
Abstract
Savanna rangelands cover large areas of southern Africa. They provide ecosystem functions and services that are essential for the livelihoods of people. However, intense land use and climate change, particularly drought, threaten biodiversity and ecosystem functions of savanna rangelands. Understanding how these factors interact is essential to inform policymakers and to develop sustainable land-use strategies. We applied three different approaches to understand the impacts of drought and grazing on rangeland vegetation: observations, experimentation and modeling. Here, we summarize and compare the main results from these approaches. Specifically, we demonstrate that all approaches consistently show declines in biomass and productivity in response to drought periods, as well as changes in community composition toward annual grasses and forbs. Vegetation recovered after drought periods, indicating vegetation resilience. However, model extrapolation until 2030 showed that vegetation attributes such as biomass and community composition did not recover to values simulated under no-drought conditions during a ten-year period following the drought. We provide policy-relevant recommendations for rangeland management derived from the three approaches. Most importantly, vegetation has a high potential to regenerate and recover during resting periods after disturbance.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Semiarid savannas cover approximately 20% of the global land area (Sankaran et al. 2005) and occupy extensive areas in the global subtropics and tropics of Africa, South America, Asia and northern Australia. Savannas are characterized by a continuous layer of grasses and forbs, interspersed with woody vegetation consisting of trees and shrubs. Fire, mammalian herbivores and pronounced seasonality with distinct dry and wet seasons have shaped the vegetation structure in savannas throughout their evolution. The coexistence of various plant life forms (Linstädter et al. 2014; Siebert and Dreber 2019), their structural heterogeneity and the unique floristic and faunistic elements (Du Toit and Cumming 1999) convey high biodiversity to savannas. Although savanna vegetation has evolved resilience strategies to cope with disturbances such as drought and herbivory (Charles-Dominique et al. 2017; Wigley et al. 2018), intense disturbances, climate change and land-use change may lead to transitions into alternative vegetation states once tipping points are exceeded (Higgins and Scheiter 2012; Pausas and Bond 2020; Staver et al. 2011). Alternative vegetation states can be wood-dominated states (Staver et al. 2011) or degraded states with low vegetation cover and little forage availability for herbivores (Oomen et al. 2016).

Savanna rangelands are influenced by complex interactions between environmental drivers, socioeconomic drivers and vegetation. They experience multiple threats through climate change and land-use change that can lead to a loss of important ecosystem functions and ecosystem services as well as to rangeland degradation
Savanna vegetation provides essential ecosystem services to people and supports their livelihood (Ferner et al. 2018; Matsika et al. 2013; Shackleton et al. 2005; Chap. 15). Direct benefits from communal rangelands (Fig. 16.1, “Ecosystem Services”) include products ranging from cattle grazing, fuelwood for cooking and heating, collection of edible plants, fruits and nuts, to harvesting of medicinal plants (Matsika et al. 2013). Tourism and recreation in national parks and game reserves may create additional revenue (Kalvelage et al. 2020). Yet, nonsustainable land-use practices combined with climate change (Fig. 16.1, “Environmental drivers”) are considerable threats to biodiversity and ecosystem services in savannas (Fig. 16.1, “Threats”). Increasing human population density (Fig. 16.1, “Socioeconomic drivers”) in most cases entails land-use intensification and rising pressure on natural resources that can reduce ecosystem resilience (Buisson et al. 2019). For instance, overstocking of rangelands with domestic livestock can lead to a decrease in rangeland productivity and cause shifts in vegetation composition toward nonpalatable species or woody encroachment (Stevens et al. 2017). Such land-use impacts may result in a degraded state not suitable for livestock (Fig. 16.1, “Adverse Effects”). Resting times as implemented in rotational grazing systems (Savory and Butterfield 2016), or the presence of browsers (Venter et al. 2017, 2018) can counteract such undesirable vegetation states. Whether degraded states are permanent, or vegetation is resilient and can recover (Fig. 16.1, “Landscape resilience”) determines if rangelands will remain available for livestock grazing.
In addition to direct land-use impacts, anthropogenic climate change (Fig. 16.1, “Environmental drivers”) exhibits pressure on savanna vegetation (IPCC 2021; Chap. 7). Rising temperatures and lower annual precipitation combined with an increasing frequency of extensive drought events are projected to negatively affect plant growth and the capacity of rangelands to support herbivores (Ruppert et al. 2015). Elevated atmospheric CO2, on the other hand, may enhance the growth of woody plants and thereby facilitate woody encroachment and transitions to wood-dominated vegetation states (Midgley and Bond 2015).
Given that savanna rangelands are highly dynamic social-ecological systems, it is challenging to predict how climate change, biodiversity loss and land-use pressure may influence future ecosystem functions and services. Interdisciplinary research approaches including scientists, policymakers and stakeholders are required to derive management recommendations and land-use strategies that ensure livelihoods of people in rural areas (Marchant 2010). Complementary research approaches can derive such understanding: (1) Observational approaches allow assessing vegetation dynamics under natural environmental conditions. (2) Experimental approaches allow manipulation of selected biotic and abiotic drivers. (3) Modeling approaches allow integration of system understanding and extrapolation of system behavior. Each approach has specific aims and strengths (Table 16.1). Combining them facilitates acquiring robust knowledge on ecosystem dynamics and deriving policy-relevant management recommendations.
In the two consecutive projects “Limpopo Living Landscapes” (LLL) and “South African Limpopo Landscapes network” (SALLnet), we combined these three approaches to understand the impacts of drought and grazing on savanna vegetation in the Limpopo province, South Africa. Here, we compiled key findings and management recommendations for their sustainable use in the face of drought, while benefiting from the complementarity of observations, experimentation and modeling. Then, we parameterized a vegetation model using data from DroughtAct, a long-term drought and grazing experiment conducted during the two projects, and projected future rangeland vegetation dynamics until 2030. Our research questions are:
-
1.
How does the vegetation in Limpopo’s semiarid rangelands respond to drought and grazing? Do the responses of our three complementary approaches (observations, experimentation and modeling) agree?
-
2.
Is rangeland vegetation resilient to drought events of different lengths, and which role does resting play in this context?
-
3.
What are recommendations for policy and decision-making to improve the resilience of savanna rangelands in the face of drought?
2 Study Area
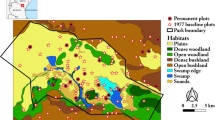
The Limpopo province occupies the Northeast of South Africa (Fig. 16.2). It exhibits considerable variation in soils, topography and climatic conditions. Mean annual precipitation (MAP) ranges between less than 200 mm and more than 1000 mm, and mean annual temperature ranges between 18°C and 28°C (New et al. 2002). The Limpopo province provides rich natural resources that allow the abundant production of agricultural goods such as livestock, vegetables, cereals, fruits and tea. While commercial farms produce these products on a large scale, the Limpopo province also hosts some of the most underprivileged rural areas of South Africa (Lehohla 2012). There, smallholder and subsistence farming rely heavily on available natural resources. Rural communities are particularly vulnerable to environmental change, disruptions such as drought and changes in the social-economic conditions (Gbetibouo 2009; Twine et al. 2003). Overall, approximately 90% of the area of the Limpopo province is utilized by rural and commercial farming, with around 10.5% of the area used for agriculture and 81% used for livestock and game (Maluleke et al. 2016). The Limpopo province also hosts conservation areas and national parks, including parts of the Kruger National Park and the Vhembe Biosphere Reserve (Pool-Stanvliet 2013), creating income through tourism. The current population in the Limpopo province is around 5.9 million people (Stats SA 2021) and growing.

Maps showing the Limpopo Province in South Africa, and the study sites of observational studies (mixed granite bushveld, mopaneveld, Sect. 16.3), and experimental and modeling studies (DroughtAct, Sects. 16.4 and 16.5). Note that the sites of observational studies represent clusters of several study sites
3 Observational Approach
3.1 Background: Observational Approaches to Study Combined Effects of Drought and Grazing
The design of observational studies commonly relies on recording temporal and spatial variation in ecosystem characteristics along local to continent-wide environmental gradients (Table 16.1). For example, study sites can be arranged along steep regional climate gradients to assess the impact of future climatic conditions via a space-for-time substitution (Blois et al. 2013). Moreover, long-term monitoring allows the detection of early warning signals for rapid ecosystem state changes (Arena et al. 2018; Buitenwerf et al. 2011). Observational studies are particularly suitable to understand drought effects on ecosystem function and services for landscapes with several land-use types. In multiple observational studies, species- and trait-based methods have helped assess the extent of taxonomic and functional responses to drought (Ruppert et al. 2015; Wigley-Coetsee and Staver 2020). However, understanding of patterns and drivers of forb communities in African rangeland systems is still limited (Siebert and Dreber 2019). Therefore, our research aimed to assess how forbs, compared to grasses, respond to drought and land-use change or rangeland intensification. We conducted observational studies across ecosystem types with varying annual precipitation. Sites were located in a semiarid mixed granite bushveld (MAP: ~630 mm, 2 sites, Fig. 16.2) and mopaneveld (MAP: ~460 mm, 3 sites, Fig. 16.2) in the Lowveld region of South Africa. The majority of results presented in this section are from two rangeland systems in the semiarid mixed granite bushveld on similar soil, located in a protected area with a high diversity of indigenous mammals (stocking rates maintained at ~0.1 livestock units per hectare, LU/ha), and a communal grazing system (>1.0 LU/ha). Results from the semiarid site were compared to the drier mopaneveld site in the Limpopo Province.
3.2 Data Collection
In our observational approach, we took advantage of a severe, two-year natural drought occurring in southern Africa in the growing seasons 2014/15 and 2015/2016 during which the rainfall for the study sites was ~330 mm below the long-term average (52% and 72% reduction relative to MAP for the two study regions). Complete floristic surveys (i.e., plant individual counts per species) were conducted in October 2016 (representing the “in-drought” survey) within permanent 1 m2 plots situated in a protected area and on communal rangeland, which was repeated in the peak growth season (i.e., January) of the postdrought year 2017. Herbaceous plants were divided into four plant functional types (PFTs): perennial grasses, annual grasses, perennial forbs, annual forbs. Directly adjacent to the permanent plots in both rangeland systems, standing plant biomass was clipped and hand-sorted into forbs and grasses. Biomass was oven-dried (70 ° C, > 48 h) and weighed (Siebert et al. 2020). We derived aboveground net primary production (ANPP) per rangeland type and year from the clipped aboveground standing biomass using the “peak standing biomass” method (Ruppert and Linstädter 2014). We assessed the relative contribution of forbs and grasses to ANPP.
3.3 Data Analysis
Herbaceous species composition across different rangeland types and rainfall was explored using Nonmetric Multidimensional Scaling (NMDS) analyses compiled in PRIMER 6 software. To assess the difference between drought and postdrought herbaceous biomass per life form, a repeated measures analysis of variance (ANOVA) combined with the Bonferroni post-hoc significance test was performed. A two-way ANOVA type Hierarchical Linear Model (HLM) was used to test the effects of rainfall year and rangeland type on variation in grass and forb diversity indices. Significant differences in response to rangeland type and rainfall year were tested using effect sizes (Cohen’s d, Ellis and Steyn 2003).
3.4 Key Results and Discussion
The two-year natural drought had significant effects on ANPP and community composition, irrespective of study sites’ MAP. Although a previous meta-analysis has shown that protected areas with a high proportion of perennial grasses are usually more resistant to drought than those dominated by annual grasses (Ruppert et al. 2015), protected areas and sites with intense grazing pressure were equally affected by the drought. However, protected areas showed a more pronounced postdrought recovery (Fig. 16.3), especially in comparison to sites with a long disturbance history (Klem 2018; Minnaar 2020).

Responses of leaf biomass ratio and the proportion of different PFTs to a 2-year drought in observations, field experiment and modeling. Leaf biomass ratio represents ANPP in observations, ANPP in field experiments and leaf biomass in the modeling approach. Time slices considered are “Pre”: year predrought; “Dr1” and “Dr2”: first and second drought year; “Post1,” “Post2,” “Post3”: three years after drought allowing vegetation recovery. For the observations, drought and postdrought data is based on (Siebert et al. 2020). Predrought biomass ratio was extrapolated using post- and predrought ratios from Van Staden (2016) and Minnaar (2020), due to a lack of predrought data for the study sites
Grass and forb biomass were equally low in both rangeland systems during drought. However, postdrought recovery differed between the two rangeland systems and between the two life forms (Fig. 16.3). Forb biomass increased significantly after the drought at both sites, although the increase was much higher in the protected area. We recorded a five-fold increase in grass biomass postdrought in the protected area, opposed to a much weaker one in the communal rangeland. The proportion of grass to forb biomass was also significantly higher in the protected area (Siebert et al. 2020).
Compared to the communal rangeland, grass species richness and diversity were lower in the protected area during the drought but increased significantly after the drought (Klem 2018). Forb richness and diversity were equally low at both sites and increased significantly postdrought, but in the protected area only. These results suggest that long-term exposure to intensive livestock grazing may deplete the herbaceous seedbank (O’Connor 1991). In the drier mopaneveld study, drought had a similar negative effect on both grass and forb species richness and diversity in a moderately grazed protected area (Minnaar 2020).
During the drought, perennial PFTs dominated the herbaceous layer in both rangeland systems. In the communal rangeland, perennial grasses had a more prostrate, lawn-like growth form, which can be connected to an improved tolerance to combined effects of drought and intensive livestock grazing (Hempson et al. 2015b). Grass and forb PFTs that persisted during the drought were often characterized by clonal growth and bud position close to the soil surface. Both rangeland systems had few annual grasses during the drought. Annual forbs increased in abundance after the drought, but only in the protected area (Klem 2018). This higher postdrought recovery of annuals is in line with the results of a global meta-analysis (Ruppert et al. 2015).
In conclusion, poor rangeland conditions resulting from long-term intensive livestock grazing in the communal rangeland could explain the observed weak positive effects of postdrought rainfall on species and functional composition of grasses (Fynn and O’Connor 2000). Both life forms displayed persistence of generalist tolerator species. Plants, such as Digitaria eriantha and various perennial forb species have a life-history strategy that tolerates defoliation through the ability to resprout from stored resources and a bud bank at or below ground level (Archibald et al. 2019), which can be ascribed to a long evolutionary history of grazing (O’Connor 1995). Although the drought in the growing seasons 2014/15 and 2015/2016 was severe, both rangeland types displayed high ecosystem resilience. However, the duration of this drought event may have been short enough for vegetation recovery after a substantial rain. Possibly, a longer period of below-average rainfall and high temperatures may have led to more pronounced effects on plant community composition (as confirmed by the experiments in Sect. 16.4).
Forb cover and biomass increased in response to postdrought rainfall. Increases in forb cover are often regarded as indicators of land degradation (Camp and Hardy 1999). In the current study, the dominance of perennial forbs in the communal rangeland system during and after drought illustrates their functional importance. Forbs are particularly important for securing forage when grass biomass is low.
3.5 Recommendations Derived from the Results and Outlook
Our results on rangeland vegetation responses to a 2-year natural drought event provided valuable insights into drought resistance of African semiarid savanna ecosystems. This ecosystem type experienced long evolutionary adaptation to grazing by large herbivores. Therefore, African grassland species have developed traits that convey tolerance to heavy browsing, grazing and defoliation by fire and allow rapid postdisturbance recovery. These patterns are similar to what we observed through their postdrought response, irrespective of the rangeland type. Furthermore, we found that the response of different herbaceous PFTs is not necessarily aligned. Changes in the species and functional composition of both PFTs are good indicators of long-term effects on the resilience of these rangeland ecosystems.
Therefore, studying the patterns of vegetation change from predrought, during drought, to postdrought conditions in an uncontrolled experimental setting could contribute to an improved understanding of the prolonged effects of drought combined with heavy grazing. Such observational approaches are even more informative when comparing results with those from experiments and model simulations.
4 Experimental Approach
4.1 Background: Experimental Approaches to Study Combined Effects of Drought and Grazing
Field experiments offer valuable opportunities to study ecological processes and the impact of climatic or land-use factors under controlled conditions (Table 16.1). Many studies have either evaluated grazing (Díaz et al. 2007; Linstädter et al. 2014) or drought (Cherwin and Knapp 2012; Tielbörger et al. 2014) as drivers for functional changes. However, field experiments focusing on combined and potentially interactive effects of rangeland management and centennial-scale drought on South Africa’s semiarid rangeland vegetation are lacking. Hence, “DroughtAct” has been specifically designed to address this research gap. The experiment also aimed at comparing the effects of drought events of different lengths (two versus six years), and to evaluate suitable rangeland management options during and after a drought. DroughtAct also contributes to the large, international and coordinated drought experiment DroughtNet (https://drought-net.colostate.edu/) which aims to assess ecosystem responses to centennial-scale drought.
4.2 Experimental Setup
The DroughtAct experiment started with a pretreatment year in the rainy season 2013/14, followed by six treatment years. It was implemented on a grazing camp of the Syferkuil experimental farm (Fig. 16.2) belonging to the University of Limpopo and was maintained as a collaborative effort of researchers from South Africa and Germany (Munjonji et al. 2020). The experiment has a full-factorial design with four-block repetitions of eight treatment plots. At the core of the experiment are four treatment combinations that were maintained for the whole duration of the experiment (six years): Ambient rainfall and grazing (D−G+, control), ambient rainfall and resting (D−G−, Fig. 16.4), drought and grazing (D+ G+, Fig. 16.4) and drought and resting (D+ G−). To directly compare the effects of a 6-year drought and a 2-year drought, we terminated the experimental drought on four plots per block after 2 years. We observed rangeland characteristics on plots with a drought history (H+) or without (H−), both under grazed and rested conditions.

Two treatments of the “DroughtAct” experiment. The left picture shows a plot under ambient rainfall and resting (D−G−) fenced with strain wire to prevent cattle grazing and chicken wire to also restrict entry of smaller mammals. The right picture shows a drought treatment in combination with grazing (D+ G+). The plot is covered by a rainout shelter and contains three movable cages to temporarily exclude grazing. Pictures by K. Behn
For the drought treatments, passive rainout-shelters were used (Yahdjian and Sala 2002). The shelters had a size of 6 x 6 m and consisted of metal constructions with transparent polycarbonate roof sheets. Sheets covered two-thirds of the area and thus reduced ambient rainfall by ca. 66%. We chose this level of reduction to reflect the site-specific rainfall history by turning a year with average rainfall into a year of centennial-scale drought (Knapp et al. 2017), following the guidelines of DroughtNet (https://drought-net.colostate.edu/). The perimeters of drought plots were trenched with plastic foil down to 1 m depth to inhibit lateral water flow that would counteract rainfall reduction (Mudongo et al. 2020). The camp was subject to moderate rotational grazing with cattle. Rested plots were fenced with strain wire to prevent grazing of cattle and with chicken wire to restrict entry of smaller mammals. We extensively checked for experimental artifacts and found that rainout shelters did not significantly alter light interception and microclimate (Mudongo et al., unpublished). No evidence of changed grazing preferences resulting from the rainout shelters was observed. With a height of at least 2 m, cattle could easily walk and graze below the roofs. The transparency of the polycarbonate shields did not provide enough shade to make the shelters a preferred resting place.
4.3 Data Collection
We assessed treatment effects on vegetation structure and composition, and on vegetation-mediated ecosystem functions and services. Here, we only report data collection for aboveground net primary production (ANPP) and functional vegetation composition, i.e., proxies that were considered in the observational study (see Sect. 16.3). At the time of peak standing biomass, we estimated ANPP of each plot. The sampling procedure differed between grazed and fenced plots (see Munjonji et al. 2020 for details). In brief, we harvested biomass on three 1 m2 harvesting quadrats on grazed plots (G+), where grazing was excluded for a given vegetation period with the aid of moveable cages (Fig. 16.4). In adjacent grazed quadrats, we assessed residual biomass with a nondestructive approach. In nongrazed plots (G−), the nondestructive approach was used, and in some years, biomass was additionally harvested in three smaller squares with a size of 0.25 m2. On each square plot, we recorded all occurring species, visually assessed their cover and measured their average height. We assigned each species to a PFT with the same a-priori approach used in the observational study. We distinguished three PFTs: perennial grasses, annual grasses and forbs. Facultative perennials and biennials were classified as perennials, following Linstädter et al. (2014).
4.4 Data Analysis
For all squares that were sampled using the nondestructive approach, we used data on species’ cover and height to estimate their relative contribution to ANPP. To this end, we applied a biomass–biovolume calibration developed for the experiment. To assess the drought and postdrought impacts, we calculated response ratios (Mackie et al. 2019) by dividing the mean value of the leaf biomass in all sampling quadrats belonging to the (post)drought treatments by the respective values of the control treatment. We put response ratios in relation to the experimental reduction of precipitation (Behn et al. 2022). The significance of the (post)drought effect was tested using ANOVA and TukeyHSD post-hoc test. For the PFTs, we calculated their relative contribution to the overall ANPP for each treatment and year.
4.5 Key Results
Our analyses of ANPP in predrought years, within-drought years and postdrought years across grazed and rested plots, and across plots with different drought duration showed marked differences in drought resistance and resilience (Behn et al. 2022). Similar as for the natural drought captured in the observational study (Sect. 16.3), the two-year experimental drought had significant negative effects on ANPP (Fig. 16.3). Compared to nondrought conditions, ANPP was on average reduced by 30% under grazed conditions in the second year of drought. Under rested conditions, the effect was even stronger with up to 50% reduction.
We found a fast recovery after the 2-year drought particularly under grazed conditions (Fig. 16.3), where already the first postdrought year did not show a significant deviation from the control. Under rested conditions, postdrought ecosystem performance remained on average below control level, even though differences were not significant from the second postdrought year on. However, the 6-year drought had devastating effects on ecosystem performance with an ANPP reduction of up to 80% and thus even exceeding the 66% reduction of precipitation. Grazing had an ambivalent role because its impacts were beneficial in the initial two years but became detrimental under ongoing drought.
Regarding the relative abundance of PFTs, there were notable differences concerning the drought duration. Perennial grasses tended to increase their relative abundance during the first years of drought and in the recovery phase after short drought at the expense of annual grasses (Fig. 16.3). With prolonged drought however, perennial grasses showed a strong decrease in relative abundance while forbs increased. Thus, species such as Chamaecrista mimosoides (L.) Greene and Monsonia angustifolia E.Mey. ex A.Rich. could be identified as relative winners of prolonged droughts. In absolute numbers, however, there were no winners when looking at biomass production per area.
4.6 Recommendations Derived from the Results and Outlook
Our results give valuable indications on both research and rangeland management practices in the face of drought. They stress the importance of a detailed understanding of the effects. Aridity and grazing management both altered ANPP and caused changes in vegetation structure and species composition. In combination, these effects impacted forage quantity, quality and palatability. Therefore, the changes caused by drought and grazing management can serve as warning signals for degradation. Management needs to be adapted and consider the ambivalent role that grazing and resting can have under different drought durations and intensities.
The DroughtAct experiment in Limpopo, South Africa, was the blueprint for a similar experiment in the Waterberg region of Namibia (Namtip project, https://www.namtip.uni-bonn.de/). The Tippex experiment of Namtip further improved the concept of DroughtAct and added a more detailed sampling approach, including the prominent role of the soil seed bank and different grazing intensities. With climate change, decreasing rainfall and changing rainfall patterns, drought experiments are crucial to understand the effects on vegetation to predict and mitigate degradation. Therefore, experiments like DroughtAct are essential due to their direct results, the data supply for vegetation modeling and to help scientists implement and improve drought experiments across the globe.
5 Modeling Approach
5.1 Background: Modeling Savanna Rangelands
Models represent properties, processes and functions of real-world systems in quantitative ways to improve system understanding. Thereby, they represent the current system understanding, but do not account for the full complexity of an ecosystem. Models are usually developed for specific research questions and only represent mechanisms considered relevant for those questions. As system understanding advances, models evolve through the inclusion of new knowledge, data and processes. Their main advantage is the applicability for a wide range of scenarios, such as past or future climate change scenarios, management scenarios, or factor combinations not considered in experiments (Table 16.1).
For savanna rangelands, several modeling approaches have been applied, including heuristic differential equation models (Baudena et al. 2010; Scheiter and Higgins 2007; van Langevelde et al. 2003), agent-based models (Fust and Schlecht 2018; Kuckertz et al. 2011) and process-based dynamic vegetation models (DVMs) (Pfeiffer et al. 2019; Scheiter and Higgins 2009). Within SALLnet, we improved and applied the individual-based dynamic vegetation models aDGVM (Scheiter and Higgins 2009) and aDGVM2 (Scheiter et al. 2013). Both models were originally developed to simulate grass-tree dynamics in savannas, but they differ in their representation of plant diversity. While aDGVM simulates dynamics of four PFTs (forest and savanna trees, C3 and C4 grasses), aDGVM2 simulates community assembly processes to create plant communities adapted to biotic and abiotic drivers.
5.2 Improving aDGVM and aDGVM2
We identified and resolved several limitations of aDGVM and aDGVM2. First, the herbaceous layer is commonly poorly represented in DVMs (Pfeiffer et al. 2019). While DVMs typically represent several woody PFTs, they aggregate grasses and forbs in C3 herbaceous and C4 herbaceous PFTs. However, when studying grazing impacts on productivity, diversity and forage quality in savanna rangelands, a representation of different grass PFTs and forbs is essential. Therefore, we included annual and perennial grass types into aDGVM2 by adjusting reproduction, carbon allocation and mortality (Pfeiffer et al. 2019). Annual grasses in aDGVM2 preferentially allocate carbon to rapid leaf growth and high seed production and die after one growing season. Perennial grasses preferentially invest carbon into root and storage compartments to enhance survival. Forbs have not yet been included in aDGVM2.
Second, in previous model versions, land-use activities such as grazing and fuelwood harvesting were poorly represented. Therefore, we included a cattle grazing model into aDGVM2 (Pfeiffer et al. 2019). In this model, cattle owners prescribe the grazing regime. Animals graze selectively with a preference for grass patches with high quantities of living leaf biomass, low ratios of dead-to-live grass biomass and high palatability. Palatability is assumed to increase with specific leaf area (SLA) and leaf nitrogen content. Thereby, grazing directly impacts the abundance of annual and perennial grasses. We further coupled aDGVM with routines to simulate grazing and fuelwood harvesting (Scheiter et al. 2019). Grazing nonselectively removes a prescribed amount of grass biomass, and fuelwood harvesting removes prescribed amounts of woody biomass. Harvesting was related to tree stem diameter, preferring trees with a stem diameter between 5 and 10 cm (Twine and Holdo 2016).
Third, aDGVM and aDGVM2 did not consider shrubs. We included shrubs into aDGVM2 (Gaillard et al. 2018) and assumed that differences between trees and shrubs are related to trade-offs between water availability, light availability and height. Trees generally invest more into height growth, which is an advantage in dense and light-limited environments. Contrastingly, shrubs with a multistemmed architecture have a higher sapwood area and improved water transport capacity. Having multiple stems entails lower height growth and a competitive disadvantage in light-limited ecosystems (Gaillard et al. 2018).
5.3 Key Results
When considering natural vegetation dynamics without land use, aDGVM results showed increases in woody biomass and biome transitions to wood-dominated vegetation states until 2099 in response to the representative concentration pathway (RCP) scenarios RCP4.5 and RCP8.5 (Martens et al. 2021; Scheiter et al. 2018). RCP4.5 is a modest-high climate mitigation scenario where carbon emissions peak toward the middle of the century, whereas RCP8.5 is a low climate mitigation scenario with high carbon emissions and energy consumption (van Vuuren et al. 2011). Biomass increases resulted from CO2 fertilization effects on tree growth. Grassland and savanna areas were most susceptible to biome transitions (Martens et al. 2021; Scheiter et al. 2018; Chap. 14). Simulations showed that vegetation dynamics lag behind environmental forcing and that observed vegetation states can deviate from those expected under given environmental conditions (Pfeiffer et al. 2020; Scheiter et al. 2020). Lagged responses result from the different velocities at which processes such as ecophysiology, population dynamics or succession operate.
Increasing grazing pressure under varying annual rainfall regimes reduced grass productivity and grass biomass at study sites in South Africa and altered the composition of the grass layer (Pfeiffer et al. 2019). Specifically, community composition shifted toward a higher abundance of annuals with increasing grazing intensity. Annual grasses became dominant once the grazing demand exceeded a critical value between 1.5 and 3 LU/ha for a rainfall gradient between 253 and 926 mm MAP, assuming a daily dry matter demand of 12.5 kg/LU/day. Such changes occurred when grazing intensity exceeded the carrying capacity of perennial grasses, i.e., when regrowth of perennial grass biomass was insufficient to cover animal demand. Consequently, the grass type preferred by grazers switched from perennials to annuals. Resting periods without animals were necessary for biomass recovery and regeneration of perennial grasses (Pfeiffer et al. 2019). Recovery periods were site-specific and up to 8 and 17 years at arid and humid sites respectively. Recovery was faster at more arid sites because these sites had a lower biomass without grazers that they had to reach, and because annual grasses that are more abundant at the arid sites recover faster than perennial grasses that are more abundant at the humid sites. Drought impacts have not been considered explicitly in these simulations, but see Sect. 16.6.1 for aDGVM2 simulations of the DroughtAct experiment.
We used optimization techniques to identify grazing and fuelwood harvesting intensities that were well-sustained by vegetation at Bushbuckridge, South Africa, and maximized the economic value of the land-use system (Scheiter et al. 2019). The economic value included, for example, milk, meat, dung and the cultural status conveyed by owning cattle. Simulations indicated that the optimal animal number was only 0.076 LU/ha whereas observed animal numbers are, for example, up to 0.75 LU/ha on the farm containing the DroughtAct experiment, 0.88 LU/ha in communal grazing lands north of Acornhoek (Parsons et al. 1997) and > 1 LU/h at a site used for the observations (Sect. 16.3). Similarly, optimal fuelwood harvesting intensities were lower than observed intensities.
5.4 Recommendations Derived from the Results and Outlook
Simulation results indicate that in the absence of land use and herbivory, grasslands and savannas are highly susceptible to woody encroachment and transition to woody vegetation states due to climate change and elevated CO2 (Martens et al. 2021; Scheiter et al. 2018). Land use (fuelwood harvesting and grazing) can alter the velocity of such transitions (Scheiter and Savadogo 2016). Vegetation changes are considerably slower than changes in environmental drivers. Therefore, vegetation will continue changing even if humanity reduces greenhouse gas emissions and manages to stabilize the climate system (Pfeiffer et al. 2020; Scheiter et al. 2020). Such delayed responses of vegetation need consideration when developing management plans for savanna rangelands.
Intense grazing strongly decreases grass biomass and productivity and shifts grass communities toward a higher abundance of annual grasses (Pfeiffer et al. 2019). According to the simulation results, vegetation is resilient and recovery is possible during resting periods without grazing. To ensure full recovery of heavily grazed vegetation (more than ca. 2.5 LU/ha) to the productivity levels expected in the absence of grazing, resting times of 5–17 years were simulated, with shorter resting times (5–8 years) for more arid sites and longer resting times (14–17 years) for more humid sites. After low-intensity grazing (less than ca. 1.2 LU/ha), resting times of less than 6 years were sufficient. Yet, to ensure sustained forage for animals, full recovery might not be necessary and sorter resting times are sufficient. Downsizing of herds, provision of additional fodder, as well as rotational herding strategies that include rangeland and cropland are possible methods to achieve resting (Pfeiffer et al. 2019, 2022).
6 Integrating Observations, Experiments and Modeling
Observation, experimentation and modeling ideally complement each other, and data and information are shared between these approaches (Fig. 16.5). Field-based observations provide insights into system functionality, identify relevant system components and are necessary for a qualitative system conceptualization. The resulting conceptual system understanding is the basis for hypothesis formulation that can be evaluated in specifically tailored experiments. The strength of observations and experiments (Sects. 16.3 and 16.4) is that they directly measure system dynamics and responses to natural environmental conditions or different experimental treatments (Table 16.1). Results from experiments provide insights allowing focus re-evaluation for further observations. They also yield system-specific quantitative data for model parameterization, calibration and benchmarking, but are often resource-limited regarding treatment numbers, factor combinations, study sites, replicates and duration. Quantitative experimental data are the basis for the deduction of system dependencies that allow the development of quantitative process models. The steps from observation to experimentation and model development signify an increasing degree of system generalization and abstraction, with a narrowing focus on processes and components deemed most relevant for the targeted questions. Process-based models (Sect. 16.5) enable scenario testing, spatiotemporal extrapolation and testing of system sensitivities and allow exploration of possible trajectories for future climate change, management scenarios or regional upscaling. Knowledge gained from models can then inform both experimentation and observation to re-evaluate the focus of further research, help generate new hypotheses and improve system understanding.

Linking observations, experiments and modeling—how different research approaches inform each other
6.1 Applying aDGVM2 to DroughtAct
Observations and experiments showed clear responses of vegetation to drought. Therefore, we replicated the DroughtAct experiment in aDGVM2 simulations (1) to test if aDGVM2 can simulate observed vegetation responses to drought and (2) to test if vegetation can recover between the end of the 6-year experimental drought in 2020 and 2030. We simulated all factor combinations described in Sect. 16.4 but used 40 instead of 4 replicates per treatment and continued simulations until 2030. Meteorological data were not available for the DroughtAct site and for future conditions to allow postdrought simulations. Data for a neighboring meteorological station had data gaps (https://www.weathersa.co.za/) and were therefore unsuitable to conduct simulations. Hence, we selected climate forcing data from climate model simulations to find the model and RCP scenario that agreed best with precipitation from station data and the EWEMBI climate product for 2008 to 2017 (Lange 2019). We found the best fit for precipitation from the CMIP5 simulations conducted with the IPSL-CM5A-LR model for RCP8.5 (Dufresne et al. 2013) combined with a transformation. Specifically, we shifted the simulated climate data by one year to the future and multiplied daily precipitation with a factor of 0.73. For drought treatments, we reduced daily rainfall by 66% for a 2-year and 6-year period starting in the growing season 2014/2015 following DroughtAct. For grazing treatments, we assumed an average number of 30 LU on the 40 ha grazing camp that includes the DroughtAct experiment, with a per-capita daily demand of 12.5 kg dry matter. We simulated a rotational grazing system with animals present during four 30-day periods per year (i.e., 120 days per year) and resting times without animals between the grazing periods. Timing of presence and absence was equal in all years. Daily biomass removal per simulated hectare was determined by randomly assigning animals to each of the 40 ha. Simulations were repeated in the absence of grazing. However, differences between simulations with and without grazing were minor as drought impacts overrode grazing impacts. We therefore only present simulations with grazing in the following.
The first two years of drought led to a steep decline in mean annual plot-level leaf biomass compared to the control scenario (Figs. 16.3, 16.6a). Biomass was reduced by ca. 40% after the second year of drought. In the 6-year drought, biomass stabilized at values of approximately 40% below the control during the 3rd to 6th years of drought. Postdrought recovery was rapid during the first two years after drought before slowing down (Fig. 16.6a). Five years after the end of the 2-year and the 6-year drought, plot-level leaf biomass was approximately 5% and 8% below the control, respectively. In 2030, plot-level biomass was still approximately 3% and 6% below the control for the 2-year and 6-year drought, respectively. At grass patch-level, biomass response to drought was similar to plot-level response (Fig. 16.6b). However, at patch-level leaf biomass fully recovered to values simulated in the control. This result indicates that surviving or newly established grass patches were resilient to drought, whereas slow recruitment and recolonization of bare ground inhibited recovery at the plot level.

Impacts of a 2-year and a 6-year drought on simulated grass biomass and PFTs, and postdrought recovery relative to nondrought control scenario. Panels show (a) biomass at the site scale, (b) biomass at the grass patch scale and (c) fraction of annual grasses in the grass population. The yellow shading indicates the 2-year drought period, the yellow and red shading the 6-year drought period and the green shading the recovery period until 2030 after the termination of drought treatments. Letters above boxes indicate the significance of the difference in the mean based on the t-test between treatment and control (n: not significant; a: p < 0.05; b: p < 0.01; c: p < 0.001)
Plot-level biomass loss during drought was caused by increased mortality of perennial grasses and lower productivity of surviving grasses. While mortality was 1–3 times higher than control during the first two drought years, it was almost 20 times higher during the 3rd and 4th drought years. Remarkably, the higher mortality persisted for five years after drought treatments and then reached values simulated for the control. Mortality of perennial grasses during drought allowed annual grasses to colonize the study site and their fraction in the population was 1.5 times higher during the first two years and 2.5 times higher from the 3rd drought year onward (Fig. 16.6c). Their abundance decreased during the recovery period but was still 1.4 and 1.8 times higher in the 2-year and 6-year drought scenarios at the end of the simulation period.
Overall, simulations indicated resilience of all considered model variables. We found no tipping point behavior or transition into an alternative vegetation state that persisted after drought. However, the velocity of recovery differed between the 2-year and the 6-year drought and between considered variables. For instance, while patch-level grass biomass recovered within 2 or 3 years after the drought treatment, attributes of community composition did not fully recover during the simulation period. This indicates that resting or at least reduced grazing intensity becomes increasingly important with increasing drought duration.
6.2 What Did We Learn from Observations, Experiments and Modeling?
The three approaches applied in LLL and SALLnet (observation, experimentation and modeling) revealed clear and consistent responses of savanna rangeland vegetation to drought and grazing. The results included decreases in biomass and productivity, changes in the relative abundances of different PFTs and recovery after drought (Fig. 16.3). However, we found disagreement in some aspects. While productivity in the experiments was relatively resistant during the first two years of drought and showed accelerating decreases in the 6-year drought, modeled biomass and productivity decreased only during the first two drought years and afterward stabilized at a lower level. We explain this model behavior by changes in resource availability, competition and community assembly. In the aDGVM2, soil water dynamics are only simulated at the 1-ha plot-level using a simple multilayer soil water bucket model instead of simulation at patch or plant level. Therefore, reduced vegetation cover due to drought mortality improves the water availability of all surviving individuals by lowering the water demand at plot level. In contrast, soil water status in the experiments and observations varies spatially due to microscale heterogeneity of soil conditions and topography.
Moreover, the aDGVM2 simulated expansion of annual grasses under drought conditions, while field-based methods showed expansion of forbs (Sects. 16.3, 16.4). Forbs are currently not represented in aDGVM2, and the modeled expansion of annual grasses during drought indicates that they fill the ecological niche of forbs in simulations. We argue that forbs need to be added to vegetation models to improve understanding of drought and grazing impacts on savanna rangelands (Siebert and Dreber 2019).
Observations, experiments and modeling showed that vegetation attributes such as biomass or productivity can recover after short drought periods, indicating resilience (Fig. 16.3). Model results suggest that vegetation can also recover after the 6-year drought (Fig. 16.6), hinting at resilience after longer drought periods. However, the rate and duration of recovery differ between vegetation attributes. In particular, community-related attributes show considerable delays when recovering after grazing or drought, primarily due to delayed community assembly and recolonization. How these model results agree with observed vegetation dynamics remains open because results related to recovery from the 6-year drought experiments are not yet available. Regular monitoring of the field sites during the following years would provide valuable information on vegetation recovery and the predictive power of the aDGVM2.
Within LLL and SALLnet, observations, experiments and modeling independently have provided valuable insights into ecosystem resistance and resilience to drought and grazing. However, added value emerged when all three activities were combined and informed each other. DroughtAct experiments allowed in-depth investigation of specific aspects that have been identified in observations, for example, the role of forbs in mitigating drought effects. Experiments allowed direct control and manipulation of environmental drivers, for example, precipitation reduction via rainout shelters, soil moisture control via trenching of experimental plots, or regulation of soil nutrient levels via fertilizer application. In observational studies, such insights are only possible by space-for-time substitution (Blois et al. 2013) or if all treatment combinations are observable. Observational studies usually lack predrought data, as drought events are not planned or easily predictable. Such predrought data are available in controlled experiments.
Field-based methods informed model development. Comparing model results with observational data revealed key aspects not captured by aDGVM2 and led to stepwise model improvement. For example, such comparisons motivated the implementation of annual and perennial grasses as distinct PFTs (Pfeiffer et al. 2019), adjustments of perennial-grass mortality during drought, consideration of buffering effects due to storage reserves during the first drought year and adjustments of postdrought colonization of bare ground (this study, Behn et al. 2022). We identified the representation of water sharing among plant individuals as a model limitation that may lead to overestimates in drought resistance once the number of plant individuals has declined due to drought mortality. A refined soil water scheme that explicitly simulates plant water availability at grass patch level may be required to reproduce the progressive decline of biomass and productivity during the 6-year drought observed in the experiment. We also found lacking forbs as functional types in the model likely influences modeled community response to drought. Using knowledge on the function and ecology of forbs gained in observations and experiments is therefore essential for future model development.
In return, the model allowed prognostic extrapolation of postdrought vegetation recovery during the 2020s. Such model-based extrapolations contribute toward synergies between observations, experiments and models, and they can provide added value to the conclusions drawn from experimental results (Behn et al. 2022). Model results can generate new hypotheses for future experimental studies. For example, we used the aDGVM2 to simulate all factorial treatment combinations realized in the DroughtAct experiment. We could conduct further simulations for factorial combinations not considered in the experiment, test additional management options, rotational grazing systems and climate change scenarios, or investigate how repeated drought affects rangelands with drought history.
6.3 Recommendations for Decision-Makers
Based on evidence from three different approaches, we derived the following conclusions and recommendations:
-
1.
Grasslands and savanna rangelands are susceptible to climate change, particularly to drought, and overgrazing. Impacts include woody encroachment and transitions to woody vegetation states, or, if heavily utilized, to declining forage quality and availability, increased erosion, and transitions to degraded states. Contrastingly, dead biomass accumulation resulting from underutilization reduces rangeland productivity.
-
2.
Recovery of productivity, biomass and community composition is possible after drought and grazing, but related to drought duration, grazing intensity and precipitation. We found rangeland resilience to short drought or low grazing, and quick recovery after such disturbances. In contrast, long drought or intense grazing requires recovery periods of at least two years, particularly if plant community composition has changed.
-
3.
Resting times without grazing or very low grazing intensity are necessary to allow the recovery of perennial grasses. Resting times of 1–2 years with sufficient precipitation are a “window of opportunity of regeneration” (Linstädter et al. 2014), and ensure long-term forage quality for grazers. Resting times of more than two years may be required in grazing systems affected by longer droughts. Rotational grazing systems or forage supply during and after drought can be applied to manage resting times. The holistic management approach (Savory and Butterfield 2016) could provide a template for grazing and resting schemes.
-
4.
Forbs have high value to secure essential ecosystem functions and services within rangeland systems and need consideration in management.
-
5.
Capacity building is crucial to make results available to relevant stakeholders. It should include training courses on field methods or modeling, stakeholder workshops and contributions to regional or global initiatives such as the South African Risk and Vulnerability Atlas, IPBES or IPCC.
7 Outlook
By replicating the DroughtAct experiments with aDGVM2 and comparing model results to the experiment, we identified several opportunities for improving the aDGVM2 for rangelands. As highlighted in Sect. 16.6, improvements should focus on the soil water model. A fine-scale representation of the soil water status allows more precise simulations of plant water availability and drought response. Additionally, aDGVM2 should be improved to capture the diversity of vegetation. Future model development could include more detailed PFTs in the herbaceous layer, such as increasers or decreasers, grass types propagating by stolons or rhizomes, geophytes, xerophytes, succulents or perennial forbs. Adaptations to disturbances such as a bud bank, water- and carbon-storage organs or chemical defenses could be included (Archibald et al. 2019; Pausas et al. 2018). The representation of the woody layer could be improved by including protection against herbivory such as thorns, spines or a cage architecture (Charles-Dominique et al. 2017; Wigley et al. 2018).
In addition, modeling animal behavior and plant-animal interactions in more detail is desirable. This includes, for example, utilization of observed timing and duration of animal presence and absence at a study site instead of using a probabilistic approach. Model development could improve the representation of herbivore selection of particular plants, the feeding duration and amount of biomass removed per plant, and the average number of plants affected per day based on observations. Rangelands are often not exclusively utilized by domesticated animals but also by wild animals. Therefore, aDGVM2 could include different herbivore functional types (Hempson et al. 2015a) with distinctive forage preferences. Finally, aDGVM2 currently does not simulate population dynamics and movement of animals. These processes are particularly relevant when modeling wild animals. Reproduction or mortality of animals could be included and related to their nutritional status using previous work (Pachzelt et al. 2013). DVMs could be coupled with agent-based models to simulate animal movement (Clemen et al. 2021; Fust and Schlecht 2018; Tang and Bennett 2010) and to simulate behavior and decision-making of pastoralists. For instance, the DECUMA household model has previously been coupled to the SAVANNA ecosystem model to study drought impacts on socioeconomic systems in Kenya (Boone et al. 2011; Boone and Galvin 2014).
Modeling drought impacts requires consideration of vegetation resistance to drought, recovery and drought-induced mortality. Modeling plant mortality is still challenging and a significant source of uncertainty in vegetation models (Hartmann et al. 2018). Ecophysiological processes, plant traits, or plant trait syndromes affect drought resistance and mortality (Sankaran 2019), and detailed knowledge of these aspects is required. A cascade of processes describes the establishment niche after disturbance (Holt 2009), including recovery of individual plants, colonization of bare ground, establishment and succession, and drives postdrought recovery. Different simulated recovery rates of grass biomass at plot- and patch-level and changes in the fractions of annual and perennial grasses after drought indicate that such processes are, to a certain degree, represented by aDGVM2. However, DVMs typically operate at large spatial scales and thus ignore small-scale processes such as seed dispersal, dynamics of the seed bank and colonization. Drought conditions can influence seed dynamics and decouple species represented in the seed bank from species in the aboveground community (Basto et al. 2018), and seed mortality increased under drought in a watering experiment (Harrison and LaForgia 2019). While those studies were not conducted in savanna rangeland systems, observations and experiments such as DroughtAct can provide information on recovery and the establishment niche. This information is valuable for parameterizing recovery and successional effects in models. In addition to increased drought risk, vegetation will experience higher atmospheric CO2 under future conditions. Elevated CO2 may fertilize plants, particularly growth of woody vegetation, and thereby modify competitive interactions between grasses, forbs and woody vegetation in savanna rangelands (Midgley and Bond 2015). Elevated CO2 might mitigate drought impacts via increased water use efficiency and enhance recovery after drought.
Our analyses focused on grazing and drought effects at a single study site. We argue that more holistic study approaches at the landscape or regional level are required to account for the multifunctionality of southern African landscapes (Rötter et al. 2021). Rural areas in southern Africa are typically used for cattle grazing, fuelwood collection and crop production. Cattle connect rangeland and cropland when feeding on crop residues or in rotational grazing systems (Pfeiffer et al. 2022). On larger scales, landscapes can be mosaics of rural areas, commercial farms, plantations, conservation areas and game reserves. Management policies need to consider these diverse land-use forms and the socioeconomic interests of stakeholders utilizing the natural resources. Combining rangeland models, crop models, economic models and agent-based models that simulate animal behavior and the decision-making of stakeholders can provide a valuable tool for decision support and the development of regional-level management policies.
The high environmental variability in savanna rangelands and the increasing likelihood of droughts make it challenging to derive management policies that ensure the sustained availability of invaluable ecosystem functions and services. Droughts can amplify the effects of grazing on vegetation and increase the risk that vegetation shifts into degraded states with low forage quantity and quality. More model simulations are required to better assess the impacts of repeated drought and different grazing strategies. Such simulations can provide insights on minimum or optimal resting times between grazing treatments or optimal resting times after drought periods of different length and therefore account for the fact that resting times of several years as assumed in our simulations might not be applicable in reality. Combining observations, experiments and models can help develop early warning signals that indicate the risk of degradation. Such indicators can then inform the developers of management intervention strategies to keep the socioecological system in a safe operating space. A model well parameterized and tested with observational and experimental data can systematically simulate a large ensemble of different management and drought scenarios. These model results can then help estimate the risk of undesirable vegetation change and degradation under different scenarios.
8 Conclusions
In the LLL and SALLnet projects, we combined observations, experiments and models to understand the response of rangeland vegetation to drought and grazing. The approaches consistently showed that drought causes substantial losses in biomass and productivity of grasses and shifts in community composition from the dominance of perennials to annuals or forbs. However, we did not find tipping point behavior and irreversible transitions to alternative vegetation states that persisted after drought. Rangelands were able to recover from drought and grazing impacts, in model results even after a 6-year drought period. However, in the model results, some vegetation attributes did not fully recover to the vegetation state simulated under no-drought conditions until 2030. We highlight the following conclusions:
-
1.
Resting times are necessary to allow vegetation recovery after grazing and drought and to ensure continued provision of essential ecosystem services to people. Recovery rates and resting times depend on the length and intensity of grazing and drought.
-
2.
Combining observations, experiments and models is essential to understand rangeland ecology and to forecast impacts of future climate change and land-use on rangeland vegetation. Using such an integrated approach, we were able to derive policy-relevant recommendations based on evidence from three approaches (see Sect. 16.6).
-
3.
Further research is required, in particular, to understand drought impacts on the complex interactions between woody vegetation, grasses and forbs, and to understand how plant traits influence resilience to grazing and drought.
-
4.
Based on our methods, we can develop early warning signals that indicate potential undesired vegetation shifts and the necessity of management intervention.
References
Archibald S, Hempson GP, Lehmann C (2019) A unified framework for plant life-history strategies shaped by fire and herbivory. New Phytol 224:1490–1503. https://doi.org/10.1111/nph.15986
Arena G, van der Merwe H, Todd SW, Pauw MJ, Milton SJ, Dean WRJ, Henschel JR (2018) Reflections, applications and future directions of long-term ecological research at Tierberg. Afr J Range Forage Sci 35:257–265. https://doi.org/10.2989/10220119.2018.1513072
Basto S, Thompson K, Grime JP, Fridley JD, Calhim S, Askew AP, Rees M (2018) Severe effects of long-term drought on calcareous grassland seed banks. Npj Clim Atmospheric Sci 1:1–7. https://doi.org/10.1038/s41612-017-0007-3
Baudena M, D’Andrea F, Provenzale A (2010) An idealized model for tree-grass coexistence in savannas: the role of life stage structure and fire disturbances. J Ecol 98:74–80
Behn K, Pfeiffer M, Mokoka V, Mudongo, E, Ruppert J, Scheiter S, Ayisi K, Linstädter A(2022): Assessing the relevance of drought duration on dryland rangelands: an experimental and modelling study. Tropentag 2022, Prague, Czech Republic, 14–16 September 2022 (Poster).https://doi.org/10.13140/RG.2.2.24307.09768
Blois JL, Williams JW, Fitzpatrick MC, Jackson ST, Ferrier S (2013) Space can substitute for time in predicting climate-change effects on biodiversity. Proc Natl Acad Sci 110:9374–9379. https://doi.org/10.1073/pnas.1220228110
Boone RB, Galvin KA (2014) Simulation as an approach to social-ecological integration, with an emphasis on agent-based modeling. In: Manfredo MJ, Vaske JJ, Rechkemmer A, Duke EA (eds) Understanding society and natural resources: forging new strands of integration across the social sciences. Springer Netherlands, Dordrecht, pp 179–202. https://doi.org/10.1007/978-94-017-8959-2_9
Boone R, Galvin K, Burn Silver S, Thornton P, Ojima D, Jawson J (2011) Using coupled simulation models to link pastoral decision making and ecosystem services. Ecol Soc 16. https://doi.org/10.5751/ES-04035-160206
Buisson E, Le Stradic S, Silveira FAO, Durigan G, Overbeck GE, Fidelis A, Fernandes GW, Bond WJ, Hermann J-M, Mahy G, Alvarado ST, Zaloumis NP, Veldman JW (2019) Resilience and restoration of tropical and subtropical grasslands, savannas, and grassy woodlands. Biol Rev 94:590–609. https://doi.org/10.1111/brv.12470
Buitenwerf R, Swemmer AM, Peel MJS (2011) Long-term dynamics of herbaceous vegetation structure and composition in two African savanna reserves. J Appl Ecol 48:238–246. https://doi.org/10.1111/j.1365-2664.2010.01895.x
Camp KGT, Hardy MB (1999) Veld condition assessment. In: Hardy MB, Hurt CR, Camp KGT, Smith JMB, Tainton NM (eds) Veld in KwaZulu-Natal. KwaZulu-Natal. Department of Agriculture and Environmental Affairs, Cedara, pp 18–31
Charles-Dominique T, Barczi J-F, Roux EL, Chamaille-Jammes S, McArthur C (2017) The architectural design of trees protects them against large herbivores. Funct Ecol 31:1710–1717. https://doi.org/10.1111/1365-2435.12876
Cherwin K, Knapp A (2012) Unexpected patterns of sensitivity to drought in three semi-arid grasslands. Oecologia 169:845–852. https://doi.org/10.1007/s00442-011-2235-2
Clemen T, Lenfers UA, Dybulla J, Ferreira SM, Kiker GA, Martens C, Scheiter S (2021) A cross-scale modeling framework for decision support on elephant management in Kruger National Park, South Africa. Ecol Inform 62:101266. https://doi.org/10.1016/j.ecoinf.2021.101266
Díaz S, Lavorel S, McINTYRE S, Falczuk V, Casanoves F, Milchunas DG, Skarpe C, Rusch G, Sternberg M, Noy-Meir I, Landsberg J, Zhang W, Clark H, Campbell BD (2007) Plant trait responses to grazing – a global synthesis. Glob Chang Biol 13:313–341. https://doi.org/10.1111/j.1365-2486.2006.01288.x
Du Toit JT, Cumming DHM (1999) Functional significance of ungulate diversity in African savannas and the ecological implications of the spread of pastoralism. Biodivers Conserv 8:1643–1661. https://doi.org/10.1023/A:1008959721342
Dufresne J-L, Foujols M-A, Denvil S, Caubel A, Marti O, Aumont O, Balkanski Y, Bekki S, Bellenger H, Benshila R, Bony S, Bopp L, Braconnot P, Brockmann P, Cadule P, Cheruy F, Codron F, Cozic A, Cugnet D, de Noblet N, Duvel J-P, Ethé C, Fairhead L, Fichefet T, Flavoni S, Friedlingstein P, Grandpeix J-Y, Guez L, Guilyardi E, Hauglustaine D, Hourdin F, Idelkadi A, Ghattas J, Joussaume S, Kageyama M, Krinner G, Labetoulle S, Lahellec A, Lefebvre M-P, Lefevre F, Levy C, Li ZX, Lloyd J, Lott F, Madec G, Mancip M, Marchand M, Masson S, Meurdesoif Y, Mignot J, Musat I, Parouty S, Polcher J, Rio C, Schulz M, Swingedouw D, Szopa S, Talandier C, Terray P, Viovy N, Vuichard N (2013) Climate change projections using the IPSL-CM5 earth system model: from CMIP3 to CMIP5. Clim Dyn 40:2123–2165. https://doi.org/10.1007/s00382-012-1636-1
Ellis SM, Steyn HS (2003) Practical significance (effect sizes) versus or in combination with statistical significance (p-values): research note. Manag Dyn J South Afr Inst Manag Sci 12:51–53. https://doi.org/10.10520/EJC69666
Ferner J, Schmidtlein S, Guuroh RT, Lopatin J, Linstädter A (2018) Disentangling effects of climate and land-use change on West African drylands’ forage supply. Glob Environ Chang 53:24–38. https://doi.org/10.1016/j.gloenvcha.2018.08.007
Fust P, Schlecht E (2018) Integrating spatio-temporal variation in resource availability and herbivore movements into rangeland management: RaMDry—an agent-based model on livestock feeding ecology in a dynamic, heterogeneous, semi-arid environment. Ecol Model 369:13–41. https://doi.org/10.1016/j.ecolmodel.2017.10.017
Fynn RWS, O’Connor TG (2000) Effect of stocking rate and rainfall on rangeland dynamics and cattle performance in a semi-arid savanna, South Africa. J Appl Ecol 37:491–507. https://doi.org/10.1046/j.1365-2664.2000.00513.x
Gaillard C, Langan L, Pfeiffer M, Kumar D, Martens C, Higgins SI, Scheiter S (2018) African shrub distribution emerges via height - sapwood conductivity trade-off. J Biogeogr 45:2815–2826
Gbetibouo GA (2009) Understanding farmers perceptions and adaptations to climate change and variability: the case of the Limpopo Basin farmers South Africa. IFPRI Discussion Paper 849. IFPRI, Washington, DC
Harrison S, LaForgia M (2019) Seedling traits predict drought-induced mortality linked to diversity loss. Proc Natl Acad Sci 116:5576–5581. https://doi.org/10.1073/pnas.1818543116
Hartmann H, Moura CF, Anderegg WRL, Ruehr NK, Salmon Y, Allen CD, Arndt SK, Breshears DD, Davi H, Galbraith D, Ruthrof KX, Wunder J, Adams HD, Bloemen J, Cailleret M, Cobb R, Gessler A, Grams TEE, Jansen S, Kautz M, Lloret F, O’Brien M (2018) Research frontiers for improving our understanding of drought-induced tree and forest mortality. New Phytol 218:15–28. https://doi.org/10.1111/nph.15048
Hempson GP, Archibald S, Bond WJ (2015a) A continent-wide assessment of the form and intensity of large mammal herbivory in Africa, vol 350. Science, p 1056
Hempson GP, Archibald S, Bond WJ, Ellis RP, Grant CC, Kruger FJ, Kruger LM, Moxley C, Owen-Smith N, Peel MJS, Smit IPJ, Vickers KJ (2015b) Ecology of grazing lawns in Africa. Biol Rev 90:979–994. https://doi.org/10.1111/brv.12145
Higgins SI, Scheiter S (2012) Atmospheric CO2 forces abrupt vegetation shifts locally, but not globally. Nature 488:209–212
Holt RD (2009) Bringing the Hutchinsonian niche into the 21st century: ecological and evolutionary perspectives. Proc Natl Acad Sci 106:19659–19665. https://doi.org/10.1073/pnas.0905137106
IPCC (2021) Climate change 2021: the physical science basis. Contribution of working group I to the sixth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Kalvelage L, Revilla Diez J, Bollig M (2020) How much remains? Local value capture from tourism in Zambezi, Namibia. Tour Geogr 1:1–22. https://doi.org/10.1080/14616688.2020.1786154
Klem J (2018) Drought responses of forb and grass communities in communal and protected rangelands. MSc dissertation, North-West University, South Africa
Knapp AK, Ciais P, Smith MD (2017) Reconciling inconsistencies in precipitation–productivity relationships: implications for climate change. New Phytol 214:41–47. https://doi.org/10.1111/nph.14381
Kuckertz P, Ullrich O, Linstädter A, Speckenmeyer E (2011) Agent-based modeling and simulation of a Pastoralnomadic land use system. Simul Notes Eur 21:147–152
Lange, S., 2019. EartH2Observe, WFDEI and ERA-interim data merged and bias-corrected for ISIMIP (EWEMBI). V. 1.1. GFZ Data Services
Lehohla P (2012) Census 2011 Census in brief. Statistics South Africa
Linstädter A, Schellberg J, Brüser K, García CAM, Oomen RJ, du Preez CC, Ruppert JC, Ewert F (2014) Are there consistent grazing indicators in drylands? Testing plant functional types of various complexity in South Africa’s grassland and savanna biomes. PLoS One 9:e104672. https://doi.org/10.1371/journal.pone.0104672
Mackie KA, Zeiter M, Bloor JMG, Stampfli A (2019) Plant functional groups mediate drought resistance and recovery in a multisite grassland experiment. J Ecol 107:937–949. https://doi.org/10.1111/1365-2745.13102
Maluleke, M., Malungani, T., Steenkamp, K. (Eds.), 2016. Limpopo environment outlook report 2016. Limpopo Provincial Government, Department of Economic Development, Environment and Tourism
Marchant R (2010) Understanding complexity in savannas: climate, biodiversity and people. Curr Opin Environ Sustain 2:101–108. https://doi.org/10.1016/j.cosust.2010.03.001
Martens C, Hickler T, Davis-Reddy C, Engelbrecht F, Higgins SI, von Maltitz GP, Midgley GF, Pfeiffer M, Scheiter S (2021) Large uncertainties in future biome changes in Africa call for flexible climate adaptation strategies. Glob Chang Biol 27:340–358. https://doi.org/10.1111/gcb.15390
Matsika R, Erasmus BFN, Twine WC (2013) Double jeopardy: the dichotomy of fuelwood use in rural South Africa. Energy Policy 52:716–725. https://doi.org/10.1016/j.enpol.2012.10.030
Midgley GF, Bond WJ (2015) Future of African terrestrial biodiversity and ecosystems under anthropogenic climate change. Nat Clim Chang 5:823–829
Minnaar, C., 2020. Drought effects on the herbaceous community structure of transformed Mopaneveld. MSc dissertation, North-West University
Munjonji L, Ayisi KK, Mudongo EI, Mafeo TP, Behn K, Mokoka MV, Linstädter A (2020) Disentangling drought and grazing effects on soil carbon stocks and CO2 fluxes in a semi-arid African savanna. Front Environ Sci 8:207. https://doi.org/10.3389/fenvs.2020.590665
New M, Lister D, Hulme M, Makin I (2002) A high-resolution data set of surface climate over global land areas. Clim Res 21:1–25
O’Connor TG (1991) Patch colonisation in a savanna grassland. J Veg Sci 2:245–254. https://doi.org/10.2307/3235957
O’Connor TG (1995) Transformation of a savanna grassland by drought and grazing. Afr J Range Forage Sci 12:53–60. https://doi.org/10.1080/10220119.1995.9647864
Oomen RJ, Ewert F, Snyman HA (2016) Modelling rangeland productivity in response to degradation in a semi-arid climate. Ecol Model 322:54–70. https://doi.org/10.1016/j.ecolmodel.2015.11.001
Pachzelt A, Rammig A, Higgins SI, Hickler T (2013) Coupling a physiological grazer population model with a generalized model for vegetation dynamics. Ecol Model 263:92–102. https://doi.org/10.1016/j.ecolmodel.2013.04.025
Parsons DAB, Shackleton CM, Scholes RJ (1997) Changes in herbaceous layer condition under contrasting land use systems in the semi-arid lowveld, South Africa. J Arid Environ 37:319–329. https://doi.org/10.1006/jare.1997.0283
Pausas JG, Bond WJ (2020) Alternative biome states in terrestrial ecosystems. Trends Plant Sci 25:250–263. https://doi.org/10.1016/j.tplants.2019.11.003
Pausas JG, Lamont BB, Paula S, Appezzato-da-Glória B, Fidelis A (2018) Unearthing belowground bud banks in fire-prone ecosystems. New Phytol 217:1435–1448. https://doi.org/10.1111/nph.14982
Pfeiffer M, Langan L, Linstädter A, Martens C, Gaillard C, Ruppert JC, Higgins SI, Mudongo EI, Scheiter S (2019) Grazing and aridity reduce perennial grass abundance in semi-arid rangelands – insights from a trait-based dynamic vegetation model. Ecol Model 395:11–22. https://doi.org/10.1016/j.ecolmodel.2018.12.013
Pfeiffer M, Kumar D, Martens C, Scheiter S (2020) Climate change will cause non-analogue vegetation states in Africa and commit vegetation to long-term change. Biogeosciences 17:5829–5847
Pfeiffer M, Hoffmann MP, Scheiter S, Nelson W, Isselstein J, Ayisi K, Odhiambo JJ, Rötter R (2022) Modeling the effects of alternative crop-livestock management scenarios on important ecosystem services for smallholder farming from a landscape perspective, Biogeosciences, 19:3935–3958. https://doi.org/10.5194/bg-19-3935-2022
Pool-Stanvliet R (2013) A history of the UNESCO man and the biosphere Programme in South Africa. South Afr J Sci 109. https://doi.org/10.1590/sajs.2013/a0035
Rötter RP, Scheiter S, Hoffmann MP, Pfeiffer M, Nelson WCD, Ayisi K, Taylor P, Feil J-H, Bakhsh SY, Isselstein J, Lindstädter A, Behn K, Westphal C, Odhiambo J, Twine W, Grass I, Merante P, Bracho-Mujica G, Bringhenti T, Lamega S, Abdulai I, Lam QD, Anders M, Linden V, Weier S, Foord S, Erasmus B (2021) Modeling the multi-functionality of African savanna landscapes under global change. Land Degrad Dev 32:2077–2081. https://doi.org/10.1002/ldr.3925
Ruppert JC, Linstädter A (2014) Convergence between ANPP estimation methods in grasslands — a practical solution to the comparability dilemma. Ecol Indic 36:524–531. https://doi.org/10.1016/j.ecolind.2013.09.008
Ruppert JC, Harmoney K, Henkin Z, Snyman HA, Sternberg M, Willms W, Linstädter A (2015) Quantifying drylands’ drought resistance and recovery: the importance of drought intensity, dominant life history and grazing regime. Glob Chang Biol 21:1258–1270. https://doi.org/10.1111/gcb.12777
Sankaran M (2019) Droughts and the ecological future of tropical savanna vegetation. J Ecol 107:1531–1549. https://doi.org/10.1111/1365-2745.13195
Sankaran M, Hanan NP, Scholes RJ, Ratnam J, Augustine DJ, Cade BS, Gignoux J, Higgins SI, Roux XL, Ludwig F, Ardo J, Banyikwa F, Bronn A, Bucini G, Caylor KK, Coughenour MB, Diouf A, Ekaya W, Feral CJ, February EC, Frost PGH, Hiernaux P, Hrabar H, Metzger KL, Prins HHT, Ringrose S, Sea W, Tews J, Worden J, Zambatis N (2005) Determinants of woody cover in African savannas. Nature 438:846–849. https://doi.org/10.1038/nature04070
Savory A, Butterfield J (2016) Holistic management: a commonsense revolution to restore our environment, 3rd edn. Island Press, Washington, DC
Scheiter S, Higgins SI (2007) Partitioning of root and shoot competition and the stability of savannas. Am Nat 170:587–601
Scheiter S, Higgins SI (2009) Impacts of climate change on the vegetation of Africa: an adaptive dynamic vegetation modelling approach (aDGVM). Glob Chang Biol 15:2224–2246
Scheiter S, Savadogo P (2016) Ecosystem management can mitigate vegetation shifts induced by climate change in West Africa. Ecol Model 332:19–27. https://doi.org/10.1016/j.ecolmodel.2016.03.022
Scheiter S, Langan L, Higgins SI (2013) Next generation dynamic global vegetation models: learning from community ecology. New Phytol 198:957–969. https://doi.org/10.1111/nph.12210
Scheiter S, Gaillard C, Martens C, Erasmus BFN, Pfeiffer M (2018) How vulnerable are ecosystems in the Limpopo province to climate change? South Afr J Bot 116:86–95. https://doi.org/10.1016/j.sajb.2018.02.394
Scheiter S, Schulte J, Pfeiffer M, Martens C, Erasmus BFN, Twine WC (2019) How does climate change influence the economic value of ecosystem Services in Savanna Rangelands? Ecol Econ 157:342–356. https://doi.org/10.1016/j.ecolecon.2018.11.015
Scheiter S, Moncrieff GR, Pfeiffer M, Higgins SI (2020) African biomes are most sensitive to changes in CO2 under recent and near-future CO2 conditions. Biogeosciences 17:1147–1167
Shackleton CM, Shackleton SE, Netshiluvhi TR, Mathabela FR (2005) The contribution and direct-use value of livestock to rural livelihoods in the Sand River catchment, South Africa. Afr J Range Forage Sci 22:127–140
Siebert F, Dreber N (2019) Forb ecology research in dry African savannas: knowledge, gaps, and future perspectives. Ecol Evol 9:7875–7891. https://doi.org/10.1002/ece3.5307
Siebert F, Klem J, Van Coller H (2020) Forb community responses to an extensive drought in two contrasting land-use types of a semi-arid Lowveld savanna. Afr J Range Forage Sci 37:53–64. https://doi.org/10.2989/10220119.2020.1726464
Stats SA (2021) Statistical release P0302: mid-year population estimates 2021
Staver AC, Archibald S, Levin S (2011) Tree cover in sub-Saharan Africa: rainfall and fire constrain forest and savanna as alternative stable states. Ecology 92:1063–1072
Stevens N, Lehmann CER, Murphy BP, Durigan G (2017) Savanna woody encroachment is widespread across three continents. Glob Chang Biol 23:235–244. https://doi.org/10.1111/gcb.13409
Tang W, Bennett DA (2010) Agent-based modeling of animal movement: a review. Geogr Compass 4:682–700. https://doi.org/10.1111/j.1749-8198.2010.00337.x
Tielbörger K, Bilton MC, Metz J, Kigel J, Holzapfel C, Lebrija-Trejos E, Konsens I, Parag HA, Sternberg M (2014) Middle-eastern plant communities tolerate 9 years of drought in a multi-site climate manipulation experiment. Nat Commun 5:5102. https://doi.org/10.1038/ncomms6102
Twine WC, Holdo RM (2016) Fuelwood sustainability revisited: integrating size structure and resprouting into a spatially realistic fuelshed model. J Appl Ecol 53:1766–1776. https://doi.org/10.1111/1365-2664.12713
Twine W, Moshe D, Netshiluvhi T, Siphugu V (2003) Consumption and direct-use values of savanna bio-resources used by rural households in Mametja, a semi-arid area of Limpopo province, South Africa. South Afr J Sci 99:467–473
van Langevelde F, van de Vijver CADM, Kumar L, van de Koppel J, de Ridder N, van Andel J, Skidmore AK, Hearne JW, Stroosnijder L, Bond WJ, Prins HH, Rietkerk MM (2003) Effects of fire and herbivory on the stability of savanna ecosystems. Ecology 84:337–350
Van Staden N (2016) Herbaceous species diversity, redundancy and resilience of Mopaneveld across different land-uses. MSc dissertation, North-West University
van Vuuren DP, Edmonds J, Kainuma M, Riahi K, Thomson A, Hibbard K, Hurtt GC, Kram T, Krey V, Lamarque J-F, Masui T, Meinshausen M, Nakicenovic N, Smith SJ, Rose SK (2011) The representative concentration pathways: an overview. Clim Chang 109:5–31. https://doi.org/10.1007/s10584-011-0148-z
Venter ZS, Hawkins H-J, Cramer MD (2017) Implications of historical interactions between herbivory and fire for rangeland management in African savannas. Ecosphere 8:e01946. https://doi.org/10.1002/ecs2.1946
Venter ZS, Cramer MD, Hawkins H-J (2018) Drivers of woody plant encroachment over Africa. Nat Commun 9:2272. https://doi.org/10.1038/s41467-018-04616-8
Wigley BJ, Fritz H, Coetsee C (2018) Defence strategies in African savanna trees. Oecologia 187:797–809. https://doi.org/10.1007/s00442-018-4165-8
Wigley-Coetsee C, Staver A (2020) Grass community responses to drought in an African savanna. Afr J Range Forage Sci 37:43–52. https://doi.org/10.2989/10220119.2020.1716072
Yahdjian L, Sala OE (2002) A rainout shelter design for intercepting different amounts of rainfall. Oecologia 133:95–101. https://doi.org/10.1007/s00442-002-1024-3
Acknowledgments
The “Limpopo Living Landscapes (LLL)” and “South African Limpopo Landscapes Network (SALLnet)” projects are part of the SPACES/SPACES II programs (Science Partnerships for the Adaptation to Complex Earth System Processes in Southern Africa), funded by the BMBF (German Federal Ministry of Education and Research, grants 01LL1304B, 01LL1304D, 01LL1802B and 01LL1802C) under FONA (Research for Sustainable Development).
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2024 The Author(s)
About this chapter

Cite this chapter
Scheiter, S., Pfeiffer, M., Behn, K., Ayisi, K., Siebert, F., Linstädter, A. (2024). Managing Southern African Rangeland Systems in the Face of Drought: A Synthesis of Observation, Experimentation and Modeling for Policy and Decision Support. In: von Maltitz, G.P., et al. Sustainability of Southern African Ecosystems under Global Change. Ecological Studies, vol 248. Springer, Cham. https://doi.org/10.1007/978-3-031-10948-5_16
Download citation
DOI: https://doi.org/10.1007/978-3-031-10948-5_16
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-10947-8
Online ISBN: 978-3-031-10948-5
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)