpenter J, Aldridge C, Boyce MS (2010) Sage-grouse habitat selection during winter in Alberta. J Wildl Manage 74:1806–1814. https://doi.org/10.2193/2009-368
Caudill D, Guttery MR, Terhune TM II, Martin JA, Caudill G, Dahlgren DK, Messmer TA (2017) Individual heterogeneity and effects of harvest on greater sage-grouse populations. J Wildl Manage 81:754–765. https://doi.org/10.1002/jwmg.21241
Chambers JC, Bradley BA, Brown CS, D’Antonio C, Germino MJ, Grace JB, Hardegree SP, Miller RF, Pyke DA (2014) Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America. Ecosystems 17:360–375. https://doi.org/10.1007/s10021-013-9725-5
Chambers JC, Maestas JD, Pyke DA, Boyd CS, Pellant M, Wuenschel A (2017) Using resilience and resistance concepts to manage persistent threats to sagebrush ecosystems and greater sage-grouse. Rangeland Ecol Manag 70:149–164. https://doi.org/10.1016/j.rama.2016.08.005
Christian JM, Wilson SD (1999) Long-term ecosystem impacts of an introduced grass in the northern Great Plains. Ecology 80:2397–2407. https://doi.org/10.1890/0012-9658(1999)080[2397:LTEIOA]2.0.CO;2
Christiansen TJ, Tate CM (2011) Parasites and diseases of greater sage-grouse. In: Knick ST, Connelly JW (eds) Greater sage-grouse: ecology and conservation of a landscape species and its habitats. Studies in avian biology 38, University of California Press, Berkeley, California, pp 113–126. https://doi.org/10.1525/California/9780520267114.003.0015
Coates PS, Casazza ML, Blomberg EJ, Gardner SC, Espinosa SP, Yee JL, Wiechman L, Halstead BJ (2013) Evaluating greater sage-grouse seasonal space use relative to leks: implications for surface use designations in sagebrush ecosystems. J Wildl Manage 77:1598–1609. https://doi.org/10.1002/jwmg.618
Coates PS, Brussee BE, Howe KB, Gustafson KB, Casazza ML, Delehanty DJ (2016a) Landscape characteristics and livestock presence influence common ravens: relevance to greater sage-grouse conservation. Ecosphere 7:e01203. https://doi.org/10.1002/ecs2.1203
Coates PS, Ricca MA, Prochazka BG, Brooks ML, Doherty KE, Kroger T, Blomberg EJ, Hagen CA, Casazza ML (2016b) Wildfire, climate, and invasive grass interactions negatively impact an indicator species by reshaping sagebrush ecosystems. Proc Natl A Sci-Biol 113:12745–12750. https://doi.org/10.1073/pnas.1606898113
Coates PS, Prochazka BG, Ricca MA, Gustafson KB, Ziegler P, Casazza ML (2017) Pinyon and juniper encroachment into sagebrush ecosystems impacts distribution and survival of greater sage-grouse. Rangeland Ecol Manag 70:25–38. https://doi.org/10.1016/j.rama.2016.09.001
Coates PS, Prochazka BG, Ricca MA, Halstead BJ, Casazza ML, Blomberg EJ, Brussee BE, Wiechman L, Tebbenkamp J, Gardner SC, Reese KP (2018) The relative importance of intrinsic and extrinsic drivers to population growth vary among local populations of greater sage-grouse: an integrated population modeling approach. Auk 135:240–261. https://doi.org/10.1642/AUK-17-137.1
Coates PS, Wann GT, Gillette GL, Ricca MA, Prochazka BG, Severson JP, Andrle KM, Espinosa SP, Casazza ML, Delehanty DJ (2019) Estimating sightability of greater sage-grouse at leks using an aerial infrared system and N-mixture models. Wildl Biol 1:1–11. https://doi.org/10.2981/wlb.00552
Coates PS, O’Neil ST, MuÑoz DA, Dwight IA, Tull JC (2021a) Sage-grouse population dynamics are adversely affected by overabundant feral horses. J Wildl Manage 85:1132–1149. https://doi.org/10.1002/jwmg.22089
Coates PS, Prochazka BG, O’Donnell MS, Aldridge CL, Edmunds DR, Monroe AP, Ricca MA, Wann GT, Hanser SE, Wiechman LA, Chenaille MP (2021b) Range-wide greater sage-grouse hierarchical monitoring framework—implications for defining population boundaries, trend estimation, and a targeted annual warning system: U.S. Geological Survey Open-File Report 2020–1154, 243 p. https://doi.org/10.3133/ofr20201154
Cole RA, Friend M (1999) Miscellaneous parasitic diseases. In: Friend M, Franson JC (eds) Field manual of wildlife diseases: general field procedures and diseases of birds. U.S. Geological Survey Biological Resources Division Information and Technical Report 1999–001, pp 249–258. https://pubs.er.usgs.gov/publication/itr19990001
Connelly JW, Browers HW, Gates RJ (1988) Seasonal movements of sage grouse in southeastern Idaho. J Wildl Manage 52:116–122. https://doi.org/10.2307/3801070
Connelly JW, Hagen CA, Schroeder MA (2011a) Characteristics and dynamics of greater sage-grouse populations. In: Knick ST, Connelly JW (eds) Greater sage-grouse: ecology and conservation of a landscape species and its habitats. Studies in avian biology 38, University of California Press, Berkeley, California, pp 53–67. https://doi.org/10.1525/California/9780520267114.003.0004
Connelly JW, Knick ST, Schroeder MA, Stiver SJ (2004) Conservation assessment of greater sage-grouse and sagebrush habitats. Unpublished report, Western Association of Fish and Wildlife Agencies, Cheyenne, Wyoming
Connelly JW, Reese KP, Garton EO, Commons-Kemner ML (2003a) Response of greater sage-grouse Centrocercus urophasianus populations to different levels of exploitation in Idaho, USA. Widl Biol 9:335–340. https://doi.org/10.2981/wlb.2003.022
Connelly JW, Reese KP, Schroeder MA (2003b) Monitoring of greater sage-grouse habitats and populations. Station Bulletin 80. University of Idaho, Moscow, USA. https://doi.org/10.5962/BHL.TITLE.153828
Connelly JW, Rinkes ET, Braun CE (2011b) Characteristics of greater sage-grouse habitats: a landscape species at micro- and macro scales. In: Knick ST, Connelly JW (eds) Greater sage‐grouse: ecology and conservation of a landscape species and its habitats. Studies in avian biology 38, University of California Press, Berkeley, pp 69–83. https://doi.org/10.1525/California/9780520267114.003.0006
Connelly JW, Schroeder MA, Sands AR, Braun CE (2000) Guidelines to manage sage grouse populations and their habitats. Wildl Soc Bull 28:967–985. https://www.jstor.org/stable/3783856
Conover MR, Roberts AJ (2017) Predators, predator removal, and sage-grouse: a review. J Wildl Manage 81:7–15. https://doi.org/10.1002/jwmg.21168
Conway CJ, Tisdale CA, Launchbaugh K, Musil D, Makela P, Roberts S (2021) The grouse & grazing project: effects of cattle grazing on sage-grouse demographic traits—2021 annual report. College of Natural Resources, University of Idaho. https://idahogrousegrazing.files.wordpress.com/2021/12/2021_annualreport.pdf
Copeland SM, Davies KW, Boyd CS, Bates JD (2021) Recovery of the herbaceous component of sagebrush steppe unimpeded by 75 years of moderate cattle grazing. Ecosphere 12:e03445. https://doi.org/10.1002/ecs2.3445
Crawford JA, Olson RA, West NE, Mosley JC, Schroeder MA, Whitson TD, Miller RF, Gregg MA, Boyd CS (2004) Ecology and management of sage-grouse and sage-grouse habitat. J Range Manage 57:2–19. https://doi.org/10.2111/1551-5028(2004)057[0002:EAMOSA]2.0.CO;2
Creutzburg MK, Halofsky JE, Halofsky JS, Christopher TA (2015) Climate change and land management in the rangelands of central Oregon. Environ Manage 55:43–55. https://doi.org/10.1007/s00267-014-0362-3
Crist MR, Chambers JC, Phillips SL, Prentice KL, Wiechman LA (eds) (2019) Science framework for conservation and restoration of the sagebrush biome: linking the department of the interior’s integrated rangeland fire management strategy to long-term strategic conservation actions. Part 2. Management applications. Gen. Tech. Rep. RMRS-GTR-389. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, Colorado, 237 pp. https://doi.org/10.2737/RMRS-GTR-389
D’Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu Rev Ecol Syst 23:63–87. https://www.jstor.org/stable/2097282
Dahlgren DK, Chi R, Messmer TA (2006) Greater sage‐grouse response to sagebrush management in Utah. Wildl Soc Bull 34:975–985. https://www-jstor-org.libproxy.uwyo.edu/stable/4134306
Dahlgren DK, Larsen RT, Danvir R, Wilson G, Thacker ET, Black TA, Naugle DE, Connelly JW, Messmer TA (2015) Greater sage-grouse and range management: insights from a 25-year case study in Utah and Wyoming. Rangeland Ecol Manag 68:375–382. https://doi.org/10.1016/j.rama.2015.07.003
Dahlgren DK, Guttery MR, Messmer TA, Caudill D, Elmore RD, Chi R, Koons DN (2016) Evaluating vital rate contributions to greater sage-grouse population dynamics to inform conservation. Ecosphere 7:e01249. https://doi.org/10.1002/ecs2.1249
Daubenmire R (1970) Steppe vegetation of Washington. Washington Agricultural Experiment Station Technical Bulletin 62, 131 p
Davies KW (2011) Plant community diversity and native plant abundance decline with increasing abundance of an exotic annual grass. Oecologia 167:481–491. https://doi.org/10.1007/s00442-011-1992-2
Davies KW, Bates JD (2014) Attempting to restore herbaceous understories in Wyoming big sagebrush communities with mowing and seeding. Restor Ecol 22:608–615. https://doi.org/10.1111/rec.12110
Davies KW, Bates JD (2020) Re-introducing fire in sagebrush-steppe experiencing decreased fire frequency: does burning promote spatial and temporal heterogeneity? Int J Wildland Fire 29:686–695. https://doi.org/10.1071/WF20018
Davies KW, Svejcar TJ, Bates JD (2009) Interaction of historical and nonhistorical disturbances maintains native plant communities. Ecol Appl 19:1536–1545. https://doi.org/10.1890/09-0111.1
Davies KW, Nafus AM, Sheley RL (2010) Non-native competitive perennial grass impedes the spread of an invasive annual grass. Biol Invasions 12:3187–3194. https://doi.org/10.1007/s10530-010-9710-2
Davies KW, Boyd CS, Beck JL, Bates JD, Svejcar TJ, Gregg MA (2011) Saving the sagebrush sea: an ecosystem conservation plan for big sagebrush plant communities. Biol Conserv 144:2573–2584. https://doi.org/10.1016/j.biocon.2011.07.016
Davies KW, Bates JD, Nafus AM (2012) Mowing Wyoming big sagebrush communities with degraded herbaceous understories: has a threshold been crossed? Rangeland Ecol Manag 65:498–505. https://doi.org/10.2111/REM-D-12-00026.1
Davis AJ, Hooten MB, Phillips ML, Doherty PF Jr (2014a) An integrated modeling approach to estimating Gunnison sage-grouse population dynamics: combining index and demographic data. Ecol Evol 4:4247–4257. https://doi.org/10.1002/ece3.1290
Davis DM, Reese KP, Gardner SC (2014b) Demography, reproductive ecology, and variation in survival of greater sage-grouse in northeastern California. J Wildl Manage 78:1343–1355. https://doi.org/10.1002/jwmg.797
Davies KW, Boyd CS, Bates JD, Hulet A (2015a) Winter grazing can reduce wildfire size, intensity, and behaviour in a shrub-grassland. Int J Wildland Fire 25:191–199. https://doi.org/10.1071/WF15055
Davies KW, Boyd CS, Johnson DD, Nafus AM, Madsen MD (2015b) Success of seeding native compared to introduced perennial vegetation for revegetating medusahead-invaded sagebrush rangeland. Rangeland Ecol Manag 68:224–230. https://doi.org/10.1016/j.rama.2015.03.004
Davies KW, Bates JD, Boyd CS (2016a) Effects of intermediate-term grazing rest on sagebrush communities with depleted understories: evidence of a threshold. Rangeland Ecol Manag 69:173–178. https://doi.org/10.1016/j.rama.2016.01.002
Davies KW, Bates JD, Boyd CS, Svejcar TJ (2016b) Prefire grazing by cattle increases postfire resistance to exotic annual grass (Bromus tectorum) invasion and dominance for decades. Ecol Evol 6:3356–3366. https://doi.org/10.1002/ece3.2127
Davies KW, Boyd CS, Bates JD (2018) Eighty years of grazing by cattle modifies sagebrush and bunchgrass structure. Rangeland Ecol Manag 71:275–280. https://doi.org/10.1016/j.rama.2018.01.002
Davies KW, Bates JD, Boyd CS (2019) Postwildfire seeding to restore native vegetation and limit exotic annuals: an evaluation in juniper-dominated sagebrush steppe. Restor Ecol 27:120–127. https://doi.org/10.1111/rec.12848
Dinkins JB, Beck JL (2019) Comparison of conservation policy benefits for an umbrella and related sagebrush-obligate species. Hum Wildl Interact 13:447–458. https://doi.org/10.26077/4ypp-vj89
Dinkins JB, Conover MR, Kirol CP, Beck JL (2012) Greater sage-grouse (Centrocercus urophasianus) select nest-sites and brood-sites away from avian predators. Auk 129:600–610. https://doi.org/10.1525/auk.2012.12009
Dinkins JB, Conover MR, Kirol CP, Beck JL, Frey SN (2014). Greater sage-grouse (Centrocercus urophasianus) select habitat based on avian predators, landscape composition, and anthropogenic features. Condor: Ornithol Appl 116:629–642. https://doi.org/10.1650/CONDOR-13-163.1
Dinkins JB, Conover MR, Kirol CP, Beck JL, Frey SN (2016a) Effects of common raven and coyote removal and temporal variation in climate on greater sage-grouse nesting success. Biol Conserv 202:50–58. https://doi.org/10.1016/j.biocon.2016.08.011
Dinkins JB, Lawson KJ, Smith KT, Beck JL, Kirol CP, Pratt AC, Conover MR, Blomquist FC (2017) Quantifying overlap and fitness consequences of migration strategy with seasonal habitat use and a conservation policy. Ecosphere 8:e01991. https://doi.org/10.1002/ecs2.1991
Dinkins JB, Duchardt CJ, Hennig JD, Beck JL (2021a) Changes in hunting season regulations (1870s–2019) reduce harvest exposure on greater and Gunnison sage-grouse. PLoS ONE 16:e0253635. https://doi.org/10.1371/journal.pone.0253635
Dinkins JB, Perry LR, Beck JL, Taylor JD (2021b) Increased abundance of the common raven within the ranges of greater and Gunnison sage-grouse: influence of anthropogenic subsidies and fire. Hum Wildl Interact 15:270–288. https://doi.org/10.26077/mv59-jy24
Dinkins JB, Smith KT, Beck JL, Kirol CP, Pratt AC, Conover MR (2016b) Microhabitat conditions in Wyoming's Sage-Grouse Core Areas: effects on nest site selection and success. PLoS ONE 11(3):e0150798. https://doi.org/10.1371/journal.pone.0150798
Doherty MK (2007) Mosquito populations in the Powder River Basin, Wyoming: a comparison of natural, agricultural and effluent coal-bed natural gas aquatic habitats. Montana State University, Bozeman, Montana, Thesis
Doherty KE, Naugle DE, Evans JS (2010) A currency for offsetting energy development impacts: horse-trading sage-grouse on the open market. PLoS ONE 5:e10339. https://doi.org/10.1371/journal.pone.0010339
Doherty KE, Naugle DE, Tack JD, Walker BL, Graham JM, Beck JL (2014) Linking conservation actions to demography: grass height explains variation in greater sage-grouse nest survival. Wildl Biol 20:320–325. https://doi.org/10.2981/wlb.00004
Doherty KE, Evans JS, Coates PS, Juliusson LM, Fedy BC (2016) Importance of regional variation in conservation planning: a rangewide example of the greater sage-grouse. Ecosphere 7:e01462. https://doi.org/10.1002/ecs2.1462
Doherty KE, Hennig JD, Dinkins JB, Griffin KA, Cook AA, Maestas JD, Naugle DE, Beck JL (2018) Understanding biological effectiveness before scaling up rangewide restoration investments for Gunnison sage-grouse. Ecosphere 9:e02144. https://doi.org/10.1002/ecs2.2144
Donnelly JP, Naugle DE, Hagen CA, Maestas JD (2016) Public lands and private waters: scarce mesic resources structure land tenure and sage-grouse distributions. Ecosphere 7:e01208. https://doi.org/10.1002/ecs2.1208
Donnelly JP, Allred BW, Perret D, Silverman NL, Tack JD, Dreitz VJ, Maestas JD, Naugle DE (2018) Seasonal drought in North America’s sagebrush biome structures dynamic mesic resources for sage-grouse. Ecol Evol 24:12492–12505. https://doi.org/10.1002/ece3.4614
Drut MS, Crawford JA, Gregg MA (1994) Brood habitat use by sage grouse in Oregon. Great Basin Nat 54:170–176. https://www.jstor.org/stable/41712827
Dumroese RK, Luna T, Richardson BA, Kilkenny FF, Runyon JB (2015) Conserving and restoring habitat for greater sage-grouse and other sagebrush-obligate wildlife: the crucial link of forbs and sagebrush diversity. Native Plants J 16:277–299. https://doi.org/10.3368/npj.16.3.276
Dusek RJ, Hagen CA, Franson JC, Budeau DA, Hofmeister EK (2014) Utilizing hunter harvest effort to survey for wildlife disease: a case study of West Nile virus in greater sage-grouse. Wildl Soc Bull 38:721–727. https://doi.org/10.1002/wsb.472
Emmons SR, Braun CE (1984) Lek attendance of male sage grouse. J Wildl Manage 48:1023–1028. https://doi.org/10.2307/3801461
Environment Canada (2014) Amended recovery strategy for the greater sage-grouse (Centrocercus urophasianus urophasianus) in Canada. Species at Risk Act Recovery Strategy Series, Environment Canada, Ottawa, Ontario, p 53
Evans CC (1986) The relationship of cattle grazing to sage grouse use of meadow habitat on the Sheldon National Wildlife Refuge. University of Nevada, Reno, Thesis
Fargione JE, Bassett S, Boucher T, Bridgham SD, Conant RT, Cook-Patton SC, Ellis PW, Falcucci A, Fourqurean JW, Gopalakrishna T, Gu H, Henderson B, Hurteau MD, Kroeger MD, Kroeger T, Lark TJ, Leavitt SM, Lomax G, McDonald RI, Megonicgal JP, Miteva DA, Richardson CJ, Sanderman J, Shoch D, Spawn SA, Veldman JW, Williams CA, Woodbury PB, Zganjar C, Baranski M, Elias P, Houghton RA, Landis E, McGlynn E, Schlesinger WH, Siikamaki JV, Sutton-Grier AE, Griscom BW (2018) Natural climate solutions for the United States. Sci Adv 4:eaat1869. https://doi.org/10.1126/sciadv.aat1869
Fedy BC, Aldridge CL, Doherty KE, O’Donnell M, Beck JL, Bedrosian B, Holloran MJ, Johnson GD, Kaczor NW, Kirol CP, Mandich CA, Marshall D, McKee G, Olson C, Swanson CC, Walker BL (2012) Interseasonal movements of greater sage-grouse, migratory behavior, and an assessment of the core regions concept in Wyoming. J Wildl Manage 76:1062–1071. https://doi.org/10.1002/jwmg.337
Fedy BC, Doherty KE, Aldridge CL, O’Donnell M, Beck JL, Bedrosian B, Gummer D, Holloran MJ, Johnson GD, Kaczor NW, Kirol CP, Mandich CA, Marshall D, Mckee G, Olson C, Pratt AC, Swanson CC, Walker BL (2014) Habitat prioritization across large landscapes, multiple seasons, and novel areas: an example using greater sage-grouse in Wyoming. Wildl Monogr 190:1–39. https://doi.org/10.1002/wmon.1014
Finch D, Schofield H, Floate KD, Kubasiewicz LM, Mathews F (2020) Implications of endectocide residues on the survival of aphodiine dung beetles: a meta-analysis. Environ Toxicol Chem 39:863–872. https://doi.org/10.1002/etc.4671
Fischer RA, Reese KP, Connelly JW (1996) Influence of vegetal moisture content and nest fate on timing of female sage grouse migration. Condor 98:868–872. https://doi.org/10.2307/1369875
Flader SL, Callicott JB, Leopold A (1992) The river of the mother of god: and other essays by Aldo Leopold. University of Wisconsin Press, Madison, Wisconsin
Fremgen AL, Hansen CP, Rumble MA, Gamo RS, Millspaugh JJ (2016) Male greater sage-grouse detectability on leks. J Wildl Manage 80:266–274. https://doi.org/10.1002/jwmg.1001
Fremgen MR, Gibson D, Ehrlich RL, Krakauer AH, Forbey JS, Blomberg EJ, Sedinger JS, Patricelli GL (2017) Necklace-style radio-transmitters are associated with changes in display vocalizations of male greater sage-grouse. Wildl Biol 2017:wlb.00236. https://doi.org/10.2981/wlb.00236
Fremgen AL, Hansen CP, Rumble MA, Gamo RS, Millspaugh JJ (2019) Weather conditions and date influence male sage grouse attendance rates at leks. Ibis 161:35–49. https://doi.org/10.1111/ibi.12598
Gamo RS, Beck JL (2017) Effectiveness of Wyoming’s sage-grouse core areas: influences on energy development and male lek attendance. Environ Manage 59:189–203. https://doi.org/10.1007/s00267-016-0789-9
Gamo RS, Carlisle JD, Beck JL, Bernard JAC, Herget ME (2013) Greater sage-grouse in Wyoming: an umbrella species for sagebrush-dependent wildlife. Wildlife Professional 7:56–59
Garton EO, Connelly JW, Horne JS, Hagen CA, Moser A, Schroeder MA (2011) Greater sage-grouse population dynamics and probability of persistence. In: Knick ST, Connelly JW (eds) Greater sage‐grouse: ecology and conservation of a landscape species and its habitats. Studies in avian biology 38, University of California Press, Berkeley, pp 293–381. https://doi.org/10.1525/California/9780520267114.003.0016
Garton EO, Wells AG, Baumgardt JA, Connelly JW (2015) Greater sage-grouse population dynamics and probability of persistence. Final report to Pew Charitable Trusts. Available at https://www.pewtrusts.org/~/media/assets/2015/04/garton-et-al-2015-greater-sagegrouse-population-dynamics-and-persistence-31815.pdf
Gelbard JL, Belnap J (2003) Roads as conduits for exotic plant invasions in a semiarid landscape. Conserv Biol 17:420–432. https://doi.org/10.1046/j.1523-1739.2003.01408.x
Gibson D, Blomberg EJ, Atamian MT, Sedinger JS (2014) Lek fidelity and movement among leks by male greater sage-grouse Centrocercus urophasianus: a capture-mark-recapture approach. Ibis 156:729–740. https://doi.org/10.1111/ibi.12192
Gibson D, Blomberg EJ, Atamian MT, Sedinger JS (2016a) Nesting habitat selection influences nest and early offspring survival in greater sage-grouse. Condor: Ornithol Appl 118:689–702. https://doi.org/10.1650/CONDOR-16-62.1
Gibson D, Blomberg EJ, Sedinger JS (2016b) Evaluating vegetation effects on animal demographics: the role of plant phenology and sampling bias. Ecol Evol 6:3621–3631. https://doi.org/10.1002/ece3.2148
Goosey HB, Smith JT, O’Neill KM, Naugle DE (2019) Ground-dwelling arthropod community response to livestock grazing: implications for avian conservation. Environ Entomol 48:856–866. https://doi.org/10.1093/ee/nvz074
Government of Canada (2021) Greater sage-grouse. https://www.canada.ca/en/environment-climate-change/services/species-risk-education-centre/greater-sage-grouse.html. Accessed 27 Aug 2021
Guttery MR, Dahlgren DK, Messmer TA, Connelly JW, Reese KP, Terletzky PA, Burkepile N, Koons DN (2013) Effects of landscape-scale environmental variation on greater sage-grouse chick survival. PLoS ONE 8:e65582. https://doi.org/10.1371/journal.pone.0065582
Hagen CA, Connelly JW, Schroeder MA (2007) A meta-analysis of greater sage-grouse Centrocercus urophasianus nesting and brood-rearing habitats. Wildl Biol 13:42–50. https://doi.org/10.2981/0909-6396(2007)13[42:AMOGSC]2.0.CO;2
Hagen CA, Sedinger JE, Braun CE (2018) Estimating sex-ratio, survival, and harvest susceptibility in greater sage-grouse: making the most of hunter harvests. Wildl Biol 2018:wlb.00362. https://doi.org/10.2981/wlb.00362.
Hamerlynck EP, Davies KW (2019) Changes in the abundance of eight sagebrush-steppe bunchgrass species 13 yr after coplanting. Rangeland Ecol Manag 72:23–27. https://doi.org/10.1016/j.rama.2018.07.001
Hansen AJ, Knight RL, Marzluff JM, Powell S, Brown K, Gude PH, Jones K (2005) Effects of exurban development on biodiversity: patterns, mechanisms, and research needs. Ecol Appl 15:1893–1905. https://doi.org/10.1890/05-5221
Hansen MJ, Wilson SD (2006) Is management of an invasive grass Agropyron cristatum contingent on environmental variation? J Appl Ecol 43:269–280. https://doi.org/10.1111/j.1365-2664.2006.01145.x
Harju SM, Coates PS, Dettenmaier SJ, Dinkins JB, Jackson PJ, Chenaille MP (2021) Estimating trends of common raven populations in North America, 1966–2018. Hum Wildl Interact 15:In press. https://doi.org/10.26077/c27f-e335
Harte J, Saleska SR, Levy C (2015) Convergent ecosystem responses to 23-year ambient and manipulated warming link advancing snowmelt and shrub encroachment to transient and long-term climate- soil carbon feedback. Glob Change Biol 21:2349–2356. https://doi.org/10.1111/gcb.12831
Heidinga L, Wilson SD (2002) The impact of an invading alien grass (Agropyron cristatum) on species turnover in native prairie. Divers Distrib 8:249–258. https://doi.org/10.1046/j.1472-4642.2002.00154.x
Heinrichs JA, O’Donnell MS, Aldridge CL, Garman SL, Homer CG (2019) Influences of potential oil and gas development and future climate on sage-grouse declines and redistribution. Ecol Appl 29:e01912. https://doi.org/10.1002/eap.1912
Hennig JD, Beck JL, Duchardt CJ, Scasta JD (2021) Variation in sage-grouse habitat quality metrics across a gradient of feral horse use. J Arid Environ 192:104550. https://doi.org/10.1016/j.aridenv.2021.104550
Holloran MJ, Heath BJ, Lyon AG, Slater SJ, Kuipers JL, Anderson SH (2005) Greater sage-grouse nesting habitat selection and success in Wyoming. J Wildl Manage 69:638–649. https://doi.org/10.2193/0022-541X(2005)069[0638:GSNHSA]2.0.CO;2
Holmes AL, Maestas JD, Naugle DE (2017) Bird responses to removal of western juniper in sagebrush-steppe. Rangeland Ecol Manag 70:87–94. https://doi.org/10.1016/j.rama.2016.10.006
Homer CG, Xian G, Aldridge CL, Meyer DK, Loveland TR, O’Donnell MS (2015) Forecasting sagebrush ecosystem components and greater sage-grouse habitat for 2050: learning from past climate patterns and Landsat imagery to predict the future. Ecol Indic 55:131–145. https://doi.org/10.1016/j.ecolind.2015.03.002
Honess RF (1982) Cestodes of grouse. In: Thorne ET, Kingston N, Jolley WR, Bergstrom RC (eds) Diseases of wildlife in Wyoming, second edition. Wyoming Game and Fish Department, Cheyenne, Wyoming, pp 161–164
Honess RF, Post G (1968) History of an epizootic in sage grouse. University of Wyoming Agricultural Experiment Station, Science Monograph 14, 32 p
Hupp JW, Braun CE (1989) Topographic distribution of sage grouse foraging in winter. J Wildl Manage 53:823–829. https://doi.org/10.2307/3809220
Huwer SL, Anderson DR, Remington TE, White GC (2008) Using human-imprinted chicks to evaluate the importance of forbs to sage-grouse. J Wildl Manage 72:1622–1627. https://doi.org/10.2193/2004-340
James JJ, Rinella MJ, Svejcar T (2012) Grass seedling demography and sagebrush steppe restoration. Rangeland Ecol Manag 65:409–417. https://doi.org/10.2111/REM-D-11-00138.1
Jenni DA, Hartzler JE (1978) Attendance at a sage grouse lek: implications for spring censuses. J Wildl Manage 42:46–52. https://doi.org/10.2307/3800688
Johnson DH (2008) In defense of indices: the case of bird surveys. J Wildl Manage 72:857–868. https://doi.org/10.2193/2007-294
Johnson GD, Boyce MS (1990) Feeding trials with insects in the diet of sage grouse chicks. J Wildl Manage 54:89–91. https://doi.org/10.2307/3808906
Kiesecker JM, Naugle DE (eds) (2017) Energy sprawl solutions: balancing global development and conservation. Island Press, Washington, D.C
King SR, Schoenecker KA, Manier DJ (2019) Potential spread of cheatgrass and other invasive species by feral horses in western Colorado. Rangeland Ecol Manag 72:706–710. https://doi.org/10.1016/j.rama.2019.02.006
Kirol CP, Beck JL, Huzurbazar SV, Holloran MJ, Miller SN (2015) Identifying greater sage-grouse source and sink habitats for conservation planning in an energy development landscape. Ecol Appl 25:968–990. https://doi.org/10.6084/m9.figshare.c.3296837.v1
Klebenow DA, Gray GM (1968) Food habits of juvenile sage grouse. J Range Manage 21:80–83. https://doi.org/10.2307/3896359
Klos PZ, Link TE, Abatzoglou JT (2014) Extent of the rain-snow transition zone in the western U.S. under historic and projected climate. Geophys Res Lett 41:4560–4568. https://doi.org/10.1002/2014GL060500
Knapp PA (1996) Cheatgrass (Bromus tectorum L) dominance in the Great Basin Desert: history, persistence, and influences to human activities. Glob Environ Change 6:37–52. https://doi.org/10.1016/0959-3780(95)00112-3
Knick ST (2011) Historical development, principal federal legislation, and current management of sagebrush habitats: implications for conservation. In: Knick ST, Connelly JW (eds) Greater sage‐grouse: ecology and conservation of a landscape species and its habitats. Studies in avian biology 38, University of California Press, Berkeley, pp 13–31. https://doi.org/10.1525/California/9780520267114.003.0002
Knick ST, Dobkin DS, Rotenberry JT, Schroeder MA, Vander Haegen WM, van Riper IIIC (2003) Teetering on the edge or too late? Conservation and research issues for avifauna of sagebrush habitats. Condor 105:611–634. https://doi.org/10.1093/condor/105.4.611
Knight RL, Wallace GN, Riebsame WE (1995) Ranching the view: subdivisions versus agriculture. Conserv Biol 9:459–461. https://www.jstor.org/stable/2386971
Knutson KC, Pyke DA, Wirth TA, Arkle RS, Pilliod DS, Brooks ML, Chambers JC, Grace JB (2014) Long-term effects of seeding after wildfire on vegetation in Great Basin shrubland ecosystems. J Appl Ecol 51:1414–1424. https://doi.org/10.1111/1365-2664.12309
Kohl KD, Connelly JW, Dearing MD, Forbey JS (2016) Microbial detoxification in the gut of a specialist avian herbivore, the greater sage-grouse. FEMS Microbiol Lett 363:fnw144. https://doi.org/10.1093/femsle/fnw144
Küchler AW (1985) Potential natural vegetation. US Geological Survey. https://doi.org/10.3133/32574
Lesica P, DeLuca TH (1996) Long-term harmful effects of crested wheatgrass on Great Plains grassland ecosystems. J Soil Water Conserv 51:408–409
Lockwood JA, Anderson-Sprecher R, Schell SP (2002) When less is more: optimization of reduced agent-area treatments (RAATs) for management of rangeland grasshoppers. Crop Prot 21:551–562. https://doi.org/10.1016/S0261-2194(01)00145-4
López-Osorio S, Chaparro-Gutiérrez JJ, Gómez-Osorio LM (2020) Overview of poultry Eimeria life cycle and host-parasite interactions. Front Vet Sci 7:384. https://doi.org/10.3389/fvets.2020.00384
Lyon AG, Anderson SH (2003) Potential gas development impacts on sage grouse nest initiation and movement. Wildl Soc Bull 31:486–491. https://www.jstor.org/stable/3784329
Mack RN (1981) Invasion of Bromus tectorum L. into western North America: an ecological chronicle. Agro-Ecosystems 7:145–165. https://doi.org/10.1016/0304-3746(81)90027-5
Maestas JD, Knight RL, Gilgert WC (2003) Biodiversity across a rural land-use gradient. Conserv Biol 17:1425–1434. https://doi.org/10.1046/j.1523-1739.2003.02371.x
Maestas JD, Campbell SB, Chambers JC, Pellant M, Miller RF (2016) Tapping soil survey information for rapid assessment of sagebrush ecosystem resilience and resistance. Rangelands 38:120–128. https://doi.org/10.1016/j.rala.2016.02.002
Martínez MI, Lumaret J-P, Ortiz Zayas R, Kadiri N (2017) The effects of sublethal and lethal doses of ivermectin on the reproductive physiology and larval development of the dung beetle Euoniticellus intermedius (Coleoptera: Scarabaeidae). Can Entomol 149:461–472. https://doi.org/10.4039/tce.2017.11
McCaffery R, Lukacs PM (2016) A generalized integrated population model to estimate greater sage-grouse population dynamics. Ecosphere 7:e01585. https://doi.org/10.1002/ecs2.1585
McCaffery R, Nowak JJ, Lukacs PM (2016) Improved analysis of lek count data using N-mixture models. J Wildl Manage 80:1011–1021. https://doi.org/10.1002/jwmg.21094
McClanahan RC (1940) Original and present breeding ranges of certain game birds in the United States. United States Bureau of Biological Survey, Wildlife Leaflet BS-158, Washington, DC
Meinke CW, Knick ST, Pyke DA (2009) A spatial model to prioritize sagebrush landscapes in the Intermountain West (U.S.A.) for restoration. Restor Ecol 17:652–659. https://doi.org/10.1111/j.1526-100X.2008.00400.x
Miller RF, Rose JA (1999) Fire history and western juniper encroachment in sagebrush steppe. J Range Manage 52:550–559. https://doi.org/10.2307/4003623
Miller RF, Wigand PE (1994) Holocene changes in semiarid pinyon-juniper woodlands: response to climate, fire, and human activities in the US Great Basin. Bioscience 44:465–474. https://doi.org/10.2307/1312298
Miller RF, Svejcar TJ, Rose JA (2000) Impacts of western juniper on plant community composition and structure. J Range Manage 53:574–585. https://doi.org/10.2458/azu_jrm_v53i6_miller
Miller RF, Bates JD, Svejcar TJ, Pierson FB, Eddleman LE (2005) Biology, ecology, and management of western juniper. Tech Bull 152. Oregon State University, Corvallis, Oregon. Available at https://ir.library.oregonstate.edu/concern/technical_reports/cz30pv075. Accessed 7 April 2022
Miller RF, Tausch RJ, MacArthur D, Johnson DD, Sanderson SC (2008) Development of post settlement piñon-juniper woodlands in the Intermountain West: a regional perspective. Research Paper Report RMRS-RP-69. USDA Forest Service, Rocky Mountain Research Station, Ft. Collins, Colorado
Monroe AP, Edmunds DR, Aldridge CL (2016) Effects of lek count protocols on greater sage-grouse population trend estimates. J Wildl Manage 80:667–678. https://doi.org/10.1002/jwmg.1050
Monroe AP, Aldridge CL, Assal TJ, Veblen KE, Pyke DA, Casazza ML (2017) Patterns in greater sage-grouse population dynamics correspond with public grazing records at broad scales. Ecol Appl 27:1096–1107. https://doi.org/10.1002/eap.1512
Monroe AP, Wann GT, Aldridge CL, Coates PS (2019) The importance of simulation assumptions when evaluating detectability in population models. Ecosphere 10:e02791. https://doi.org/10.1002/ecs2.2791
Muñoz DA, Coates PS, Ricca MA (2021) Free-roaming horses disrupt greater sage-grouse lekking activity in the Great Basin. J Arid Environ 184:104304. https://doi.org/10.1016/j.jaridenv.2020.104304
Natural Resources Conservation Service [NRCS] (2021) A decade of science support in the sagebrush biome. United States Department of Agriculture, Washington, D.C. 32 p. https://doi.org/10.32747/2021.7488985
Naugle DE, Aldridge CL, Walker BL, Doherty KE, Matchett MR, McIntosh J, Cornish TE, Boyce MS (2005) West Nile virus and sage-grouse: what more have we learned? Wildl Soc Bull 33:616–623. https://doi.org/10.2193/0091-7648(2005)33[616:WNVASW]2.0.CO;2
Naugle DE, Allred BW, Jones MO, Twidwell D, Maestas JD (2020) Coproducing science to inform working lands: the next frontier in nature conservation. Bioscience 70:90–96. https://doi.org/10.1093/biosci/biz144
Naugle DE, Doherty KE, Walker BL, Holloran MJ, Copeland HE (2011) Energy development and greater sage-grouse. In: Knick ST, Connelly JW (eds) Greater sage‐grouse: ecology and conservation of a landscape species and its habitats. Studies in avian biology 38, University of California Press, Berkeley, pp 489–503. https://doi.org/10.1525/California/9780520267114.003.0021
Naugle DE, Maestas JD, Allred BW, Hagen CA, Jones MO, Falkowski MJ, Randall B, Rewa CA (2019) CEAP quantifies conservation outcomes for wildlife and people on western grazing lands. Rangelands 41:211–217. https://doi.org/10.1016/j.rala.2019.07.004
Neel LA (1980) Sage grouse response to grazing management in Nevada, Thesis, University of Nevada, Reno
Newton RE, Tack JD, Carlson JC, Matchett MR, Fargey PJ, Naugle DE (2017) Longest sage-grouse migratory behavior sustained by intact pathways. J Wildl Manage 81:962–972. https://doi.org/10.1002/jwmg.21274
Nichols JD, Thomas L, Conn PB (2009) Inferences about landbird abundance from count data: recent advances and future directions. In: Thomson DL, Cooch EG, Conroy MJ (eds) Modeling demographic processes in marked populations. Springer, New York, New York, USA, pp 201–235
Oh KP, Aldridge CL, Forbey JS, Dadabay CY, Oyler-McCance SJ (2019) Conservation genomics in the sagebrush sea: population divergence, demographic history, and local adaptation in sage-grouse (Centrocercus spp.). Genome Biol Evol 11:2023–2034. https://doi.org/10.1093/gbe/evz112
Olsen AC, Severson JP, Maestas JD, Naugle DE, Smith JT, Tack JD, Yates KH, Hagen CA (2021) Reversing tree expansion in sagebrush steppe yields population-level benefit for imperiled grouse. Ecosphere 12:e03551. https://doi.org/10.1002/ecs2.3551
Oyler-McCance SJ, Quinn TW (2011) Molecular insights into the biology of greater sage-grouse. In: Knick ST, Connelly JW (eds) Greater sage‐grouse: ecology and conservation of a landscape species and its habitats. Studies in avian biology 38, University of California Press, Berkeley, pp 85–94. https://doi.org/10.1525/california/9780520267114.003.0006
Oyler-McCance SJ, St. John J, Taylor SE, Apa AD, Quinn TW (2005) Population genetics of Gunnison sage-grouse: implications for management. J Wildl Manage 69:630–637.https://doi.org/10.2193/0022-541X(2005)069[0630:PGOGSI]2.0.CO;2
Palmquist KA, Schlaepfer DR, Bradford JB, Lauenroth WK (2016) Mid-latitude shrub steppe plant communities: climate change consequences for soil water resources. Ecology 97:2342–2354. https://doi.org/10.1002/ecy.1457
Pennington VE, Schlaepfer DR, Beck JL, Bradford JB, Palmquist KA, Lauenroth WK (2016) Sagebrush, greater sage-grouse, and the occurrence and importance of forbs. West N Am Nat 76:298–312. https://doi.org/10.3398/064.076.0307
Peterson JG (1970) The food habits and summer distribution of juvenile sage grouse in central Montana. J Wildl Manage 34:147–155. https://doi.org/10.2307/3799502
Peterson MJ (2004) Parasites and infectious diseases of prairie grouse: should managers be concerned? Wildl Soc Bull 32:35–55. https://doi.org/10.2193/0091-7648(2004)32[35:PAIDOP]2.0.CO;2
Pierson FB, Bates JD, Svejcar TJ, Hardegree SP (2007) Runoff and erosion after cutting western juniper. Rangeland Ecol Manag 60:285–292. https://doi.org/10.2111/1551-5028(2007)60[285:RAEAECW]2.0.CO;2
Pratt AC, Smith KT, Beck JL (2017) Environmental cues used by greater sage-grouse to initiate altitudinal migration. Auk: Ornithol Adv 134:628–643. https://doi.org/10.1642/AUK-16-192.1
Pratt AC, Smith KT, Beck JL (2019) Prioritizing seasonal habitats for comprehensive conservation of a partially migratory species. Glob Ecol Conserv 17:e00594. https://doi.org/10.1016/j.gecco.2019.e00594
Post G (1951) Effects of toxaphene and chlordane on certain game birds. J Wildl Manage 15:381–386. https://doi.org/10.2307/3796581
Prochazka BG, Coates PS, Ricca MA, Casazza ML, Gustafson KB, Hull JM (2017) Encounters with pinyon-juniper influence riskier movements in greater sage-grouse across the Great Basin. Rangel Ecol Manag 70:39–49. https://doi.org/10.1016/j.rama.2016.07.004
Pyke DA, Knick ST, Chambers JC, Pellant M, Miller RF, Beck JL, Doescher PS, Schupp EW, Roundy BA, Brunson M, McIver JD (2015b) Restoration handbook for sagebrush steppe ecosystems with emphasis on greater sage-grouse habitat—Part 2. Landscape level restoration decisions. U.S. Geological Survey Circular 1418, 21 p. https://doi.org/10.3133/cir1418.
Pyke DA, Chambers JC, Pellant M, Knick ST, Miller RF, Beck JL, Doescher PS, Schupp EW, Roundy BA, Brunson M, McIver JD (2015a) Restoration handbook for sagebrush steppe ecosystems with emphasis on greater sage-grouse habitat—Part 1. Concepts for understanding and applying restoration: U.S. Geological Survey Circular 1416, 44 p. https://doi.org/10.3133/cir1416
Pyke DA, Chambers JC, Pellant M, Miller RF, Beck JL, Doescher PS, Roundy BA, Schupp EW, Knick ST, Brunson M, McIver JD (2017) Restoration handbook for sagebrush steppe ecosystems with emphasis on greater sage-grouse habitat—Part 3. Site level restoration decisions. U.S. Geological Survey Circular 1426, 62 p. https://doi.org/10.3133/cir1426
Randall KJ, Ellison MJ, Yelich JV, Price WJ, Johnson TN (2022) Managing forbs preferred by greater sage-grouse and soil moisture in mesic meadows with short-duration grazing. Rangeland Ecol Manag 82:66–75. https://doi.org/10.1016/j.rama.2022.02.008
Reese KP, Connelly JW (1997) Translocation of sage grouse Centrocercus urophasianus in North America. Wildlife Biol 3:235–241
Reeves MC, Moreno AL, Bagne KE, Running SW (2014) Estimating climate change effects on net primary production of rangelands in the United States. Clim Change 126:429–442. https://doi.org/10.1007/s10584-014-1235-8
Reinhart JR, Filippelli S, Falkowski M, Allred B, Maestas JD, Carlson JC, Naugle DE (2020) Quantifying pinyon-juniper reduction within North America’s sagebrush ecosystem. Rangeland Ecol Manag 73:420–432. https://doi.org/10.1016/j.rama.2020.01.002
Renne RR, Schlaepfer DR, Palmquist KA, Bradford JB, Burke IC, Lauenroth WK (2019) Soil and stand structure explain shrub mortality patterns following global change–type drought and extreme precipitation. Ecology 100:e02889. https://doi.org/10.1002/ecy.2889
Romme WH, Allen CD, Bailey JD, Baker WL, Bestelmeyer BT, Brown PM, Eisenhart KS, Floyd ML, Huffman DW, Jacobs BF, Miller RF (2009) Historical and modern disturbance regimes, stand structures, and landscape dynamics in pinon–juniper vegetation of the western United States. Rangeland Ecol Manag 62:203–222. https://doi.org/10.2111/08-188R1.1
Rowland MM, Wisdom MJ, Suring LH, Meinke CW (2006) Greater sage-grouse as an umbrella species for sagebrush-associated vertebrates. Biol Conserv 129:323–335. https://doi.org/10.1016/j.biocon.2005.10.048
Sanderson JS, Beutler C, Brown JR, Burke I, Chapman T, Conant RT, Derner JD, Easter M, Fuhlendorf SD, Grissom G, Herrick JE, Liptzin D, Morgan JA, Murph R, Pague C, Rangwala I, Ray D, Rondeau R, Schulz T, Sullivan T (2020) Cattle, conservation, and carbon in the western Great Plains. J Soil Water Conserv 75:5A-12A. https://doi.org/10.2489/jswc.75.1.5A
Sauls HS (2006) The role of selective foraging and cecal microflora in sage-grouse nutritional ecology. Thesis, University of Montana, Missoula, Montana
Scasta JD, Hennig JD, Beck JL (2018) Framing contemporary U.S. wild horse and burro management processes in a dynamic ecological, sociological, and political environment. Hum Wildl Interact 12:31–45. https://doi.org/10.26077/2fhw-fz24
Schlaepfer DR, Lauenroth WK, Bradford JB (2012) Consequences of declining snow accumulation for water balance of mid-latitude dry regions. Glob Change Biol 18:1988–1997. https://doi.org/10.1111/j.1365-2486.2012.02642.x
Schlaepfer DR, Taylor KA, Pennington VE, Nelson KN, Martyn TE, Rottler CM, Lauenroth WK, Bradford JB (2015) Simulated big sagebrush regeneration supports predicted changes at the trailing and leading edges of distribution shifts. Ecosphere 6:1–31. https://doi.org/10.1890/ES14-00208.1
Schroeder MA, Aldridge CL, Apa AD, Bohne JR, Braun CE, Bunnell SD, Connelly JW, Deibert PA, Gardner SC, Hilliard MA, Kobriger GD, McAdam SM, McCarthy CW, McCarthy JJ, Mitchell DL, Rickerson EV, Stiver SJ (2004) Distribution of sage-grouse in North America. Condor 106:363–376. https://doi.org/10.1093/condor/106.2.363
Schroeder MA, Young JR, Braun CE (2020) Greater Sage-Grouse (Centrocercus urophasianus), version 1.0. In: Poole AF, Gill FB (eds) Birds of the world. Cornell Lab of Ornithology, Ithaca, New York. https://doi.org/10.2173/bow.saggro.01
Sedinger JS, White GC, Espinosa S, Partee ET, Braun CE (2010) Assessing compensatory versus additive harvest mortality: an example using greater sage-grouse. J Wildl Manage 74:326–332. https://doi.org/10.2193/2009-071
Severson JP, Hagen CA, Maestas JD, Naugle DE, Forbes JT, Reese KP (2016) Effects of conifer expansion on greater sage-grouse nesting habitat selection. J Wildl Manage 81:86–95. https://doi.org/10.1002/jwmg.21183
Severson JP, Coates PS, Prochazka BG, Ricca MA, Casazza ML, Delehanty DJ (2019) Global positioning system tracking devices can decrease greater sage-grouse survival. Condor: Ornithol Appl 121:1–15. https://doi.org/10.1093/condor/duz032
Shyvers JE, Walker BL, Noon BR (2018) Dual-frame lek surveys for estimating greater sage-grouse populations. J Wildl Manage 82:1689–1700. https://doi.org/10.1002/jwmg.21540
Shyvers JE, Walker BL, Oyler-McCance SJ, Fike JA, Noon BR (2019) Genetic mark-recapture analysis of winter faecal pellets allow estimation of population size in sage grouse Centrocercus urophasianus. Ibis 162:749–765. https://doi.org/10.1111/ibi.12768
Silverman NL, Allred BW, Donnelly JP, Chapman TB, Maestas JD, Wheaton JM, White J, Naugle DE (2019) Low-tech riparian and wet meadow restoration increases vegetation productivity and resilience across semiarid rangelands. Restor Ecol 27:269–278. https://doi.org/10.1111/rec.12869
Sinai NL, Coates PS, Andrle KM, Jefferis C, Sentíes-Cué CG, Pitesky ME (2017) A serosurvey of greater sage-grouse (Centrocercus urophasianus) in Nevada, USA. J Wildl Dis 53:136–139. https://doi.org/10.7589/2015-10-285
Smith KT, Beck JL (2018) Sagebrush treatments influence annual population change for greater sage-grouse. Restor Ecol 26:497–505. https://doi.org/10.1111/rec.12589
Smith KT, Beck JL, Pratt AC (2016) Does Wyoming’s Core Area Policy protect winter habitats for greater sage-grouse? Environ Manage 58:585–596. https://doi.org/10.1007/s00267-016-0745-8
Smith JT, Tack JD, Berkeley LI, Szczypinski M, Naugle DE (2018a) Effects of rotational grazing management on nesting greater sage-grouse. J Wildl Manage 82:103–112. https://doi.org/10.1002/jwmg.21344
Smith JT, Tack JD, Doherty KE, Allred BW, Maestas JD, Berkeley LI, Dettenmaier SJ, Messmer TA, Naugle DE (2018b) Phenology largely explains taller grass at successful nests in greater sage-grouse. Ecol Evol 8:356–364. https://doi.org/10.1002/ece3.3679
Smith KT, Beck JL, Kirol CP (2018c) Reproductive state leads to intraspecific habitat partitioning and survival differences in greater sage-grouse: implications for conservation. Wildl Res 45:119–131. https://doi.org/10.1071/WR17123
Smith KT, Pratt AC, LeVan JR, Rhea AM, Beck JL (2019) Reconstructing greater sage-grouse chick diets: diet selection, body condition, and food availability at brood-rearing sites. Condor: Ornithol Appl 121:duy012. https://doi.org/10.1093/condor/duy012
Smith JT, Allred BW, Boyd CS, Carlson JC, Davies KW, Hagen CA, Naugle DE, Olsen AC, Tack JD (2020) Are sage-grouse fine-scale specialists or shrub-steppe generalists? J Wildl Manag 84:759–774. https://doi.org/10.1002/jwmg.21837
Smith JT, Allred BW, Boyd CS, Davies KW, Jones MO, Kleinhesselink AR, Maestas JD, Morford SL, Naugle DE (2022) The elevational ascent and spread of exotic annual grass dominance in the Great Basin, USA. Divers Distrib 28:83–96. https://doi.org/10.1111/ddi.13440
Smith KT, LeVan JR, Chalfoun AD, Christiansen TJ, Harter SR, Oberlie S, Beck, JL (2023) Response of greater sage-grouse to sagebrush reduction treatments in Wyoming big sagebrush. Wildl Monogr 212:e1075. https://doi.org/10.1002/wmon.1075
Spence ES, Beck JL, Gregory AJ (2017) Probability of lek collapse is lower inside sage-grouse Core Areas: effectiveness of conservation policy for a landscape species. PLoS ONE 12:e0185885. https://doi.org/10.1371/journal.pone.0185885
Stevens BS, Connelly JW, Reese KP (2012a) Multi-scale assessment of greater sage-grouse fence collision as a function of site and broad scale factors. J Wildl Manage 76:1370–1380. https://doi.org/10.1002/jwmg.397
Stevens BS, Reese KP, Connelly JW, Musil DD (2012b) Greater sage-grouse and fences: does marking reduce collisions? Wildl Soc Bull 36:297–303. https://doi.org/10.1002/wsb.142
Stewart G, Hull AC (1949) Cheatgrass (Bromus tectorum L.)—an ecologic intruder in southern Idaho. Ecology 30:58–74. https://doi.org/10.2307/193227
Stiver SJ, Apa AD, Bohne J, Bunnell SD, Deibert P, Gardner S, Hilliard M, McCarthy C, Schroeder MA (2006) Greater sage-grouse comprehensive conservation strategy. Unpublished Report, Western Association of Fish and Wildlife Agencies, Cheyenne, Wyoming, 442 pp
Stiver SJ, Rinkes ET, Naugle DE, Makela PD, Nance DA, Karl JW (eds) (2015) Sage-grouse habitat assessment framework: a multiscale assessment tool. Technical reference 6710-1. Bureau of Land Management and Western Association of Fish and Wildlife Agencies, Denver, Colorado
Stohlgren TJ, Schnase JL (2006) Risk analysis for biological hazards: what we need to know about invasive species. Risk Anal 26:163–173. https://doi.org/10.1111/j.1539-6924.2006.00707.x
Street PA (2020) Greater sage-grouse habitat and demographic responses to grazing by non-native ungulates. Dissertation, University of Nevada, Reno. http://hdl.handle.net/11714/7709
Swanson S, Wyman S, Evans C (2015) Practical grazing management to maintain or restore riparian functions and values on rangelands. J Rangeland Appl 2:1–28. https://thejra.nkn.uidaho.edu/index.php/jra/article/view/20/39
Syphard AD, Radeloff VC, Hawbaker TJ, Stewart SI (2009) Conservation threats due to human-caused increases in fire frequency in Mediterranean-climate ecosystems. Conserv Biol 23:758–769. https://doi.org/10.1111/j.1523-1739.2009.01223.x
Tack JD, Jakes AF, Jones PF, Smith JT, Newton RE, Martin BH, Hebblewhite M, Naugle DE (2019) Beyond protected areas: private lands and public policy anchor intact pathways for multi-species wildlife migration. Biol Cons 234:18–27. https://doi.org/10.1016/j.biocon.2019.03.017
Tausch RJ, West NE, Nabi AA (1981) Tree age and dominance patterns in Great Basin pinyon-juniper woodlands. J Range Manage 34:259–264. https://doi.org/10.2307/3897846
Taylor RL, Walker BL, Naugle DE, Mills LS (2012) Managing multiple vital rates to maximize greater sage-grouse population growth. J Wildl Manage 76:336–347. https://doi.org/10.1002/jwmg.267
Taylor RL, Tack JD, Naugle DE, Mills LS (2013) Combined effects of energy development and disease on greater sage-grouse. PLoS ONE 8:e71256. https://doi.org/10.1371/journal.pone.0071256
Thompson KM, Holloran MJ, Slater SJ, Kuipers JL, Anderson SH (2006) Early brood-rearing habitat use and productivity of greater sage-grouse in Wyoming. West N Am Nat 66:332–342. https://doi.org/10.3398/1527-0904(2006)66[332:EBHUAP]2.0.CO;2
Urza AK, Weisberg PJ, Chambers JC, Board D, Flake SW (2019) Seeding native species increases resistance to annual grass invasion following prescribed burning of semiarid woodlands. Biol Invasions 21:1993–2007. https://doi.org/10.1007/s10530-019-01951-9
U.S. Fish and Wildlife Service [USFWS] (2013) Greater Sage-grouse (Centrocercus urophasianus) Conservation Objectives: Final Report. U.S. Fish and Wildlife Service, Denver Colorado
U.S. Fish and Wildlife Service [USFWS] (2014) Endangered and threatened wildlife and plants: threatened status for Gunnison sage-grouse. Fed Reg 79:69191–69310
U.S. Fish and Wildlife Service [USFWS] (2015) Endangered and threatened wildlife and plants; 12-month finding on a petition to list the Greater sage-grouse (Centrocercus urophasianus) as an endangered or threatened species. Proposed Rule. Federal Register 80:59858–59942
Van Lanen NJ, Green AW, Gorman TR, Quattrini LA, Pavlacky DC Jr (2017) Evaluating efficacy of fence markers in reducing greater sage-grouse collisions with fencing. Biol Conserv 213:70–83. https://doi.org/10.1016/j.biocon.2017.06.030
Veblen KE, Pyke DA, Aldridge CL, Casazza ML, Assal TJ, Farinha MA (2014) Monitoring of livestock grazing effects on Bureau of Land Management land. Rangeland Ecol Manage 67:68–77. https://doi.org/10.2111/REM-D-12-00178.1
Walker BL, Naugle DE (2011) West Nile virus ecology in sagebrush habitat and impacts on greater sage‐grouse populations. In: Knick ST, Connelly JW (eds) Greater sage‐grouse: ecology and conservation of a landscape species and its habitats. Studies in Avian Biology 38, University of California Press, Berkeley, pp 127–142. https://doi.org/10.1525/California/9780520267114.003.0010
Walsh DP, White GC, Remington TE, Bowden DC (2004) Evaluation of the lek-count index for greater sage-grouse. Wildl Soc Bull 32:56–68. https://doi.org/10.2193/0091-7648(2004)32[56:EOTLIF]2.0.CO;2
Wambolt CL, Harp AJ, Welch BL, Shaw N, Connelly JW, Reese KP, Braun CE, Klebenow DA, McArthur ED, Thompson JG, Torrell LA, Tanaka JA (2002) Conservation of greater sage-grouse on public lands in the western U.S.: implications of recovery and management policies. Policy Analysis Center for Western Public Lands Policy Paper SG-02-02, Caldwell, Idaho, 41 p
Wann GT, Coates PS, Prochazka BG, Severson JP, Monroe AP, Aldridge CL (2019) Assessing lek attendance of male greater sage-grouse using fine-resolution GPS data: implications for population monitoring of lek mating grouse. Popul Ecol 61:183–197. https://doi.org/10.1002/1438-390X.1019
Wann GT, Braun CE, Aldridge CL, Schroeder MA (2020) Rates of ovulation and reproductive success estimated from hunter-harvested greater sage-grouse in Colorado. J Fish Wildl Manag 11:151–163. https://doi.org/10.3996/072019-JFWM-063
Watchorn RT, Maechtle T, Fedy BC (2018) Assessing the efficacy of fathead minnows (Pimephales promelas) for mosquito control. PLoS ONE 13:e0194304. https://doi.org/10.1371/journal.pone.0194304
West NE, Provenza FD, Johnson PS, Owens MK (1984) Vegetation change after 13 years of livestock grazing exclusion on sagebrush semidesert in west central Utah. J Range Manage 37:262–264. https://doi.org/10.2307/3899152
Western Governors’ Association (2020) A toolkit for invasive annual grass management in the West. Available at: https://westgov.org/images/editor/FINAL_Cheatgrass_Toolkit_July_2020.pdf. Accessed 7 April 2022
Wilken E, Nava FJ, Griffith G (2011) North American terrestrial ecoregions—Level III. Commission for Environmental Cooperation, Montreal, Canada
Wolfe AL, Broughton JM (2016) Chapter 14 - Bonneville Basin avifaunal change at the Pleistocene/Holocene transition: evidence from Homestead Cave. In: Oviatt CG, Shroder JF (eds) Lake Bonneville: a scientific update. Developments in earth surface processes 20, pp 371–419. https://doi.org/10.1016/B978-0-444-63590-7.00014-7
Young JR, Braun CE, Oyler-McCance SJ, Aldridge CL, Magee PA, Schroeder MA (2020) Gunnison sage-grouse (Centrocercus minimus), version 1.0. In: Rodewald PG (ed) Birds of the World. Cornell Lab of Ornithology, Ithaca, New York. https://doi.org/10.2173/bow.gusgro.01
Zablan MA, Braun CE, White GC (2003) Estimation of greater sage-grouse survival in North Park, Colorado. J Wildl Manage 67:144–154. https://doi.org/10.2307/3803070
Ziska LH, Reeves JB III, Blank B (2005) The impact of recent increases in atmospheric CO2 on biomass production and vegetative retention of cheatgrass (Bromus tectorum): implications for fire disturbance. Glob Change Biol 11:1325–1332. https://doi.org/10.1111/j.1365-2486.2005.00992.x
Zou L, Miller SN, Schmidtmann ET (2006) Mosquito larval habitat mapping using remote sensing and GIS: implications of coalbed methane development and West Nile virus. Med Entomol 43:1034–1041. https://doi.org/10.1093/jmedent/43.5.1034
Zwickel FC, Schroeder MA (2003) Grouse of the Lewis and Clark expedition, 1803 to 1806. Northwest Nat 84:1–19. https://doi.org/10.2307/3536717
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2023 The Author(s)
About this chapter

Cite this chapter
Beck, J.L. et al. (2023). Sage-Grouse. In: McNew, L.B., Dahlgren, D.K., Beck, J.L. (eds) Rangeland Wildlife Ecology and Conservation . Springer, Cham. https://doi.org/10.1007/978-3-031-34037-6_10
Download citation
DOI: https://doi.org/10.1007/978-3-031-34037-6_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-34036-9
Online ISBN: 978-3-031-34037-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)