
Abstract
This chapter deals with the conventional upstream fish passage technologies developed for a safe fish migration at run-of-river hydropower plants (HPPs). It covers the factors influencing successful migration, current status, practices and challenges, novel developments, recommendations and outlook. Main focusses of this chapter are on the attractiveness of fishways and bypass facilities based on FIThydro study results. Although upstream fish passage technologies are well developed, there are still research gaps on their effectiveness related to their attractiveness and passabilities. FIThydro fills such research gaps by providing tools, methods, devices and best practice examples. This chapter covers the field studies at the case study HPP Guma with pool and weir fishway in Spain and Schiffmühle with vertical-slot and nature-like fishways in Switzerland and a combination of field, laboratory and numerical simulation studies of vertical-slot fishway in France. The results of these three studies contribute to better fishway design for a range of fish species and hydraulic conditions.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others
5.1 Passability: Fish Swimming Behaviour and Fishpass Preferences (e.g. Schiffmühle)
5.1.1 Introduction
Migration and movements of fishes can be observed on a daily, seasonal or annual basis. Individuals move and migrate over short or long distances. Movement is considered to be a change of location or habitat within a river, and is mainly to seek basic resources (food, shelter) or as a reaction to avoid predators. Dingle (2014) emphasizes that most movements take place within a defined area or home range. Foraging (movement in search of resources) or commuting (movement in search of resources, typically daily) are typical examples (Dingle 2014). The size of the home range depends on an individual’s swimming ability and behaviour. In general, movement is a repetitive behaviour in time and space. Migration, however, is different from movement. Migration is not carried out to seek available resources (food, habitat), but is a distinct and specialized behaviour involving leaving one habitat and settling in another outside the home range (Dingle 2014). Cyprinid fishes (Lucas and Baras 2001) and salmonids change habitats often during their life cycle, and both show homing behaviour, i.e. the behaviour of spawners returning to the streams or spawning areas in which they spent their early life stages and which is therefore considered to be a suitable habitat for reproduction (Wootton 1990). To fulfil their natural life cycle, fishes depend on an intact migration corridor that is not artificially fragmented.
Considering the need for movement and migration, it is very important for hydropower to be produced in a way that allows fish to use the various habitats and to switch between them in a river system. River fragmentation by dams and its effects on fishes is a worldwide challenge.
New legislation in EU and Switzerland therefore requires fish migration facilities at hydropower plants (HPP) to be built according to the latest standards, focusing on target species while also allowing non-target species to fully ascend the devices built for upstream migration (fish ladders, nature-like bypass systems) or descend them (physical screens and angled bar racks with bypasses).
Success monitoring must also be carried out in order to verify the fish passage efficiency of upstream and downstream fishpasses. Evaluating a fishway’s efficiency after construction is crucial ensuring that the structure serves its purpose and for making any necessary adjustments (Noonan et al. 2012). Therefore, this study deals with the performance evaluation of upstream and downstream fishpasses installed at the case study HPP Schiffmühle in Switzerland by conducting a fish monitoring.
5.1.2 Case Study Hydropower Plant Schiffmühle, River Limmat, Switzerland
There are two run-of-river HPPs Schiffmühle on the River Limmat, which is the outflow of Lake Zurich: the main powerhouse and the residual flow HPPs. The residual flow HPP served as a case study in the FIThydro project (https://www.fithydro.eu/schiffmuhle/) and all the descriptions below refer to this plant. The design discharge of the HPP is 14 m3/s, and the head is 3.17 m. Its electricity generation capacity is 0.5 MW (bevel gear bulb turbine). The 75 m long upstream migration facility for upstream migrating fishes is a combination of a nature-like fishpass (NL) and a vertical-slot (VS) fishpass (Figs. 5.1 and 5.2). The entrance into the NL pass is 36 m downstream of the turbine flow outlet (entrance angle about 40 degrees). The entrance into the VL pass, which has a 6.3% slope, is located 2 m downstream of the turbine flow, with an entrance angle of 90 degrees. Total discharge of the pass is 500 l/s. After 10 pools in the VS pass and 12 pools in the NL pass, the two passes merge and an additional 12 pools follow in the upstream direction until the exit.

HPP Schiffmühle, River Limmat. VS: vertical-slot pass, NL: nature-like pass, EX: exit of fishpass, CF: counting facility of the fishpass, HS: horizontal bar rack, ENBY: entrance bypass, EXBY: exit bypass

a PIT-tagging antenna in the nature-like fishpass, b antenna in the vertical-slot pass, c turbine inlet with horizontal bar rack and d close-up photo of bypass system
The technical device for downstream migration is a Horizontal Bar Rack-Bypass System (HBR-BS) (Figs. 5.1 and 5.2, also see Sect. 7.2). The rack is positioned almost parallel to the main flow, with a lateral intake. The rack is 14.6 m long and 1.82 m high, and the clear bar spacing is 20 mm. The bars have rectangular profiles. The average velocity in front of the screen is 0.5 m/s at the design discharge. It is intended to guide fish into a bypass with two openings in a vertical chamber at different water depths (bottom and water surface). There is a 25 cm diameter pipe bypass to lead the fish to the downstream part of the HPP. The discharge of the bypass pipe is 170 l/s.
This study answers the following research questions:
-
What is the passage efficiency of fishpasses for upstream migration fishes and how much time does a successful passage take?
-
Do fishes have a special preference for one of the two entrances?
-
Are all fish species and all fish sizes able to negotiate the fish passes successfully, and what are their migration seasons?
-
Are downstream migrating individuals able to find the entrance into the bypass?
5.1.3 Methods
PIT-tagging was used to detect the migration of fishes. Two antennae were installed at each entrance, one of them as close as possible to the entrance, and a second in the fourth or fifth pool of the ladder. One additional antenna was installed in the upper part of the fish ladder where fish leave the fishpass. In order to detect downstream migration, one antenna was placed in the bypass pipe close to the entrance.
Individuals were tagged with HDX (half-duplex) Pit-tags (Oregon RFID, Texas Instruments ISO 11784/11785) and measured (total length). 23 mm (individuals > 150 mm) and 12 mm tags were used. A total of 3087 individuals were tagged between September 2017 and September 2019. Tagged fish were allowed to recover and then released 210 m downstream of the nature-like fishpass entrance on the left riverbank. Data treatment was carried out with R (R development core team 2013).
Attraction efficiency was defined as the proportion of individuals tagged that were subsequently detected at the first antenna at the fishpass entrance. Entrance efficiency was the number of fish detected at the second antenna divided by the number of fish registered at the entrance antenna.
Fish passage efficiency was defined by dividing the number of fish exiting the fishway by the number of fish detected at the second antenna at the pass entrance (Bunt et al. 2012). Passage time was calculated as the time from the last detection at the entrance antennae to successful exit.
5.1.4 Passage Efficiency
A total of 13 fish species were tagged within the group of the 2890 individuals caught in the pass counting basin. The rheophilic barbel (Barbus barbus), chub (Squalius cephalus) and spirlin (Alburnoides bipunctatus) representative of schooling fishes, were the most abundant individuals tagged. Other tagged fish species were brown trout, perch (Perca fluviatilis), bullhead, gudgeon, dace, bleak (Alburnus alburnus), nase (Chondrostoma nasus), roach, rudd and pumpkinseed.
Most of the tagged fish were between 90 and 260 mm long. 67.3% of the tagged individuals were redetected after tagging (attraction efficiency), 95.5% of them entered the fishpasses. Other studies have documented smaller attraction values (32.9% by Benitez et al. 2018), or approximately 30% for all entrances at the HPP Rheinfelden in the River Rhine (Peter et al. 2016). In contrast, Noonan et al. (2012) published average attraction efficiencies of 65.1%; however, most of these studies were carried out on salmonids, which have a high motivation to migrate. A meta-analysis by Bunt et al. (2012) also documented average attraction efficiencies of 66% (different fish species and types of migration facilities). The obtained values in the Limmat can therefore be classified in the mid-range but clearly higher than in the study in the Rhine. Fish used different entrances into the fishpasses. Bleak showed no preference for one of the entrances, but dace did show a clear preference. The attraction efficiency for dace for the VS fishpass was only 9.8%: conversely, 79.3% preferred the NL fishpass. The same clear preference was also observed for roach (Rutilus rutilus, 69.1%). Chub and perch likewise preferred the nature-like fishpass, however distinctly less. Species that preferred the VS fishpass were barbel and, to a lesser extent, spirlin.
The entrance efficiency over all species was 98.5% in the VS fishpass and 87.9% in the NL fishpass, indicating a high efficiency in this study. The passage efficiencies were 78.5% for the VS and 73.6% for the NL fishpasses, respectively and the passage efficiency for the whole fishpass was 81.4% or all species. Such high efficiencies are comparable to the values published by Benitez et al. (2018) with 86.3%, but clearly higher than those published by Noonan et al. (2012, value 41.7%) and Bunt et. al. (2012) (values 45–70%). Finally, the passage efficiencies at the Schiffmühle HPP have to be assessed as good.
5.1.5 Migration Time
Four main periods were observed for the time of migration, coinciding with the time the tagged fish were released. Many ascents were observed after each release of fishes. The four periods were October 2017, May–June 2018, September-mid November 2018, and July–November 2019. The effect of the release date on the date of registration by the antennae was obvious. The tagged and released fish continued their migration without interruption. However, between December and April the antennae registered almost no fishes. The migration activity in winter is thus very low.
The values for the median and the minimum time needed for the passage are very meaningful. For most of the fish species, 50% of all individuals needed less than 60 min for the passage. Spirlin generally took well over 60 min. However, the passage time of the fastest spirlin was only 15 min (Table 5.1).
The passage times in the NL fishpass were somewhat longer than in the VS fishpass. The median passage time increased by 25 min (not including spirlin). The spirlin had a clearly longer passage time. A chub had the fastest passage, with 6.4 min to reach the exit of the fishpass (Table 5.2).
In general, the time needed for the upstream passage is short and the observed passage time of about one hour for more than 50% of the individuals cannot be regarded as an impairment of migration.
5.1.6 Migration Distance
769 fish (25% of the tagged individuals at the HPP Schiffmühle) were detected at the fishpass facility of the HPP Aue, which is situated 7.03 km upstream of the HPP Schiffmühle. All of them had to successfully pass the HPP Kappelerhof, which is 2.17 km upstream. Mainly dace (73.5%), roach (62.9%) and nase (60%) tagged at HPP Schiffmühle migrated as far as HPP Aue. However, 37.1% of the bleak also migrated the 7.03 km distance.
5.1.7 Downstream Migration
Over the whole observation period a total of 445 tagged fish descended downstream. Only two fish used the bypass for downstream migration. Other migration corridors were the NL fishpass (N = 56 individuals), the VS fishpass (N = 122 individuals) and unknown corridors (N = 265 individuals). The unknown corridors include the downstream migration over the spillway, through the turbine or through the main HPP at Schiffmühle (Fig. 5.1).
Most of the individuals used unknown migration corridors. The bypass was often clogged with wood affecting the bypass flow and discharge. Furthermore, the attraction flow to the bypass seems inefficient and a recirculation zone possibly affects fish searching and finding the bypass entrance. Moreover, the acceleration at the beginning of the pipe was far too high. The bypass at HPP Schiffmühle is therefore far from fully functional. Fishes migrating downstream used alternative corridors.
5.1.8 Conclusions
Knowledge of the passage efficiency at each single dam is very important for assessing the cumulative impacts of 10 consecutive dams in the River Limmat. Low passage efficiency can have a detrimental impact on populations. PIT-tagging studies are a very useful tool to assess upstream migration facilities. Attraction efficiencies, passage efficiencies and passage time are important parameters for evaluating success. Our results demonstrate that Both vertical slot and nature-like fishpasses at HPP Schiffmühle function well for upstream moving fish with high attractiveness, entrance and passage efficiencies. Species-specific preferences were observed for the available entrances. Having more than one entrance into a fish ladder can therefore be an advantage in rivers with a broad range of species. Tagged fish continued their migration without interruption, proving that the attraction of the fishpass was good. A considerable number of individuals migrated further upstream in the River Limmat over a distance of 5.1 km. An important parameter for assessing swimming performance in the fishpass is the time needed for the passage. The time taken for the ascent was generally low.
The bypass for downstream migration was barely used at all, and serious problems were detected in terms of clogging of the bypass entrance, unfavourable flow conditions around the bypass and high acceleration at the entrance into the pipe. This result indicates that design, location and operation of a bypass system are of prime importance for a successful implementation and high fish guidance efficiency of HBR-BS. Therefore, bypass system needs optimization.
Overall, the present findings have a wide range of applications for other similarly sized HPPs and will serve as a basis for an optimal design of fishpasses and HBR-BS for various fish species.
5.1.9 Acknowledgements
We thank the Limmatkraftwerke AG (Andreas Doessegger, Peter Rotenfluh, Christoph Froelich and his team), the fishing club (Pachtvereinigung Stausee Wettingen) with Reto Wittwer and his team for the support for the fish tagging, and Tabea Kropf (Sektion Jagd und Fischerei des Kantons Aargau).
5.2 Assessing the Effectiveness of Migration Facilities in Guma Iberian Testcase
5.2.1 Introduction
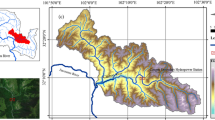
The FIThydro testcase Guma is situated in the Duero River, northwest part of Spain (Fig. 5.3). The Duero River basin presents a high degree of fragmentation with more than 140 small hydropower plants (HPP) (below 10 MW in EU) and 23 large HPP in the Spanish part of the basin, as well as nearly 5200 other obstacles for irrigation, domestic and industrial water supply (www.chduero.es). The presence of these obstacles causes a high disruption of the fluvial longitudinal connectivity, which hinders or even prevents fish migration, among other associated environmental problems related to discharge abstraction and sediment management (Branco et al. 2017; Nilsson et al. 2005).

Location of Guma testcase and associated facilities
The hydrology of the Duero River in the river section affected by Guma HPP is characterized by low flows in summer (exacerbated by upstream water diversions for irrigation) and medium to high flows during winter and early spring, associated with the rainy season and snow-melting episodes. The river reach is dominated by small and medium-size potamodromous rheophilic endemic cyprinids, such as Iberian barbel (Luciobarbus bocagei), northern straight-mouth nase (Pseudochondrostoma duriense), northern Iberian chub (Squalius carolitertii), and Pyrenean gudgeon (Gobio lozanoi). Their populations are currently suffering an important decrease (specially nase, categorized as endangered (IUCN, 2018)), whereas alien species are increasing their presence (mainly the bleak (Alburnus alburnus)). River discharge during cyprinid upstream migration (April–July) is between 15 and 30 m3/s. The river reach belongs to the Epipotamon zone (Illies and Botoseanu 1963) with an average altitude of around 810 m a.s.l. and corresponds to C6 category (Rosgen and Silvey 1996).
The Guma HPP is operated by Salto de Vadocondes S.A. (SAVASA). It is a run-of-river HPP with a total height of 8.85 m, an installed capacity of 2.25 MW, and 2 Kaplan turbines (Fig. 5.3). Due to the run-of-river configuration, there is no legal requirement for maintaining a minimum environmental flow through the dam. In the right bank of the dam, there is a pool and weir type fishway composed of 36 cross-walls with submerged notches and bottom orifices. Through the fishway runs a discharge of 0.25–0.50 m3/s. This discharge ensures proper operation of the fishway and it is additionally supported by a supplementary attraction flow of an additional channel near the fishway entrance (Table 5.3).
The main FIThydro objective in this testcase was to evaluate the effectiveness of the fishway and the overall performance of the facility for bidirectional fish migration. The Testcase is representative for the most common HPP configurations in Mediterranean environments; therefore, its results are of interest to advance in the understanding of bidirectional fish migration in the Mediterranean area. In addition, this information will help operators to fulfill the requirements of cost-effective energy production and, at the same time, meet the environmental requirements and targets under European legislation.
5.2.2 Materials and Methods
To evaluate the effectiveness of migration facilities, native fish species (Luciobarbus bocagei, Pseudochondrostoma duriense, Squalius carolitertii) were PIT tagged. Iberian barbel individuals represented more than 90% of the sample. Three sets of experiments were carried out: (1) Fishway evaluation under free conditions (fish released in the river), (2) fishway evaluation under confined conditions (fish locked in the fishway), and (3) punctual downstream fish movement analysis in the fishway.
The fish samplings for the first and second experiments were carried out from May to October 2018 and 2019 (at least one per month), collecting fish from different origins (i.e. downstream, upstream, and inside the fishway). In both experiments, fish movements were recorded using a pass-through antenna system, which consisted of four antennas installed in the fishway, covering notch and orifice passage, and connected to a dedicated reader (ORFID® Half Duplex multiplexer reader).
For the first experiments (free conditions), 754 fish were tagged using different release places (411 downstream and 343 upstream, and in both river banks, while in the second set of experiments (confined conditions, n = 153) fish were locked inside the fishway for less than 24 h.
The third set of experiments (downstream movement analysis) was carried out between June and October 2020 (with monthly frequency). They consisted of the capture of all fish inside the fishway, their release downstream outside the fishway, the installation of a close mesh in the middle side turning pool of the fishway, and the counting of fish upstream and downstream of the mesh after 24 h, to identify upstream and downstream movements of fish.
All procedures were carried out following national and community legislation and ethical guidelines about research with animals (Directive 2010/63/UE, and Spanish Act RD 53/2013).
5.2.3 Results
Upstream Migration
For experiments under free conditions, 204 fish from 411 downstream released fish, located and entered the fishway (50% in 2018 and 31% in 2019), of which 129 had success in the fishway ascent (61%). Additionally, 17 fish released in 2018 located and entered the fishway in 2019, which would increase the proportion of location in the subsequent year. Fish spent a median of 9 days (InterQuartile range: 5–19 days) locating the fishway. Both fishway location and passage success were influenced by the origin of the fish. In addition, for the Iberian barbel, fish length significantly affected fishway location (median for those that did not locate the fishway of 123 mm versus 142 mm for successful location); however, fish length did not show significant differences in the ascent success. Fishway location and ascent success were also related to environmental variables, i.e. total river discharge, water level difference between upstream and downstream the dam, and water temperature. There was a period of peak movement mainly during the second half of June and the first days of July, related to changes in the river discharge and water temperature (Fig. 5.4). Regarding the transit time, the global median time for ascending the fishway was 3.4 h (median transit time per meter of height ascended was 26 min/m; 31 pools with water drops of 0.25 m, i.e. a total water head assessed near 8 m). The transit time varied significantly according to the fish origin, which seems to indicate a higher motivation for the ascent of those fish from upstream origin.

Location and ascent success of Guma fishway, related to river discharge and water temperature. Water head refers to the difference between water level upstream and downstream. Between 30/10/2018 and 01/04/2019 data logger was running but there were no data at the antennas
For confined experiments, more than 90% of fishpassed the fishway successfully and the median transit time to ascend 2.25 m in height was lower than 23 min (median transit time per meter of height ascended was 10.2 min/m). Fish length had a significant effect on ascent time, with the larger fish the faster to ascend the fishway. Fish in confined experiments were faster than fish in free experiments, presenting speeds similar to those obtained in a vertical slot fishway with the same discharge and under confined conditions (see details in Bravo-Córdoba et al. 2018).
Downstream Migration
The collected data disclosed that the Guma fishway can be used as a downstream migration route. Several fish moved upstream and downstream were identified through the fishway during the tracking period. Regarding fish that were released specifically upstream Guma dam (2018 and 2019), 42% (144/343) were found in the fishway and 64% (92/144) of these fish completed the downstream movement (Fig. 5.5). About monthly samplings in the fishway (third experiment, spring–autumn 2020), for the two main sampled species (Luciobarbus bocagei and Alburnus alburnus), there was a relevant proportion of fish that entered the fishway with downstream migration direction (Table 5.4).

Main results of downstream migration trials in Guma fishway
5.2.4 Conclusions
-
Guma fishway is used not only for upstream migration but also for downstream migration and other types of movements related to the search of refuge or feeding purposes.
-
Fish were found using the fishway during most of the year, except in winter and their movements were mainly related to water temperature and peak discharge events.
-
The fishway does not seem to cause an important migration delay and it is suitable for upstream migration, at least for Iberian barbel.
-
Origin and length of the fish were identified as relevant factors. Fish from upstream origin showed a higher motivation for ascending and smaller fish showed a lower ascending success. A high degree of immature sizes was found using the fishway, thus, the lower success of small fish could be related to the absence of reproduction motivation.
5.3 Adaptation of Vertical Slot Fishways to Multi Fish Species with Macro-Roughness
5.3.1 Introduction
Vertical Slot Fishways (VSF) are technical solutions developed from more than 30 years for upstream fish migration. As they have been designed at first for salmonids who have high swimming capacities, they are not always suitable for other fish species. Adaptive technical solutions have been developed with the objective to facilitate the upstream migration for all the fish species and in particular for the small species (Albayrak et al. 2020).
5.3.2 Methods and Results
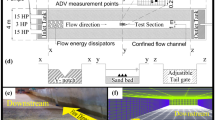
A basic Vertical Slot Fishway geometry is established (Fig. 5.6) by an analysis of the geometry of VSFs built over the last 20 years, from which we defined an ‘average’ geometry.

Pool configuration and axes of references (Z is the vertical axis)
The main geometric ratios of the pool are the L/b which is fixed to 10 with L the length of the pool and b the width of the slot, B/b which varies from 9 to 5.67 with B the pool width. The baffles in the pools have the following ratios: A/b = 2 and a/b = 1.3. To ensure the migration of individuals or species with low swimming ability, these fishways can be improved by inserting elements such as vertical cylinders, sills in the slot, bottom roughness or flexible structures, which were suggested by Tarrade et al. (2011), Calluaud et al. (2014) or Ballu et al. (2019). The influence of added elements on the flow features is highlighted by laboratory and field experiments and flow numerical simulations. They were carried out for several configurations and for a wide range of channel slopes, pool widths and flow discharges. The modification of the flow topology, discharge coefficients and turbulence features were evaluated by ADV and water depth measurements. The results presented herein may be used to define a predictive law that helps engineers ensure the greatest effectiveness of VSFs with the presence of elements (Ballu et al. 2019). For example, macro-roughness are more and more often positioned at the bottom of fishways and they can facilitate the passage of benthic species through the crossing device (Branco et al. 2015). These are useful to reduce bottom velocity and create hydraulic shelters. However, the presence of macro-roughness is not yet taken into account when designing a VSF.
The fish migration efficiency varies with the flow topologies which are described in details in Calluaud et al. (2014). In particular, two salient flow topologies can be generated depending on the length to width ratios of the pools and on the slope, called the first flow pattern (FP1) and the second flow pattern (FP2). The FP1 provides better efficiency for the upstream migration and is preferred for the design of a VSF. A mapping of the flow topology (Fig. 5.7) as a function of the pool width, the slope s expressed in %, and discharge is given for three densities of macro-roughness’s, noted dr (dr is the surface occupied by the macro-roughness divided by the surface of the bottom of the pool). The density dr is defined as the ratio between the area covered by elements with macro roughness and the total area on which the elements are positioned.

Mapping of the transition zones between the type 1 topology and the type 2 topology as a function of the macro-roughness density
For the three macro-roughness densities, the graph of Fig. 5.7 delimits the transition zones of flow pattern for each parameter. It can be seen that for all configurations, an increase in the slope of the fishway forces the flow to adopt a FP2. In agreement with Wang et al. (2010) the flow discharge has no significant impact on the flow topology for dr = 0%. Since macro-roughness are introduced at the bottom of the fishway, the transition between FP1 and FP2 spreads over a wider range of pool width. This effect is amplified when the density of macro-roughness increases, where the change from FP1 to transition occurs at a greater B/b and the change from transition to FP2 at a smaller B/b. For instance, for Q = 23 l/s and s = 5%, the flow changes from FP1 to transition at B/b = 7 for dr = 0%, at B/b = 7.67 for dr = 10%, and finally at B/b = 8.33 for dr = 15%. Likewise, the flow changes to FP2 for B/b = 6.67 for dr = 0%, B/b = 5.67 for dr = 10% and B/b smaller than 5.67 for dr = 15%. The flow discharge has an important role with dr = 10% and dr = 15%, because it increases also the transition area between the two Flow Patterns. As the slope grows, the flow shifts from FP1 to transition for higher values of B/b when dr = 0% and dr = 10%. However, this trend tends to be reversed when dr = 15%.
The influences of discharge, pool slope, pool width and density of macro-roughness on the discharge coefficient Cd are illustrated in Fig. 5.8. The discharge coefficient Cd (−) is the ratio between the measured discharge Q (m3/s) and the theoretical discharge, which depends on the water depth at the centre of the pool and is expressed as follows, where g is the acceleration due to gravity (ms−2), b is the width of the slot (m), h0 is the water depth (m) at the centre of the pool, Δh the head difference (m) between successive basins, and Q is the discharge (m3/s).

Evolution of the discharge coefficient Cd with: a the flow discharge, b the slope and c the pool width
The uncertainty is reported on each plot using error bars. The mean value of relative uncertainty on the Cd is about 7% with a coverage factor k = 2.
Figure 5.8a shows that the value of Cd is not significantly affected by the change in Q. The biggest variation of Cd with the Q is measured when dr = 0% and is about 0,03 ± 0,1 with k = 2. In contrast, an increase in the slope creates a significant decrease of Cd, with a minimum of −20% ± 7% with k = 2 and dr = 15% (Fig. 5.8b). As regards to the influence of B/b (Fig. 5.8c), there is also a significant decrease of Cd when the pools become wider. The reduction of the discharge coefficient between B/b = 5.67 and B/b = 9 is about −15% ± 8% with k = 2.
The mean evolution of Cd values between s = 5% and s = 12.5% was calculated for each of the four widths studied. With macro-roughness, an increase in pool width reveals a more important influence of the slope on the discharge coefficient value. For example, for B/b = 5.67 the relative decrease in Cd between s = 5% and s = 12.5% is about 4% for dr = 10% and 15% against more than 12% for B/b = 9. Without macro-roughness, it remains on average around 10%. Consequently, in the presence of macro-roughness and for small pool widths, the discharge coefficient will hardly be affected by a change in the channel slope.
To quantify the influence of macro-roughness density on the kinematic quantities of the flow, a comparison of the velocity and turbulent kinetic energy profiles (TKE) obtained for a pool width B/b = 6.67, a slope s = 7.5% and a flow rate Q = 23L/s in the without macro-roughness configuration, dr = 10% and dr = 15% is performed. When B/b = 6.67, the topology is a transient type in the three configurations (smooth floor, dr = 10% and 15%). The graphs in Fig. 5.9 show the values of the velocity norm and turbulent kinetic energy \(\frac{{\mathrm{k}}_{3\mathrm{D}}}{{{\mathrm{V}}_{\mathrm{d}}}^{2}}\), for one transversal profile at mid depth in the configuration B/b = 6.67.

Three-dimensional mean velocity \(\frac{\Vert {{\varvec{V}}}_{3{\varvec{D}}}\Vert }{{{\varvec{V}}}_{{\varvec{d}}}}\) and Turbulent Kinetic Energy \(\frac{{k}_{3D}}{{{V}_{d}}^{2}}\) according to the macro-roughness densities for a pool width of B/b = 6.67
The presence of macro-roughness induces a decrease of the Turbulent Kinetic Energy k3D/Vd2. For the two macro-roughness densities, between Y/b = 0 et Y/b = 2, k3D/Vd2is reduced by an average of 40% ± 7% compared to the smooth floor configuration. On the 4 ≤ Y/b ≤ 6 interval, this reduction is estimated at an average 26% ± 7%. It should be noted that on the first interval, it is the streamwise component kU/Vd2 which mainly contributes to the decrease in the global Turbulent Kinetic Energy k3D/Vd2, while it is equally distributed between kV/Vd2et kW/Vd2on the second interval. On the other hand, the impact of increased density of macro-roughness is negligible.
For this configuration (B/b = 6,67), the flow is transient regardless of the macro-roughness density. The area near the wall opposite the slot (\(4\le \frac{Y}{b}\le 6\)) is then subject to large velocity variations: it alternates between type 1 flow and type 2 flow. The x component of the velocity is the more impacted by these variations, because this part of the profile is located in the jet in type 2 flow (highly positive velocity) and in the recirculation zone in the case of type 1 flow (negative velocity). With macro-roughness, the amplitude of these variations seems reduced.
The lower part of the profile (0 ≤ Y/b ≤ 2) is located in a recirculation zone. The presence of macro-roughness seems to create a constraint, independent of density, which decreases the amplitude of vertical and transverse velocity fluctuations.
The turbulent kinetic energy \(\frac{{k}_{3D}}{{{V}_{d}}^{2}}\) is reduced by an average of 25% ± 15%. Areas of strong influence are mostly located on either side of the jet (Y/b ≤ 2 et ≥ 4) and the three fluctuating components contribute equally to this decrease.
5.3.3 Conclusions
Vertical Slot Fishways (VSF) have been studied in this work to modify them and allow the upstream passage of not only salmonids but also species with small swimming capacities. Adaptive technical solutions have been developed and tested like the adjunction of one or three cylinders in the pool, sills in the slots, macro-roughness in the bottom of the pool or flexible cylinders inside the pool. The technical solutions proposed have shown some large modifications of the hydraulics and in particular the flow quantities which govern the fish motion (Velocity, Velocity Gradient, Turbulent Kinetic Energy and Dissipation), are reduced highly with the adjunction of elements. Values of the discharge coefficient for these different configurations are given to help the engineers to design such vertical slot fishways. A methodology is proposed which defines first the topology of the flow from the geometric parameters and finally predicts the discharge coefficients, (Ballu et al. 2019). These different options could be used to modify existing, poorly functioning fishways and could allow the upstream migration of a greater number of fish species, at a lower cost than building a completely new fishway.
References
Albayrak I, Boes R, Beck C, Meister J, David L, Lemkecher F, Chatellier L, Courret D, Pineau G, Calluaud D, Larrieu T, Sagnes P, Geiger F, Rutschmann P (2020) D3.4 – Enhancing and customizing technical solutions for fish migration. FIThydro Project Report, https://www.fithydro.eu/deliverables-tech/
Ballu A, Calluaud D, Pineau G, David L (2019) Experimental-based methodology to improve the design of Vertical Slot Fishways. J Hydraul Eng 145(9):04019031. https://doi.org/10.1061/(ASCE)HY.1943-7900.0001621
Benitez JP, Dierckx A, Nzau Matondo B, Rollin X, Ovidio M (2018) Movement behaviours of potamodromous fish within a large anthropised river after the reestablishment of the longitudinal connectivity. Fish Res 207:140–149
Branco P, Amaral SD, Ferreira MT, Santos JM (2017) Do small barriers affect the movement of freshwater fish by increasing residency? Sci Total Environ 581:486–494. https://doi.org/10.1016/j.scitotenv.2016.12.156
Bravo-Córdoba FJ, Sanz-Ronda FJ, Ruiz-Legazpi J, Valbuena-Castro J, Makrakis S (2018) Vertical slot versus submerged notch with bottom orifice: Looking for the best technical fishway type for Mediterranean barbels. Ecol Eng 122:120–125. https://doi.org/10.1016/j.ecoleng.2018.07.019
Bunt CM, Castro-Santos T, Haro A (2012) Performance of fish passage structures at upstream barriers to migration. River Res Appl 28(4):457–478
Calluaud D, Pineau G, Texier A, David L (2014) Modification of vertical slot fishway flow with a supplementary cylinder. J Hydraul Res 52(5):614–629
Dingle H (2014) Migration. Oxford University Press, Oxford, The biology of life on the move, p 326
Illies J, Botoseanu L (1963) Problèmes et méthodes de la classification et de la-zonation écologique des eaux courantes, considérées surtout-du point de vue faunistique. SIL Commun 12:1–57. https://doi.org/10.1080/05384680.1963.11903811
IUCN (2018) The IUCN red list of threatened species (www.iucnredlist.org). [WWW Document]
Lucas MC, Baras E (2001) Migration of freshwater fishes. Blackwell Science, London, p 420
Nilsson C, Reidy CA, Dynesius M, Revenga C (2005) Fragmentation and flow regulation of the world’s large river systems. Science 308(5720):405–408. https://doi.org/10.1126/science.1107887
Noonan MJ, Grant JWA, Jackson CD (2012) A quantitative assessment of fish passage efficiency. Fish Fish 13(4):450–464
Peter A, Mettler R, Schölzel N (2016) Kurzbericht zum Vorprojekt “PIT-Tagging Untersuchungen am Hochrhein – Kraftwerk Rheinfelden”, p 45
Rosgen DL, Silvey HL (1996) Applied river morphology. Wildland Hydrology, Pagosa Springs, Colorado, USA
Tarrade L, Pineau G, Calluaud D, Texier A, David L, Larinier M (2011) Detailed experimental study of hydrodynamic turbulent flows generated in vertical slot fishways. J Environ Fluid Mech 11(1):1–21
Wang R, David L, Larinier M (2010) Contribution of experimental fluid mechanics to the design of vertical slot fish passes. Knowl Manag Aquat Ecosyst 396:02
Wootton RJ (1990) Ecology of teleost fishes. Chapman & Hall, London, p 404
Acknowledgments
We specifically thank Juan Carlos Romeral de la Puente (SAVASA) for the availability of the HPP facilities and for his continuous support, as well as Confederación Hidrográfica del Duero (Duero Water Authority) and Fishing Service of the regional government of Castilla y León for their legal and technical support.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2022 The Author(s)
About this chapter

Cite this chapter
Peter, A. et al. (2022). The Attractiveness of Fishways and Bypass Facilities. In: Rutschmann, P., et al. Novel Developments for Sustainable Hydropower. Springer, Cham. https://doi.org/10.1007/978-3-030-99138-8_5
Download citation
DOI: https://doi.org/10.1007/978-3-030-99138-8_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-99137-1
Online ISBN: 978-3-030-99138-8
eBook Packages: EngineeringEngineering (R0)