
Abstract
Perceptual priming can stabilize the phenomenal appearance of multistable visual displays (Leopold, Wilke, Maier, & Logothetis, Nature Neuroscience, 5, 605–609, 2002). Prior exposure to such displays induces a sensory memory of their appearance, which persists over long intervals and intervening stimulation, and which facilitates renewed perception of the same appearance. Here, we investigated perceptual priming for the apparent rotation in depth of ambiguous structure-from-motion (SFM) displays. Specifically, we generated SFM objects with different three-dimensional shapes and presented them in random order and with intervening blank periods. To assess perceptual priming, we established the probability that a perceived direction of rotation would persist between successive objects. In general, persistence was greatest between identical objects, intermediate between similar objects, and negligible between dissimilar objects. These results demonstrate unequivocally that sensory memory for apparent rotation is specific to three-dimensional shape, contrary to previous reports (e.g., Maier, Wilke, Logothetis, & Leopold, Current Biology, 13, 1076–1085, 2003). Because persistence did not depend on presentation order for any pair of objects, it provides a commutative measure for the similarity of object shapes. However, it is not clear exactly which features or aspects of object shape determine similarity. At least, we did not find simple, low-level features (such as volume overlap, heterogeneity, or rotational symmetry) that could have accounted for all observations. Accordingly, it seems that sensory memory of SFM (which underlies priming of ambiguous rotation) engages higher-level representations of object surface and shape.
Similar content being viewed by others

Perceptual priming is an implicit form of memory, involving neither conscious recollection nor active maintenance (Wiggs & Martin, 1998). Typically, perceptual priming persists over longer periods (minutes to days) and across many intervening stimuli (Schacter, Wig, & Stevens, 2007). Although the behavioral effects of priming are often facilitatory (i.e., prior exposure enhances performance, Schacter et al., 2007), its presumed neural correlates are typically suppressive (“repetition suppression”; Desimone, 1996; Grill-Spector, Henson, & Martin, 2006; but see Segaert, Weber, De Lange, Petersson, & Hagoort, 2013). These characteristics distinguish priming from perceptual adaptation, which persists over shorter periods (seconds), is disrupted by intervening stimuli, and reduces both behavioral performance and neural responses (Clifford et al., 2007; Webster, 2011). The perceptual functions of neither priming nor adaptation are well understood, but both may facilitate perceptual learning in the context of perceptual inference (Clifford et al., 2007; Friston, 2005; Hohwy & Kallestrup, 2008). Priming may help ensure object persistence in the face of occlusions or eye movements (Flombaum & Scholl, 2006; Hollingworth & Franconeri, 2009; Richard, Luck, & Hollingworth, 2008; Tas, Moore, & Hollingworth, 2012).
When physical stimulus features are altered between presentations, some of the neural effects of perceptual priming may diminish, thereby revealing the neural sites of “feature-specific representations” (Schacter et al., 2007). For example, changing object features such as shape, viewpoint, illumination, size and position diminishes “repetition suppression” more in earlier than in higher visual areas (Grill-Spector et al., 1999; Schacter, Dobbins, & Schnyer, 2004; Vuilleumier, Henson, Driver, & Dolan, 2002; Vuilleumier, Schwartz, Duhoux, Dolan, & Driver, 2005). Following the same logic, selective adaptation has been used to identify neural representations of size and orientation of gratings (Blakemore & Campbell, 1969), three-dimensional shape (Preston, Kourtzi, & Welchman, 2009), face identity (Leopold, O’Toole, Vetter, & Blanz, 2001), and other object features (reviewed by Malach, 2012). Interestingly, the neural correlates of feature-specific priming and feature-specific adaptation may not be identical (Schwiedrzik et al., in press).
A particularly prominent form of perceptual priming has long been observed in the context of ambiguous, multistable visual displays (Adams, 1954; Leopold, Wilke, Maier, & Logothetis, 2002; Orbach, Ehrlich, & Heath, 1963; Ramachandran & Anstis, 1983). The instantaneous appearance of such displays is exquisitely sensitive to how they were perceived at earlier points in time (Brascamp et al., 2008; Pastukhov & Braun, 2008). As a result, facilitatory, long-lasting perceptual priming can stabilize the appearance of an ambiguous display almost indefinitely, provided the experimental paradigm minimizes interference from perceptual adaptation (i.e., the suppressive effects of prior exposure) (Leopold et al., 2002; Pastukhov & Braun, 2008; Pearson & Brascamp, 2008). The same putative mechanism has also been termed a sensory memory of multistable displays (Pearson & Brascamp, 2008).
Facilitatory priming builds up in seconds, decays over minutes (Brascamp et al., 2008; Pastukhov & Braun, 2008), and persists across intervening stimuli (Maier, Wilke, Logothetis, & Leopold, 2003). Priming effects with similar time courses have been documented also with unambiguous displays (Bar & Biederman, 1999; Campana, Cowey, & Walsh, 2002; Tanaka & Sagi, 1998). Accordingly, results obtained with ambiguous displays may well generalize to other forms of perceptual priming (Pearson & Brascamp, 2008).
Here we capitalize on the susceptibility of multistable perception to facilitatory priming in order to characterize the stimulus selectivity of such priming effects. For ambiguous objects rotating in depth, perceptual facilitation is known to be specific to retinal location (Chen & He, 2004; Knapen, Brascamp, Adams, & Graf, 2009) and to axis of rotation (Maier et al., 2003). Surprisingly, changes in the shape, size, angular speed, or color of such objects does not seem to reduce perceptual facilitation (Chen & He, 2004; Maier et al., 2003); see Table 1. This lack of specificity in the perceptual facilitation of ambiguous objects contrasts with the extensive specificity (mentioned above) to shape, viewpoint, illumination, size and position of the neural suppression of non-ambiguous objects (Grill-Spector et al., 1999; Schacter et al., 2004; Vuilleumier et al., 2002; Vuilleumier et al., 2005).
To resolve this issue and to establish whether or not the perceptual facilitation of ambiguous objects exhibits a comparable degree of specificity as the neural suppression of non-ambiguous objects, we re-investigated perceptual priming with ambiguous objects rotating in depth (structure-from-motion, or SFM; Sperling & Dosher, 1994; Wallach & O’Connell, 1953). We presented a variety of three-dimensional objects in randomized order, changing object shape, adding or subtracting object parts, replacing hollow with solid objects, or vice versa, between presentations. To minimize suppressive effects between successively presented shapes, we interspersed sufficiently long blank intervals (Adams, 1954; Leopold et al., 2002; Orbach et al., 1963; Ramachandran & Anstis, 1983). To assess perceptual facilitation, we established the average perceptual appearance of any given ambiguous object as a function of the type and appearance of preceding objects.
Our results show that perceptual facilitation is highly specific to object shape and depends strongly on the correspondence between preceding and succeeding objects. The strength of perceptual facilitation varies with the overall similarity between object shapes and with their volumetric properties (hollow or solid). As these results could not be explained by shared low-level features, it would seem that sensory memory of SFM involves higher-level representations of object surfaces and shapes. We conclude that perceptual facilitation provides a novel and sensitive means for probing the perceptual representation of three-dimensional shape. The present results contradict a previous report that sensory memory of SFM lacks shape-specificity (Maier et al., 2003).
General method
Observers
All participants had normal or corrected-to-normal vision. Apart from the authors, observers were naive to the purpose of the experiments and were paid for their participation. Procedures were approved by the medical ethics board of the Otto-von-Guericke Universität, Magdeburg: “Ethik-Komission der Otto-von-Guericke-Universität an der Medizinischen Fakultät.”
Apparatus
Stimuli were generated with MATLAB using the Psychophysics Toolbox (Brainard, 1997) and displayed on a CRT screen (Iiyama VisionMaster Pro 514, iiyama.com) with a spatial resolution of 1,600 × 1,200 pixels and refresh rate of 100 Hz. The viewing distance was 73 cm, so that each pixel subtended approximately 0.019°. In all experiments, background luminance was kept at 36 cd/m2 and environmental luminance at 80 cd/m2.
Stimuli
All shapes measured 5.7° in height. The diameter of the individual dots was 0.057° and their luminance was 110 cd/m2. Rotation speed was kept constant at 0.2 Hz.
Procedure
The presentation schedule used in all experiments is depicted schematically in Fig. 1a (see also Movie 1). The stimuli were presented intermittently with presentation intervals labeled as T on (0.5, 1, 2.5 s) and blank intervals labeled as T off (1 s). The duration of T on interval was chosen for each observer such as to bring the average probability of survival of the illusory rotation (P surv, across all trials) into the [.7, .8] range. P surv is the probability that the same direction of illusory rotation would be reported in two successive trials T(i) and T(i + 1).

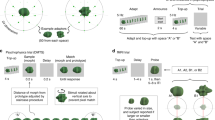
Schematic procedure. (a) Each block consisted of 120 T on and 120 T off intervals, T on ∈ [0.5, 1, 2.5 s], T off = 1 s. The stimuli were presented in pseudorandom order, ensuring that all shape pairs occurred equally often. Observers continuously reported on the perceived direction of rotation. See also Movie 1. (b) The probability that the same shape was repeated remained ~.25 for all lags (only lags up to 20 are shown)
A single block consisted of 120 T on and 120 T off intervals. Shapes were presented in pseudo-random order, ensuring that all shape pairs occurred equally often (one-back history randomization). The probability that same shape was presented both on trial T(i) and on trial T(i + Lag) was approximately .25 for all lag values (Fig. 1b, t test for mean of distribution μ = .25, p > .05).
Experiment 1: Various shapes
Our first experiment aimed to assess the extent to which sensory memory of SFM is specific to shape. To maximize the chances of observing shape-specific effects, we used four very different shapes, termed “hourglass,” “spinning top,” “tilted cross,” and “bent band” (see Fig. 2a and Movies 2–5). These four shapes were presented intermittently and in pseudorandom order, ensuring that all possible pairs of shapes succeeded each other equally often (see the General Method and Fig. 1). Observers reported the direction of illusory rotation of each shape.

Experiment 1. (a) Static snapshots of the stimuli used in the experiment, as if they were seen on the screen (front view, x–y plane). In the main text, the shapes are referred as “hourglass,” “spinning top,” “tilted cross,” and “bent band” (from left to right). See Movies 2–5. (b) Probability that the same illusory rotation was reported on two consecutive trials, as a function of the shapes presented during trials T(i) and T(i + 1)
Our chosen measure for the sensory memory for illusory rotation was P surv, which is the probability that same direction of illusory rotation was reported in two successive trials T(i) and T(i + 1). For convenience, we sometimes term the preceding trials T(i) “prime” trials and the succeeding trials T(i + 1) “probe” trials. Values of P surv > .5 indicated perceptual persistence due to sensory memory, whereas values of P surv < .5 indicated perceptual suppression due to neural fatigue (Kang & Blake, 2010; Pastukhov & Braun, 2011). Values near .5 were more difficult to interpret: they may have indicated either the complete absence of priming effects, or the cancellation of positive and negative effects. Note that due to observer-specific bias (Carter & Cavanagh, 2007) baseline level P surv may be different from .5. Our analysis concentrated on relative effects, sidestepping this issue.
Method
Five observers (three females, two males), including the first and second authors, participated in this experiment. Trials with no responses (no keypress) or with multiple responses (multiple keypresses) during the T on(i) and T off(i) intervals were discarded (4.5 % of total). For further details, please refer to the General Method.
Structure-from-motion (SFM) stimuli consisted of 500 dots distributed randomly over the surfaces of three shapes (“hourglass,” “spinning top” and “bent band” stimuli) or throughout the volume of one shape (“tilted cross”). Please see Fig. 2a and Movies 2–5.
In studying sensory memory, observer-specific bias presents a possible confound (Carter & Cavanagh, 2007), as it would increase perceptual stability of the favored rotational appearance even in the absence of memory. Note, however, that any such effect would apply equally to all sequences of shapes and thus would tend to decrease, not increase, congruency effects (i.e., differences between similar and dissimilar sequences of shapes).
In the present series of experiments, observer-specific bias did not significantly influence the results. Observer-specific bias may be defined as
where N left and N right are number of trials with reported rotation, respectively, to the left and to the right. Bias values range from 0 (both rotations reported equally often) to 1 (same rotation reported on all trials). Our observers exhibited uniformly moderate bias values (.42 ± .07). A two-way analysis of variance (ANOVA) of bias values, with Observer Identity and Shape Identity as independent factors, showed no significant main effects [F(4, 12) = 2.37, p = .11, for observer; F(3, 12) = 1.39, p = .3, for shape], demonstrating that bias values were uniform across observers and shapes. In addition, bias values were not correlated with perceptual stability \( \left[ {P_{\mathrm{surv}}^{\mathrm{same}}\left( {\mathrm{observer},\mathrm{shape}} \right):R=.28,p=.24} \right] \). In conclusion, there is no evidence that observer- or shape-specific bias influenced the reported results.
Results and discussion
The results of Experiment 1 revealed that shape identity strongly influenced the persistence of illusory rotation (Fig. 2b). When identical shapes were presented in succession, P surv was consistently higher than when two different shapes were used: \( P_{\mathrm{surv}}^{\mathrm{same}}=.86\pm .06 \), as compared to \( P_{\mathrm{surv}}^{\mathrm{diff}}=.56\pm .08 \) [right-tailed t test, t(4) = 3.4, p = .02].
We analyzed history effects using Identity of “Prime” i, Identity of “Probe” i + 1, and Prime–Probe Congruency as independent factors. We also included Observer Identity as an independent factor. Although we attempted to obtain comparable perceptual stability with all observers (by adjusting the duration of T on intervals), some residual differences in the average P surv remained. Including Observer Identity as an independent factor allowed us to focus the analysis on differences between conditions (within each observer), rather than on differences between individual observers.
A four-way ANOVA showed the strongest main effect for Prime–Probe Congruency [F(1, 53) = 43, p < 10–7]. The effect of Probe Identity was marginally significant [F(3, 53) = 3, p = .04] and the effect of Prime Identity was not significant [F(3, 53) = 1.6, p = .9].
In addition, we performed a two-way ANOVA separately for each pair of shapes. Here, the independent factors were Prime–Probe Congruency and Observer Identity. The results, which are summarized in Table 2, revealed significant effects of Prime–Probe Congruency in all cases [all F values are above the Bonferroni-corrected critical value of F(1, 11) = 10.3]. However, the size of these effects varied greatly between pairs. The largest difference between congruent and noncongruent shapes was observed for the pairing of “bent band” and “spinning top.” In this case, there was no detectable persistence of rotation between either “bent band” and “spinning top” [P surv = .46 ± .06, t(3) = −0.45, p = .68] or “spinning top” and “bent band” [P surv = .51 ± .08, t(3) = 0.09, p = .68; t test for mean of distribution μ = .5]. In contrast, the smallest difference was observed for the pairing of “tilted cross” and “bent band,” in which case there was significant persistence of rotation between both “tilted cross” and “bent band” [P surv = .84 ± .07, t(3) = 10.2, p = .002] and between “bent band” and “tilted cross” [P surv = .81 ± .06, t(3) = 9.9, p = .002; t test for mean of distribution μ = .5].
The comparison of these two pairings (“bent band” with “spinning top” versus “bent band” with “tilted cross”) seems to suggest that the persistence of illusory rotation depended on the similarity of the shapes in question. Whereas “bent band” and “tilted cross” were similar shapes (and exhibited persistence), “bent band” and “spinning top” were not (and did not exhibit persistence). Accordingly, we hypothesize that it is the similarity of SFM shapes that determines the strength of prime/probe interactions.
Maier et al. (2003) demonstrated that the sensory memories induced by ambiguous displays are robust to the intervening presentation of other kinds of ambiguous displays at the same location. We wondered whether the same is true for sensory memories induced by different SFM shapes. To answer this question, we repeated the ANOVA for longer lags, restricting the independent factors to Prime–Probe Congruency and Observer Identity. The analysis results are plotted in Fig. 3a separately for cases in which shape i + lag was either an “hourglass” or “spinning top” (solid line) or was either a “bent band” or “tilted cross” (dashed line). For lags of up to 14 trials, the probability that same direction of illusory rotation was reported on trial i – lag and on trial i was significantly higher for identical than for dissimilar shapes. This surprising result suggests that shape-specific sensory memories for illusory rotation are robust to the intervening presentation of multiple other SFM shapes at the same location.

Prime–Probe Congruency effect for longer lags. (a) Main effect of Prime–Probe Congruency for various lags (two-way ANOVA with Prime–Probe Congruency and observer identity as independent factors). Solid line: T(i + Lag) shape was either “hourglass” or “spinning top.” Dashed line: T(i + Lag) shape was either , “bent band” and “tilted cross.” (b, c) Survival probability when the same shape (\( P^{{same}}_{{surv}} \), open circles) or a different shape (\( P^{{diff}}_{{surv}} \), filled circles) was presented at trial T(i + Lag). (b) T(i + Lag) shape was either a “bent band” or “tilted cross.” (c) T(i + Lag) shape was either an “hourglass” or “spinning top”
Curiously, for rotationally asymmetric shapes (“bent band” and “spinning top”), the effect on P surv of an identical shape having been presented on an earlier trial exhibited a periodicity of approximately five trials (Fig. 3b, open circles, “bent band” and “spinning top” were analyzed together). The survival probability was significantly higher for lags of multiples of five trials (5, 10, 15, and 20) than for the survival probability of an immediately preceding lags (4, 9, 14, and 19) [t(38) = 3.6, p = .0008, two sample t test]. No such periodicity was evident for rotationally symmetric shapes (“spinning top” and “hour glass” analyzed together, Fig. 3c) or when shapes were dissimilar (Fig. 3b, filled circles). Accordingly, it seems likely that this periodicity reflected an additional stabilization by a sensory memory of illusory depth. Because the rotational frequency was 0.2 Hz, shapes appeared at the same angle of rotation every five trials (at least for most observers). Thus, when a prime shape was succeeded by a probe shape after four intervening trials, shape-specific memories of both illusory rotation and illusory depth were able to enhance P surv.
Experiment 2: Band stimuli
The results of our first experiment suggested that the similarity of two shapes influenced the persistence of illusory rotation between them. In our second experiment, we sought to investigate shape similarity in a more systematic manner. To this end, we chose an ordered set of shapes in which each shape was “contained within” all preceding shapes: “sphere,” “quad band,” “dual band,” and “single band” (see Fig. 4a and Movies 6–9). Since any two could be transformed into each other by adding or subtracting “bands,” the number of additions or subtractions separating two shapes provided a straightforward measure of “similarity.”

Experiment 2: Band stimuli. (a) Static snapshots of the stimuli, as if seen on the screen (top row: front view, x–y plane) and from above (bottom row: top view, x–z plane). Black lines show the 0° angle of rotation. See also Movies 6–9. (b) Probability of survival of the direction of illusory rotation as a function of shapes presented on two consecutive trials T(i) and T(i + 1). (c) Main effect of Prime–Probe Congruency for various lags (two-way ANOVA with Prime–Probe Congruency and Observer Identity as independent factors). The analysis was restricted to the sphere or quad-band shapes. (d) Probability of survival of illusory rotation as a function of the “dissimilarity” measure between two shapes. This measure was defined as 0 for the same shape, 1 for the most similar shapes (e.g., sphere and quad-band, dual band and single band), 2 for less similar shapes (e.g., sphere and dual band, quad band and single band), and 3 for the most different shapes (sphere and single band). The stability of illusory depth is inversely proportional to the dissimilarity measure, Spearman rank correlation R = –.76, p < 10–14
Method
Five observers (three females, two males), including the second author, participated in the experiment.
SFM stimuli consisted of 500 dots distributed randomly over the surface of four shapes: a sphere, a quad band, a dual band, and a single band (see Fig. 4a and Movies 6–9). Trials with no responses or multiple responses were discarded (<1 % of total). For further details, please refer to the General Method.
Observer-specific bias was moderate (.19 ± .07) and uniform. A two-way ANOVA, with Observer Identity and Shape Identity as independent factors, showed no significant main effects: F(4, 12) = 2.86, p = .07 (observer); F(3, 12) = 0.81, p = .51 (shape). Bias strength was not correlated with perceptual stability: R = −.27, p = .24. See the Method section of Experiment 1 for details.
Results and discussion
As expected, the results confirmed that persistence of illusory rotation from one shape to another depended on the similarity of the shapes in question (Fig. 4b). Although P surv was consistently largest for identical shapes (\( P^{{same}}_{{surv}} \) = .87 ± .04), it remained significantly above chance when different but similar shapes were used \( \left( {P_{\mathrm{surv}}^{\mathrm{diff}}=.74\pm .06} \right) \) [right-tailed t test: t(4) = 5.6, p = .002].
As in Experiment 1, we performed a four-way ANOVA with Observer Identity, Identity of “Prime” T(i), Identity of “Probe” T(i + 1), and Prime–Probe Congruency as independent factors. As before, the strongest main effect was obtained for Prime–Probe Congruency [F(1, 68) = 24.4, p < 10–5], with no significant effect of either Probe Identity [F(3, 68) = 0.59, p = .63] or Prime Identity [F(3, 68) = 0.61, p = .61]. Similarly, analysis of the Prime–Probe Congruency effect showed that it remained significant for up to 17 trials (Fig. 4c, two-way ANOVA with Observer Identity and Prime–Probe Congruency as independent factors, with analysis restricted to sphere or quad-band shapes). A multitrial analysis performed on single-band shapes showed a periodicity of five trials (data not shown), similar to that observed for the “bent band” and “tilted cross” in Experiment 1.
Assigning Indices 1 through 4, respectively, to the “single band,” “dual band,” “quad band,” and “sphere,” we could use the absolute value of the difference between indices as a (crude) measure of the dissimilarity of two shapes. In this way, “single band” and “sphere” were assigned a dissimilarity index of 3, whereas a “dual band” and a “single band” have a dissimilarity index of 1. Two identical shapes have a dissimilarity index of zero.
When persistence of illusory rotation is plotted against this dissimilarity index (Fig. 4d), a significant and strong correlation is obtained (R = –.76, p < 10–14; Spearman rank correlation), confirming the suspected importance of similarity between shapes. Of course, a more sophisticated measure of dissimilarity might have resulted in an even stronger correlation.
Inverting the argument, we can use persistence of illusory rotation as an empirical measure for the similarity of two shapes. To this end, we again performed a two-way ANOVA separately for each pair of shapes (Table 3). The results suggest that a “dual band” is more similar to a “single band” than to a “quad band,” whereas a “quad band” is almost indistinguishable from a “sphere.” Such an analysis offers a general and potentially useful measure of similarity for the perceptual representations of various SFM shapes.
Experiment 3: Surface versus volume
The previous results suggest that sensory memory of SFM can be used to probe the structure of perceptual representations of SFM shapes. Identical shapes (with fully overlapping representations) are expected to result in greater persistence (P surv) than similar shapes (with partially overlapping representations), which in turn are expected to result in greater persistence than dissimilar shapes (with non-overlapping representations). In other words, the persistence of sensory memory should provide an empirical measure for the overlap of perceptual representations.
As a first application of this novel method, we wondered whether the representation of SFM shapes is based on surfaces or on volumes. The perceptual difference between SFM shapes defined exclusively by surface dots (“hollow shapes”) and SFM shapes defined by dots distributed throughout their volume (“solid shapes”) would seem to be slight, as the salient differences between shapes are defined by their respective surfaces. Accordingly, one might expect that surface properties, rather than hollowness or solidity, dominate the perceptual representation of SFM shapes.
To address this issue, we investigated sensory memory between “solid sphere,” “solid cylinder,” “hollow sphere,” and “hollow cylinder” (see Movie 6 and Movies 10–12). If perceptual representations are surface-based, one would expect persistence between spheres and spheres, or between cylinders and cylinders, irrespective of their hollowness or solidity. If, however, perceptual representations are volume-based, one would expect the opposite outcome. Expect for the shapes used, the experimental procedure was identical to Experiments 1 and 2.
Method
Five observers (three females, two males), including the first and second authors, participated in the experiment.
Structure-from-motion (SFM) stimuli consisted of 500 dots distributed randomly over the surfaces of two shapes (“hollow sphere, “hollow cylinder” stimuli) or throughout the volume of two shapes (“filled sphere,” “filled cylinder” stimuli). Please see Movie 6 and Movies 10–12. Trials with no responses or multiple responses were discarded (<1 % of total). For further details, please refer to the General Method.
Observer-specific bias was moderate (.22 ± .06) and uniform. A two-way ANOVA, with Observer Identity and Shape Identity as independent factors, showed no significant main effects: F(4, 12) = 1.43, p = .28 (observer); F(3, 12) = 0.33, p = .8 (shape). Bias strength was not correlated with perceptual stability: R = −.05, p = .84. See the Method section of Experiment 1 for details.
Results and discussion
Surprisingly, the results revealed that persistence of illusory rotation depended more on the solidity or hollowness of a SFM shape than on its form (sphere or cylinder) (Fig. 5a). When two hollow (or two solid) shapes were presented in succession, P surv was consistently higher \( \left( {P_{\mathrm{surv}}^{{\mathrm{same}\ \mathrm{fill}}}=.79\pm .06} \right) \) than when two spheres (or two cylinders) followed each other \( \left( {P_{\mathrm{surv}}^{{\mathrm{same}\ \mathrm{shape}}}=.72\pm .06} \right) \). To examine these results in more detail, we performed a seven-way analysis of variance with the following independent factors: Observer Identity, Prime Form (sphere or cylinder), Probe Form, Prime Fill (hollow or solid), Probe Fill, Form Congruency, and Fill Congruency. The results are summarized in Table 4 and confirm that the fill congruency is the only significant factor. Analysis over multiple trials showed that fill congruency remained a significant factor for lags of approximately 20 trails (Fig. 5b, two-way ANOVA with Observer Identity and Fill Congruency as independent factors). In contrast to Experiments 1 and 2, the effect of form congruency was not significant.

Experiment 3: Surface versus full volume. (a) Probability of survival of the direction of illusory rotation, as a function of the object presented on two consecutive trials T(i) and T(i + 1). Stability of the illusory rotation depends primarily on the stability of the surface/filled volume characteristic. (b) Main effect of prime–probe fill congruency for various lags (two-way ANOVA with Prime–Probe Fill Congruency and Observer Identity as independent factors)
However, two-way analyses of variance performed separately for each pair of shapes (Table 5) suggested that form (sphere or cylinder) did play a role, even if only for hollow shapes [F(1, 11) = 13.9, p = .002]. For solid shapes, the interaction was not significant [F(1, 11) = 1.7, p = .21]. Apparently, hollow volumes of different form are perceptually more dissimilar than solid volumes of different form. The interaction between solid and hollow shapes was consistently significant, except for the pairing of solid sphere with hollow cylinder.
Combined analysis of results
For any given object pair and observer, the degree of priming is measured most directly by \( P^{{same}}_{{surv}} \) of the apparent rotation. However, across observers, a given object pair (e.g., A→B) exhibits somewhat different degrees of priming. Accordingly, a comparison of different object pairs (e.g., A→B with C→D) is best accomplished in statistical terms. To correct for variability across observers and experiments, we normalized the priming by object A on object B (A→B) with respect to priming by object on itself (B→B). Specifically, we used a pairwise t test to compare priming A→B with priming B→B, across observers. The p value of this comparison provides a statistical measure of relative priming. In terms of this measure, the degree of priming A→B (relative to B→B) may be compared to the degree of priming C→D (relative to D→D).
Since relative priming depended on object order, we also defined an alternative measure termed symmetric priming. This measure compared priming between A→B and B→A, on the one hand, and A→A and B→B, on the other hand, with a two-sample t test.
In comparing different experiments, we wondered whether the results might depend systematically on low-level object properties. In particular, we were curios whether the observed priming effects could be explained in terms of a low-level representation of local velocity gradients, either in three-dimensional space or in the image plane. (For simplicity, we are going to conflate these two possibilities—three-dimensional space and image plane—and formulate the argument in terms of three-dimensional volumes.) In such a low-level representation, a probe object would be composed of numerous local component volumes, which would be primed independently. The overall degree of priming would then depend on the fraction of component volumes that are stimulated both by a preceding (prime) and a succeeding (probe) object. In other words, the degree of priming should depend on the “volume overlap” between the prime and probe objects (Fig. 6a). Note that, in our displays, a given component volume would always be stimulated by the same velocity, because all objects rotate with the same angular velocity.

Combined analysis of the results: Relative priming. (a) “Volume overlap” between prime and probe objects. Gray dots indicate the volumes of prime object T(i) and probe object T(i + 1), and black dots mark the part of volume T(i + 1) that overlaps volume T(i). (b) Relative priming A→B as a function of relative overlap (see the text for details)
Two variants of this scenario seem plausible. First, the degree of priming might depend on the total number of primed component volumes, irrespective of the number of unprimed component volumes. Second, the degree of priming might depend on the relative number of primed and unprimed component volumes. Accordingly, we computed both the total overlap (intersection V prime ∩ V probe) and the relative overlap (intersection V prime ∩ V probe divided by V probe) of all object pairs. Finally, as relative overlap depended on object order, we also defined a symmetric measure of volume overlap, which we termed symmetric overlap (intersection V prime ∩ V probe divided by union V prime ∪ V probe).
Somewhat to our surprise, relative priming failed to correlate significantly with either total overlap or relative overlap (R = −.03, p = .85, Fig. 6b). This was in spite of the fact that fractional overlap values ranged from a few percent (for combinations of “bent band” or “tilted cross” with hollow objects such as “spinning top” or “hourglass”) to 100 % (for identical solid objects). Even when the results of each experiment were analyzed separately, no significant correlations were observed: R = −.38, p = .23 (Exp. 1); R = −.21, p = .52 (Exp. 2); R = .39, p = .2 (Exp. 3).
Yet another hypothesis considered was that object similarity might depend on the degree of rotational symmetry. However, pairs of rotationally symmetric objects (e.g., “sphere” and “cylinder”) or rotationally asymmetric objects (e.g., “bent band” and “tilted cross”) did not prime each other to any greater degree than mixed pairs (e.g., “diamond” and “tilted cross,” or “sphere” and “band”), as the reader can see by closely inspecting Fig. 6b. In addition, priming between solid and hollow objects (spheres or cylinders) tended to be weak, although both were rotationally symmetric.
Computing similarity from distributions of other local motion components proved equally unsuccessful, no matter whether using local motions in the image plane, local motions perpendicular to the image, or local motions perpendicular to the shape surface. No significant correlation between any of these similarity measures and symmetric priming was obtained (all Rs < .18, p > .5).
The only clear systematic trend in our results was independence of object order. In other words, priming was comparable for A→B and B→A in all cases, as is shown in Fig. 7a. The correlation between P surv(A→B) and P surv(B→A) was highly significant (R = .97, p < 10–10) and a paired-sample t test showed no significant order effect for any object pair (with either the Bonferroni corrected [α = .0028] or uncorrected [α = .05] criterion).

Combined analysis of the results: Symmetric priming. (a) Persistence of apparent rotation in “forward” order, P surv(A→B) versus “reverse” order, P surv(B→A). The dark square marks the object pair depicted on Fig. 6a (sphere–band). (b) Symmetric priming as a function of symmetric overlap (see the text for details)
In view of the startling symmetry (independence of object order) of the observed priming effects, we conclude that the degree of priming offers a commutative measure for the “similarity” of two objects. We further surmise that this operative “similarity” simply depends on the number of features (whatever they may be) that are shared between prime and probe objects. It certainly does not depend on the relative fraction of features that are primed, as any fractional dependence would depend on object order (and thus not be symmetric).
The notion of “volume overlap” acquired some explanatory value when symmetric measures were used. We found that symmetric priming was correlated significantly with symmetric overlap when all experiments are considered (R = .76, p = .0002; Fig. 7b). However, when each experiment was considered separately, the correlation was significant only for Experiment 2: R = .34, p = .5 (Exp. 1); R = .88, p = .02 (Exp. 2); R = .4, p = .43 (Exp. 3).
In conclusion, the degree of priming depends on the commutative “similarity” between prime and probe objects. Our results do not reveal on what object features or properties this “similarity” depends, except that it does not seem to depend on low-level (local) properties such as volume overlap, shape heterogeneity, or depth symmetry. At the same time, the dependence of priming on volumetric object properties (solid or hollow) shows that the relevant features are not entirely detached from the physical instantiation and thus are not at a quasi-semantic level (prototypical cylinder, sphere, etc.). Accordingly, we surmise that perceptual priming engages an intermediate-level representation of shapes and surfaces.
General discussion
Perceptual priming is of interest as an implicit memory system (Grill-Spector et al., 2006; Schacter et al., 2007; Wiggs & Martin, 1998). A particular form of perceptual priming occurs in ambiguous displays, where past appearance strongly biases future perception (Adams, 1954; Leopold et al., 2002; Orbach et al., 1963; Ramachandran & Anstis, 1983). We studied this “sensory memory” (Pastukhov & Braun, 2008; Pearson & Brascamp, 2008) for perceptual appearance, aiming to throw light on the underlying sensory representations and on the relation to other priming phenomena (e.g., perceptual adaptation, repetition suppression).
Specifically, we generated the appearance of three-dimensional objects rotating in depth with structure-from-motion (SFM) displays. A sensory memory of the apparent rotation persisted over longer periods (>1 min) and multiple intervening objects (>15), significantly biasing the apparent rotation of subsequent objects. Most interestingly, the degree of persistence was modulated strongly by object shape, with the largest effects obtained for identical object pairs, intermediate effects for similar object pairs, and little or no effect for dissimilar objects pairs. These results demonstrate unequivocally that sensory memory of SFM is specific for object shape, contrary to an earlier report (Maier et al., 2003).
Probing sensory memory
The observed pattern of specificities afforded some clues as to the nature of the representations that underlie sensory memory. Admittedly, the relation between behavioral manifestations of adaptation/priming and their correlates in neuroimaging data or single cell responses is complex (Grill-Spector et al., 2006; Henson & Rugg, 2003; Sawamura, Orban, & Vogels, 2006). In addition, the neural substrates of neither sensory memory in multistable displays (Brascamp, Kanai, Walsh, & Van Ee, 2010; Braun & Mattia, 2010; Sterzer & Rees, 2008) nor facilitatory priming in general are known (Grill-Spector et al., 2006; Henson & Rugg, 2003). Nonetheless, our results do help us to obtain a general characterization of representations involved in sensory memory of illusory rotation.
On the one hand, sensory memory did not seem to engage low-level representations of local component volumes. If this had been the case, the volume overlap between successive objects should have predicted the priming effects, which it did not. On the other hand, sensory memory also did not seem to engage high-level representations of canonical objects shapes (ring, sphere, cylinder, etc.). For if this had been the case, hollow and solid objects of the same shape should have interacted strongly, which they did not. Instead, sensory memory seemed to involve intermediate-level representations of object surfaces and/or shapes. Presumably, a variety of surface and/or shape properties such as hollowness, solidity, rotational symmetry or asymmetry, or nature and degree of articulation all have a part to play.
Importantly, sensory memory proved independent of object order and thus provided a commutative measure of “similarity” between objects. To our mind, this symmetry suggests several things: Firstly, the effect of sensory memory grows with the degree of overlap between the sensory representations of successive objects, with similar objects overlapping more, dissimilar objects overlapping less. Secondly, sensory representations of object shape involve a large variety surface and/or shape properties, so that the overlap can vary continuously along multiple dimensions of similarity/dissimilarity. If only a few properties had been involved, the overlap would have varied discontinuously. Thirdly, the various surface/shape properties form a functionally homogeneous representation, in that they contribute comparably to sensory memory. If particular properties had dominated, memory effects would again have varied discontinuously.
Comparing perceptual adaptation
The experimental approach adopted here is analogous to the use of perceptual adaptation as a probe for neural and perceptual representations. In perceptual adaptation, prolonged viewing (>10 s) of a particular object reduces neural responsiveness to, and distorts the perception of, subsequent presentations of the same or similar objects. The former effect has been used widely to probe neural object representation with fMRI (Huk & Heeger, 2002; Preston et al., 2009) and the latter effect was proven valuable in characterizing the representations of features such as numerosity (Burr & Ross, 2008) or face identity (Leopold et al., 2001). In case of ambiguous displays, the latter effect biases perception in favor of alternative (unadapted) appearances (Nawrot & Blake, 1989; Pastukhov & Braun, in press; Wolfe, 1984).
The combined effects of facilitatory priming and of perceptual adaptation on the ambiguous appearance of multistable displays have been modeled in terms of a single, sensory representation (Noest, van Ee, Nijs, & van Wezel, 2007; Noest & van Wezel, 2012). However, it is equally possible to model these effects in terms of an interaction between sensory and memory representations (Gigante, Mattia, Braun, & Del Giudice, 2009). It is far from evident that facilitatory priming and perceptual adaptation exhibit identical or even comparable specificities (and thus engage similar representations). In fact, the negative motion aftereffect induced by SFM is not shape-specific (Nawrot & Blake, 1991), in contrast to the positive sensory memory investigated here. A recent report dissociating the neural correlates of facilitatory priming and of perceptual adaptation points to a similar conclusion (Schwiedrzik et al., in press). To settle this issue, further work comparing the respective shape-specificities of facilitatory priming and perceptual adaptation is required.
Neural substrates of sensory memory
The neural substrates of sensory memory for ambiguous appearance have been studied in several situations. The sensory memory for apparent rotation of SFM displays can be disrupted by TMS of cortical area hMT/V5 (Brascamp et al., 2010), which is known to play a prominent role in processing visual motion (Born & Bradley, 2005; Campana et al., 2002; Orban, 2011; Rokers, Cormack, & Huk, 2009; Tootell et al., 1995). In the context of binocular rivalry, a sensory memory for faces appears to engage the fusiform face area (FFA), as well as several fronto-parietal regions (Sterzer & Rees, 2008).
The neural representation of three-dimensional shape is thought to involve the superior part of lateral occipital complex (LOC; Kourtzi, Erb, Grodd, & Bülthoff, 2003; Kourtzi & Kanwisher, 2001; Murray, Olshausen, & Woods, 2003) and/or the superior temporal sulcus (Beer, Watanabe, Ni, Sasaki, & Andersen, 2009); see Orban (2011) for a review.
Of course, the present findings—sensory memory for apparent rotation is specific to three-dimensional shape—imply that three-dimensional rotation and three-dimensional shape are not represented entirely independently and must exhibit some degree of “binding.” About the mechanisms of such “binding” one can only speculate at this point. Elsewhere, we have argued that a “nested” attractor dynamics would provide a plausible substrate for cooperativity within heterogeneous representations, especially when these may be anatomically dispersed (Braun & Mattia, 2010; Gigante et al., 2009).
Related forms of sensory memory
A sensory memory that resolves perceptual ambiguity and favors perceptual stability is observed also in several other contexts (besides the ambiguous SFM displays investigated here). For example, the perception of ambiguous motion in Ternus displays is biased by object shape and even by semantic properties (Kramer & Rudd, 1999; Shechter, Hochstein, & Hillman, 1988; Yu, 2000). Similarly, the perception of object continuity in the stream/bounce paradigm is also influenced by shape and other object properties (Caplovitz, Shapiro, & Stroud, 2011; Feldman & Tremoulet, 2006; Kawachi, Kawabe, & Gyoba, 2011). Presumably, these types of sensory memory help to ensure object persistence when visibility is interrupted due to occlusion (Flombaum & Scholl, 2006; Hollingworth & Franconeri, 2009) or during eye movements (Richard et al., 2008; Tas et al., 2012). In all of these instances, sensory memory is not object-selective in an “all-or-nothing” manner. Instead, the effect of prior exposure is gradual and grows with the similarity of two objects.
Specificity to features other than shape?
As we mentioned, previous studies have concluded that sensory memory of apparent rotation does not depend on object properties such as shape, size, or color (Chen & He, 2004; Maier et al., 2003). Our successful demonstration of shape specificity raises the question whether more refined experimental approaches might reveal also specificities to object size or color.
In ambiguous displays, object shape is comparatively persistent in that reversals of apparent shape are prohibited even when prompted by stimulus transients (Pastukhov & Braun, 2013; Pastukhov, Vonau, & Braun, 2012). By comparison, object rotation is far more “flexible” in that reversals of apparent rotation are readily prompted by stimulus transients (Pastukhov et al., 2012; Stonkute, Braun, & Pastukhov, 2012). Accordingly, object shape may be treated as an invariant and may carry particular weight in determining the continuity and persistence of objects.
On the other hand, more sensitive experimental approaches may well reveal specificities to object size or color. For example, in the case of ambiguous object translation (streaming/bouncing), object size, color, or luminance are just as disambiguating as object shape (Caplovitz et al., 2011; Feldman & Tremoulet, 2006; Kawachi et al., 2011).
Conclusions
Sensory memory of apparent rotation in structure-from-motion (SFM) is specific for object shape. This specificity provides a sensitive measure for the “similarity” of three-dimensional shapes and a novel probe into the nature of its sensory representation.
References
Adams, P. A. (1954). The effect of past experience on the perspective reversal of a tridimensional figure. The American Journal of Psychology, 67, 708–710.
Bar, M., & Biederman, I. (1999). Localizing the cortical region mediating visual awareness of object identity. Proceedings of the National Academy of Sciences, 96, 1790–1793.
Beer, A. L., Watanabe, T., Ni, R., Sasaki, Y., & Andersen, G. J. (2009). 3-D surface perception from motion involves a temporal–parietal network. European Journal of Neuroscience, 30, 703–713. doi:10.1111/j.1460-9568.2009.06857.x
Blakemore, C., & Campbell, F. W. (1969). On the existence of neurones in the human visual system selectively sensitive to the orientation and size of retinal images. The Journal of Physiology, 203, 237–260.
Born, R. T., & Bradley, D. C. (2005). Structure and function of visual area MT. Annual Review of Neuroscience, 28, 157–189. doi:10.1146/annurev.neuro.26.041002.131052
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision, 10, 433–436. doi:10.1163/156856897X00357
Brascamp, J. W., Kanai, R., Walsh, V., & Van Ee, R. (2010). Human middle temporal cortex, perceptual bias, and perceptual memory for ambiguous three-dimensional motion. Journal of Neuroscience, 30, 760–766. doi:10.1523/JNEUROSCI.4171-09.2010
Brascamp, J. W., Knapen, T. H. J., Kanai, R., Noest, A. J., Van Ee, R., & Van den Berg, A. V. (2008). Multi-timescale perceptual history resolves visual ambiguity. PLoS One, 3, e1497. doi:10.1371/journal.pone.0001497
Braun, J., & Mattia, M. (2010). Attractors and noise: Twin drivers of decisions and multistability. NeuroImage, 52, 740–751. doi:10.1016/j.neuroimage.2009.12.126
Burr, D., & Ross, J. (2008). A visual sense of number. Current Biology, 18, 425–428. doi:10.1016/j.cub.2008.02.052
Campana, G., Cowey, A., & Walsh, V. (2002). Priming of motion direction and area V5/MT: A test of perceptual memory. Cerebral Cortex, 12, 663–669.
Caplovitz, G. P., Shapiro, A. G., & Stroud, S. (2011). The maintenance and disambiguation of object representations depend upon feature contrast within and between objects. Journal of Vision, 11(1), 1–14. doi:10.1167/11.14.1
Carter, O., & Cavanagh, P. (2007). Onset rivalry: Brief presentation isolates an early independent phase of perceptual competition. PLoS One, 2, e343. doi:10.1371/journal.pone.0000343
Chen, X., & He, S. (2004). Local factors determine the stabilization of monocular ambiguous and binocular rivalry stimuli. Current Biology, 14, 1013–1017. doi:10.1016/j.cub.2004.05.042
Clifford, C. W. G., Webster, M. A., Stanley, G. B., Stocker, A. A., Kohn, A., Sharpee, T. O., & Schwartz, O. (2007). Visual adaptation: Neural, psychological and computational aspects. Vision Research, 47, 3125–3131. doi:10.1016/j.visres.2007.08.023
Desimone, R. (1996). Neural mechanisms for visual memory and their role in attention. Proceedings of the National Academy of Sciences, 93, 13494–13499.
Feldman, J., & Tremoulet, P. D. (2006). Individuation of visual objects over time. Cognition, 99, 131–165. doi:10.1016/j.cognition.2004.12.008
Flombaum, J. I., & Scholl, B. J. (2006). A temporal same-object advantage in the tunnel effect: Facilitated change detection for persisting objects. Journal of Experimental Psychology. Human Perception and Performance, 32, 840–853. doi:10.1037/0096-1523.32.4.840
Friston, K. J. (2005). A theory of cortical responses. Philosophical Transactions of the Royal Society B, 360, 815–836. doi:10.1098/rstb.2005.1622
Gigante, G., Mattia, M., Braun, J., & Del Giudice, P. (2009). Bistable perception modeled as competing stochastic integrations at two levels. PLoS Computational Biology, 5, e1000430. doi:10.1371/journal.pcbi.1000430
Grill-Spector, K., Henson, R., & Martin, A. (2006). Repetition and the brain: Neural models of stimulus-specific effects. Trends in Cognitive Sciences, 10, 14–23. doi:10.1016/j.tics.2005.11.006
Grill-Spector, K., Kushnir, T., Edelman, S., Avidan, G., Itzchak, Y., & Malach, R. (1999). Differential processing of objects under various viewing conditions in the human lateral occipital complex. Neuron, 24, 187–203.
Henson, R. N. A., & Rugg, M. D. (2003). Neural response suppression, haemodynamic repetition effects, and behavioural priming. Neuropsychologia, 41, 263–270. doi:10.1016/S0028-3932(02)00159-8
Hohwy, J., & Kallestrup, J. (Eds.). (2008). Being reduced. Oxford, UK: Oxford University Press. doi:10.1093/acprof:oso/9780199211531.001.0001
Hollingworth, A., & Franconeri, S. L. (2009). Object correspondence across brief occlusion is established on the basis of both spatiotemporal and surface feature cues. Cognition, 113, 150–166. doi:10.1016/j.cognition.2009.08.004
Huk, A. C., & Heeger, D. J. (2002). Pattern-motion responses in human visual cortex. Nature Neuroscience, 5, 72–75. doi:10.1038/nn774
Kang, M.-S., & Blake, R. (2010). What causes alternations in dominance during binocular rivalry? Attention, Perception, & Psychophysics, 72, 179–186. doi:10.3758/APP.72.1.179
Kawachi, Y., Kawabe, T., & Gyoba, J. (2011). Stream/bounce event perception reveals a temporal limit of motion correspondence based on surface feature over space and time. Perception, 2, 428–439. doi:10.1068/i0399
Knapen, T., Brascamp, J., Adams, W. J., & Graf, E. W. (2009). The spatial scale of perceptual memory in ambiguous figure perception. Journal of Vision, 9(13), 16.1–12. doi:10.1167/9.13.16
Kourtzi, Z., Erb, M., Grodd, W., & Bülthoff, H. H. (2003). Representation of the perceived 3-D object shape in the human lateral occipital complex. Cerebral Cortex, 13, 911–920.
Kourtzi, Z., & Kanwisher, N. (2001). Representation of perceived object shape by the human lateral occipital complex. Science, 293, 1506–1509. doi:10.1126/science.1061133
Kramer, P., & Rudd, M. (1999). Visible persistence and form correspondence in Ternus apparent motion. Perception & Psychophysics, 61, 952–962.
Leopold, D. A., O’Toole, A. J., Vetter, T., & Blanz, V. (2001). Prototype-referenced shape encoding revealed by high-level aftereffects. Nature Neuroscience, 4, 89–94. doi:10.1038/82947
Leopold, D. A., Wilke, M., Maier, A., & Logothetis, N. K. (2002). Stable perception of visually ambiguous patterns. Nature Neuroscience, 5, 605–609. doi:10.1038/nn851
Maier, A., Wilke, M., Logothetis, N. K., & Leopold, D. A. (2003). Perception of temporally interleaved ambiguous patterns. Current Biology, 13, 1076–1085.
Malach, R. (2012). Targeting the functional properties of cortical neurons using fMR-adaptation. NeuroImage, 62, 1163–1169. doi:10.1016/j.neuroimage.2012.01.002
Murray, S. O., Olshausen, B. A., & Woods, D. L. (2003). Processing shape, motion and three-dimensional shape-from-motion in the human cortex. Cerebral Cortex, 13, 508–516.
Nawrot, M., & Blake, R. (1989). Neural integration of information specifying structure from stereopsis and motion. Science, 244, 716–718.
Nawrot, M., & Blake, R. (1991). The interplay between stereopsis and structure from motion. Perception & Psychophysics, 49, 230–244.
Noest, A. J., van Ee, R., Nijs, M. M., & van Wezel, R. J. A. (2007). Percept-choice sequences driven by interrupted ambiguous stimuli: A low-level neural model. Journal of Vision, 7(8), 10. doi:10.1167/7.8.10
Noest, A. J., & van Wezel, R. J. A. (2012). Dynamics of temporally interleaved percept–choice sequences: Interaction via adaptation in shared neural populations. Journal of Computational Neuroscience, 32, 177–195. doi:10.1007/s10827-011-0347-7
Orbach, J., Ehrlich, D., & Heath, H. A. (1963). Reversibility of the Necker cube: I. An examination of the concept of “satiation of orientation. Perceptual and Motor Skills, 17, 439–458.
Orban, G. A. (2011). The extraction of 3-D shape in the visual system of human and nonhuman primates. Annual Review of Neuroscience, 34, 361–388. doi:10.1146/annurev-neuro-061010-113819
Pastukhov, A., & Braun, J. (2008). A short-term memory of multi-stable perception. Journal of Vision, 8(13), 7.1–14. doi:10.1167/8.13.7
Pastukhov, A., & Braun, J. (2011). Cumulative history quantifies the role of neural adaptation in multistable perception. Journal of Vision, 11(10), 12. doi:10.1167/11.10.12
Pastukhov, A., & Braun, J. (in press). Disparate time-courses of adaptation and facilitation in multi-stable perception. Learning & Perception. doi:10.1556/LP.4.2012.1.7
Pastukhov, A., & Braun, J. (2013). Structure-from-motion: Dissociating perception, neural persistence, and sensory memory of illusory depth and illusory rotation. Attention, Perception, & Psychophysics, 75, 322–340. doi:10.3758/s13414-012-0390-0
Pastukhov, A., Vonau, V., & Braun, J. (2012). Believable change: Bistable reversals are governed by physical plausibility. Journal of Vision, 12(1), 17. doi:10.1167/12.1.17
Pearson, J., & Brascamp, J. W. (2008). Sensory memory for ambiguous vision. Trends in Cognitive Sciences, 12, 334–341. doi:10.1016/j.tics.2008.05.006
Preston, T. J., Kourtzi, Z., & Welchman, A. E. (2009). Adaptive estimation of three-dimensional structure in the human brain. Journal of Neuroscience, 29, 1688–1698. doi:10.1523/JNEUROSCI.5021-08.2009
Ramachandran, V. S., & Anstis, S. M. (1983). Perceptual organization in moving patterns. Nature, 304, 529–531. doi:10.1038/304529a0
Richard, A. M., Luck, S. J., & Hollingworth, A. (2008). Establishing object correspondence across eye movements: Flexible use of spatiotemporal and surface feature information. Cognition, 109, 66–88. doi:10.1016/j.cognition.2008.07.004
Rokers, B., Cormack, L. K., & Huk, A. C. (2009). Disparity- and velocity-based signals for three-dimensional motion perception in human MT+. Nature Neuroscience, 12, 1050–1055. doi:10.1038/nn.2343
Sawamura, H., Orban, G. A., & Vogels, R. (2006). Selectivity of neuronal adaptation does not match response selectivity: A single-cell study of the FMRI adaptation paradigm. Neuron, 49, 307–318. doi:10.1016/j.neuron.2005.11.028
Schacter, D. L., Dobbins, I. G., & Schnyer, D. M. (2004). Specificity of priming: A cognitive neuroscience perspective. Nature Reviews Neuroscience, 5, 853–862. doi:10.1038/nrn1534
Schacter, D. L., Wig, G. S., & Stevens, W. D. (2007). Reductions in cortical activity during priming. Current Opinion in Neurobiology, 17, 171–176. doi:10.1016/j.conb.2007.02.001
Schwiedrzik, C. M., Ruff, C. C., Lazar, A., Leitner, F. C., Singer, W., & Melloni, L. (2012). Untangling perceptual memory: Hysteresis and adaptation map into separate cortical networks. Cerebral Cortex (New York, N.Y.: 1991). doi:10.1093/cercor/bhs396
Segaert, K., Weber, K., De Lange, F. P., Petersson, K. M., & Hagoort, P. (2013). The suppression of repetition enhancement: A review of fMRI studies. Neuropsychologia, 51, 59–66. doi:10.1016/j.neuropsychologia.2012.11.006
Shechter, S., Hochstein, S., & Hillman, P. (1988). Shape similarity and distance disparity as apparent motion correspondence cues. Vision Research, 28, 1013–1021.
Sperling, G., & Dosher, B. A. (1994). Depth from motion. In T. V. Papathomas, A. G. C. Chubb, & E. Kowler (Eds.), Early vision and beyond (pp. 133–142). Cambridge, MA: MIT Press.
Sterzer, P., & Rees, G. (2008). A neural basis for percept stabilization in binocular rivalry. Journal of Cognitive Neuroscience, 20, 389–399. doi:10.1162/jocn.2008.20039
Stonkute, S., Braun, J., & Pastukhov, A. (2012). The role of attention in ambiguous reversals of structure-from-motion. PLoS One, 7, e37734. doi:10.1371/journal.pone.0037734
Tanaka, Y., & Sagi, D. (1998). A perceptual memory for low-contrast visual signals. Proceedings of the National Academy of Sciences, 95, 12729–12733.
Tas, A. C., Moore, C. M., & Hollingworth, A. (2012). An object-mediated updating account of insensitivity to transsaccadic change. Journal of Vision, 12(11), 18. doi:10.1167/12.11.18
Tootell, R. B. H., Reppas, J. B., Dale, A. M., Look, R. B., Sereno, M. I., Malach, R., … Rosen, B. R. (1995). Visual motion aftereffect in human cortical area MT revealed by functional magnetic resonance imaging. Nature, 375, 139–141. doi:10.1038/375139a0
Vuilleumier, P., Henson, R. N., Driver, J., & Dolan, R. J. (2002). Multiple levels of visual object constancy revealed by event-related fMRI of repetition priming. Nature Neuroscience, 5, 491–499. doi:10.1038/nn839
Vuilleumier, P., Schwartz, S., Duhoux, S., Dolan, R. J., & Driver, J. (2005). Selective attention modulates neural substrates of repetition priming and “implicit” visual memory: Suppressions and enhancements revealed by FMRI. Journal of Cognitive Neuroscience, 17, 1245–1260. doi:10.1162/0898929055002409
Wallach, H., & O’Connell, D. N. (1953). The kinetic depth effect. Journal of Experimental Psychology, 45, 205–217.
Webster, M. A. (2011). Adaptation and visual coding. Journal of Vision, 11(5), 3.1–23. doi:10.1167/11.5.3
Wiggs, C. L., & Martin, A. (1998). Properties and mechanisms of perceptual priming. Current Opinion in Neurobiology, 8, 227–233.
Wolfe, J. M. (1984). Reversing ocular dominance and suppression in a single flash. Vision Research, 24, 471–478. doi:10.1016/0042-6989(84)90044-0
Yu, K. (2000). Can semantic knowledge influence motion correspondence? Perception, 29, 693–707. doi:10.1068/p3063
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Rights and permissions
About this article
Cite this article
Pastukhov, A., Füllekrug, J. & Braun, J. Sensory memory of structure-from-motion is shape-specific. Atten Percept Psychophys 75, 1215–1229 (2013). https://doi.org/10.3758/s13414-013-0471-8
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-013-0471-8