
Abstract
Infant studies have suggested that the detection of biological motion (BM) might be an innate capacity, based on newborns’ spontaneous preference for BM. However, it is unclear if, like adults, infants recognize humans from BM and are able to build the representation of bodies and faces. To address this issue, we tested whether exposure to BM influences subsequent face recognition in 3- to 8-month-old infants. After familiarization with a point-light walker (PLW) of either a female or a male, the infant’s preference for female and male faces was measured. If infants can build the representation of not only the body but also the face from PLWs, the familiarization effect of gender induced by the PLW might be generalized to faces. We found that infants at 7 to 8 months looked for longer at the face whose gender was opposite to that of the PLW, whereas 3- to 4- and 5- to 6-month-old infants did not. These results suggest that infants can access the representation of humans from BM and extract gender, which is shared across bodies and faces, from at least 7 to 8 months of age.
Similar content being viewed by others

Humans are highly sensitive to the actions of others, which carries important information for social cognition. A perceptual phenomenon that demonstrates such sensitivity to human movements is biological motion (BM; Johansson, 1973), in which the motion of only a few light dots attached to body parts evokes a vivid perception of a moving human. We can extract much information from BM, such as the walker’s direction (Bertenthal & Pinto, 1994; Mather et al., 1992), gender (Kozlowski & Cutting, 1977; Mather & Murdoch, 1994), emotions (Dittrich et al., 1996), action categories (Dittrich, 1993), and social interaction (Neri et al., 2006).
Infant studies on BM have revealed that 4-month-old infants (Fox & McDaniel, 1982), newborn human infants (Bardi et al., 2011, 2014; Simion et al., 2008), and newly hatched chicks (Vallortigara et al., 2005) show a preference for point-light walkers (PLWs) rather than inverted or scrambled PLWs or random motion. These results suggest that infants have a predisposition to orienting toward the movement of biological organisms, as well as toward faces (Goren et al., 1975; Morton & Johnson, 1991; Valenza et al., 1996). Thus, BM perception is likely to be an innate capacity.
However, it is not clear whether infants, like adults, recognize humans from BM—that is, whether infants can build the representation of humans including bodies and faces from the dynamics of BM. Spontaneous preference for PLWs clearly demonstrates that infants can detect BM; however, this does not necessarily indicate that they recognize humans from PLWs. It has been proposed that BM perception might be mediated by subcortical areas in early infancy (Hirai & Senju, 2020; Simion et al., 2011), whereas it is mediated by cortical areas relevant to social cognition in adults (Engell & McCarthy, 2013; Grossman & Blake, 2002; Sokolov et al., 2018; Vaina et al., 2001). Thus, it is possible that the spontaneous preference for BM in early infancy is an automatic behavior that is not accompanied by recognition of humans.
The ability to extract information from BM has also been shown to develop during infancy. The ability to discriminate between walking and running PLWs (Booth et al., 2002) and walkers’ directions (Kuhlmeier et al., 2010) develops by 3 and 6 months, respectively. The gender of PLWs can be distinguished by around 6 months (Tsang et al., 2018) and categorized at 9 months (Johnson et al., 2021). However, the presence of these abilities to distinguish or categorize different types of PLWs also does not necessarily indicate that infants are able to recognize humans from PLWs; the results mean that infants can detect the difference in the dynamics between the two types of PLWs. Some studies have shown that infants can detect social information from point-light displays (PLDs) such as the direction of attention at 12 months of age (Furuhata & Shirai, 2015; Yoon & Johnson, 2009) and interaction between two PLDs at 14 months (Galazka et al., 2014). Detecting social information might indirectly suggest that infants are able to access conceptual knowledge of humans from BM. However, there is no study that directly proves infants are capable of recognizing from BM not only the body shape of humans but conceptual knowledge of them. Whether PLWs can be recognized as humans is a fundamental problem since the PLW stimulus is widely used to investigate social cognition. Infants’ ability to access the representation of humans from PLWs must be established in order to determine whether they have similar BM recognition to adults.
To examine whether infants can recognize humans from BM, the present study tested whether exposure to BM influences subsequent face recognition in 3- to 8-month-old infants. We used the familiarization and novelty preference paradigm, in which, after familiarization with a specific stimulus, infants prefer a novel stimulus over the familiarized stimulus. In our experiment, either a female or male PLW was presented repeatedly in the familiarization phase; subsequently, female and male faces were presented simultaneously (Experiment 1). If infants can recognize humans from PLWs, the familiarization effect of gender induced by PLWs might be generalized to face preference; that is, infants might prefer faces whose gender is opposite to that of the PLW. In this paradigm, infants are required to distinguish the gender of PLWs and recognize them as humans. Since PLWs do not include any information about faces, generalization from PLWs to faces would not occur unless infants are able to access the representation of humans from PLWs and to build the representation of not only the body but also the face of humans. Thus, this paradigm allows us to test infant’s abilities to extract from PLWs the representation of humans consisting of the face and the body.
Infants’ ability to distinguish the gender of PLWs has already been revealed with a familiarization paradigm using PLWs and faces (Johnson et al., 2021), which used a similar method to ours. However, their study did not examine the recognition of humans from PLWs. In their method, infants were first familiarized with several pairs of a face stimulus and a PLW of the same gender. After the learning, a face and two PLWs (male and female) were presented simultaneously and infants were tested to see whether they could detect a gender-mismatched PLW (whose gender was different from that of the face). Because infants learned the association of faces and PLWs of the same gender prior to the test, whether they recognize humans including the face and the body from PLWs cannot be tested through this paradigm (i.e., infants do not need to be able to extract the face and body representation from PLWs to detect the gender-mismatched PLWs). Infants only need to categorize the gender of PLWs and the gender of faces independently because they learned the association between PLWs and faces during the experiment. Although Johnson et al. (2021) demonstrate that infants can categorize the dynamics of PLWs of different gender, the results do not necessarily indicate that infants can recognize from PLWs the gender which is shared across faces and bodies and is not limited to PLWs. By contrast, in our method, infants are familiarized only to a PLW without such association learning, and hence they must be able to build the face and body representation only from PLWs so that generalization from PLWs to faces is possible. Our method allows us to examine if infants can recognize from BM the gender that is independent of specific stimuli.
Moreover, in another experiment, to investigate whether local information of the PLWs influences subsequent face preference, infants were familiarized with inverted PLWs (Experiment 2). BM perception is known to be disrupted when a PLW is inverted (Pavlova & Sokolov, 2000; Sumi, 1984). Thus, if the familiarization effect is generalized from PLWs to faces in Experiment 1 because infants recognize PLWs as humans, the generalization would disappear when a PLW is inverted. However, if the generalized effect in Experiment 1 is due to local information of PLWs, such as differences in spans or speed of dot motion between female and male PLWs, the generalization will remain even when a PLW is inverted.
We examined 3- to 8-month-old infants because various abilities to extract information from BM (Booth et al., 2002; Kuhlmeier et al., 2010; Tsang et al., 2018) and brain responses to it (Hirai & Hiraki, 2005; Lisboa et al., 2020) develop during this period and we predicted that the recognition of humans from BM might also develop during this period.
Methods
Participants
The final sample consisted of 120 Japanese infants in Experiment 1. The infants were in three age groups: 3–4 months, 5–6 months, and 7–8 months. In the female-PLW condition, 20 3- to 4-month-old infants (12 female, age range: 97–134 days, mean age = 120.9 days), 20 5- to 6-month-old infants (seven female, age range: 137–194 days, mean age = 171.1 days), and 20 7- to 8-month-old infants (seven female, age range: 200–254 days, mean age = 228.1 days) were tested. An additional 42 infants were tested but removed from the analysis because of fussiness (n = 12; 3–4 months: five infants, 5–6 months: three infants, 7–8 months: four infants), side bias (n = 13; 3–4 months: 12 infants, 5–6 months: one infant; looking time at one side of the monitor was more than 90% of the total looking time in the test trials), or failure to familiarize (n = 20; 3–4 months: seven infants, 5–6 months: seven infants, 7–8 months: six infants; the average looking time in the last two familiarization trials was more than 90% of that in the first two trials). In the male-PLW condition, 20 3- to 4-month-old infants (13 female, age range: 86–132 days, mean age = 114.8 days), 20 5- to 6-month-old infants (nine female, age range: 148–194 days, mean age = 177.3 days), and 20 7- to 8-month-old infants (11 female, age range: 195–254 days, mean age = 232.5 days) were tested. An additional 55 infants were tested but removed from the analysis because of fussiness (n = 9; 3–4 months: six infants, 5–6 months: one infant, 7–8 months: two infants), side bias (n = 20; 3–4 months: 20 infants), or failure to familiarize (n = 28; 3–4 months: 11 infants, 5–6 months: seven infants, 7–8 months: 10 infants).
The final sample consisted of 40 infants aged 7–8 months in Experiment 2. In the female-PLW condition, 20 infants (10 female, age range: 200–252 days, mean age = 229.2 days) were tested. An additional 12 infants were tested but removed from the analysis because of failure to familiarize. In the male-PLW condition, 20 infants (10 female, age range: 201–254 days, mean age = 234.3 days) were tested. An additional 13 infants were tested but excluded from the analysis because of failure to familiarize.
The exclusion criteria were decided before data collection and were based on previous studies (for side bias: Otsuka et al., 2013; Tsang et al., 2018; for failure to familiarize: Nakashima et al., 2019; Otsuka et al., 2013; Skelton et al., 2017). The attrition rates were relatively high: 41% for the female-PLW condition and 47% for the male-PLW condition in Experiment 1, and 37% for female-PLW condition and 39% for the male-PLW condition in Experiment 2. It is likely to be because the PLW stimuli were attractive for infants and many did not reach the familiarization criterion. However, there are studies whose attrition rates are equal to or higher than our study because of a high attraction of stimuli and failure to familiarize or habituate (Arterberry & Yonas, 1988; Cohen & Cashon, 2001; Otsuka et al., 2009), and thus the attrition rate in the present study does not seem to be unusually high compared with others. Moreover, when we analyzed data including infants that failed to familiarize, the same results were obtained as when those infants were excluded (see Additional analysis in Supplementary Information).
The sample size was calculated to achieve a power of 0.8 with an effect size of 0.65 (Cohen’s d), based on a study examining infants’ ability to discriminate the gender of faces (Quinn et al., 2002) and various infant studies that measured looking time (Oakes, 2017).
Infants were recruited by advertisements on local newspapers distributed in Tokyo, Japan, and on the lab website. All participants were full term at birth and healthy at the time of the test. This study was conducted according to the Declaration of Helsinki and was approved by the ethics committee of Chuo University. Written informed consent was obtained from all the parents.
Apparatus and stimuli
Stimuli were displayed on a 21.5-inch LCD monitor (BenQ GW2270-T, 1,920 × 1,080 pixel, 60 Hz) with PsychoPy (Version 1.85.6; Peirce et al., 2019). Each infant sat on a parent’s lap in front of the monitor in a dark room. The viewing distance was 40 cm. A camera (Logicool C920R) was located below the monitor to record the infant’s behavior and sounds. An experimenter could observe the behavior of infants through another monitor connected to the camera.
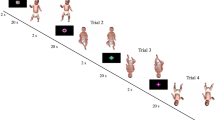
Two PLW stimuli were created from the motion capture data of the female and male walkers (Fig. 1a). The data were selected from the Carnegie Mellon University Motion Capture Database (http://mocap.cs.cmu.edu; file number: “49_01” as female and “90_22” as male). The motion capture data were converted to a point-light display in the frontal view using the Biological Motion Toolbox (van Boxtel & Lu, 2013) in MATLAB (The MathWorks, Natick, MA, USA). The PLW stimuli comprised 16 circular white dots (0.3° in diameter, 246 cd/m2) placed on the main joints, such as the shoulders, hips, and feet. The PLWs appeared to walk forward on a treadmill. A single walking cycle was extracted from the original motion capture data and was repeated for 10 s. The duration of a walking cycle was approximately 1.4 s and 1.2 s for the female and male PLWs, respectively. The transition from the last frame back to the first frame was smoothed such that the walking action looped seamlessly. The stimuli were exported as a movie presented on PsychoPy. The maximum width and height of both the female and male PLWs were 6.3° and 15.4°, respectively.

Stimuli and procedure. The PLW stimuli (a) comprised 16 dots placed on the main joints, which appeared to walk forward on a treadmill. In a familiarization trial (b, left), either the female or the male PLW was presented for 10 s and the trial was repeated six times. In a test trial (b, right), female and male faces were presented side by side for 10 s. The test trial was repeated twice and the position of the female and male faces was reversed. The test phase was conducted before (pretest) and after (posttest) the familiarization. In Experiment 2, PLWs were inverted in the familiarization phase
Two face stimuli (female and male) were created by morphing face pictures of Japanese adults taken in frontal view. Face images of five females and five males were used to create each face stimulus. The stimulus contained outer features, such as hair, ears, and neck. The size of the face stimuli was 5.6° in width × 6.6° in height for the female face and 5° in width × 6.9° in height for the male face.
Procedure
The visual stimuli were presented on a grey background (33.7 cd/m2). In Experiment 1, an upright PLW stimulus was presented at the center of the screen during the familiarization phase (Fig. 1b). The familiarization phase consisted of six trials. Each trial lasted until the infant looked away from the monitor for at least 3 s or until the trial reached a 10-s maximum. This familiarization time (a total of 60 s) is the same as that used in previous studies examining gender recognition of faces (Quinn et al., 2002) and PLWs (Johnson et al., 2021). In the female-PLW condition, the female PLW was presented; in the male-PLW condition, the male PLW was presented.
The test phase was conducted before (pretest) and after (posttest) familiarization, in which female and male face stimuli were displayed side by side on the screen (Fig. 1b). The face stimuli were located at 12.8° left and right from the center of the monitor. Each test phase (i.e., pretests and posttests) consisted of two trials, with each trial lasting 10 s. The positions of the female and male faces were reversed across the two trials, and the order of the two trials was counterbalanced across infants. The same test phase was conducted for female- and male-PLW conditions. Before each trial in the familiarization and test phases, a cartoon was presented at the center of the screen with a short beeping sound. Each trial began after the infants looked at the cartoon. The cartoon appeared repeatedly for 500 ms at 500-ms intervals.
In Experiment 2, the stimuli and procedure were the same as those in Experiment 1, except that the PLWs were inverted during the familiarization phase.
Data coding and analysis
We measured the duration for which infant looked at the stimuli. An observer who was naïve to the stimulus identity scored the looking time from off-line videos. In the familiarization trials, the observer pressed a key while infants looked at the central area of the screen where the PLW stimulus appeared. The key was released when infants looked away. In the test trials, the observer pressed a key while infants looked at the face presented on the left side and pressed another key while infants looked at the face on the right side. When the infant looked away from the stimuli, no recording was made. The looking time in the two test trials was summed for each face stimulus. To test whether the familiarization effect induced by PLWs was generalized to face recognition, preference for a novel gender was calculated by dividing the looking time for face stimuli whose gender was opposite to that of the familiarized PLW by the total looking time in the test trials (i.e., the sum of the looking time for both face stimuli).
In Experiment 1, to assess the difference in preference for a novel gender between the pretests and posttests, a three-way mixed analysis of variance (ANOVA), with test phase (pretests and posttests) as a within-participant factor and PLW gender (female and male) and age group (3–4, 5–6, and 7–8 months) as between-participant factors, was used. To quantify the effect observed in the ANOVA, we also calculated a Bayes factor indicating the likelihood of the hypothesis (familiarization effect (post–pre) is different between 3- to 6-month-olds and 7- to 8-month-olds; see Results) over the null hypothesis. In addition, to assess whether the preference for a novel gender was greater than the chance level (50%), a two-sided one-sample t test was used. The significance level was corrected using the false discovery rate (FDR) for multiple comparisons (12 tests). To assess the looking time in the familiarization phase, a three-way mixed ANOVA, with familiarization trials (six trials) as a within-participant factor and PLW gender and age group as between-participant factors, was used. In Experiment 2, the same tests were used as in Experiment 1, except that two-way ANOVAs without the age-group factor were used for preference for a novel gender and the looking time in the familiarization phase, and the FDR correction was used for four t tests.
All data have been made publicly available at the Open Science Framework and can be accessed online (https://osf.io/qtzfg/?view_only=6c8de9d19b1f4308a2331e1320cb3585). Materials for this study are available by emailing the corresponding author.
Results
Experiment 1
In the familiarization phase, the looking time was reduced as the trials advanced, and the overall looking time was longer for the male PLW than for the female PLW (Fig. S1a in Supplementary Information). A three-way ANOVA showed a significant main effect of familiarization trials, F(5, 570) = 79.48, p < .001, η2 = .25, and a significant main effect of PLW gender, F(1, 114) = 4.87, p < .05, η2 = .01, but no other effects. The greater interest in male PLWs compared with female PLWs observed here is consistent with previous studies (Johnson et al., 2021; Tsang et al., 2018).
Figure 2 shows preference for a novel gender in the test phases for each age group. A three-way ANOVA showed a significant main effect of age group, F(2, 114) = 3.83, p < .05, η2 = .03, and a significant interaction between test phase and age group, F(2, 114) = 3.15, p < .05, η2 = .03, but no other effects. Tests for simple effects of test phase in each age group showed that preference for a novel gender was stronger in the posttest than in the pretest in 7- to 8-month-olds, F(1, 39) = 18.54, p < .001, η2 = .11, but there was no effect of test phase in 5- to 6-month-olds, F(1, 39) = 0.13, p = .73, η2 = .002, and in 3- to 4-month-olds, F(1, 39) = 0.48, p = .49, η2 = .007. A Bayes factor testing a hypothesis that the familiarization effect (post–pre) is different between 3- to 6-month-olds and 7- to 8-month-olds against the null hypothesis was 3.14.

Results of Experiment 1. Preference for a novel gender in the pretests and posttests is shown for each age group. In a graph of each age group, the two left bars show the results for the female PLW condition and the right bars show the results for the male PLW condition. Error bars indicate a 95% confidence interval of the mean. Asterisks represent significant differences (**p < .01). The light-gray lines indicate the individual data. (Color figure online)
In 7- to 8-month-old infants, preference for a novel gender was greater than chance level (50%) in the posttest for both female and male PLW conditions (Fig. 2), female PLW: t(19) = 3.87, p < .01, q (FDR) < .05, d = 0.87; male PLW: t(19) = 3.20, p < .01, q < .05, d = 0.72, but not in the pretest, female PLW: t(19) = 1.41, p = .18, d = 0.31; male PLW: t(19) = −0.47, p = .64, d = −0.11. In 5- to 6-month-old and 3- to 4-month-old infants, such an effect was not observed. In infants at 5 to 6 months, preference for a novel gender did not differ from the chance level in either the pretest, female PLW: t(19) = −0.31, p = .76, d = −0.07; male PLW: t(19) = −0.95, p = .36, d = −0.21, or posttest, female PLW: t(19) = −0.94, p = .36, d = −0.21; male PLW: t(19) = −0.86, p = .40, d = −0.19. Similarly, in infants at 3 to 4 months, preference for a novel gender did not differ from the chance level in both pretest, female PLW: t(19) = 1.00, p = .28, d = 0.22; male PLW: t(19) = 0.41, p = .68, d = 0.09, and posttest, female PLW: t(19) = −0.26, p = .85, d = −0.06; male PLW: t(19) = 0.10, p = .92, d = 0.02.
Taken together, these results indicate that, after familiarization with a PLW, 7- to 8-month-old infants looked for longer at the face whose gender was opposite to the PLW. However, in 5- to 6-month-old and 3- to 4-month-old infants, familiarization with a PLW did not influence subsequent face recognition.
Experiment 2
In Experiment 2, 7- to 8-month-old infants were investigated using inverted PLWs. Looking time in the familiarization phase reduced as the trials advanced and did not differ between female and male PLWs (Fig. S1b). ANOVA showed a significant main effect of familiarization trials, F(5, 190) = 35.58, p < .001, η2 = .33, but no effect of PLW gender, F(1, 38) = 0.27, p = .606, η2 = .002, and the interaction, F(5, 190) = 0.25, p = .937, η2 = .002.
Preference for a novel gender in the test phases was shown in Fig. 3. ANOVA shows that there was no main effect, test phase: F(1, 38) = 0.09, p = .77, η2 = .001; PLW gender: F(1, 38) = 2.12, p = .15, η2 = .03, and no interaction, F(1, 38) = 0.96, p = .33, η2 = .01. The preference for a novel gender did not differ from the chance level in either the pretest, female PLW: t(19) = −0.52, p = .55, d = −0.12; male PLW: t(19) = −0.03, p = .98, d < −0.01, or posttest, female PLW: t(19) = −1.22, p = .20, d = −0.27; male PLW: t(19) = 1.07, p = .30, d = 0.24. Thus, the preference for the novel gender’s face in the posttest, observed in Experiment 1, disappeared when the PLW was inverted in the familiarization trials.

Results of Experiment 2. Preference for a novel gender in the pretests and posttests is shown for the female and male PLW conditions. Error bars indicate a 95% confidence interval of the mean. The light-gray lines indicate the individual data. (Color figure online)
Discussion
The present study examined whether infants are capable of recognizing humans from BM and building the body and face representation, as adults do, by testing whether information extracted from PLWs is generalized to subsequent face recognition. After familiarization with a PLW, 7- to 8-month-old infants looked for longer at the face of the novel gender, indicating that a familiarization effect induced by PLWs is generalized to face recognition and that 7- to 8-month-old infants are able to match gender across PLWs and faces. Since PLWs do not contain any information about faces, this generalization can occur only if infants distinguish PLWs of different gender and build the representation of not only the body but the face from PLWs. Thus, the present results suggest that 7- to 8-month-old infants can extract gender information and access the representation of humans consisting of the body and face from the dynamic of PLWs. Although infants have been shown to prefer spontaneously BM over non-BM (Bardi et al., 2011, 2014; Fox & McDaniel, 1982; Simion et al., 2008) and to be able to distinguish or categorize several types of PLWs (Booth et al., 2002; Johnson et al., 2021; Kuhlmeier et al., 2010; Tsang et al., 2018), these findings do not provide evidence that they can recognize humans from PLWs as adults do. Our results suggest that 7- to 8-month-old infants have the ability to process BM similar to that of adults in terms of being able to recognize conceptual knowledge of humans from PLWs.
When familiarized with an inverted PLW, the bias of looking time toward the novel-gender face was not observed. This suggests that the generalization from PLWs to faces observed in Experiment 1 is not due to local information of PLWs, such as differences in spans or speed of dot motion between female and male PLWs.
Johnson et al. (2021) examined infants’ ability to recognize gender of PLWs using a method similar to ours, a familiarization paradigm using PLWs and faces. However, they did not examine the recognition of humans from PLWs. Since the infants learned the association of faces and PLWs during the experiment, they did not need to extract the face and body representation from PLWs. Thus, whether infants can recognize the representation of humans from PLWs cannot be tested through their paradigm, unlike our study. Another study (Tsang et al., 2018) also examined infants’ ability to recognize PLWs’ gender with the habituation method, but only PLW stimuli without faces were used. Although these studies (Johnson et al., 2021; Tsang et al., 2018) have revealed that infants can categorize the dynamics of PLWs based on gender, we cannot conclude from the results that infants can recognize the gender of humans that is independent of specific stimuli like PLWs. Our results demonstrate that 7- to 8-month-old infants are able to extract from BM the gender which is shared across bodies and faces.
The ability to detect gender information across visual and auditory modalities has been shown to emerge between 9 and 12 months; infants at these ages can match faces and voices based on gender (Hillairet de Boisferon et al., 2015; Poulin-Dubois et al., 1994; Richoz et al., 2017). Representation of gender extracted from PLWs and faces, which tested in the present study, is likely to emerge earlier than that from faces and voices. This is possibly because modality-specific representation of gender develops earlier than multimodal representation of gender.
It has been shown that infants exhibit spontaneous preference for female faces (Quinn et al., 2002; Quinn et al., 2008). However, no bias in looking time between female and male faces was observed in the pretest of the present study. One possibility for this could be that gender of faces to which infants in our study were exposed was not strongly biased toward females. Infants’ preference between female versus male faces depends on gender of their primary caregivers (Quinn et al., 2002). Another possibility is that female-face preference might be somewhat weak in Asian faces. When using Asian faces, one study found female-face preference (Liu et al., 2015), while another study found no bias in preference, even in infants who are not White or who are mixed-race (Kim et al., 2015). This might be because the difference in facial features between females and males is less in Asian than White (Hopper et al., 2014).
In 3- to 6-month-old infants, familiarization with PLWs did not influence subsequent face recognition. There are two possible reasons why generalization was not observed: (1) 3- to 6-month-old infants could not recognize humans from PLWs, or (2) they could recognize humans from PLWs but could not distinguish the gender of PLWs. Our paradigm cannot determine which is the case; thus, we cannot conclude whether younger infants are able to recognize PLWs as humans. Our results only suggest that from at least 7 months of age, infants can distinguish the PLWs’ gender and recognize humans from PLWs.
However, previous studies suggest that infants can process gender information from human bodies by at least 5 to 6 months of age. Infants at 5 months can recognize gender from body shape (Hock et al., 2015), and infants at 6 months and older can discriminate between female and male PLWs (Tsang et al., 2018). These results suggest that 5- to 6-month-old infants, in which the familiarization effect is not generalized from PLWs to faces in our study, might be able to process gender information from PLWs. Thus, it is possible that the absence of generalization in 5- to 6-month-old infants is because they cannot recognize humans from PLWs, rather than being unable to distinguish PLWs of different gender.
Moreover, there are results implying that infants at around 6 months cannot recognize PLWs as human bodies. Infants at 9 months of age are sensitive to occlusion information of PLWs (Bertenthal et al., 1984); some dots on PLW’s arms and feet will be occluded when they pass behind the unseen body, and the appropriate occlusion of these dots enhances perception of PLWs as a person’s motion in adults (Proffitt et al., 1984). However, 5- and 7.5-month-old infants do not show such sensitivity, suggesting that infants at these ages might not recognize PLWs as human bodies (Bertenthal et al., 1984). In addition, infants aged 9 months and older can recognize the difference between human bodies of natural and unnatural structures only when the body stimulus moves in a biologically possible way (Christie & Slaughter, 2010). However, such an enhancement of human body recognition by biological movement was not observed in 6-month-old infants, suggesting that infants at this age cannot integrate biological movement with the human body. Given these findings, it is possible that recognition of humans from PLWs might emerge from around 7 to 9 months of age, although further studies are needed to test this possibility.
Our findings suggest that the development of BM perception in infancy consists of two distinct stages: The first is the detection of BM (Bardi et al., 2011, 2014; Fox & McDaniel, 1982; Simion et al., 2008), which is presumably without the recognition of humans from BM; the second is BM processing accompanied by recognition of humans. This view is closely related to the two-process model of BM processing, proposing that BM perception is underlaid by the two systems that process local and global information (Chang & Troje, 2009; Hirai & Senju, 2020), derived from the two-process model of infant face processing (Morton & Johnson, 1991). The local system detects the local foot motion of PLWs preattentively. This system is thought to be mediated in subcortical areas and emerges early in development, which explains the preference for PLWs observed in newborns (Bardi et al., 2011, 2014; Simion et al., 2008). The global system processes body form information carried by the global motion of PLWs, which is mediated in cortical areas and requires postnatal learning. The present results suggest that the early mechanism to detect local information of BM might be independent from the recognition of BM as humans and that such recognition is related to the global system and acquired through postnatal visual experiences. At around 1 year of age, infants acquire the ability to process higher-level information on BM, such as the direction of the PLW’s attention at 12 months (Furuhata & Shirai, 2015; Yoon & Johnson, 2009) and interaction between two PLWs at 14 months (Galazka et al., 2014). Abilities to process such social information could be acquired based on the recognition of humans from BM.
The development of neural mechanisms underlying BM perception is also consistent with the findings of the present study. In adults, BM processing is mediated by occipitotemporal areas, including the right posterior superior temporal sulcus and the fusiform face area (Engell & McCarthy, 2013; Grossman & Blake, 2002; Sokolov et al., 2018; Vaina et al., 2001). Several studies have shown that adult-like brain responses to BM are observed from 7 to 8 months (Hirai & Hiraki, 2005; Lisboa et al., 2020). However, in infants at 5 months (Marshall & Shipley, 2009) or 10 weeks (Reid et al., 2019), the ERP response to PLWs is largely different from that of adults and older infants. Thus, the right posterior temporal areas, possibly related to social cognition, are likely to develop around 7 to 8 months, and the development of these areas could underlie the interaction between BM and face recognition observed in 7- to 8-month-old infants.
Data availability
All data have been made publicly available at the Open Science Framework and can be accessed online (https://osf.io/75wau/?view_only=64c30a98ecf64502aad075ee2ad1a1fc). None of the experiments was preregistered.
Code availability
Experimental code is available upon request to the corresponding author.
References
Arterberry, M. E., & Yonas, A. (1988). Infants’ sensitivity to kinetic information for three-dimensional object shape. Perception & Psychophysics, 44, 1–6. https://doi.org/10.3758/BF03207466
Bardi, L., Regolin, L., & Simion, F. (2011). Biological motion preference in humans at birth: Role of dynamic and configural properties. Developmental Science, 14, 353–359. https://doi.org/10.1111/j.1467-7687.2010.00985.x
Bardi, L., Regolin, L., & Simion, F. (2014). The first time ever I saw your feet: inversion effect in newborns’ sensitivity to biological motion. Developmental Psychology, 50, 986–993. https://doi.org/10.1037/a0034678
Bertenthal, B. I., & Pinto, J. (1994). Global processing of biological motions. Psychological Science, 5, 221–225. https://doi.org/10.1111/j.1467-9280.1994.tb00504.x
Bertenthal, B. I., Proffitt, D. R., Spetner, N. B., & Thomas, M. A. (1984). The development of infant sensitivity to biomechanical motions. Child Development, 56, 531–543. https://doi.org/10.2307/1129742
Booth, A. E., Pinto, J., & Bertenthal, B. I. (2002). Perception of the symmetrical patterning of human gait by infants. Developmental Psychology, 38, 554–563. https://doi.org/10.1037/0012-1649.38.4.554
Chang, D. H., & Troje, N. F. (2009). Characterizing global and local mechanisms in biological motion perception. Journal of Vision, 9(5), 8. https://doi.org/10.1167/9.5.8
Christie, T., & Slaughter, V. (2010). Movement contributes to infants’ recognition of the human form. Cognition, 114, 329–337. https://doi.org/10.1016/j.cognition.2009.10.004
Cohen, L. B., & Cashon, C. H. (2001). Do 7-month-old infants process independent features or facial configurations? Infant and Child Development, 10, 83–92. https://doi.org/10.1002/icd.250
Dittrich, W. H. (1993). Action categories and the perception of biological motion. Perception, 22, 15–22. https://doi.org/10.1068/p220015
Dittrich, W. H., Troscianko, T., Lea, S. E., & Morgan, D. (1996). Perception of emotion from dynamic point-light displays represented in dance. Perception, 25, 727–738. https://doi.org/10.1068/p250727
Engell, A. D., & McCarthy, G. (2013). Probabilistic atlases for face and biological motion perception: An analysis of their reliability and overlap. NeuroImage, 74, 140–151. https://doi.org/10.1016/j.neuroimage.2013.02.025
Fox, R., & McDaniel, C. (1982). The perception of biological motion by human infants. Science, 218, 486–487. https://doi.org/10.1126/science.7123249
Furuhata, N., & Shirai, N. (2015). The development of gaze behaviors in response to biological motion displays. Infant Behavior and Development, 38, 97–106. https://doi.org/10.1016/j.infbeh.2014.12.014
Galazka, M. A., Roché, L., Nyström, P., & Falck-Ytter, T. (2014). Human infants detect other people’s interactions based on complex patterns of kinematic information. PLoS One, 9, Article e112432. https://doi.org/10.1371/journal.pone.0112432
Goren, C. C., Sarty, M., & Wu, P. Y. (1975). Visual following and pattern discrimination of face-like stimuli by newborn infants. Pediatrics, 56, 544–549. https://doi.org/10.1542/peds.56.4.544
Grossman, E. D., & Blake, R. (2002). Brain areas active during visual perception of biological motion. Neuron, 35, 1167–1175. https://doi.org/10.1016/S0896-6273(02)00897-8
Hillairet de Boisferon, A., Dupierrix, E., Quinn, P. C., Loevenbruck, H., Lewkowicz, D. J., Lee, K., & Pascalis, O. (2015). Perception of multisensory gender coherence in 6- and 9-month-old infants. Infancy, 20, 661–674. https://doi.org/10.1111/infa.12088
Hirai, M., & Hiraki, K. (2005). An event-related potentials study of biological motion perception in human infants. Cognitive Brain Research, 22, 301–304. https://doi.org/10.1016/j.cogbrainres.2004.08.008
Hirai, M., & Senju, A. (2020). The two-process theory of biological motion processing. Neuroscience and Biobehavioral Reviews, 111, 114–124. https://doi.org/10.1016/j.neubiorev.2020.01.010
Hock, A., Kangas, A., Zieber, N., & Bhatt, R. S. (2015). The development of sex category representation in infancy: Matching of faces and bodies. Developmental Psychology, 51, 346–352. https://doi.org/10.1037/a0038743
Hopper, W. J., Finklea, K. M., Winkielman, P., & Huber, D. E. (2014). Measuring sexual dimorphism with a race–gender face space. Journal of Experimental Psychology: Human Perception and Performance, 40, 1779. https://doi.org/10.1037/a0037743
Johansson, G. (1973). Visual perception of biological motion and a model for its analysis. Perception & Psychophysics, 14, 201–211. https://doi.org/10.3758/BF03212378
Johnson, S. P., Dong, M., Ogren, M., & Senturk, D. (2021). Infants’ identification of gender in biological motion displays. Infancy, 26, 798–810. https://doi.org/10.1111/infa.12406
Kim, H. I., Johnson, K. L., & Johnson, S. P. (2015). Gendered race: Are infants’ face preferences guided by intersectionality of sex and race? Frontiers in Psychology, 6, 1330. https://doi.org/10.3389/fpsyg.2015.01330
Kozlowski, L. T., & Cutting, J. E. (1977). Recognizing the sex of a walker from a dynamic point-light display. Perception & Psychophysics, 21, 575–580. https://doi.org/10.3758/BF03198740
Kuhlmeier, V. A., Troje, N. F., & Lee, V. (2010). Young infants detect the direction of biological motion in point-light displays. Infancy, 15, 83–93. https://doi.org/10.1111/j.1532-7078.2009.00003.x
Lisboa, I. C., Queirós, S., Miguel, H., Sampaio, A., Santos, J. A., & Pereira, A. F. (2020). Infants’ cortical processing of biological motion configuration–A fNIRS study. Infant Behavior and Development, 60, 101450. https://doi.org/10.1016/j.infbeh.2020.101450
Liu, S., Xiao, N. G., Quinn, P. C., Zhu, D., Ge, L., Pascalis, O., & Lee, K. (2015). Asian infants show preference for own-race but not other-race female faces: The role of infant caregiving arrangements. Frontiers in Psychology, 6, 593. https://doi.org/10.3389/fpsyg.2015.00593
Marshall, P. J., & Shipley, T. F. (2009). Event-related potentials to point-light displays of human actions in 5-month-old infants. Developmental Neuropsychology, 34, 368–377. https://doi.org/10.1080/87565640902801866
Mather, G., & Murdoch, L. (1994). Gender discrimination in biological motion displays based on dynamic cues. Proceedings of the Royal Society B: Biological Sciences, 258, 273–279. https://doi.org/10.1098/rspb.1994.0173
Mather, G., Radford, K., & West, S. (1992). Low-level visual processing of biological motion. Proceedings of the Royal Society B: Biological Sciences, 249, 149–155. https://doi.org/10.1098/rspb.1992.0097
Morton, J., & Johnson, M. H. (1991). CONSPEC and CONLERN: A two-process theory of infant face recognition. Psychological Review, 98, 164–181. https://doi.org/10.1037/0033-295X.98.2.164
Nakashima, Y., Yamaguchi, M. K., & Kanazawa, S. (2019). Development of center-surround suppression in infant motion processing. Current Biology, 29, 3059–3064. https://doi.org/10.1016/j.cub.2019.07.044
Neri, P., Luu, J. Y., & Levi, D. M. (2006). Meaningful interactions can enhance visual discrimination of human agents. Nature Neuroscience, 9, 1186–1192. https://doi.org/10.1038/nn1759
Oakes, L. M. (2017). Sample size, statistical power, and false conclusions in infant looking-time research. Infancy, 22, 436–469. https://doi.org/10.1111/infa.12186
Otsuka, Y., Konishi, Y., Kanazawa, S., & Yamaguchi, M. K. (2009). The effect of occlusion on motion integration in infants. Journal of Experimental Psychology: Human Perception and Performance, 35, 72. https://doi.org/10.1037/0096-1523.35.1.72
Otsuka, Y., Motoyoshi, I., Hill, H. C., Kobayashi, M., Kanazawa, S., & Yamaguchi, M. K. (2013). Eye contrast polarity is critical for face recognition by infants. Journal of Experimental Child Psychology, 115, 598–606. https://doi.org/10.1016/j.jecp.2013.01.006
Pavlova, M., & Sokolov, A. (2000). Orientation specificity in biological motion perception. Perception & Psychophysics, 62, 889–899. https://doi.org/10.3758/BF03212075
Peirce, J., Gray, J. R., Simpson, S., MacAskill, M., Höchenberger, R., Sogo, H., Kastman, E., & Lindeløv, J. K. (2019). PsychoPy2: Experiments in behavior made easy. Behavior Research Methods, 51, 195–203. https://doi.org/10.3758/s13428-018-01193-y
Poulin-Dubois, D., Serbin, L. A., Kenyon, B., & Derbyshire, A. (1994). Infants intermodal knowledge about gender. Developmental Psychology, 30, 436–442. https://doi.org/10.1037/0012-1649.30.3.436
Proffitt, D. R., Bertenthal, B. I., & Roberts, R. J. (1984). The role of occlusion in reducing multistability in moving point-light displays. Perception & Psychophysics, 36, 315–323. https://doi.org/10.3758/BF03202783
Quinn, P. C., Yahr, J., Kuhn, A., Slater, A. M., & Pascalis, O. (2002). Representation of the gender of human faces by infants: A preference for female. Perception, 31, 1109–1121. https://doi.org/10.1068/p3331
Quinn, P. C., Uttley, L., Lee, K., Gibson, A., Smith, M., Slater, A. M., & Pascalis, O. (2008). Infant preference for female faces occurs for same-but not other-race faces. Journal of Neuropsychology, 2, 15–26. https://doi.org/10.1348/174866407X231029
Reid, V. M., Kaduk, K., & Lunn, J. (2019). Links between action perception and action production in 10-week-old infants. Neuropsychologia, 126, 69–74. https://doi.org/10.1016/j.neuropsychologia.2017.11.005
Richoz, A. R., Quinn, P. C., Hillairet de Boisferon, A., Berger, C., Loevenbruck, H., Lewkowicz, D. J., Lee, K., Dole, M., Caldara, R., & Pascalis, O. (2017). Audio-visual perception of gender by infants emerges earlier for adult-directed speech. PLoS One, 12, Article e0169325. https://doi.org/10.1371/journal.pone.0169325
Simion, F., Regolin, L., & Bulf, H. (2008). A predisposition for biological motion in the newborn baby. Proceedings of the National Academy of Sciences, USA, 105, 809–813. https://doi.org/10.1073/pnas.0707021105
Simion, F., Giorgio, E. D., Leo, I., & Bardi, L. (2011). The processing of social stimuli in early infancy: From faces to biological motion perception. Progress in Brain Research, 189, 173–193. https://doi.org/10.1016/B978-0-444-53884-0.00024-5
Skelton, A. E., Catchpole, G., Abbott, J. T., Bosten, J. M., & Franklin, A. (2017). Biological origins of color categorization. Proceedings of the National Academy of Sciences, USA, 114, 5545–5550. https://doi.org/10.1073/pnas.1612881114
Sokolov, A. A., Zeidman, P., Erb, M., Ryvlin, P., Friston, K. J., & Pavlova, M. A. (2018). Structural and effective brain connectivity underlying biological motion detection. Proceedings of the National Academy of Sciences, USA, 115, E12034–E12042. https://doi.org/10.1073/pnas.1812859115
Sumi, S. (1984). Upside-down presentation of the Johansson moving light-spot pattern. Perception, 13, 283–286. https://doi.org/10.1068/p130283
Tsang, T., Ogren, M., Peng, Y., Nguyen, B., Johnson, K. L., & Johnson, S. P. (2018). Infant perception of sex differences in biological motion displays. Journal of Experimental Child Psychology, 173, 338–350. https://doi.org/10.1016/j.jecp.2018.04.006
Vaina, L. M., Solomon, J., Chowdhury, S., Sinha, P., & Belliveau, J. W. (2001). Functional neuroanatomy of biological motion perception in humans. Proceedings of the National Academy of Sciences, USA, 98, 11656–11661. https://doi.org/10.1073/pnas.191374198
Valenza, E., Simion, F., Macchi Cassia, V., & Umiltà, C. (1996). Face preference at birth. Journal of Experimental Psychology: Human Perception and Performance, 22, 892–903. https://doi.org/10.1037/0096-1523.22.4.892
Vallortigara, G., Regolin, L., & Marconato, F. (2005). Visually inexperienced chicks exhibit spontaneous preference for biological motion patterns. PLoS Biology, 3, 1312–1316. https://doi.org/10.1371/journal.pbio.0030208
van Boxtel, J. J., & Lu, H. (2013). A biological motion toolbox for reading, displaying, and manipulating motion capture data in research settings. Journal of Vision, 13(12), 7. https://doi.org/10.1167/13.12.7
Yoon, J. M., & Johnson, S. C. (2009). Biological motion displays elicit social behavior in 12-month-olds. Child Development, 80, 1069–1075. https://doi.org/10.1111/j.1467-8624.2009.01317.x
Funding
This study was supported by Japan Society for the Promotion of Science (JSPS) KAKENHI Grant 19K14479 (Y.N.) and 17H06343 (M.K.Y.).
Author information
Authors and Affiliations
Contributions
Y.N., S.K., and M.K.Y. designed research; Y.N. performed research; Y.N. analyzed data; and Y.N., S.K., and M.K.Y. wrote the paper.
Corresponding author
Ethics declarations
Conflict of interest
We have no conflicts of interest.
Ethics approval
This study was conducted according to the Declaration of Helsinki and was approved by the ethics committee of Chuo University.
Consent to participate and for publication
Written informed consent was obtained from all the parents for their children’s participation and for publication in a scientific article.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
(DOCX 219 kb)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Nakashima, Y., Kanazawa, S. & Yamaguchi, M.K. Recognition of humans from biological motion in infants. Atten Percept Psychophys 85, 2567–2576 (2023). https://doi.org/10.3758/s13414-023-02675-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-023-02675-8