
Abstract
The context’s role in Pavlovian conditioning depends on the trial spacing during training, with massed trials revealing a function akin to that of discrete stimuli, and spaced trials revealing a modulatory function (Urcelay & Miller, Journal of Experimental Psychology. Animal Behavior Processes, 36, 268–280, 2010). Here we examined the contextual determinants of a common but largely ignored effect: attenuated conditioned responding with extended reinforced training (i.e., a postpeak performance deficit [PPD]). Contextual sources of PPDs were investigated in four fear-conditioning experiments with rats. In Experiment 1, as the number of reinforced trials increased, conditioned responding decreased, even when testing occurred outside the training context. Experiment 2 revealed opposing influences of context on the PPD based on trial spacing, which interacted with whether testing occurred in the training context. This finding reconciles Experiment 1’s results with previous data (Bouton, Frohardt, Sunsay, Waddell, & Morris, Journal of Experimental Psychology. Animal Behavior Processes, 34, 223–236, 2008). Experiment 3 suggested that extended training with these parameters did not lead to habituation to conditioned or unconditioned stimuli. In Experiment 4, few or many massed training trials were followed orthogonally by context extinction or no context extinction. After many pairings, context extinction reduced the PPD (i.e., enhanced responding), suggesting a competitive role of the context. These results, together with prior data suggesting that context modulates expressions of the PPD, are consistent with the view that contexts can play two distinctly different roles.
Similar content being viewed by others
Recently, a surge in interest has taken place concerning the nature of and the mechanisms underlying the processing of contextual information (e.g., Mesquita, Feldman Barrett, & Smith, 2010). Inherent to this quest is the observation that contexts sometimes compete with other stimuli or responses for behavioral control, and sometimes modulate the retrieval of memories (Urcelay & Miller, 2010). For example, Rescorla (1968) saw that rats were behaviorally influenced less by the predictive relationship between a signal and a brief footshock when additional, unsignaled footshocks were administered during the intertrial intervals (ITIs) than when unsignaled shocks were omitted. Presumably, this occurred because the context became associated with the footshock and competed with (i.e., blocked) the signal, thereby preventing high levels of responding to the signal. In contrast to Rescorla’s observation, Godden and Baddeley (1975) reported an experiment in which the test context facilitated the expression of learning. Diver subjects had to learn a list of 40 words either on land or underwater and were later tested in a context that either matched or did not match that of study. Memory recall was much better when subjects were tested in the same context in which they had studied, irrespective of whether the study context was on land or underwater. In this example, the context modulated the expression of learned behavior; that is, the context seems to have had a facilitative rather than a competitive role.
Although several functions of contextual information are widely acknowledged (Balsam, 1985; Bouton, 2010; Hall & Mondragón, 1998; Holland & Bouton, 1999; Miller & Schachtman, 1985; Urcelay & Miller, 2010), little is known about the conditions that favor one or the other of these functions being revealed. Gaining knowledge concerning these determinants is critical because, at a theoretical level, the two broad functions highlighted above (i.e., competitive vs. modulatory) have previously been treated separately, but rarely in an integrated fashion. For example, the Rescorla and Wagner (1972) model assumes that contexts play a competitive role similar to any other cue present in the experiment; thus, this model is poorly equipped to handle such phenomena as experimental extinction, which depends critically on the context of extinction for its expression (Bouton, 1993). In contrast, models that assume that contexts have a modulatory role on learning are better able to handle extinction and its recovery (i.e., interference effects; Bouton, 1993), but they have difficulties in explaining how contexts compete with discrete stimuli for behavioral control.
Studies demonstrating competition with the context have usually been conducted with short training regimens. With extended training, the picture becomes more difficult to understand, because contexts seem to engage in modulatory functions (Balsam, 1985). Despite the frequent absence of measurable differences between contexts in associative strength, contexts can differentially control the expression of excitatory conditioning in a powerful way (e.g., Balaz, Capra, Kasprow, & Miller, 1982). Given that all learning necessarily occurs in a particular environment, it is imperative to better understand the conditions that determine whether the context will compete with or modulate responding to discrete stimuli. In particular, we were interested in assessing the influence of contextual cues after extended training, which is known to result in a decrease from peak responding achieved with fewer trials (Pavlov, 1927). In a recent example of this postpeak performance deficit (PPD), using an aversively motivated task with rats as subjects, Bouton, Frohardt, Sunsay, Waddell, and Morris (2008) observed that with extended training, conditioned responding decreased in a context-dependent manner (i.e., the decrease was observed only in the training context), suggesting that the context played a modulatory role. Their interpretation of these findings was that subjects seem to develop “some kind of inhibition or adaptation process” that diminishes the effectiveness of footshock as an unconditioned stimulus (US) and that the habituation is context specific (Bouton et al., 2008, p. 234). Consistent with Bouton’s earlier view on how contexts function (1993; see also Spear, 1973, 1978), this interpretation of the PPD assumes that the training context has a modulatory effect on the expression of the overtrained conditioned stimulus (CS)–US association that underlies the PPD. In the absence of the contextual cues that were present during training, the PPD disappears at test (see also Pickens, Golden, Adams-Deutsch, Nair, & Shaham, 2009, for an extension of this interpretation to the effects of the passage of time, understood as change in the temporal context).
As mentioned above, contexts seem to be able to both impair control of behavior by a discrete cue (i.e., associative competition) and facilitate expression of memories concerning discrete cues, and these functions depend critically on the spacing of the training trials (Urcelay & Miller, 2010). In Bouton et al.’s (2008) experiments, trials during training were widely spaced (see below). It is not surprising, then, that these experiments revealed evidence favoring a modulatory function of the context. In contrast, other accounts of the PPD have assumed that it arises because the context competes with the target cue for behavioral control. Such is the case with the comparator hypothesis (Miller & Matzel, 1988), a model that assumes that the training context functions like a discrete stimulus, predicting that the PPD is caused by competition between the training context and the target stimulus. This model is silent concerning the modulatory role of the context. Within this framework, the competitive role of the training context can be inferred during tests of the target stimulus in a neutral context; otherwise, the putative associative strength of the context would be expected to summate with responding to the target stimulus. A consequence of this analysis is that, according to the comparator hypothesis, the PPD will be observed better when the target is tested outside rather than inside the training context, where it was observed by Bouton et al. (2008; see also Pickens et al., 2009).
In the present research, we sought to document the PPD effect in a lick suppression preparation and to determine whether this effect occurs when testing is conducted in an associatively neutral context (Exp. 1). In addition, because Experiment 1 revealed a PPD effect even when testing was conducted outside of the training context, we sought to assess the source of the discrepancy with Bouton et al.’s (2008) results by testing after PPD training in both the training context and a neutral context. Critically, we hypothesized that these opposite effects were caused by the use of different spacings of training trials (Urcelay & Milller, 2010), so that test context was orthogonal to training with massed trials (as in Exp. 1) or with widely spaced trials (approximating those used by Bouton et al., 2008, and Pickens et al., 2009). These two prior experiments agreed in suggesting that the PPD is context dependent, but they were constrained by the parameters used during training. Because a myriad of alternative explanations based on habituational processes might explain subsets of the data from Experiments 1 and 2, in Experiment 3 we directly tested whether in this paradigm the PPD results from CS or US habituation. In the absence of a suggestion that the present parameters led to CS or US habituation, in Experiment 4 we directly tested whether manipulating the associative strength of the context through extinction can alleviate the PPD.
Experiment 1
The objective of Experiment 1 was to document the basic PPD effect in our lick suppression preparation. In particular, we assessed whether this effect could be observed when testing was conducted outside the training context, a finding that would contrast with the results by Bouton et al. (2008) showing recovery from the PPD when the test is conducted outside of the context of PPD training. Four groups of rats received 5, 10, 20, or 50 pairings of a clicker with a mild footshock. All groups were trained using relatively massed trials, which result in higher context conditioning and facilitate the stimulus-like (i.e., competitive) function of the context (Urcelay & Miller, 2010). To equate the number of sessions and to preclude alternative explanations based on differing retention intervals between training and testing or different numbers of sessions (see Gottlieb, 2008, for a discussion), all subjects received five sessions of reinforced training. However, the different groups received different numbers of trials in each session (see Table 1). Each of our groups received one reinforced trial every 10 min (on average) of exposure to the training context.
Method
Subjects
The subjects were 24 female and 24 male experimentally naïve Sprague-Dawley young adult rats, bred in our colony. The mean body weights were 215 g (range 188–249 g) for females and 315 g (range 281–352 g) for males. Subjects were individually housed and maintained on a 16-h light/8-h dark cycle, with experimental sessions occurring roughly midway through the light portion. All subjects were handled for 30 s three times per week from weaning until the initiation of the study. Subjects had free access to food in the home cage. One week prior to initiation of the experiment, water availability was progressively reduced to 20 min per day, provided approximately 2 h after any scheduled treatment.
Apparatus
Six identical copies of each of two different types of experimental chambers were used. Chamber Rectangular (R) was a clear, Plexiglas, rectilinear chamber, measuring 23.0 × 8.5 × 12.5 cm (length × width × height). The floor was constructed of 0.48-cm-diameter stainless-steel rods, spaced 1.5 cm apart, center to center. The rods were connected by NE-2 neon bulbs that allowed a constant-current footshock to be delivered by means of a high-voltage AC circuit in series with a 1.0-MΩ resistor. Each copy of Chamber R was housed in a separate light- and sound-attenuating environmental isolation chest, which was dimly illuminated by a 2-W (nominal at 120 VAC) incandescent bulb driven at 60 VAC. The houselight was mounted on the ceiling of the environmental chest, approximately 26 cm from the center of the experimental chamber.
Chamber V-shaped (V) was a 22.1-cm-long box in a truncated V-shape (25.3 cm in height, 21.3 cm wide at the top, 5.1 cm wide at the bottom). The floor and sides were constructed of stainless steel sheets, and the ceiling was constructed of clear Plexiglas. The floor of each chamber consisted of two parallel metal plates, each 2.0 cm wide, with a 1.1-cm gap between them. Each V-shaped chamber was housed in its own environmental isolation chest, which was dimly illuminated by a 7.5-W (nominal at 120 VAC) incandescent houselight driven at 60 VAC mounted on an inside wall of the environmental chest, approximately 30 cm from the center of the experimental chamber. The light entering the animal chamber was primarily that reflected from the roof of the environmental chest, which was composed of white insulating material. The light intensities in the two types of chambers were approximately equal due to the differences in opaqueness of the walls in Chambers R and V.
Each chamber (R and V) could be equipped with a water-filled lick tube that extended 1 cm from the rear of a cylindrical niche, 4.5 cm in diameter, left–right centered in one short wall, with its axis perpendicular to the wall, and positioned with its center 4.25 cm above the floor of the chamber. Each niche had a horizontal infrared photobeam traversing it parallel to the wall on which the niche was mounted, 1 cm in front of the lick tube. In order to drink from the tube, subjects had to insert their heads into the niche, thereby breaking the infrared photobeam. Thus, we could record when subjects had their heads in the niche with the water tube. Ordinarily, they did this only when they were drinking. Disruption of ongoing drinking by a test stimulus served as our dependent variable.
Each chamber (R and V) was also equipped with three 45-Ω speakers mounted on the inside walls of the isolation chests. These speakers could deliver a complex tone 8 dB (C-scale) above background (76 dB, produced mainly by a ventilation fan), a click (6/s) 8 dB (C) above background, or a white-noise stimulus 8 dB (C scale) above background. In this experiment, only the clicks were used as the target stimulus (X). A 0.8-mA, 0.5-s footshock, which served as the US, could be delivered through the chamber floors. In Experiment 1, the CS was 30 s in duration.
Procedure
Subjects were randomly assigned to one of four groups based on the number of reinforced trials experienced during training: Group 5, Group 10, Group 20, and Group 50, counterbalanced for sex (ns = 12). Phase 1 was conducted in one (training) context (V or R, counterbalanced), and all other treatments in the remaining (test) context in a counterbalanced manner within groups. This was done to minimize differential fear of the test context summating with fear to the CS.
Acclimation
On Days 1 and 2, preexposure to the test context was conducted during daily 60-min sessions. Water-filled lick tubes were available during these sessions.
Phase 1
Prior to Phase 1, the lick tubes were removed. On Days 3–7, subjects in Group 5 received 1 X → US trial on each day during a daily 10-min session in the training chambers. Subjects in Group 10 experienced 2 X → US trials on each day, within a 20-min session. Subjects in Group 20 experienced 4 X → US trials on each day, within a 40-min session. Finally, subjects in Group 50 experienced 10 X → US trials on each day, within a 100-min session. Thus, the time spent inside the training context between USs was held constant. The overall mean ITI (from CS offset to CS onset) for all groups was 9.5 min. For all groups, the first trial occurred 5.5-min into the session, and the US was presented during the 0.5 s directly after termination of the CS. The order of treatment was rotated every day among the groups. On the first day of training, the running order was Groups 5, 10, 20, and 50. On the second day of training, the order was Groups 10, 20, 50, and 5. On the third day of training, the order was Groups 20, 50, 5, and 10. On the fourth day of training, the order was Groups 50, 5, 10, and 20. On the fifth day of training, the order was the same as on Day 3. This was done to avoid confounding the time of day when subjects were run with group assignment.
Reacclimation
On Days 8 and 9, the water lick tubes were reinserted, and subjects were allowed to drink during daily 60-min sessions in the test context. This treatment was intended to restabilize baseline levels of drinking. These sessions did not include any nominal stimulus presentations. The order of reacclimation was counterbalanced across groups.
Testing
On Day 10, with the lick tubes present in the test context, all subjects were tested for suppression to X. This was accomplished by presenting CS X immediately upon completion of the first five cumulative seconds of licking (as measured by the total amount of time the infrared photobeam was disrupted). Thus, all subjects were drinking at the time of CS onset. The time to complete this initial five cumulative seconds of licking (pre-CS scores) and the time to complete five additional seconds after the onset of the test CS (CS score) were recorded. Test sessions were 16 min in duration, with a ceiling score of 15 min being imposed on the time to complete five cumulative seconds of drinking in the presence of the test CS.
Data analysis
Latencies to initiate and resume drinking during the CS were transformed to log (base 10) scores to approximate a normal distribution of scores within groups, thereby facilitating the use of parametric statistics. An alpha level of .05 (two-tailed) was adopted for all statistical tests. Following the convention of our laboratory, subjects that took more than 60 s to complete their first five cumulative seconds of licking (i.e., prior to CS onset), thereby exhibiting an unusual reluctance to drink in the test context, were eliminated from all analyses. In this experiment, no subject met this criterion. The transformed data were analyzed with a one-way ANOVA on number of training trials, followed by planned comparisons that used the error term from the omnibus ANOVA. When appropriate, Cohen’s f is reported below to indicate effect size (Myers & Well, 2003, p. 210).
Results and discussion
As can be observed in Fig. 1, increasing the number of reinforced trials decreased suppression to the CS at test, as suggested by the lower latencies to resume drinking in the group that received 50 reinforced trials, even when testing was conducted outside of the training context (i.e., in an associatively neutral context). This observation is confirmed by the following analysis. A one-way ANOVA conducted on the pre-CS transformed latencies failed to detect an effect of group, F(3, 44) = 2.74, p > .05. A similar ANOVA conducted on the transformed latencies to resume drinking after the onset of the CS revealed differences among the groups, F(3, 44) = 3.69, MSE = 0.24, Cohen’s f = 0.41. Planned comparisons showed that the group that experienced the greater number of training trials (Group 50) suppressed less than did Group 5, F(1, 44) = 7.93, Group 10, F(1, 44) = 7.88, and Group 20, F(1, 44) = 6.10. Groups 5, 10, and 20 did not differ from each other, all ps > .70. Thus, this experiment showed that conditioned lick suppression with the present parameters is sufficiently sensitive to produce the PPD effect, even when testing is conducted outside of the training context.

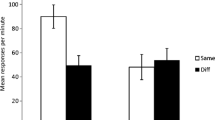
Mean times to complete five cumulative seconds of drinking upon presentation of the target CS in Experiment 1. See Table 1 for the treatments of the groups
Experiment 2
Superficially, the observations of Experiment 1 are inconsistent with the results of Bouton et al. (2008) and Pickens et al. (2009), because we observed a reliable PPD effect outside of the training context, whereas the results of Bouton et al. and Pickens et al. suggested that recovery from the PPD is observed when the test is conducted outside of the physical or temporal context of training. As mentioned in the introduction, Urcelay and Miller’s (2010) observations become relevant for the present discussion because a comparison of the parameters used in Experiment 1 with those used by Bouton et al. and Pickens et al. shows that they were indeed widely different. For example, in their experiment, Pickens et al. exposed subjects to the context for 1,440 min and to the stimulus for 50 min, which, according to a rough indicator of the relative spacing of trials (C/T; Gibbon & Balsam, 1981) results in a ratio of 28.8. Similarly, Bouton et al. exposed their subjects for a total of 1,740 min to the context and for 48 min to the stimulus, which results in a C/T ratio of 36.25 (note that in both experiments mentioned above, the instrumental baseline was trained in the same context in which cue conditioning occurred). In our Experiment 1, during training subjects in Group 50 experienced 500 min of exposure to the training context and 25 min of exposure to the CS, resulting in a C/T ratio of 20, which suggests that our parameters were more massed than those of Bouton et al. and Pickens et al.
In Experiment 2, we tested the view that the discrepancy between Experiment 1 and Bouton et al.’s (2008) data was driven by differences in our treatment of the context. We included a group that received relatively massed training and testing in both the acquisition context and an equivalently treated nontraining context. In addition, a separate group was similarly trained, but with spaced trials, as used by Bouton et al. and by Pickens et al. (2009). This design gets at the heart of the problem because, if there is an interaction between the spacing of training trials during the PPD and the context of test, both assumptions about the function of contextual information would be shown to be incorrect under select conditions, thereby substantiating the argument of Urcelay and Miller (2010) that both functions may coexist and selectively dominate behavior at extreme regions in the parametric space.
Each of two groups of rats received 50 reinforced trials with each of two stimuli (X and Y) in separate contexts. Group Massed experienced training with relatively massed trials similar to those used in Experiment 1 (9.5-min ITI). Group Spaced experienced training with more widely spaced trials (19.5-min ITI). Each stimulus was subsequently tested either in the training context (as Bouton et al., 2008, did) or in a different context (as we did in Exp. 1). The design of Experiment 2 is shown in Table 2.
Method
Subjects and apparatus
The subjects were 24 female and 24 male experimentally naïve Sprague-Dawley young adult rats, bred in our colony. The mean body weights were 196 g (range 173–217 g) for females and 292 g (range 215–338 g) for males. Subjects were housed, maintained, handled, and water deprived like those in the prior experiment. The apparatus was also the same as in Experiment 1. CSs X and Y were a 30-s click train and a 30-s high-frequency complex tone (i.e., an equal mix of two simple tones at 3000 and 3200 Hz), counterbalanced.
Procedure
The subjects were randomly assigned to one of two groups based on the spacing of trials during training (massed vs. spaced), counterbalanced for sex (ns = 12). The contexts (V and R) were counterbalanced within groups in the role of the training context, with the other context serving as the test context in Condition Different (see Table 2). During acclimation, reacclimation, and test, one quarter of the subjects in each group were run at a given time; during training, the massed and spaced groups were run separately because their sessions differed in duration.
Acclimation
Before the beginning of training, a 2-day preexposure to both contexts was conducted on Days 1 and 2. Subjects were exposed daily to each context (A and B) during 30-min sessions. Water-filled lick tubes were available during these two days, and the order of exposure was counterbalanced within groups and reversed from Day 1 to Day 2.
Phase 1
Prior to Phase 1, the lick tubes were removed from the training chambers. On Days 3, 5, 7, 9, and 11, all subjects experienced 10 daily X→US trials in Context A. Subjects in the massed group experienced these trials within a 100-min session, whereas subjects in the spaced group experienced these trials within a 200-min session. The overall mean ITIs (from CS termination to CS onset) for all trials were 9.5 min for Group Massed and 19.5 min for Group Spaced. The US was presented during the 0.5 s directly after termination of the CS. On Days 4, 6, 8, 10, and 12, subjects were exposed for a similar amount of time to the alternate context (B) and experienced 10 Y→US trials on each day with parameters similar to those used during the training of CS X.
Reacclimation
On Days 13, 14, and 15, the lick tubes were reinserted, and subjects were allowed to drink during daily 30-min sessions in each of the two contexts. This treatment was intended to restabilize baseline levels of drinking. These sessions did not include any nominal stimulus presentations.
Test
On Day 16, with the lick tubes inserted, all subjects were tested for suppression to CS X as in the previous experiment. Half of the subjects in each group were tested in the training context for X (A), whereas the remaining subjects were tested in the alternative context (B). Test sessions were 16 min in duration, with a ceiling score of 15 min being imposed on the time to complete five cumulative seconds of drinking in the presence of the test CS. Thus, all subjects received 15 min of exposure to CS X. On Day 17, subjects were tested for suppression to CS Y in the same context in which X had been tested. This allowed for the possibility of conducting a within-subjects comparison between testing in the same or in a different context, relative to the context in which the test CS had been trained. In other words, subjects that experienced testing of X in the same context during the first test experienced a test of Y in a different context from that in which Y had been trained (i.e., testing occurred in the same context where X had been trained). Subjects that experienced X in a different context during the first test were subsequently tested on Y in the same context (the context where Y had been trained).
Data analysis
The suppression data were transformed to log10 scores as in previous studies. The transformed data were analyzed first with a between-subjects ANOVA (including Order of Testing as a factor) and subsequently with a mixed ANOVA, with Trial Spacing (massed vs. spaced; between subjects) and Test Context (same vs. different; within subjects) as factors.
Results and discussion
The main results of Experiment 2 are shown in Fig. 2. Critically, there was a crossover interaction between the spacing of trials during training (massed vs. spaced) and the context of the test (same vs. different). Subjects that experienced training with parameters similar to those used in Experiment 1 (i.e., relatively massed training trials) revealed more responding inside the training context than outside, consistent with the view that these parameters endowed the context with associative properties akin those of a discrete cue. In addition, these results suggest that relatively massed training trials can still reveal modulation of cues by contexts in excitatory fear conditioning. That is, although both contexts had similar associative strength, subjects displayed more responding in the training context than in the alternative context. We will discuss this further in the General Discussion. However, when training was conducted with spaced trials, the opposite pattern emerged. Conditioned responding was lower after extended training when testing was conducted in the training context instead of in the alternative context, which is similar to the findings of Bouton et al. (2008) and Pickens et al. (2009). Presumably, widely spaced trials during training prevented the observation of context-dependent fear conditioning observed in the group that received relatively massed training trials, and instead revealed modulatory influences opposite to those observed with massed training. These impressions were supported by the following analysis.

Mean times to complete five cumulative seconds of drinking upon presentation of the target CSs X and Y in Experiment 2. Subjects received massed or spaced training and were tested in the same or a different context, relative to training. See Table 2 for the treatments of the groups
Two rats were eliminated from the study prior to testing due to poor health, and the data of two other rats were lost due to an experimenter error. A t test was used to compare Group Massed and Group Spaced with respect to their latencies for consuming water for five cumulative seconds after being placed in the test context and before CS onset on each day of testing. These two t tests failed to reveal differences on either day (both ts < 1.5), thereby indicating that there were no appreciable differences in baseline drinking and in fear of the two contexts. A 2 (same first vs. different first) × 2 (massed vs. spaced) × 2 (same vs. different) ANOVA on latencies to resume drinking in the presence of X and Y, with same versus different constituting a within-subjects variable, did not detect any effect of the order of testing (same first vs. different first), p > .85, or an interaction of order with any other variable, ps > .10. Consequently, the subsequent analyses of suppression to CS X and Y log latencies were collapsed across order of testing. A 2 (massed vs. spaced) × 2 (same vs. different) mixed ANOVA conducted on latencies to resume drinking in the presence of the test stimulus detected a crossover interaction, F(1, 42) = 13.56, Cohen’s f = 0.55, without detecting either a main effect of the context of testing (p > .63) or a main effect of trial spacing (p > .87).Footnote 1
Planned comparisons were conducted to determine the source of the critical two-way interaction and to test specific hypotheses. Suppression in Group Spaced was greater when testing was conducted in a different context, relative to the same context used in training, F(1, 42) = 8.63, which replicates Bouton et al.’s (2008) observation of an attenuated PPD effect when testing is conducted outside of the training context. In contrast, suppression in Group Massed was greater when testing was conducted in the training context (same) rather than in an alternative context (different), F(1, 42) = 5.15, which suggests positive modulation by the training context in the absence of differences in context–US associations. This observation is consistent with the results of Balaz, Capra, Hartl, and Miller (1981), in which superior conditioned responding was observed inside the training context (relative to an alternative context that had a similar associative history) in the absence of differences in conditioned responding to the contexts.
Experiment 3
The results of Experiment 1 indicated that testing in the training context is not necessary for the PPD to be observed. This is consistent with predictions based on the assumption that the context, acting like a discrete stimulus, can play a competitive role in the PPD. The results of Experiment 2 suggested that competition is most apt to occur with massed training trials, as the modulatory role of the training context on the cue is most apt to be observed when trials are spaced. Before directly extinguishing the context in order to assess whether the stimulus-like function observed in Experiment 1 is sensitive to direct manipulation of the associative strength of the context, we needed to corroborate that the PPD effect in the present protocol is not due to (1) habituation of the CS or the US, or (2) a generalization decrement resulting from extended training in one context and testing in a different context. According to a habituation account, subjects display the PPD because, due to the repetitive nature of extended reinforced training, they habituate to the CS (Ayres, Berger-Gross, Kohler, Mahoney, & Stone, 1979), to the US (Annau & Kamin, 1961), or to both, which results in attenuated responding to the CS during the test. For example, Bouton et al. (2008) interpreted their PPD effect in terms of context-dependent US habituation, because they observed that the PPD waned when they tested outside of the context in which it was trained. Although this interpretation is consistent with our observations in Experiment 2, particularly for Group Spaced, the opposite pattern was observed for Group Massed. In previous research, habituation has sometimes been found to be context specific (e.g., Archer & Sjödén, 1979; Evans & Hammond, 1983) and sometimes not (e.g., Hall & Channell, 1985; Hall & Honey, 1989; Marlin & Miller, 1981). Hence, these prior studies did not clearly discriminate between habituation-based explanations and an analysis that assumes cue-like properties for the context. Thus, we needed to assess whether parameters similar to those of Experiment 1 can result in a context-independent form of US habituation and whether this explains the PPD decrement when testing is conducted outside of the training context.
A second alternative account of Experiment 1 is that the decreased responding observed during testing outside the training context was the result of generalization decrement. According to this account, with extended training the target stimulus becomes configured with the context, and therefore removal of contextual elements by testing in an associatively neutral context results in decreased responding to the target. Thus, this explanation predicts that administering extensive training with a compound of two stimuli AX and testing one element (i.e., X) in a separate context should result in a similar, if not a larger, decrease in responding due to generalization decrement. In addition, habituation to auditory stimuli such as the CS in Experiments 1 and 2 has been observed to progress faster when the salience of the to-be-habituated stimulus is greater than the background noise (Davis, 1974). Consequently, if the PPD is due to habituation, it should be greater after training with a compound of cues AX. Therefore, according to both the generalization decrement and habituation accounts of the results of Experiment 1, conducting extended reinforced training with a compound of cues (AX) should increase the PPD. Similarly, if US habituation increases with the number of trials, extended training with a compound of discrete cues should result in a decrease in conditioned responding similar to that seen in Experiment 1 to one cue alone. This is predicted because, if the memory representation of the US is habituated (i.e., devalued) at the time of testing, responding should be relatively weak (Holland & Rescorla, 1975), regardless of how many cues were presented during training.
Alternatively, if the relatively massed training used in Experiment 1 made the context an effective competitor with the target cue at the time of testing, its effect should be weakened if a second stimulus is also presented during training. Using parameters that maximize the cue-like properties of the context, the PPD should be observed when only one cue is trained, but not when two cues are simultaneously trained. This assertion is based on previous research with cue competition phenomena. Given conventional spaced trials, recovery from both overshadowing (Bellingham & Gillette, 1981; Stout, Arcediano, Escobar, & Miller, 2003) and blocking (Azorlosa & Cicala, 1988) has been observed with extended training, which constitutes the opposite of a PPD effect. In addition, in the absence of extended training, overshadowing is not observed with highly massed training trials (i.e., short ITIs), regardless of whether the CSs are short (Stout, Chang, & Miller, 2003) or long (Sissons, Urcelay, & Miller, 2009). Thus, we next decided to test the prediction that extended training would result in recovery from another cue competition effect, overexpectation (Rescorla, 1970). In overexpectation, two cues are separately reinforced during a first phase, and in a second phase are reinforced together. Critically, in Experiment 3 we directly manipulated the number of trials during Phase 2 of training, which is the phase that establishes the overexpectation (i.e., the competition due to compound pairings). If the results of Experiment 1 were due to habituation-like effects, extended Phase 2 training in an overexpectation design using similar parameters should result in a similar, or even greater, response reduction, because habituation and overexpectation should summate. Similarly, if extended training results in less conditioned responding in an alternate context because of generalization decrement, conducting extended training with a compound of cues should reveal little conditioned responding to the target cue (X) after removal at test of the alternate stimulus (A) and the training context. However, if the PPD is due to context competition resulting from the use of relatively massed training trials, extended Phase 2 training with a compound cue AX should increase responding (i.e., overexpectation and the PPD should counteract each other; see the General Discussion for elaboration) because the nontarget discrete stimulus (i.e., A) and the context nullify each other in their capacity to compete with the target.
As depicted in Table 3, subjects were assigned to one of four groups, depending on the type of Phase 2 training (elemental [X] or compound [AX]) and on the number of trials received during this phase (12 or 60 trials). In order to assess the interaction between overexpectation and the PPD, we implemented some parametric variations from Experiment 1 that would better approximate the typical parameters for overexpectation. We modeled these parameters after Blaisdell, Denniston, and Miller (2001). In their experiments, Blaisdell et al. observed overexpectation when using 10-s cues and more trials (24) than the five trials used in Experiment 1. In order to make our parameters more similar to theirs, we used 10-s (instead of 30-s) cues and a slightly more intense footshock (1.0 mA). All subjects received 6 X–US and 6 A–US trials in Phase 1. During Phase 2, subjects experienced either 12 or 60 reinforced trials of X alone (elemental) or of an AX compound (overexpectation treatment).
Method
Subjects and apparatus
The subjects were 24 female and 24 male experimentally naïve Sprague-Dawley young adult rats, bred in our colony. The mean body weights were 220 g (range 196–275 g) for females and 324 g (range 296–363 g) for males. Subjects were housed, maintained, handled, and water deprived like those in the prior experiments. The apparatus was the same as in Experiment 1. CS X was a 10-s click train, and CS A was a 10-s high-frequency complex tone (i.e., an equal mix of two simple tones at 3000 and 3200 Hz).
Procedure
Subjects were randomly assigned to one of four groups based on the number of reinforced Phase 2 trials experienced during training (12 vs. 60) and the type of training in Phase 2 (elemental [X] vs. overexpectation [AX]). Group assignments were counterbalanced for sex (ns = 12). Phases 1 and 2 were conducted in one context (training), and all other treatments in the remaining context (test) in a counterbalanced manner within groups. Reacclimation, testing, and data analysis were conducted in the same manner as in Experiment 1.
Acclimation
Following Blaisdell et al. (2001), only 1 day of preexposure to the test context was conducted. Subjects were exposed to the testing chamber during a 60-min session with water lick tubes available.
Phase 1
Phases 1 and 2 were conducted with the lick tubes removed from the training chambers. On Days 2–4, all subjects were exposed to identical Phase 1 training, consisting of two reinforced trials of each X and A per daily 60-min session. A–US trials occurred 6 and 34 min into each Phase 1 session, whereas X–US trials occurred at 18 and 49 min into each Phase 1 session.
Phase 2
On Days 5–8, subjects in the “12” condition were exposed to three trials, at 6, 18, and 36 min into each daily 45-min session. Subjects in the “60” condition were exposed to 15 trials, at 6, 18, 36, 54, 67, 78, 96, 115, 130, 147, 159, 175, 187, 200, and 216 min into each daily 225-min session. The mean ITI for all trials in all groups was 15 min. Subjects in the AX condition were exposed to Phase 2 training consisting of reinforced AX trials (the number of which was dependent on group assignment [12 or 60]). Subjects in the X condition were exposed to training similar to the subjects in the AX condition, except that stimulus A was absent.
Results and discussion
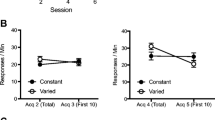
The results of Experiment 3 are shown in Fig. 3. In the “12” condition, a marginal decrement due to overexpectation training (AX) was observed relative to a control that experienced elemental reinforced training (X). This pattern was clearly reversed after extended Phase 2 pairings. The 60-X group showed a marked decline in behavioral control relative to the 12-X group, reflecting a strong PPD, which replicates that seen in Experiment 1 and extends it to somewhat different parameters (CS duration and number of trials). However, the subjects that experienced overexpectation training responded strongly to the target cue, even after many reinforced trials (Group 60-AX). Responding in this group increased with extended reinforced training, which is opposite to the pattern observed in Group 60-X. Thus, these results indicate that neither CS or US habituation nor generalization decrement due to testing in a different context can account for the results of Experiment 1.

Mean times to complete five cumulative seconds of drinking upon presentation of the target CSs in Experiment 3. Differential treatment began on Trial 13. See Table 3 for the treatments of the groups
No subject took more than 60 s to complete its first five cumulative seconds of licking prior to CS onset. A 2 × 2 factorial ANOVA with Amount of Training During Phase 2 (12 vs. 60) and Type of Training (elemental [X] vs. compound [AX]) as factors conducted on the pre-CS transformed latencies did not yield any main effects or interactions, all Fs < 1. A similar ANOVA conducted on the transformed latencies to resume drinking after presentation of the target CS revealed no main effects but a significant crossover interaction, F(1, 44) = 14.61, MSE = 0.07, Cohen’s f = 0.53. The interaction without significant main effects indicates that, as predicted, the two treatments (overexpectation and PPD training) had opposite effects on behavioral control by the target. Planned comparisons revealed a PPD effect indexed by more suppression in Group 12-X than in Group 60-X, F(1, 44) = 15.11. A comparison between the two groups that experienced few trials yielded a nonsignificant suggestion of an overexpectation effect, F(1, 44) = 2.44, p = .12. Importantly, compound conditioning during Phase 2 (overexpectation training) resulted in a reduction in the deficit observed with elemental training after many reinforced trials: Group 60-AX suppressed more than did Group 60-X, F(1, 44) = 14.77, p < .01, which suggests that extending reinforced training results in a decrement that is not due to generalization decrement or to habituation to the CS or US. Instead, it seems attributable to an associative interaction between the target cue and the training context. If the PPD resulted from generalization decrement or habituation, the opposite pattern should have been observed between Groups 60-X and 60-AX. That responding to X after extended compound training did not result in a PPD effect, as was observed with elemental training, is problematic for explanations based on habituation or generalization decrement processes.
Experiment 4
In Experiment 4, we sought further evidence regarding the competitive role of the context at the time of testing by directly manipulating the associative status of the training context after PPD training was completed. (In the General Discussion, we provide a theoretical basis for competition at the time of testing.) A simple prediction that follows from an account of the PPD based on the assumption that context competition between the target CS and the context occurs at the time of testing is that posttraining extinction of the training context should enhance behavioral control after 50 training trials, because such extinction should render the context unable to compete with the CS for behavioral control at test. Note that this prediction is conceptually similar to the recovery from cue competition effects such as overshadowing (e.g., Kaufman & Bolles, 1981) that is observed after extinction of the overshadowing cue; this effect is now often called retrospective revaluation. In Experiment 4, we aimed to replicate the PPD effect of Experiments 1 and 3 and to directly test this prediction. In addition, we tested the specificity of this posttraining manipulation by including two additional groups that only received five training trials. If recovery from the PPD after context extinction is due to some nonspecific increase in responding or to the mere passage of time, we should also observe an increase in responding after few training trials. We routinely observe greater suppression in our preparation than the levels of suppression observed in Group 5 in Experiment 1: Thus, our procedure should be sensitive to such effects. However, if the PPD that arises with massed training depends on the progressive engagement of the context with extended training, an increasing stimulus control as a result of context extinction should only be observed in the group that experienced the PPD training (50 trials), but not after minimal training (five trials).
We used a factorial design in which subjects experienced either five or 50 reinforced trials using the same procedure as in Experiment 1, and were orthogonally assigned to minimal (20 min) or extensive (600 min) posttraining context extinction (see Table 4). With minimal extinction, we anticipated decreased behavioral control after 50 reinforced trials relative to five reinforced trials (i.e., a PPD effect). In the groups that received analogous treatment but experienced extensive context extinction, we expected an increase in behavioral control after both five and 50 training trials. However, the recovery should be larger after 50 than after five reinforced trials, because the influence of context–US associations should be appreciably weaker in the five-trial condition, which would leave little room for extinction to alter conditioned responding.
Method
Subjects and apparatus
The subjects were 24 female and 24 male experimentally naïve Sprague-Dawley young adult rats, bred in our colony. The mean body weights were 209 g (range 186–240 g) for females and 307 g (range 267–340 g) for males. The subjects were housed, maintained, handled, and water deprived as those in previous experiments. The apparatus was the same as in Experiments 1 and 3.
Procedure
Subjects were randomly assigned to one of four groups (ns = 12) based on the number of reinforced trials experienced during training (5 vs. 50) and the posttraining context extinction (control [C] vs. extinction [E]) treatment. Phases 1 and 2 were conducted in one context (training) and all other treatments in the remaining context (test) in a counterbalanced manner within groups. Acclimation, reacclimation, testing, and data analysis were conducted in exactly the same manner as in Experiment 1.
Phase 1
Phases 1 and 2 were conducted with the lick tubes removed from the training chambers. On Days 3–8 (with the exception of Day 6, on which no treatment occurred due to a power failure), during daily 10-min sessions, subjects in Groups 5C and 5E received 1 X → US trials on each day. Subjects in Groups 50C and 50E experienced 10 X → US trials on each day, within a 100-min session. All other details were similar to those of Experiment 1.
Phase 2
During Days 9–12, subjects experienced simple exposure to the training context. This treatment was expected to reduce the strength of the context–US association, and perhaps of the CS–context association; therefore, the critical manipulation was the amount of exposure to the training context. Subjects in Groups 5E and 50E (the extinction groups) were exposed daily for 150 min to the training context. Every 30 min, the sound-attenuating enclosures were opened to ensure that the subjects were awake. Subjects in the control condition (Groups 5C and 50C) experienced one daily 5-min exposure to the training context in order to equate subjects for overall handling and retention interval between training and testing. These 5 min of exposure were not expected to have a large impact on the PPD, on the basis of prior research in our laboratory (Schachtman, Brown, Gordon, Catterson, & Miller, 1987) that indicated that at least 480 min of context extinction is necessary for observing retrospective revaluation based on posttraining context extinction.
Results and discussion
The results of Experiment 4 are depicted in Fig. 4. The two groups that received minimal exposure to the training context (control condition) behaved similarly to their counterparts in Experiments 1 and 3. That is, extending the number of reinforced trials decreased behavioral control by the CS. However, massive posttraining exposure to the training context alone attenuated this difference, and did so selectively in subjects that received extended training, not in subjects that received minimal training. These observations were confirmed by the following analyses.

Mean times to complete five cumulative seconds of drinking upon presentation of the target CS in Experiment 4. See Table 4 for the treatments of the groups
One subject from Group 5C was excluded because of an equipment malfunction on the test day. No subject took more than 60 s to complete its first five cumulative seconds of licking on the test day. A 2 × 2 factorial ANOVA with Amount of Training (5 vs. 50 trials) and Training Context Extinction (control [C] vs. extinction [E]) as the main factors conducted on the pre-CS scores revealed no main effects or interaction (largest F < 1), suggesting no baseline differences between the groups in generalized fear of the test context. A similar ANOVA conducted on the transformed latencies in the presence of the CS revealed a main effect of amount of training, F(1, 43) = 9.72, Cohen’s f = 0.43, a main effect of extinction, F(1, 43) = 17.99, Cohen’s f = 0.60, and an interaction, F(1, 43) = 6.80, MSE = 0.13, Cohen’s f = 0.35. Planned comparisons revealed that Group 50C suppressed drinking less than did Group 5C, F(1, 43) = 16.03, replicating the central result of Experiment 1. As anticipated on the basis that the PPD arises from competition with the context at the time of testing, massive context extinction attenuated the PPD. That is, Group 50E suppressed more than did Group 50C, F(1, 43) = 23.99. A smaller effect of context extinction was expected after only five training trials, and the direction of the ordinal difference between the means was consistent with this expectation. Although responding was numerically stronger in Group 5E than in Group 5C, this difference did not achieve statistical significance, F(1, 43) = 1.30, p = .25.
The results of Experiment 4 add to those of Experiments 1 and 3 in suggesting that, at least with the present parameters, the PPD can be observed when testing is conducted outside of the training context, and that manipulations that presumably affect the associative status of the training context (such as the presentation of another discrete stimulus during training, which competes with the context’s potential, or direct extinction of the training context) can reverse the PPD.
General discussion
The present experiments investigated contextual determinants of the PPD, which is a decrement in behavioral control that arises with extended pairings of a single CS with a US. Although this effect has now been documented in a wide range of preparations, most theories do not account for it, and those that do so appeal to a diverse array of mechanisms. In particular, we were interested in two families of explanations of the PPD that assume that the training context is a critical determinant of the PPD, but for different reasons. Thus, following Urcelay and Miller (2010), we hypothesized that at least two different roles of the context may be engaged in this phenomenon, and that the spacing of trials during training is critical in engaging one role or the other. Moreover, the contextual role engaged by massed or spaced training trials determines the expression of the PPD’s context dependency. Below, we summarize the main results of these experiments and discuss them in the light of different theoretical approaches.
We first sought to determine whether the PPD can be observed when testing is conducted outside of the training context, as anticipated by the sometimes-competing-retrieval model (SOCR; Stout & Miller, 2007), which accounts for the PPD by assuming that the training context plays a role akin to a discrete stimulus (see below). In Experiment 1, we documented the basic PPD effect while conducting training with relatively massed trials and testing outside of the training context. Although this result was consistent with an explanation of the PPD that attributes to the training context a stimulus-like function, we noted that recent findings in other laboratories have documented an effect opposite the one we found in Experiment 1: namely, that the PPD is best observed when tests are conducted in the training context. In Experiment 2, we trained two groups with either short spacing of trials, as in Experiment 1, or widely spaced trials, which were used in the studies by Bouton and colleagues (2008; see also Pickens et al., 2009) in which opposite findings were observed. To test for a possible interaction between the spacing of training trials and the test location, we tested each group inside and outside the context of training. Consistent with both the findings of Experiment 1 and those opposite to our findings, we found an interaction between the spacing of training trials and the testing location. These two experiments, taken together, provide a satisfactory account of these diverse findings. Because habituation-like processes and generalization decrement offer alternative explanations of portions of the observations in Experiments 1 and 2, in Experiment 3 we tested whether the PPD would also decrease conditioned responding to a stimulus that underwent extended compound (i.e., overexpectation) training. The results of Experiment 3 agree with recent reports (Garfield & McNally, 2009) in showing that, contrary to what we observed with extensive training of a single CS (Exps. 1 and 2), extensive reinforcement of the target CS in compound with another CS does not result in the PPD, but rather in recovery from the cue competition observed with few training trials (see the introduction to Exp. 3). Finally, assuming that the context can function as a CS to compete with discrete CSs, in Experiment 4 we tested whether the PPD effect (with relatively massed training) is sensitive to changes in the associative status of the training context, achieved through an extinction treatment. Consistent with a stimulus-like assumption concerning the role of the context, massive posttraining extinction of the training context resulted in recovery from the PPD.
Although the assumption that the context functions as a CS fares well in explaining the results of Experiments 1, 3, and 4, we noted in the introduction that it fails to anticipate recent demonstrations that the PPD is attenuated when testing occurs outside of the training context, relative to when testing occurs in the training context (Bouton et al., 2008; Pickens et al., 2009). To understand these discrepant findings, we appealed to recent experiments in Pavlovian fear conditioning that have suggested that only with relatively massed training trials does the context compete for behavioral control with the target cue. When trials are spaced, the competitive influence wanes, and a modulatory (i.e., occasion-setting) role of the context is revealed (Urcelay & Miller, 2010). This view assumes that the training context competes with the target and causes the PPD when trials are massed. When trials are spaced, the PPD may occur because of error-driven loss of attention (Pearce & Hall, 1980; see Kaye & Pearce, 1984, for evidence consistent with this mechanism, in particular after extended reinforced training), and the context should serve to modulate the CS–US association that results in the PPD.Footnote 2 Consistent with this theory, in Experiment 2 we observed that both the spacing of trials during extended reinforced training and the context of testing (i.e., the training context or outside the training context) are critical determinants of the expression of the PPD. That is, if training is conducted with relatively massed trials (Group Massed in Exp. 2), the training context has a moderately high associative value and seems to compete with the target CS–US association for behavioral control. This competition by the context is best revealed when testing is conducted outside the target training context; otherwise, responding based on the context–US association may summate with responding to the target CS. In contrast, spacing the training trials revealed a different role of the context. Presumably, with extended training and the use of spaced trials, the CS-evoked conditioned responding decreases as a result of extended trainingFootnote 3 in a context-dependent manner, in which case testing in the training context reveals the PPD, but testing outside the training context results in recovery from the PPD (Bouton et al., 2008).
Contexts have been hypothesized to play several, fundamentally different roles in associative learning (Balsam, 1985; Bouton, 2010; Holland & Bouton, 1999; Miller & Schachtman, 1985), but theories of learning have usually adopted one of two general functions. Some theories assume that contexts function in a way that is similar to any other discrete stimulus (Gallistel & Gibbon, 2000; Gibbon & Balsam, 1981; Mackintosh, 1975; Pearce & Hall, 1980; Rescorla & Wagner, 1972; Solomon & Corbit, 1974). For example, opponent-processes theory (OPT; Solomon & Corbit, 1974; also see Schull, 1979) and Wagner’s (1981) SOP model both offer a plausible account of the PPD that is based on cue-like properties of the context, but neither fully explains the results observed in Experiments 1–4. By different processes, both predict that the training context progressively decreases responding to the target stimulus; thus, both predict that the PPD should only be observed inside the context of training, not outside. According to OPT, conditioned responding results from the interaction between two processes, a and b, that act in opposition. The a process represents the reaction to the presentation of the US, has a fast response, and is a monotonic function of the US intensity. The b process, on the contrary, is a sluggish process evoked by the a process that is inhibitory to the reaction evoked by the a process. With extended training, the b process decreases in latency of onset and increases in intensity and duration, resulting in a progressive cancellation of the a process. Responding (or the emotional reaction to the US, in Solomon and Corbit’s language) is the algebraic summation of the two processes, and therefore decreases with extended training. However, the b process is triggered not only by the a process activated by the CS, but also by the a process activated by the context as a result of context–US associations (Solomon & Corbit, 1974, p. 136). SOP anticipates the development of the PPD inside the training context because, according to this model, the context becomes associated with the CS with repeated exposures, and therefore subsequently retrieves a representation of the CS into state A2. This retrieved representation of the CS in A2 precludes activation of the CS into A1, thereby decreasing activation of the US representation into A2, and consequently decreasing responding. In Experiments 1, 3, and 4 we tested the target CS in a second, neutral context and in the absence of US presentations, so the PPD effect would not be anticipated by either OPT or SOP. That is, in the absence of the training context, the b process (or activation of the CS into A2, according to SOP) should not have been anticipatory to CS onset. Of note, we used highly distinct contexts that extensive prior research in our laboratory has demonstrated are well discriminated by rats (e.g., Laborda, Witnauer, & Miller, 2011; Schachtman et al., 1987). OPT and SOP predict the PPD effect only if testing is conducted in the same context where training was conducted, a prediction that received support in Experiment 2, but only when the spacing of trials during training was large. The pattern of results that resembles OPT’s and SOP’s predictions only occurred in the spaced-trial condition, a finding that is challenging for these models because they predict that the context dependency of the PPD should be better observed with massed training trials. Thus, although these models explain parts of the data, they do not provide a full account of the findings across all four experiments.
According to a habituation account of the PPD, with extended training the subjects’ sensitivity to CS or US presentations decreases, and consequently, responding during test is reduced. In general, there are few grounds on which to suggest that, with the use of the present parameters, the data from these experiments are explicable in terms of habituation. First, habituation has been observed to be context specific (Archer & Sjödén, 1979; Evans & Hammond, 1983; but see Hall & Channell, 1985; Hall & Honey, 1989; Marlin & Miller, 1981), and in Experiments 1, 3, and 4 we conducted testing in a different context than had been used for training. Second, and more important for the present discussion, habituation usually does not decrease with massive extinction of the context as administered in Experiment 4, which resulted in a recovery from the PPD. In fact, Marlin and Miller (1981, Exp. 4; see also Baker & Mercier, 1982; Hall & Minor, 1984) saw increases in habituation after extinction of the context where habituation had previously been administered. However, Jordan, Strasser, and McHale (2000) found the opposite effect using lick suppression, which is the same dependent measure used here, so we could not categorically reject the possibility that habituation to the CS was responsible for the PPD we observed in Experiment 1. In addition, the results of Experiment 1 (and the recovery observed in Exp. 4 after context extinction) can be also accommodated by an explanation in terms of decreases in stimulus effectiveness with increased exposure to the stimulus (Hall, 2003), which is recovered after being associatively activated during context extinction (Exp. 4).
Experiment 3 was designed to address the possibility that the PPD resulted from habituation to the CS or from generalization decrement (see the introduction to Exp. 3) due to testing in a different context, by investigating the PPD in a cue competition procedure. On the basis of SOCR (Stout & Miller, 2007) and of previous data suggesting that cue competition effects such as overshadowing (Bellingham & Gillette, 1981; Stout, Arcediano, et al., 2003) and blocking (Azorlosa & Cicala, 1988) are attenuated with extended training, we investigated whether overexpectation, another cue competition effect, would show a similar recovery after extended training. We observed a rather weak overexpectation effect with few trials but, importantly, an increase (rather than a decrease) in responding with extended training. In other words, we observed the PPD with elemental training but not after compound (overexpectation) training. The loss of the PPD with compound training agrees with a recent report in which different amounts of Phase 2 overexpectation were assessed using freezing as dependent measure (Garfield & McNally, 2009). Thus, the results of Experiment 3 allowed us to reject, at least with rather massed training parameters, the view that the PPD observed in Experiment 1 was due to habituation to the CS or the US. A second purpose of Experiment 3 was to test the plausibility of an explanation of Experiment 1 in terms of generalization decrement. That is, the low responding after PPD training observed in Experiment 1 could be due to the context change subtracting stimulus attributes from the X–context compound that received extensive training to testing X alone. If this change in stimulus during the transition from training to test explained the low responding observed in Experiment 1 after PPD training, a similar (or larger) decrease should be observed when the PPD training was conducted with a compound of cues, as was the case with overexpectation treatment. Because the opposite effect was observed (administering PPD training with a compound AX and testing X in a different context resulted in increased rather than decreased responding to the target), we concluded that generalization decrement is unlikely to explain the pattern of results we observed across these experiments (see also Exp. 2).
Taken together, Experiments 1, 3, and 4 suggest that an important determinant of the PPD is the training context, which competes with the target cue for control of behavior, as is predicted by SOCR (Stout & Miller, 2007), which assumes that the context functions like a discrete stimulus to compete with the target stimulus. Figure 5 depicts a comparison of the results of all groups observed in Experiments 1, 3, and 4 with simulations of SOCR, Stout and Miller’s mathematical implementation of the extended comparator hypothesis. We only present simulations of Experiments 1, 3, and 4 here because these experiments were all inspired by the predictions of SOCR, on the assumption that the context plays a role akin to a discrete stimulus when trials are relatively massed. In these simulations, we used a hill-climbing procedure to obtain the best-fitting parameters that would account for all 12 groups in Experiments 1, 3, and 4. This procedure is widely used in cognitive science (e.g., Denton, Kruschke, & Erickson, 2008; Kruschke, 2006; Smith & Minda, 1998). SOCR’s statistical fit was assessed by computing the sum of the squared difference between predicted and observed group means (i.e., the sum of squared errors [SSE]) across all 12 groups in these experiments. The set of parameters that produced the lowest SSE was determined using a hill-climbing algorithm. The hill-climbing algorithm searched the set of parameter configurations immediately adjacent to the previous best-fitting set of parameters (or the starting set of parameters), ultimately moving through parameter space in the direction of steepest descent on SSE. The program was terminated when it failed to find an immediately adjacent configuration of parameters that produced lower SSE. The hill-climbing algorithm was repeated three times with three pseudorandom starting points in parameter space, with the constraint that they be far apart. We report the results of the simulation that produced the lowest SSE. In order to make the simulations readily comparable with the log10 latency to consume water for five cumulative seconds in the presence of the test CS, we used a scaling procedure that would allow for direct comparisons between the observed and simulated data. After Larrauri and Schmajuk (2008), we assumed that lick suppression is proportional to the response potential of X (RX), as predicted by SOCR. In addition, we assumed that the lick suppression predicted by such proportional scaling is added to the scale’s minimum [i.e., log10(5), where 5 s was the minimum time necessary to consume water for 5 s—that is, no suppression]. Thus, in the present simulations, we approximated the lick suppression (LS) scale via the following equation:
where k 4 is a free parameter used to approximate the log10 latency scale measured in our lick suppression situation. The final parameters used in all these simulations were k 1 = 0.05, k 2 = 1, and k 3 = 1. The salience of the target X was 0.65, the salience of A was 1, that of the context was 0.1, and that of the US was 0.5. As can be observed in Fig. 5, SOCR’s simulations provide a relatively accurate description of the results that we observed across all three experiments. More importantly, SOCR predicts all of the critical ordinal differences in these three experiments.

Simulations and the results of Experiments 1, 3, and 4 using SOCR with parameters that were obtained using a hill-climbing procedure and scaled to a log-second scale for comparisons with the actual data (see the text for details)
Although SOCR provides a good fit of Experiments 1, 3, and 4, the pattern of results observed in Experiment 2 goes beyond the predictions of the model. Experiment 2 contrasted one acquisition variable (spacing of trials) with one test variable (same vs. different) and revealed a crossover interaction between these variables. This pattern is problematic for current theories of learning. This is perhaps due to the fact that, to date, theories of learning have assumed only one or the other of the two different roles about contextual information; in fact, most theories have assumed a competitive role for contextual information. This assumption about contextual information has allowed these models to correctly predict the detrimental effects of trial massing, US preexposure, and contingency degradation treatments, among other treatments. In contrast, other theories have assumed a modulatory role for the context on memory retrieval. Models derived from the human literature, such as Tulving’s encoding specificity principle (Tulving & Thomson, 1973), and analogues based on animal research (Bouton, 1993; Spear, 1973, 1978) have assumed that the context is a retrieval aid for memory (i.e., a memory modulator). In recent experiments, Urcelay and Miller (2010) revealed one variable that determines which of these two functions of the context will be most pronounced by manipulating the spacing of the training trials. The present Experiment 2 extends these findings by demonstrating these two functions of the context, which oppositely depend on the spacing of trials during PPD training. When the PPD was trained with moderately massed trials, testing outside the training context revealed the PPD, but the opposite result was observed when training was conducted with rather spaced trials. Thus, the notion that contexts can play fundamentally different roles depending on certain parameters (as indicated by the crossover interaction observed in Exp. 2; see also Exp. 1 of Urcelay & Miller, 2010) represents a new challenge that theories of learning need to embrace.
Experiment 2 suggests that, as Bouton et al. (2008) have observed, in certain circumstances (such as when training is conducted with widely spaced trials) the PPD is observed when the test is conducted in the training context, but not outside the training context. This is consistent with an account of the PPD in terms of context-specific processes that are likely to arise when widely spaced trials are used during training. Perhaps the greatest value of Experiment 2 is that it identifies one parameter (i.e., the spacing of trials) that determines the ordinal relationship in conditioned responding between testing in the same context as training and in a different context. This pattern of results is certainly challenging for most of the theories of which we are aware, because they anticipate either no PPD effect, a stronger PPD in the training context (e.g., Solomon & Corbit, 1974; Wagner, 1981), or a stronger PPD outside of the training context (Stout & Miller, 2007). The results of Experiment 2 also shed light on a longstanding issue in the Pavlovian conditioning literature: whether a change in context between training and testing enhances or decreases conditioned responding to a stimulus. Whereas some researchers have observed a decrease in behavioral control with a change in context from that of training (Balaz et al., 1981; Hall & Honey, 1990), others have observed increases in behavioral control after changes in context (Bouton et al., 2008; Kaye & Mackintosh, 1990). Our Experiment 2 revealed that both outcomes can be observed, and it isolated one variable that critically determines which outcome will occur. Overall, the pattern observed in Experiment 2 complements recent findings (Urcelay & Miller, 2010) that have suggested that contexts can have different (sometimes opposing) influences on learned behavior, depending on the spacing of trials used during training.
Finally, we wish to consider whether the competitive and modulatory roles of contexts are mutually exclusive or additive. That is, the variable that we identified, amount of context exposure, that determines which role the context plays suggests that these two roles represent the extreme portions of a relative temporal dimension. In Experiment 2, after massed training we tested in the training context or in a different context, but importantly, the two contexts were equal in their associative strength. If the use of relatively massed trials causes the context to function like a competing stimulus at the expense of any modulation, then responding should have been equal, independent of the context of test, given the within-subjects nature of the experiment (i.e., the associative status of the context was matched within groups). The observation that responding was differential despite equal associative strengths suggests that the modulatory function adds to the associative function, because the modulatory function is observed even when the associative status of the context is high. This conclusion is logically consistent with the conclusions reached by Balaz and colleagues (1981; Balaz et al., 1982). In their experiments, modulation was observed after context preexposure or extinction, in the absence of summation between context and CS associations. Further studies will surely be needed to clarify these relationships. What we can conclude with confidence, on the basis of the present results and those of Urcelay and Miller (2010), is that (1) contexts can engage in at least two functions and (2) the conditions of training, such as the spacing of training trials, have an important role in determining which function, if any, will be prevalent in controlling behavior.
Notes
An additional 2 (massed vs. spaced) × 2 (same vs. different) between-subjects analysis conducted on the first test alone also revealed a crossover interaction (p < .05) in the absence of main effects (ps > .34), suggesting that the results presented did not derive from the within-subjects nature of the analysis, making these results comparable to those of Experiments 1–3, in which only between-subjects designs were used.
This explanation is clearly speculative. However, we favor it here, given that we found no evidence of habituation with these parameters.
The precise mechanism of this context-dependent PPD observed with widely spaced training trials is unclear. Above, we theorized that the mechanism is error-driven decreases in attention, as proposed by the Pearce–Hall (1980) model. Bouton and colleagues have referred to “adaptation,” “inhibition,” and “habituation.” The crossover interactions observed in Experiments 2 and 3 challenge a purely “nonassociative” habituation explanation, or at least suggest that, if these results do arise from habituation, we currently understand little about its mechanisms. In addition, Experiment 2 revealed higher responding after PPD training with a spacing of trials similar to that of Experiment 1 (Group Massed) when testing was conducted inside the training context, which strongly argues against an explanation of the PPD in terms of habituation.
A role for inhibition is difficult to capture formally (or intuitively) because the CS and US were always paired, and therefore a situation in which an expectation was not fulfilled (e.g., negative contingency) was never operationally met. Rosas, Callejas Aguilera, Ramos Álvarez, and Fernández Abad (2006) proposed an attentional theory of context processing that builds on Bouton’s (1993), although the attentional mechanism that we have speculated about here is not directed toward the context, but rather to the cue.
References
Annau, Z., & Kamin, L. J. (1961). The conditioned emotional response as a function of intensity of the US. Journal of Comparative and Physiological Psychology, 54, 428–432.
Archer, T., & Sjödén, P. (1979). Neophobia in taste-aversion conditioning: Individual differences and effects of contextual changes. Physiological Psychology, 7, 364–369.
Ayres, J. J. B., Berger-Gross, P., Kohler, E. A., Mahoney, W. J., & Stone, S. (1979). Some orderly nonmonotonicities in the trial-by-trial acquisition of conditioned suppression: Inhibition with reinforcement? Animal Learning & Behavior, 7, 174–180.
Azorlosa, J. L., & Cicala, G. A. (1988). Increased conditioning in rats to a blocked CS after the first compound trial. Bulletin of the Psychonomic Society, 26, 254–257.
Baker, A. G., & Mercier, P. (1982). Extinction of the context and latent inhibition. Learning and Motivation, 13, 391–416.
Balaz, M. A., Capra, S., Hartl, P., & Miller, R. R. (1981). Contextual potentiation of acquired behavior after devaluing direct context–US associations. Learning and Motivation, 12, 383–397.
Balaz, M. A., Capra, S., Kasprow, W. J., & Miller, R. R. (1982). Latent inhibition of the conditioning context: Further evidence of contextual potentiation of retrieval in the absence of context–US associations. Animal Learning & Behavior, 10, 242–248.
Balsam, P. D. (1985). The functions of context in learning and performance. In P. D. Balsam & A. Tomie (Eds.), Context and learning (pp. 1–21). Hillsdale: Erlbaum.
Bellingham, W. P., & Gillette, K. (1981). Attenuation of overshadowing as a function of nondifferential compound conditioning trials. Bulletin of the Psychonomic Society, 18, 218–220.
Blaisdell, A. P., Denniston, J. C., & Miller, R. R. (2001). Recovery from the overexpectation effect: Contrasting performance-focused and acquisition-focused models of retrospective reevaluation. Animal Learning & Behavior, 29, 367–380.
Bouton, M. E. (1993). Context, time, and memory retrieval in the interference paradigms of Pavlovian learning. Psychological Bulletin, 114, 80–99. doi:10.1037/0033-2909.114.1.80
Bouton, M. (2010). The multiple forms of “context” in associative learning theory. In B. Mesquita, L. Feldman Barrett, & E. R. Smith (Eds.), The mind in context (pp. 233–258). New York: Guilford Press.
Bouton, M., Frohardt, R., Sunsay, C., Waddell, J., & Morris, R. (2008). Contextual control of inhibition with reinforcement: Adaptation and timing mechanisms. Journal of Experimental Psychology. Animal Behavior Processes, 34, 223–236.
Davis, M. (1974). Signal-to-noise ratio as a predictor of startle amplitude and habituation in the rat. Journal of Comparative and Physiological Psychology, 86, 812–825.
Denton, S. E., Kruschke, J. K., & Erickson, M. A. (2008). Rule-based extrapolation: A continuing challenge for exemplar models. Psychonomic Bulletin & Review, 15, 780–786. doi:10.3758/PBR.15.4.780
Evans, J., & Hammond, G. (1983). Differential generalization of habituation across contexts as a function of stimulus significance. Animal Learning & Behavior, 11, 431–434.
Gallistel, C. R., & Gibbon, J. (2000). Time, rate, and conditioning. Psychological Review, 107, 289–344. doi:10.1037/0033-295X.107.2.289
Garfield, J. B. B., & McNally, G. P. (2009). The effects of FG7142 on overexpectation of Pavlovian fear conditioning. Behavioral Neuroscience, 123, 75–85.
Gibbon, J., & Balsam, P. (1981). Spreading association in time. In C. M. Locurto, H. S. Terrace, & J. Gibbon (Eds.), Autoshaping and conditioning theory (pp. 219–253). New York: Academic Press.
Godden, D. R., & Baddeley, A. D. (1975). Context-dependent memory in two natural environments: On land and underwater. British Journal of Psychology, 66, 325–331.
Gottlieb, D. A. (2008). Is the number of trials a primary determinant of conditioned responding? Journal of Experimental Psychology. Animal Behavior Processes, 34, 185–201.
Hall, G. (2003). Learned changes in the sensitivity of stimulus representations: Associative and nonassociative mechanisms. Quarterly Journal of Experimental Psychology, 56B, 43–55.
Hall, G., & Channell, S. (1985). Differential effects of contextual change on latent inhibition and on the habituation of an orienting response. Journal of Experimental Psychology. Animal Behavior Processes, 11, 470–481.
Hall, G., & Honey, R. C. (1989). Contextual effects in conditioning, latent inhibition, and habituation: Associative and retrieval functions of contextual cues. Journal of Experimental Psychology. Animal Behavior Processes, 15, 232–241. doi:10.1037/0097-7403.15.3.232
Hall, G., & Honey, R. C. (1990). Context-specific conditioning in the conditioned-emotional-response procedure. Journal of Experimental Psychology. Animal Behavior Processes, 16, 271–278. doi:10.1037/0097-7403.16.3.271
Hall, G., & Minor, H. (1984). A search for context–stimulus associations in latent inhibition. Quarterly Journal of Experimental Psychology, 36B, 145–169.
Hall, G., & Mondragón, E. (1998). Contextual control as occasion setting. In N. A. Schmajuk & P. C. Holland (Eds.), Occasion setting: Associative learning and cognition in animals (pp. 199–222). Washington, DC: American Psychological Association. doi:10.1037/10298-007
Holland, P. C., & Bouton, M. E. (1999). Hippocampus and context in classical conditioning. Current Opinion in Neurobiology, 9, 195–202.
Holland, P. C., & Rescorla, R. A. (1975). The effect of two ways of devaluing the unconditioned stimulus after first- and second-order appetitive conditioning. Journal of Experimental Psychology. Animal Behavior Processes, 1, 355–363.
Jordan, W. P., Strasser, H. C., & McHale, L. (2000). Contextual control of long-term habituation in rats. Journal of Experimental Psychology. Animal Behavior Processes, 26, 323–339.
Kaufman, M. A., & Bolles, R. C. (1981). A nonassociative aspect of overshadowing. Bulletin of the Psychonomic Society, 18, 318–320.
Kaye, H., & Mackintosh, N. J. (1990). A change of context can enhance performance of an aversive but not of an appetitive conditioned response. Quarterly Journal of Experimental Psychology, 42B, 113–134.
Kaye, H., & Pearce, J. M. (1984). The strength of the orienting response during Pavlovian conditioning. Journal of Experimental Psychology. Animal Behavior Processes, 10, 90–109.
Kruschke, J. K. (2006). Locally Bayesian learning with applications to retrospective revaluation and highlighting. Psychological Review, 113, 677–699. doi:10.1037/0033-295X.113.4.677
Laborda, M. A., Witnauer, J. E., & Miller, R. R. (2011). Contrasting AAC and ABC renewal: The role of context associations. Learning & Behavior, 39, 46–56. doi:10.3758/s13420-010-0007-1
Larrauri, J. A., & Schmajuk, N. A. (2008). Attentional, associative, and configural mechanisms in extinction. Psychological Review, 115, 640–676.
Mackintosh, N. J. (1975). A theory of attention: Variations in the associability of stimuli with reinforcement. Psychological Review, 82, 276–298. doi:10.1037/h0076778
Marlin, N. A., & Miller, R. R. (1981). Associations to contextual stimuli as a determinant of long term habituation. Journal of Experimental Psychology. Animal Behavior Processes, 7, 313–333.
Mesquita, B., Feldman Barrett, L., & Smith, E. R. (2010). The mind in context. New York: Guilford Press.
Miller, R. R., & Matzel, L. D. (1988). The comparator hypothesis: A response rule for the expression of associations. In G. H. Bower (Ed.), The psychology of learning and motivation (Vol. 22, pp. 51–92). Orlando: Academic Press.
Miller, R. R., & Schachtman, T. R. (1985). The several roles of context at the time of retrieval. In P. D. Balsam & A. Tomie (Eds.), Context and learning (pp. 167–194). Hillsdale: Erlbaum.
Myers, J. L., & Well, A. D. (2003). Research design and statistical analysis (2nd ed.). Mahwah: Erlbaum.
Pavlov, I. P. (1927). Conditioned reflexes (G. V. Anrep, Trans.). London, U.K.: Oxford University Press.
Pearce, J. M., & Hall, G. (1980). A model for Pavlovian learning: Variations in the effectiveness of conditioned but not of unconditioned stimuli. Psychological Review, 87, 532–552. doi:10.1037/0033-295X.87.6.532
Pickens, C. L., Golden, S. A., Adams-Deutsch, T., Nair, S. G., & Shaham, Y. (2009). Long-lasting incubation of conditioned fear in rats. Biological Psychiatry, 65, 881–886.
Rescorla, R. A. (1968). Probability of shock in the presence and absence of CS in fear conditioning. Journal of Comparative and Physiological Psychology, 66, 1–5.
Rescorla, R. A. (1970). Reduction in the effectiveness of reinforcement after prior excitatory conditioning. Learning and Motivation, 1, 372–381.
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In A. H. Black & W. F. Prokasy (Eds.), Classical conditioning II: Current research and theory (pp. 64–99). New York: Appleton-Century-Crofts.
Rosas, J. M., Callejas Aguilera, J. E., Ramos Álvarez, M. M., & Fernández Abad, M. J. (2006). Revision of retrieval theory of forgetting: What does make information context-specific? International Journal of Psychology and Psychological Therapy, 6, 147–166.
Schachtman, T. R., Brown, A. M., Gordon, E. L., Catterson, D. A., & Miller, R. R. (1987). Mechanisms underlying retarded emergence of conditioned responding following inhibitory training: Evidence for the comparator hypothesis. Journal of Experimental Psychology. Animal Behavior Processes, 13, 310–322.
Schull, J. (1979). A conditioned opponent theory of Pavlovian conditioning and habituation. In G. H. Bower (Ed.), The psychology of learning and motivation (Vol. 13, pp. 57–90). New York: Academic Press.
Sissons, H. T., Urcelay, G. P., & Miller, R. R. (2009). Overshadowing and CS duration: Counteraction and a reexamination of the role of within-compound associations in cue competition. Learning & Behavior, 37, 254–268.
Smith, J. D., & Minda, J. P. (1998). Prototypes in the mist: The early epochs of category learning. Journal of Experimental Psychology: Learning, Memory, and Cognition, 24, 1411–1436. doi:10.1037/0278-7393.24.6.1411
Solomon, R. L., & Corbit, J. D. (1974). An opponent-process theory of motivation: I Temporal dynamics of affect. Psychological Review, 81, 119–145.
Spear, N. E. (1973). Retrieval of memories in animals. Psychological Review, 80, 163–194. doi:10.1037/h0034326
Spear, N. E. (1978). The processing of memories: Forgetting and retention. Hillsdale: Erlbaum.
Stout, S. C., Arcediano, F., Escobar, M., & Miller, R. R. (2003a). Overshadowing as a function of trial number: Dynamics of first- and second-order comparator stimuli. Learning & Behavior, 31, 85–97.
Stout, S. C., Chang, R., & Miller, R. R. (2003b). Trial spacing is a determinant of cue interaction. Journal of Experimental Psychology. Animal Behavior Processes, 29, 23–38.
Stout, S. C., & Miller, R. R. (2007). Sometimes competing retrieval (SOCR): A formalization of the comparator hypothesis. Psychological Review, 114, 759–783.
Tulving, E., & Thomson, D. M. (1973). Encoding specificity and retrieval processes in episodic memory. Psychological Review, 80, 352–373. doi:10.1037/h0020071
Urcelay, G. P., & Miller, R. R. (2010). Two roles of the context in Pavlovian fear conditioning. Journal of Experimental Psychology. Animal Behavior Processes, 36, 268–280. doi:10.1037/a0017298
Wagner, A. R. (1981). SOP: A model of automatic memory processing in animal behavior. In N. E. Spear & R. R. Miller (Eds.), Information processing in animals: Memory mechanisms (pp. 5–47). Hillsdale: Erlbaum.
Author Notes
This research was supported by National Institute of Mental Health Grant 33881. The authors thank Joseph Alesandro, Eric Curtis, Mario Laborda, Bridget McConnell, Gonzalo Miguez, Cody Polack, Sarah Sterling, and Yumu Tanaka for their comments on an earlier version of the manuscript. G.P.U. is currently at the Behavioural and Clinical Neuroscience Institute and the Department of Experimental Psychology, University of Cambridge.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Urcelay, G.P., Witnauer, J.E. & Miller, R.R. The dual role of the context in postpeak performance decrements resulting from extended training. Learn Behav 40, 476–493 (2012). https://doi.org/10.3758/s13420-012-0068-4
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13420-012-0068-4