
Abstract
Psychopathic behavior has long been attributed to a fundamental deficit in fear that arises from impaired amygdala function. Growing evidence has demonstrated that fear-potentiated startle (FPS) and other psychopathy-related deficits are moderated by focus of attention, but to date, no work on adult psychopathy has examined attentional modulation of the amygdala or concomitant recruitment of relevant attention-related circuitry. Consistent with previous FPS findings, here we report that psychopathy-related differences in amygdala activation appear and disappear as a function of goal-directed attention. Specifically, decreased amygdala activity was observed in psychopathic offenders only when attention was engaged in an alternative goal-relevant task prior to presenting threat-relevant information. Under this condition, psychopaths also exhibited greater activation in selective-attention regions of the lateral prefrontal cortex (LPFC) than did nonpsychopaths, and this increased LPFC activation mediated psychopathy’s association with decreased amygdala activation. In contrast, when explicitly attending to threat, amygdala activation did not differ in psychopaths and nonpsychopaths. This pattern of amygdala activation highlights the potential role of LPFC in mediating the failure of psychopathic individuals to process fear and other important information when it is peripheral to the primary focus of goal-directed attention.
Similar content being viewed by others


Psychopathy is a common and severe psychopathological disorder affecting approximately 1 % of the general population and 15 %–25 % of incarcerated male offenders (Hare, 1996; Neumann & Hare, 2008). Despite psychopathic individuals’ good intelligence and an absence of Axis I psychopathology (aside from substance abuse; Hart & Hare, 1989), they display an inability to form genuine relationships with parents, teachers, friends, or lovers; limited and superficial affective processing, especially with respect to anticipatory anxiety and remorse; an impulsive behavioral style involving a general failure to evaluate anticipated actions and inhibit the inappropriate ones; and a chronic antisocial lifestyle that entails great costs to society as well as for the affected individual (e.g., incarceration; Cleckley, 1941). Although both affective and behavioral characteristics are important elements of psychopathy, the affective deficits have traditionally been considered the root cause of the psychopath’s problems.
Affective deficits in psychopathy have most often been understood in the context of the low-fear model (Lykken, 1957), which posits that the psychopath’s deficit is mediated by an amygdala-based deficiency (Blair, 2003; Marsh & Cardinale, in press; Patrick, 1994; Viding et al., 2012). In adult samples, whereas some neuroimaging evidence has suggested that psychopathic individuals display less amygdala activation than controls during aversive conditioning, moral decision-making, social cooperation, and memory for emotionally salient words (Birbaumer et al., 2005; Glenn, Raine, & Schug, 2009; Harenski, Harenski, Shane, & Kiehl, 2010; Kiehl et al., 2001; Rilling et al., 2007), other results have indicated that psychopathic individuals display greater amygdala reactivity when viewing emotionally salient scenes and emotionally evocative faces (Carre, Hyde, Neumann, Viding, & Hariri, 2012; Muller et al., 2003). Thus, existing research in psychopathy does not indicate the presence of a reliable amygdala deficit, though such deficits may be revealed for psychopathic individuals under specific experimental circumstances.
Given the potential inconsistency in psychopathy-related amygdala deficiency, it may be that this traditional view of a primary amygdala deficit in psychopathy undervalues the role that cognitive–affective and cortical–subcortical brain interactions have in modulating the complex etiological and phenotypic manifestations of psychopathy. Recent theoretical and empirical models of psychopathy have attempted to integrate cognitive and affective processes and their influence on prototypic psychopathic behavior (Blair, 2007; Kiehl, 2006; Moul, Killcross, & Dadds, 2012; Newman & Baskin-Sommers, 2011). Hence, as deficits in a number of cognitive processes have been implicated in these formulations, there is increasing empirical support for the hypothesis that psychopaths have a core deficit in attention, particularly the adaptive deployment of selective attention (see Newman & Baskin-Sommers, 2011, for a review).
Work in this area has indicated that psychopaths are impaired in the ability to alter top-down goal-directed behavior to incorporate information from salient bottom-up stimuli (including threat cues), and that this inability to modulate behavior results directly from a failure to reallocate attention away from the goal-relevant task toward salient, but task-irrelevant, stimuli (Patterson & Newman, 1993). Moreover, the affective dysfunction observed in psychopaths can be explained by this deficit in adaptively switching between top-down and bottom-up deployment of selective attention. For instance, psychopathic offenders display poor passive avoidance and weak electrodermal responses to punishment cues when they are focused on approach-related goals, but these differences disappear when avoidance learning takes precedence (Arnett, Smith, & Newman, 1997; Newman & Kosson, 1986). Similarly, psychopaths display normal fear-potentiated startle (FPS) when explicitly attending to threat cues, but the same threat cues elicit significantly smaller startle responses in psychopaths than nonpsychopaths when attention is already engaged by another stimulus or task (Baskin-Sommers, Curtin, & Newman, 2011; Newman, Curtin, Bertsch, & Baskin-Sommers, 2010). Thus, across these diverse experimental paradigms, psychopathic offenders display normal responses to affective information when it is part of their primary task, but their reactions to the same affective stimuli are deficient if they have allocated attention to other goal-relevant stimuli (see Newman & Baskin-Sommers, 2011, for review).
Synthesizing these findings, Baskin-Sommers et al. (2011) proposed that an early attention bottleneck is the proximal mechanism underlying deficient emotion reactivity in psychopathy (see also Newman & Baskin-Sommers, 2011). The rationale for the attention bottleneck stems from models that characterize early selective attention as a “fixed bottleneck” where information is processed in serial, and once the bottleneck is established, it blocks the processing of peripheral information that is not goal relevant (Driver, 2001). Such a bottleneck would confer an advantage for psychopaths in filtering potential distractors, but at the cost of undermining the ability to attend to multiple ongoing streams of information. Ultimately, this trade-off would result in a tendency to overlook potential threat and other important information unless it is directly related to their goal-directed focus of attention. Thus, according to the attention bottleneck model, psychopaths are insensitive to threat cues not because they are incapable of fear responses, but because their failure to reallocate attention to affective stimuli while engaged in goal-directed behavior renders them oblivious to these affective cues.
To directly test the attention bottleneck hypothesis, Baskin-Sommers et al. (2011) measured FPS during an instructed fear paradigm that crossed focus of attention (threat vs. alternative focus) with early versus late presentation of the goal-relevant cues. As predicted, psychopaths’ deficit in FPS was nonexistent under conditions that focused attention on the threat-relevant dimension of the stimuli (i.e., threat-focus conditions), but it was pronounced when threat-relevant cues were peripheral to the primary focus of attention (i.e., alternative-focus conditions). More specifically, the psychopathic deficit was only apparent in the early-alternative-focus condition, in which threat cues were presented after the alternative goal-directed focus had been established. That is, psychopathic individuals displayed a deficit in threat processing when their attention was already engaged in a goal-directed focus, and they did not display a threat-processing deficit under any other circumstances.
Despite specific support for the attention bottleneck model and substantial evidence regarding the pivotal role of attention in the affective and inhibitory deficits associated with psychopathy (Blair & Mitchell, 2009; Newman & Baskin-Sommers, 2011), investigators have yet to examine the influence of this early selective-attention bias on emotion processing in psychopathy using neuroimaging. Given the prominence of the low-fear model of psychopathy, much of the extant imaging work in this population has examined amygdala functioning; however, only one study to date has examined amygdala activation to threat-related stimuli as a function of attention, albeit in children with callous–unemotional traits rather than psychopathy per se (White et al., 2012). Also, no study has specifically assessed recruitment of the amygdala using a task explicitly designed to elicit psychopaths’ early selective-attention deficits. Furthermore, despite the amygdala’s prominence in most etiological models of psychopathy, the neuroimaging literature in this area is actually quite small and has yielded somewhat mixed conclusions (see Koenigs, Baskin-Sommers, Zeier, & Newman, 2011, for a review). Although a number of factors (e.g., sample, experimental context) may influence the reliability of a psychopathy-related amygdala deficit, one possibility is that these inconsistencies may be explained by attentional engagement. For example, given the association between FPS and amygdala activation (Davis, Falls, Campeau, & Kim, 1993; Grillon, Ameli, Goddard, Woods, & Davis, 1994), the Baskin-Sommers et al. (2011) FPS findings raise the possibility that amygdala dysfunction in psychopathic offenders may be modulated by an early attention bottleneck. However, direct investigation would be required in order to determine whether the precise attentional manipulations that undermine FPS in psychopathy are also associated with psychopathy-related deficits in amygdala activation.
In addition to assessing whether manipulating attentional engagement modulates amygdala activation in psychopathy, neuroimaging offers the opportunity more specifically to investigate recruitment of top-down attention circuitry that may instantiate the attention bottleneck evident in psychopaths. Basic neuroscience models (Bishop, 2007; Corbetta, Patel, & Shulman, 2008; Dux, Ivanoff, Asplund, & Marois, 2006) and models of psychopathy (Blair, 2003; Kiehl, 2006) highlight the role of the prefrontal cortices (e.g., orbitofrontal, lateral prefrontal) in directing attention toward task-relevant information and inhibiting processing of salient distracting information. Notably, recent work has explicitly identified lateral prefrontal cortex (LPFC) as the site of an information-processing bottleneck (Dux et al., 2006), leading Newman and Baskin-Sommers (2011) to speculate that regions of the LPFC may be central to the early attention bottleneck operating in psychopathy. Moreover, engagement of the LPFC is thought to be specifically necessary to override the amygdala-based automatic processing of threat-related information (Davidson, 2000; Dolcos & McCarthy, 2006; Ochsner et al., 2004; Siegle, Steinhauer, Thase, Stenger, & Carter, 2002). Thus, we hypothesized that LPFC in particular may mediate psychopaths’ deficits in processing multiple streams of information. Although this LPFC-mediated attention bottleneck model provides a promising and potentially crucial perspective on psychopathy, more direct evidence concerning the neural underpinnings of abnormal attention–emotion interactions in psychopathy is required to substantiate the model.
The primary aim of this study was to address the paucity of neuroimaging data available for specifying the neural mechanisms mediating the attention-related modulation of emotion processing in psychopathy. Toward this end, we administered the instructed fear paradigm developed by Baskin-Sommers et al. (2011) in the context of a functional magnetic resonance imaging study. More specifically, by employing the same experimental manipulations that had enabled Baskin-Sommers et al. (2011) to specify the attention bottleneck, we aimed to identify the neural correlates associated with the early attention bottleneck. Paralleling results from the Baskin-Sommers et al. (2011) study, we predicted that psychopathy-related abnormalities associated with the attention bottleneck would be specific to the early-alternative-focus condition. In light of the fact that psychopathic participants displayed less FPS than did controls specifically in the early-alternative-focus condition, we predicted that psychopathy would also be associated with weaker amygdala-related differentiation to threat and safety cues in this condition, but not in the threat-focus or late-alternative-focus conditions. Furthermore, on the basis of the attention bottleneck model, we predicted that psychopathy would be associated with greater activation in regions of LPFC in the early-alternative-focus condition, reflecting a strong goal-directed focus and a failure to disengage attention from the alternative focus (letter) cue when faced with a subsequent salient threat stimulus (red box). We further predicted that this activation in LPFC would mediate the reduced amygdala-related differentiation associated with psychopathy.
Method
Subjects
A group of 71 white male prisoners from a medium-security prison in Southern Wisconsin between the ages of 18 and 45 were asked to participate in this study. Elements of consent were presented individually to all of the participants in verbal and written form. Participants were informed that their decision to take part in the project or to refuse would have no influence on their status within the correctional system. Of the 71, 66 of the prisoners agreed to participate (93 %). Participants were excluded if they were age 45 or older; currently used psychotropic medication; had clinical diagnoses of schizophrenia, bipolar disorder, or psychosis not otherwise specified (NOS); had contraindications for magnetic resonance (MR) scanning (metal in body, unable to lie still in scanner, history of neurological disorder or head injury, or claustrophobia); scored below the 4th-grade reading level on achievement tests administered by the Department of Corrections; or had an estimated IQ score of less than 70 on the Shipley Institute of Living Scale (Zachary, 1986). Participants older than 45 years of age were excluded because the expression of psychopathic traits (i.e., impulsivity) has been found to decline with advancing age (Hare et al., 1990; Harpur & Hare, 1994), and also because some research has suggested that memory and cognitive functioning begins to decline starting around age 45 (Singh-Manoux et al., 2012).
All participants were assessed using file information and a semistructured interview that lasted approximately 60 min and provided sufficient information to diagnose psychopathy using the Psychopathy Checklist–Revised (PCL-R; Hare, 2003). The PCL-R contains 20 items that are rated 0, 1, or 2 according to the degree to which a characteristic was present: significantly (2), moderately (1), or not at all (0). Numerous sources have documented the reliability and validity of the PCL-R (Hare, 2003).
In total, three participants were dropped due to poor alignment of the structural and functional images, six because of movement artifacts, one due to claustrophobia, six because of shock device malfunction, and one (nonpsychopath) because amygdala activation exceeded three standard deviations from the mean. The final sample consisted of 24 psychopaths and 25 nonpsychopaths (see Table 1 for descriptive information about the sample).
Instructed fear task
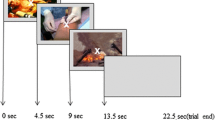
The task consisted of four conditions, with 60 trials per condition. The task conditions crossed focus of attention and timing of goal-relevant information, yielding the early-alternative-focus, early-threat-focus, late-alternative-focus, and late-threat-focus conditions (Fig. 1). In the two threat-focus conditions, participants attended to the color of a box predicting shock administration (red boxes = potential shock; green boxes = safe, no chance of shock). In the two alternative-focus conditions, participants were instructed to attend to the case of a letter stimulus, and the case of the letter was unrelated to administration of electric shocks. Each of the four conditions was blocked, with the order counterbalanced across participants.

Schematic of the task. The instructed fear task consisted of four conditions in which focus of attention and timing of attentional focus were manipulated: (A) early alternative focus, (B) late alternative focus, (C) late threat focus, and (D) early threat focus. During threat focus, participants attended to the color of a box predicting shock administration (red = potential shock, as depicted in (A) and (D); green = safe, as depicted in (B) and (C)). For the alternative focus, participants were instructed to attend to the case of a letter stimulus. In each trial, the box (red or green) and letter (uppercase “N” or lowercase “n”) were presented sequentially, with the order varying according to condition. During early trials, the stimulus to be attended to was presented first, and during late trials, it was presented second. Shocks were presented during the last 200 ms of the blank screen on 20 % of the potential threat trials. Finally, participants pressed a button indicating whether a probe word, “red” or “green” for threat focus and “upper or “lower” for alternative focus, matched the stimulus presented during that trial
On every trial in all conditions, a fixation cross was displayed for 400 to 1,200 ms (M = 800 ms) at the beginning of each trial. Then participants saw two stimuli presented sequentially: a box (red or green) and a letter (uppercase “N” or lowercase “n”), presented sequentially. The order of stimulus appearance varied by condition. The first stimulus (either a box or a letter) appeared alone for 200 ms, and then the second stimulus (the letter or box, whichever had not been presented first) appeared concurrently with the first stimulus for another 200 ms. Following the offset of the stimuli, a blank screen appeared for 1,200 ms. After the blank screen, for 1,600 ms, a descriptive word appeared on the screen, prompting participants to respond on the basis of either the first or the second stimulus, according to the condition. In the threat-focus conditions, the word “red” or “green” was presented, and participants pressed one of two buttons to indicate whether the word described the color of the box during that trial. In the alternative-focus conditions, the word “upper” or “lower” appeared, and participants pressed one of two buttons to indicate whether the word described the case of the letter during that trial. Participants were asked to make a buttonbox response according to the nature of each condition.
In all conditions, participants were given instructions about which stimulus was goal-relevant and which signaled threat (shock) or safety (no shock). The intensity of shocks was calibrated to participants’ subjective tolerance (a procedure administered prior to the task; see the Electrical Stimulation section). If a shock was presented, it occurred during the last 200 ms of the blank screen. Shocks were presented on 20 % of the threat trials (six trials per condition).
Finally, across conditions the timing of the task-relevant focus of attention (early or late) was manipulated to examine the effects of the early attention bottleneck. As mentioned above, this was accomplished by presenting the task-relevant stimulus either first or second in the trial sequence. As is shown in Fig. 1, the early-alternative-focus condition was the only one that engaged attention prior to presenting the threat-relevant information. As such, this condition was the only one expected to reveal a psychopathy-related deficit in amygdala activation.
Electrical stimulation
Shocks were delivered via an AC source (Contact Precision Instruments, Boston, MA) through two Ag/AgCl electrodes placed over the right tibial nerve over the right medial malleolus (right ankle). To control for individual differences in shock sensitivity, the intensity of shocks received during the experimental session was calibrated to the participants’ individual subjective shock sensitivities.
This procedure was conducted immediately prior to the start of the instructed fear paradigm. Participants were administered a series of electric shocks of increasing intensity and rated the intensity of the shock on a scale from 0 (no sensation) to 10 (painful but tolerable). The intensity of the shock administered allowed us to determine two intensity anchors: the first intensity that a participant considered uncomfortable and the maximum intensity level that could be tolerated. The series was terminated when a self-reported intensity level of 10 was reached. This intensity was used for all administrations during the instructed fear task.
Image acquisition
Data were collected on the grounds of the prison using the Mind Research Network (MRN) mobile 1.5-T Siemens Avanto scanner. The MRN mobile MRI scanner is equipped with advanced SQ gradients (max slew rate 200 T/m/s, 346 T/m/s vector summation, rise time 200 µs) and a 12-element head coil (www.mrn.org/facilities/mobile-mri-scanning-facility). EPI echoplanar functional images were collected according to the following parameters: TR/TE (2,000/39 ms, flip angle 75º, FOV 24 × 24 cm, 64 × 64 matrix, 3.75 × 3.75 mm in-plane resolution, 5-mm slice thickness, 27 axial oblique slices). Head motion was limited by using padding. Participants with motion greater than 3 mm were excluded from analysis. High-resolution MPRAGE structural images were obtained to cover the whole brain with 128 1-mm sagittal slices, 8º flip angle and 24-cm FOV.
fMRI data preprocessing and analysis
All fMRI data preprocessing and analysis was conducted with the AFNI software (Cox, 1996). After image reconstruction, rigid-body motion correction was done in three translational and three rotational directions, with all images being registered to the fifth functional image in the time series. The amount of motion in these directions was estimated and then included as a regressor in the subsequent analyses. Participants with more than 3 mm displacement were excluded from the final sample. The MR signal was normalized to a mean of 100 (arbitrary units). Functional time series were deconvolved using a tent function. The mean percentage of signal change for images 2–5 following each stimulus onset was calculated. A spatial blur of 4 mm full width at half maximum was applied, and images were resampled to 1 mm3 voxels. Anatomical images for each participant were transformed to Talairach space, and each functional run was warped to the Talairach-transformed anatomical image. One participant did not have usable anatomical scans, so the functional images were transformed to a standard Talairach template. Threat minus safe difference scores were calculated for each condition for each participant and used as the dependent variable in all analyses.
Whole-brain group analysis
Our main a priori hypothesis was that psychopaths would exhibit normal levels of amygdala activation during all conditions except the early-alternative-focus condition. To test this hypothesis, we conducted an analysis of variance (ANOVA) structured to replicate the startle findings reported by Baskin-Sommers et al. (2011). This voxel-wise ANOVA compared activation between psychopaths and nonpsychopaths for the early-alternative-focus condition against the average of the three remaining conditions, as had been done in the previous startle work. Participant was included as a random factor in the model.Footnote 1 For all of the whole-brain voxel-wise analyses, cluster thresholding based on Monte Carlo simulations calculated with AFNI’s AlphaSim program was applied in order to correct for multiple comparisons. Using a voxel-based p < .025 and nearest-neighbor selection criteria, accounting for spatial correlation, clusters greater than 562 mm3 achieved a whole-brain corrected p value of <.05.
Subsequent post-hoc analyses (using the same cluster-thresholding criteria) were designed to clarify the additional circuitry instantiating aberrant attentional processes for psychopaths in the early-alternative-focus condition. To accomplish this, a voxel-wise t test was calculated comparing activation for psychopaths and nonpsychopaths in the early-alternative-focus condition.
Results
Amygdala activation as a function of attentional focus
As predicted by the attention bottleneck hypothesis, in the early-alternative-focus condition psychopaths exhibited differential activation in the right amygdala as compared to nonpsychopaths (peak: x = 20, y = –6, z = –13; see Fig. 2, as well as Table 2, where data are presented separately for threat and safe for all four attention conditions). In this condition, in which attention was engaged in a neutral task prior to presentation of the task-irrelevant threat cue, nonpsychopaths exhibited relatively greater amygdala activation, particularly in comparison with psychopaths. Also consistent with the attention-based predictions, we found no evidence of psychopathy-related differences in amygdala activation in the remaining three conditions (separately or combined; see Fig. 2 and Table 2).

Results of paired comparisons demonstrating decreased activation of the right amygdala and extended amygdala to threat among psychopaths only when attention is previously engaged by another task. (A) Brain image depicting a significant Group × Condition interaction for the early-alternative-focus condition relative to the average of the three remaining conditions (y = –6). (B) Bar graph depicting the mean threat-minus-safe percentage of signal change for each condition, based on the right amygdala cluster demonstrating a significant group difference for the early-alternative-focus condition. Error bars represent standard errors of the means. As can be seen, during early-alternative-focus trials, decreased activation in psychopaths as compared to nonpsychopaths was present in the right amygdala
Attention-related circuitry: Early-alternative-focus condition
As predicted, group differences in amygdala activation were specific to the early-alternative-focus condition. Thus, follow-up analyses examining attention-related circuitry focused on this condition. A whole-brain t test comparing activation for psychopaths and nonpsychopaths during early alternative focus revealed significant group differences in three left LPFC clusters: the inferior frontal gyrus (IFG: x = –49, y = 14, z = 0; BA 47), middle frontal gyrus (MFG: x = –49, y = 36, z = –4; BA 47), and superior frontal gyrus (SFG: x = –23, y = 48, z = –3; BA 10) (see Fig. 3). The direction of these effects indicated that psychopaths were recruiting neural regions in the dorsolateral (SFG/BA 10) and mid-ventrolateral PFC (mVLPFC; IFG and MFG/BA 47) that are associated with selective attention to a greater extent than did the nonpsychopaths.

Lateral prefrontal cortex (LPFC) group differences and relations with amygdala activation for the early-alternative-focus condition. The brain images (A, B) and bar graph (C) present group differences in left LPFC activation from a voxel-wise t test (higher values on the scale indicate greater activation in psychopaths than in nonpsychopaths). Greater activation for psychopaths as compared to nonpsychopaths is present in (A) the IFG (x = –49, y = 14, z = 0; BA 47) and MFG (x = –49, y = 36, z = –4; BA 47), as well as (B) SFG (x = –23, y = 48, z = –3; BA 10). Error bars in the bar graph (C) represent standard errors of the means
To aid in interpreting the interaction between psychopathy status and recruitment of LPFC regions, simple regressions were conducted between LPFC regions and the amygdala within each group separately.Footnote 2 LPFC activation significantly predicted amygdala activation in psychopaths but not in nonpsychopaths for all three LPFC regions (Fig. 4). For psychopaths, the r values for the IFG, MFG, and SFG were –.44, –.73, and –.49 (all ps < .05), respectively. For the nonpsychopaths, the corresponding IFG, MFG, and SFG r values were .38, –.05, and –.14 (all ps > .05). A test of the significance of the difference between the independent rs (Cohen & Cohen, 1983) revealed that the magnitudes of the LPFC–amygdala correlations were significantly different for the two groups for the two clusters in mVLPFC/BA 47—that is, IFG and MFG (p = .004 for both)—but not for the dorsolateral SFG cluster (p = .19).Footnote 3

Correlations between lateral prefrontal cortex (LPFC) and amygdala for early alternative focus. The scatterplots depict simple regressions predicting amygdala activation with the mid-ventrolateral PFC/BA 47 (IFG, MFG) and the dorsolateral PFC (SFG) for the early-alternative-focus condition separately for both groups. All correlations between PFC regions and the amygdala were significant for the psychopaths (all ps < .05), but not for the nonpsychopaths (all ps > .05)
Finally, we examined whether LPFC activation mediated the relationship between psychopathy and amygdala activation. To test mediation, we used an SPSS macro designed to implement a simultaneous multiple mediator bootstrapping model (Preacher & Hayes, 2004, 2008). Two mediator models were run, once with the LPFC regions combined and once with the three regions (IFG, SFG, and MFG) as separate mediators, to examine the total and direct effects of psychopathy on amygdala activation, as well as the total and specific indirect effects of psychopathy on amygdala activation through the prefrontal regions. All models were based on 2,000 samples, producing 95 % (bias-corrected and accelerated) confidence intervals that did not contain zero. The regression models indicated that (1) psychopathy significantly predicted LPFC activation [in combined LPFC, B = 0.07, SE B = 0.01, t = 4.51, p < .001; in IFG, B = 0.09, SE B = 0.02, t = 3.87, p < .001; in SFG, B = 0.05, SE B = 0.02, t = 2.48, p = .01; and in MFG, B = 0.10, SE B = 0.03, t = 3.84, p < .001]; (2) significant direct effects of LPFC activation on amygdala activation were apparent, but only for the combined LPFC and the specific mVLPFC/BA 47 region [combined LPFC, B = –1.02, SE B = 0.31, t = –3.33, p < .001; IFG, B = 0.28, SE B = 0.24, t = 1.16, p = .25; SFG, B = –0.32, SE B = 0.29, t = –1.09, p = .28 ; MFG, B = –0.80, SE B = 0.23, t = –3.43, p < .001]; (3) the total effect of psychopathy on amygdala activation was significant (B = –0.09, SEB = 0.03, t = –2.35, p = .02); (4) the direct effect of psychopathy status on amygdala activation was not significant (B = –0.02, SEB = 0.04, t = –.57, p = .57); and (5) the indirect effects of psychopathy on amygdala through the combined LPFC (z = –2.22, p = .03) region and mVLPFC/BA 47 (z = –2.63, p = .01) were significant, whereas the indirect effects through IFG (z = –1.16, p = .25) and SFG (z = –1.04, p = .30) were not significant. The overall regression model was significant [F(4, 44) = 7.26, R 2 adj = .34, p < .001]. Together, this pattern of results indicates full mediation when considering the impact of the combined LPFC region, and more specifically mVLPFC/BA 47, on the association between psychopathy and amygdala activation.
Discussion
In light of growing evidence that attention moderates the core behavioral and affective deficits associated with psychopathy, we attempted to extend these findings to amygdala dysfunction because of its prominence in prevailing models of psychopathy (Blair, 2007; Glenn et al., 2009; Patrick, 2007). Paralleling results for fear-potentiated startle (Baskin-Sommers et al., 2011), we found unambiguous evidence that psychopaths’ amygdala dysfunction can be moderated by focus of attention. Moreover, we found that the association between psychopathy and amygdala activation was mediated by regions of LPFC with established links to top-down attention. Thus, in conjunction with previous findings, the present results provide strong evidence for the crucial role of top-down attention in moderating the impact of bottom-up emotion and inhibitory cues among psychopathic offenders.
The role of attention on psychopathy-related differences in amygdala activation
As is predicted by the attention bottleneck model, psychopathy-related deficits in amygdala activation were specific to experimental conditions that engaged attention in goal-relevant processing prior to presenting threat-relevant information. Once they were focused on goal-relevant stimuli (i.e., the letter stimulus), psychopathic offenders displayed significantly less amygdala activation to subsequently presented threat stimuli than did nonpsychopaths, indicating impaired recruitment of task-relevant regions during serial information processing. Also as predicted, this deficit disappeared when threat cues were presented prior to establishing a goal-directed focus of attention (i.e., late alternative focus) and when the conflict between processing goal-relevant and threat-relevant stimuli was eliminated (i.e., under threat-focused conditions). Thus, the present findings corroborate the crucial role of attention in moderating the impact of bottom-up emotion on psychopathic offenders, and further suggest that this attentional moderation specifically influences engagement of the amygdala.
Also addressing the effects of attention on threat processing, albeit in youth with moderate levels of callous–unemotional traits rather than adult psychopathic offenders, White et al. (2012) manipulated attentional load in a primary task while presenting task-irrelevant faces. More specifically, White et al. presented fearful and neutral faces (i.e., goal-irrelevant stimuli) that were flanked by lines that varied in vertical orientation. Participants were instructed to indicate whether or not the lines were parallel (i.e., lines were the goal-relevant stimuli), and attentional load was manipulated by altering the difficulty of this discrimination. Consistent with the attention bottleneck proposal, youth with elevated callous–unemotional traits displayed less affect-mediated amygdala activation to the task-irrelevant fearful faces than did controls when the primary task attentional load was low, though no group difference was found in the high-load condition. White et al. interpreted the absence of group differences in the high-load condition as being inconsistent with the bottleneck hypothesis. However, the lack of group difference in the high-load condition is in keeping with work demonstrating that even in healthy individuals, when attention is fully occupied by high perceptual load, processing of peripheral cues is minimal (Lavie, Hirst, de Fockert, & Viding, 2004). Thus, the opportunity to observe individual differences, particularly in amygdala activation, under such conditions was necessarily limited (Bishop, Jenkins, & Lawrence, 2007; Hsu & Pessoa, 2007; Pessoa, Padmala, & Morland, 2005). In light of such findings, White et al.’s results for the low-load condition appear to provide a more sensitive evaluation of attention-related effects on amygdala activation. Overall, then, the pattern of psychopathy-related results showing deficient amygdala activation to peripheral emotion stimuli when participants are engaged in another goal appears to be consistent with the attention bottleneck model (see Sadeh & Verona, 2008).
Together, these studies provide initial support for attentional moderation of emotion processing in psychopathy and related traits. To focus on the present study, as we noted above, the pattern of psychopathy-related differences in amygdala activation corresponded precisely to our a priori hypotheses. At a more general level, however, the fact that participants, including nonpsychopaths, failed to display significantly more amygdala activation on threat than on safe trials across many of the attention conditions was not expected (see Table 2). Nevertheless, subsequent investigation of this issue revealed a number of theoretical and methodological considerations that put these findings in perspective and diminish concern regarding their implications for the present results. First, a number of other investigations of cognitive–emotion interactions in psychopaths and other antisocial samples (e.g., Marsh et al., 2011; Passamonti et al., 2010; White et al., 2012) have reported negative amygdala activations. Second, the relatively weak amygdala activation found among both psychopaths and nonpsychopaths in our instructed fear paradigm is consistent with results from a recent meta-analysis by Mechias, Etkin, and Kalisch (2010), who failed to find consistent amygdala activation in healthy controls across an array of instructed fear paradigms. Third, growing evidence is showing that attentional load dampens amygdala activation (Bishop et al., 2007; Hsu & Pessoa, 2007; Pessoa et al., 2005). With this in mind, it is worth noting that the threat stimuli in our study were embedded in a cognitive task that required participants to (1) maintain a top-down set to focus on particular stimuli, (2) categorize the stimuli, (3) preserve the categorization in working memory, and (4) make a rapid decision at the end of each trial. Thus, relative to tasks that involve passive viewing of emotion stimuli, our study involved significant cognitive demands that might have played a key role in reducing amygdala activation. Despite these constraints on overall amygdala activation, we propose that the relative level of amygdala activation in this study provides a valid estimate of fear magnitude. In support of this proposal, we note that previous research employing the same paradigm yielded nearly identical psychopathy-related differences in fear responses (but without the negative activations) when fear was indexed using fear-potentiated startle rather than amygdala activation (Baskin-Sommers et al., 2011).
The role of LPFC activation and top-down attention on amygdala activation in psychopathy
In light of compelling evidence that a top-down focus of attention limits crucial information processing and moderates the core deficits in psychopathy, the neurological underpinnings of the attention bottleneck in psychopathy need to be characterized. Thus, we examined regions in prefrontal cortex that were associated with psychopathy-related differences in response to bottom-up threat cues after attention had been allocated to goal-directed stimuli. Consistent with the attention bottleneck model, psychopathy was associated with differential activation of the left LPFC under conditions that established an alternative focus of attention before threat-relevant stimuli were presented (i.e., in the early-alternative-focus condition). In contrast, in the face of salient task-irrelevant threat cues, controls exhibited weaker left LPFC activation. Further supporting the potential importance of left LPFC for the attention bottleneck and its potential impact on threat processing in psychopathy, follow-up analyses revealed that the inverse relationship between LPFC and amygdala activation was significantly greater for psychopaths than for nonpsychopaths. Importantly, the association between psychopathy and reduced amygdala activation was completely mediated by activation in left LPFC, particularly BA 47, a region of mid-ventrolateral PFC (Petrides, 2005).
LPFC has been broadly implicated in selective attention (Badre, Poldrack, Paré-Blagoev, Insler, & Wagner, 2005; Bishop, Duncan, Brett, & Lawrence, 2004; Corbetta & Shulman, 2002; Miller & Cohen, 2001; Rossi, Pessoa, Desimone, & Ungerleider, 2009), including selection of task-relevant information (Bode & Haynes, 2009; Desimone & Duncan, 1995; Duncan, 2006). Of particular relevance for the present work, regions of left LPFC have been identified as the source of an information-processing bottleneck in which interference occurs when individuals attempt to complete multiple tasks (Dux et al., 2006). Interpreted in this light, and consistent with the Baskin-Sommers et al. (2011) proposal, the enhanced left LPFC activation evident in psychopathic offenders may be related to a more pronounced bottleneck in processing multiple streams of information, particularly when one set of information is directly relevant to task demands, and another is not. For psychopaths, this bottleneck appears to enhance their resistance to distraction, but at the cost of processing secondary and peripheral information that may be crucial for regulating maladaptive responses.
The specificity of the increased psychopathy-related LPFC activation to mVLPFC, and BA 47 in particular, is consistent with previous work that has identified the mVLPFC as critical for active selection and maintenance of stimuli requiring further processing (Bunge, 2004; Burgess & Braver, 2010; Hampshire, Duncan, & Owen, 2007; Petrides, 2005). Importantly, BA 47 is recruited to support stimulus-driven orienting, but only for stimuli relevant for the ongoing task, not for task-irrelevant cues (Hampshire et al., 2007; Kincade, Abrams, Vastafiev, Shulman, & Corbetta, 2005; Natale, Marzi, & Macaluso, 2009). This portion of LPFC is also recruited when multiple attributes of a stimulus compete for attentional resources and task demands necessitate the selection of a specific attribute (Hedden & Gabrieli, 2010; Nelson, Reuter-Lorenz, Sylvester, Jonides, & Smith, 2003). Left mVLPFC has also been found to play a causal role in minimizing the interference of distracting task-irrelevant information in order to meet task demands (Wais, Kim, & Gazzaley, 2012). Indeed, individuals who are faster at resolving interference from competing stimulus attributes exhibit greater activation in BA 47 (Egner, 2011). These findings indicate that this portion of LPFC serves to enhance attentional engagement with goal-directed stimuli for the purpose of selecting the most relevant information in a given context.
Thus, mVLPFC, and BA 47 specifically, has been implicated in processes consistent with our interpretation that psychopaths show greater LPFC activation during the early-alternative-focus condition because of enhanced goal-directed allocation of attention (e.g., to the case of the letter, here) in the face of salient task-irrelevant information (i.e., the red box threat cue). Assuming that this region is important for proactive attentional control and maintenance of task-relevant response sets, disruption of activation in this region may allow for bottom-up processes, including diversion of attention to task-irrelevant salient stimuli, to take precedence. In contrast, psychopaths, who show greater activation of BA 47 when confronted with a competing and salient task-irrelevant stimulus, show reduced bottom-up interference effects, as indexed by decreased amygdala activation to the distracting threat cue. Thus, these data suggest that psychopaths are predisposed to focus on primary goals and tasks to the exclusion of goal-irrelevant information, and that this exaggerated selective attention may be mediated by the mVLPFC. More broadly, this pattern of attention selection is consistent with behavioral work indicating that psychopaths are often superior in ignoring distractors and other task-irrelevant information (Hiatt, Schmitt, & Newman, 2004; Newman, Schmitt, & Voss, 1997; Zeier, Maxwell, & Newman, 2009). In light of this dynamic, psychopathic individuals are prone to display an imbalance of selective attention that favors proximal top-down priorities over potentially relevant but unexpected considerations that do modulate the goal-directed behavior of others (see MacCoon, Wallace, & Newman, 2004, for more details).
The overall pattern of results, particularly those from the present study, leads us to propose that the abnormal selective attention in psychopathy stems from “top-down” LPFC mediation of attention during goal-directed behavior. First, psychopaths’ deficit in threat-related amygdala activation was found only during the condition in which attention was already engaged in a goal-directed task, highlighting the specific role of a top-down focus of attention in revealing the psychopath’s deficit. Second, mVLPFC mediated the inverse relationship between psychopathy and amygdala activation. That is, after controlling statistically for the influence of mVLPFC on amygdala activation, psychopathy-related differences in amygdala activation were no longer significant. Finally, as has already been noted (Newman & Baskin-Sommers, 2011), psychopaths’ excessive selective attention is evident with nonemotional as well emotional distractors, further reducing the likelihood that psychopathy-related differences in LPFC activation reflect an emotion-specific deficiency in bottom-up activation. Such evidence provides compelling support for the attention bottleneck model of psychopathy. However, it should be noted that, whereas the present study supported our a priori hypotheses regarding the specific influence of attention-related limitations on emotion processing in psychopathy, others have proposed that the attention abnormalities in psychopathy and differences in LPFC activation are an indirect consequence of deficiencies in “bottom-up” amygdala-related processes (Blair & Mitchell, 2009; Kiehl, 2006). For example, models by Moul et al. (2012) and Blair and Mitchell (2009) have suggested that deficient amygdala functioning may account for most of the extant observations of impaired attention–emotion interactions, particularly given that the subcomponents of the amygdala are related to attentional selection (e.g., basolateral nuclei of the amygdala; Moul et al., 2012). Thus, further research will be needed to characterize the precise contributions of top-down versus bottom-up processes of attention and emotion, and also to understand how these processes operate in different indices of psychopathy (e.g., callous–unemotional traits vs. psychopathy).
Limitations
Although the findings provide strong support for attentional moderation of the neural circuitry underlying threat processing, a number of limitations of the present study must be noted. First, in light of research indicating that laboratory findings among European American offenders often do not generalize to African American offenders (Baskin-Sommers, Newman, Sathasivam, & Curtin, 2010), our sample was limited to adult Caucasian prisoners. The sample also consisted solely of male prisoners. In general, the homogeneous nature of the sample may limit its generalizability to other populations, including younger individuals with traits on the psychopathy spectrum. Also, six participants (9 % of the sample) were dropped due to excessive movement. Although this is a relatively large portion of the sample, it is consistent with other studies in which we have administered electrical stimulation (Balderston, Schultz, & Helmstetter, 2011). Finally, the sample size was somewhat modest, but the results were consistent with the hypotheses, and power appears to have been sufficient to detect group differences.
Conclusions
Challenging the long-standing view that psychopathy involves a core amygdala-mediated deficit in threat and other emotion processing (Blair, 2007; Lykken, 1995; Patrick, 2007), this study provides strong evidence that psychopaths’ amygdala-mediated fear deficit appears and disappears as a function of attention-related priorities. Moreover, the data suggest that psychopaths’ failure to recruit the amygdala in response to salient emotion cues while pursuing immediate goals may be a manifestation of a more general LPFC-instantiated attention bottleneck that severely limits the processing of potentially important peripheral information (Newman & Baskin-Sommers, 2011). The existence of such an early attention bottleneck that undermines the processing of emotion and inhibitory stimuli could provide a compelling explanation for psychopaths’ chronically dysregulated behavior, including their callous disregard for the rights and well-being of others (Newman & Lorenz, 2003). Moreover, to the extent that psychopaths’ core information-processing deficits are moderated by attention, rather than absolute (Newman & Baskin-Sommers, 2011), it may be possible to reduce their antisocial behavior and improve their self-regulation by using newly developed methods for training attention (Baskin-Sommers & Newman, in press; Browning, Holmes, Murphy, Goodwin, & Harmer, 2010).
Notes
Analyses of the amygdala data were also conducted separately for each of the four conditions. These data are presented in the Supplementary materials.
For consistency, the amygdala cluster used in these correlations was also extracted from the between-groups t test for the early-alternative-focus condition. Although it was slightly more ventral, the amygdala cluster yielding significant group differences for this contrast was largely overlapping with that from the initial a priori ANOVA described above (peak: x = 19, y = –7, z = –9). The cluster size for the original ANOVA was 1,211 mm3, whereas for the early-alternative-focus condition t test, the cluster size was 1,257 mm3. A conjunction analysis revealed an overlap of 915 mm3.
We also assessed whether group moderated the association between LPFC and amygdala activation by using moderated regression (Aiken & West, 1991). After entering group and mean-centered LPFC activation into the model predicting amygdala activation (separately for each LPFC cluster), the interaction between group and LPFC was significant for IFG (β = –.40, p = .004) and MFG (β = –.29, p = .04), but not for SFG (β = –.15, p = .29).
References
Aiken, L. S., & West, S. G. (1991). Multiple regression: Testing and interpreting interactions. Newbury Park, CA: Sage.
Arnett, P. A., Smith, S. S., & Newman, J. P. (1997). Approach and avoidance motivation in psychopathic criminal offenders during passive avoidance. Journal of Personality and Social Psychology, 72, 1413–1428.
Badre, D., Poldrack, R. A., Paré-Blagoev, E. J., Insler, R. Z., & Wagner, A. D. (2005). Dissociable controlled retrieval and generalized selection mechanisms in ventrolateral prefrontal cortex. Neuron, 47, 907–918. doi:10.1016/j.neuron.2005.07.023
Balderston, N. L., Schultz, D. H., & Helmstetter, F. J. (2011). The human amygdala plays a specific role in the detection of novelty. NeuroImage, 55, 1889–1898.
Baskin-Sommers, A. R., Curtin, J. J., & Newman, J. P. (2011). Specifying the attentional selection that moderates the fearlessness of psychopathic offenders. Psychological Science, 22, 226–234. doi:10.1177/0956797610396227
Baskin-Sommers, A. R., & Newman, J. P. (2013). Differentiating the cognition-emotion interactions that characterize psychopathy versus externalizing disorders. In M. D. Robinson, E. Harmon-Jones, & E. R. Watkins (Eds.), Handbook of Cognition and emotion (pp. 501–520). New York, NY: Guilford Press.
Baskin-Sommers, A. R., Newman, J. P., Sathasivam, N., & Curtin, J. J. (2010). Evaluating the generalizability of a fear deficit in African American offenders. Journal of Abnormal Psychology, 120, 71–78. doi:10.1037/a0021225
Birbaumer, N., Veit, R., Lotze, M., Erb, M., Hermann, C., Grodd, W., & Flor, H. (2005). Deficient fear conditioning in psychopathy: A functional magnetic resonance imaging study. Archives of General Psychiatry, 62, 799–805. doi:10.1001/archpsyc.62.7.799
Bishop, S. J. (2007). Neurocognitive mechanisms of anxiety: An integrative account. Trends in Cognitive Science, 11, 307–316. doi:10.1016/j.tics.2007.05.008
Bishop, S. J., Duncan, J., Brett, M., & Lawrence, A. D. (2004). Prefrontal cortical function and anxiety: Controlling attention to threat-related stimuli. Nature Neuroscience, 7, 184–188. doi:10.1038/nn1173
Bishop, S. J., Jenkins, R., & Lawrence, A. D. (2007). Neural processing of fearful faces: Effects of anxiety are gated by perceptual capacity limitations. Cerebral Cortex, 17, 1595–1603.
Blair, R. J. (2003). Neurobiological basis of psychopathy. The British Journal of Psychiatry, 182, 5–7.
Blair, R. J. (2007). The amygdala and ventromedial prefrontal cortex in morality and psychopathy. Trends in Cognitive Sciences, 11, 387–392. doi:10.1016/j.tics.2007.07.003
Blair, R. J., & Mitchell, D. G. (2009). Psychopathy, attention and emotion. Psychological Medicine, 39, 543–555. doi:10.1017/S0033291708003991
Bode, S., & Haynes, J. D. (2009). Decoding sequential stages of task preparation in the human brain. NeuroImage, 45, 606–613. doi:10.1016/j.neuroimage.2008.11.031
Browning, M., Holmes, E. A., Murphy, S. E., Goodwin, G. M., & Harmer, C. J. (2010). Lateral prefrontal cortex mediates the cognitive modification of attentional bias. Biological Psychiatry, 67, 919–925. doi:10.1016/j.biopsych.2009.10.031
Bunge, S. A. (2004). How we use rules to select actions: A review of evidence from cognitive neuroscience. Cognitive, Affective, & Behavioral Neuroscience, 4, 564–579. doi:10.3758/CABN.4.4.564
Burgess, G. C., & Braver, T. S. (2010). Neural mechanisms of interference control in working memory: Effects of interference expectancy and fluid intelligence. PLoS One, 5, e12861. doi:10.1371/journal.pone.0012861
Carre, J. M., Hyde, L. W., Neumann, C. S., Viding, E., & Hariri, A. R. (2012). The neural signatures of distinct psychopathic traits. Social Neuroscience, 8, 122–135.
Cleckley, H. (1941). The mask of sanity: An attempt to reinterpret the so-called psychopathic personality. St. Louis, MO: Mosby.
Cohen, J., & Cohen, P. (1983). Applied multiple regression/correlation analysis for the behavioral sciences (2nd ed.). Hillsdale, NJ: Erlbaum.
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting system of the human brain: From environment to theory of mind. Neuron, 58, 306–324. doi:10.1016/j.neuron.2008.04.017
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews Neuroscience, 3, 201–215. doi:10.1038/nrn755
Cox, R. W. (1996). AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Computers and Biomedical Research, 29, 162–173.
Davidson, R. J. (2000). Affective style, psychopathology, and resilience: Brain mechanisms and plasticity. American Psychologist, 55, 1196–1214.
Davis, M., Falls, W. A., Campeau, S., & Kim, M. (1993). Fear-potentiated startle: A neural and pharmacological analysis. Behavioral and Brain Research, 58, 175–198.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18, 193–222. doi:10.1146/annurev.ne.18.030195.001205
Dolcos, F., & McCarthy, G. (2006). Brain systems mediating cognitive interference by emotional distraction. Journal of Neuroscience, 26, 2072–2079.
Driver, J. (2001). A selective review of selective attention research from the past century. British Journal of Psychology, 92, 53–78.
Duncan, J. (2006). EPS Mid-Career Award 2004: Brain mechanisms of attention. Quarterly Journal of Experimental Psychology, 59, 2–27. doi:10.1080/17470210500260674
Dux, P. E., Ivanoff, J., Asplund, C. L., & Marois, R. (2006). Isolation of a central bottleneck of information processing with time-resolved FMRI. Neuron, 52, 1109–1120. doi:10.1016/j.neuron.2006.11.009
Egner, T. (2011). Right ventrolateral prefrontal cortex mediates individual differences in conflict-driven cognitive control. Journal of Cognitive Neuroscience, 23, 3903–3913.
Glenn, A. L., Raine, A., & Schug, R. A. (2009). The neural correlates of moral decision-making in psychopathy. Molecular Psychiatry, 14, 5–6. doi:10.1038/mp.2008.104
Grillon, C., Ameli, R., Goddard, A., Woods, S. W., & Davis, M. (1994). Baseline and fear-potentiated startle in panic disorder patients. Biological Psychiatry, 35, 431–439.
Hampshire, A., Duncan, J., & Owen, A. M. (2007). Selective tuning of the blood oxygenation level-dependent response during simple target detection dissociates human frontoparietal subregions. Journal of Neuroscience, 27, 6219–6223. doi:10.1523/jneurosci.0851-07.2007
Hare, R. D. (1996). Psychopathy: A clinical construct whose time has come. Criminal Justice and Behavior, 23, 25–54.
Hare, R. D. (2003). Manual for Revised Psychopathy Checklist. Toronto, ON, Canada: Multi-Health Systems.
Hare, R. D., Harpur, T. J., Hakistan, A. R., Forth, A. E., Hart, S. D., & Newman, J. P. (1990). The revised Psychopathy Checklist: Reliability and factor structure. Psychological Assessment, 2, 338–341.
Harenski, C. L., Harenski, K. A., Shane, M. S., & Kiehl, K. A. (2010). Aberrant neural processing of moral violations in criminal psychopaths. Journal of Abnormal Psychology, 119, 863–874.
Harpur, T. J., & Hare, R. D. (1994). Assessment of psychopathy as a function of age. Journal of Abnormal Psychology, 103, 604–609. doi:10.1037/0021-843X.103.4.604
Hart, S. D., & Hare, R. D. (1989). Discriminant validity of the Psychopathy Checklist in a forensic psychiatric population. Psychological Assessment, 1, 211–218.
Hedden, T., & Gabrieli, J. D. E. (2010). Shared and selective neural correlates of inhibition, facilitation, and shifting processes during executive control. NeuroImage, 51, 421–431. doi:10.1016/j.neuroimage.2010.01.089
Hiatt, K. D., Schmitt, W. A., & Newman, J. P. (2004). Stroop tasks reveal abnormal selective attention among psychopathic offenders. Neuropsychology, 18, 50–59. doi:10.1037/0894-4105.18.1.50
Hsu, S.-M., & Pessoa, L. (2007). Dissociable effects of bottom-up and top-down factors in the processing of unattended fearful faces. Neuropsychologia, 45, 3075–3086.
Kiehl, K. A. (2006). A cognitive neuroscience perspective on psychopathy: Evidence for paralimbic system dysfunction. Psychiatry Research, 142, 107–128. doi:10.1016/j.psychres.2005.09.013
Kiehl, K. A., Smith, A. M., Hare, R. D., Mendrek, A., Forster, B. B., Brink, J., & Liddle, P. F. (2001). Limbic abnormalities in affective processing by criminal psychopaths as revealed by functional magnetic resonance imaging. Biological Psychiatry, 50, 677–684.
Kincade, J. M., Abrams, R. A., Vastafiev, S. V., Shulman, G. L., & Corbetta, M. (2005). An event-related functional magnetic resonance imaging study of voluntary and stimulus-driven orienting of attention. The Journal of Neuroscience, 25, 4593–4604. doi:10.1523/jneurosci.0236-05.2005
Koenigs, M., Baskin-Sommers, A., Zeier, J., & Newman, J. P. (2011). Investigating the neural correlates of psychopathy: A critical review. Molecular Psychiatry, 16, 792–799. doi:10.1038/mp.2010.124
Lavie, N., Hirst, A., de Fockert, J. W., & Viding, E. (2004). Load theory of selective attention and cognitive control. Journal of Experimental Psychology. General, 133, 339–354. doi:10.1037/0096-3445.133.3.339
Lykken, D. T. (1957). A study of anxiety in the sociopathic personality. Journal of Abnormal and Social Psychology, 55, 6–10.
Lykken, D. T. (1995). The antisocial personalities. Hillsdale, NJ: Erlbaum.
MacCoon, D. G., Wallace, J. F., & Newman, J. P. (2004). The context-appropriate allocation of attentional capacity to dominant and non-dominant cues. In R. F. Baumeister & K. D. Vohs (Eds.), Handbook of self-regulation: Research, theory, and application (pp. 422–466). New York, NY: Guilford Press.
Marsh, A. A., & Cardinale, E. M. (in press). When psychopathy impairs moral judgments: Neural responses during judgments about causing fear. Social, Cognitive, and Affective Neuroscience. doi:10.1093/scan/nss097
Marsh, A. A., Finger, E. C., Fowler, K. A., Jurkowitz, I. N., Schechter, J. C., Yu, H. H., & Blair, R. J. R. (2011). Reduced amygala-orbitofrontal connectivity during moral judgments in youths with distruptive behavior disorders and psychopathic traits. Psychiatry Research, 194, 279–286.
Mechias, M.-L., Etkin, A., & Kalisch, R. (2010). A meta-analysis of instructed fear studies: Implications for conscious appraisal of threat. NeuroImage, 49, 1760–1768.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefrontal cortex function. Annual Review of Neuroscience, 24, 167–202. doi:10.1146/annurev.neuro.24.1.167
Moul, C., Killcross, S., & Dadds, M. R. (2012). A model of differential amygdala activation in psychopathy. Psychological Review, 119, 789–806.
Muller, J. L., Sommer, M., Wagner, V., Lange, K., Taschler, H., Roder, C. H., & Hajak, G. (2003). Abnormalities in emotion processing within cortical and subcortical regions in criminal psychopaths: evidence from a functional magnetic resonance imaging study using pictures with emotional content. Biological Psychiatry, 54, 152–162.
Natale, E., Marzi, C. A., & Macaluso, E. (2009). fMRI correlates of visuo-spatial reorienting investigated with an attention shifting double-cue paradigm. Human Brain Mapping, 30, 2367–2381. doi:10.1002/hbm.20675
Nelson, J. K., Reuter-Lorenz, P. A., Sylvester, C.-Y. C., Jonides, J., & Smith, E. A. (2003). Dissociable neural mechanisms underlying response-based and familiarity-based conflict in working memory. Proceedings of the National Academy of Science, 100, 11171–11175. doi:10.10173/pnas.1334125100
Neumann, C. S., & Hare, R. D. (2008). Psychopathic traits in a large community sample: Links to violence, alcohol use, and intelligence. Journal of Consulting and Clinical Psychology, 76, 893–899.
Newman, J. P., & Baskin-Sommers, A. R. (2011). Early selective attention abnormalities in psychopathy: Implications for self-regulation. In M. I. Posner (Ed.), Cogntive neuroscience of attention (2nd ed., pp. 421–440). New York, NY: Guilford Press.
Newman, J. P., Curtin, J. J., Bertsch, J. D., & Baskin-Sommers, A. R. (2010). Attention moderates the fearlessness of psychopathic offenders. Biological Psychiatry, 67, 66–70. doi:10.1016/j.biopsych.2009.07.035
Newman, J. P., & Kosson, D. S. (1986). Passive avoidance learning in psychopathic and nonpsychopathic offenders. Journal of Abnormal Psychology, 95, 252–256.
Newman, J. P., & Lorenz, A. R. (2003). Response modulation and emotion processing: Implications for psychopathy and other dysregulatory psychopathology. In R. J. Davidson, K. Scherer, & H. H. Goldsmith (Eds.), Handbook of affective sciences (pp. 422–446). Oxford, UK: Oxford University Press.
Newman, J. P., Schmitt, W. A., & Voss, W. D. (1997). The impact of motivationally neutral cues on psychopathic individuals: Assessing the generality of the response modulation hypothesis. Journal of Abnormal Psychology, 106, 563–575.
Ochsner, K. N., Ray, R. D., Cooper, J. C., Robertson, E. R., Chopra, S., Gabrieli, J. D. E., & Gross, J. J. (2004). For better or for worse: Neural systems supporting the cognitive down- and up-regulation of negative emotion. NeuroImage, 23, 483–499. doi:10.1016/j.neuroimage.2004.06.030
Passamonti, L., Fairchild, G., Goodyer, I. M., Hurford, G., Hagan, C. C., Rowe, J. B., & Calder, A. J. (2010). Neural abnormalities in early-onset conduct disorder. Archives of General Psychiatry, 67, 729–738.
Patrick, C. J. (1994). Emotion and psychopathy: Startling new insights. Psychophysiology, 31, 319–330.
Patrick, C. J. (2007). Getting to the heart of psychopathy. In H. Herve & J. C. Yuille (Eds.), The psychopath: Theory, research, and social implications (pp. 207–252). Hillsdale, NJ: Erlbaum.
Patterson, C. M., & Newman, J. P. (1993). Reflectivity and learning from aversive events: Toward a psychological mechanism for the syndromes of disinhibition. Psychological Review, 100, 716–736. doi:10.1037/0033-295X.100.4.716
Pessoa, L., Padmala, S., & Morland, T. (2005). Fate of unattended fearful faces in the amygdala is determined by both attentional resources and cognitive modulation. NeuroImage, 28, 249–255. doi:10.1016/j.neuroimage.2005.05.048
Petrides, M. (2005). Lateral prefrontal cortex: architectonic and functional organization. Philosophical Transactions of the Royal Society B, 360, 781–795.
Preacher, K. J., & Hayes, A. F. (2004). SPSS and SAS procedures for estimating indirect effects in simple mediation models. Behavior Research Methods, Instruments, & Computers, 36, 717–731. doi:10.3758/BF03206553
Preacher, K. J., & Hayes, A. F. (2008). Asymptotic and resampling strategies for assessing and comparing indirect effects in multiple mediator models. Behavior Research Methods, 40, 879–891. doi:10.3758/BRM.40.3.879
Rilling, J. K., Glenn, A. L., Jairam, M. R., Pagnoni, G., Goldsmith, D. R., Elfenbein, H. A., & Lilienfeld, S. O. (2007). Neural correlates of social cooperatin and non-cooperation as a function of psychopathy. Biological Psychiatry, 61, 1260–1271.
Rossi, A. F., Pessoa, L., Desimone, R., & Ungerleider, L. G. (2009). The prefrontal cortex and the executive control of attention. Experimental Brain Research, 192, 489–497. doi:10.1007/s00221-008-1642-z
Sadeh, N., & Verona, E. (2008). Psychopathic personality traits associated with abnormal selective attention and impaired cognitive control. Neuropsychology, 22, 669–680. doi:10.1037/a0012692
Siegle, G. J., Steinhauer, S. R., Thase, M. E., Stenger, V. A., & Carter, C. S. (2002). Can’t shake that feeling: fMRI assessment of sustained amygdala activity in response to emotional information in depressed individuals. Biological Psychiatry, 51, 693–707.
Singh-Manoux, A., Kivimaki, M., Glymour, M. M., Elbaz, A., Berr, C., Ebmeier, K. P., & Dugravot, A. (2012). Timing of onset of cognitive decline: Results from Whitehall II prospective cohort study. British Medical Journal, 344, d7622.
Viding, E., Sebastian, C. L., Dadds, M. R., Lockwood, P., Cecil, C., DeBrito, S., & McCrory, E. (2012). Amygdala response to pre-attentive masked fear is associated with callous–unemotional traits in children with conduct problems. The American Journal of Psychiatry, 169, 1109–1116. doi:10.1176/appi.ajp.2012.12020191
Wais, P. E., Kim, O. Y., & Gazzaley, A. (2012). Distractibility during episodic retrieval is exacerbated by perturbation of left ventrolateral prefrontal cortex. Cerebral Cortex, 22, 717–724. doi:10.1093/cercor/bhr160
White, S. F., Marsh, A. A., Fowler, K. A., Schechter, J. C., Adalio, C., Pope, K., & Blair, R. J. (2012). Reduced amygdala response in youths with disruptive behavior disorders and psychopathic traits: Decreased emotional response versus increased top-down attention to nonemotional features. The American Journal of Psychiatry, 169, 750–758.
Zachary, R. A. (1986). Shipley Institute of Living Scale: Revised Manual. Los Angeles, CA: Western Psychological Service.
Zeier, J. D., Maxwell, J. S., & Newman, J. P. (2009). Attention moderates the processing of inhibitory information in primary psychopathy. Journal of Abnormal Psychology, 118, 554–563. doi:10.1037/a0016480
Author note
This study was supported by a grant from the University of Wisconsin–Madison/University of Wisconsin–Milwaukee IRIGP, and by NIMH Grant Nos. MH053041 (to J.P.N.), MH070539 (to K.A.K.), NIDA DA026505 (to K.A.K.), MH086809 (to C.L.L.), and T32-MH018931 (to A.R.B.-S.). We thank the Wisconsin Department of Corrections, and especially Dr. Kevin Kallas and Deputy Warden Tom Nickel, for making this research possible.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(PDF 159 kb)
Rights and permissions
About this article
Cite this article
Larson, C.L., Baskin-Sommers, A.R., Stout, D.M. et al. The interplay of attention and emotion: top-down attention modulates amygdala activation in psychopathy. Cogn Affect Behav Neurosci 13, 757–770 (2013). https://doi.org/10.3758/s13415-013-0172-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13415-013-0172-8