
Abstract
Barley plants (cv. Giza 2000) are infected by Drechslera teres which causes net blotch disease symptoms and yield losses. Plants were treated with commercial molecular products such as Trichoderma asperilium (T34), eugenol compared to non-traditional compounds (potassium silicate, nano-selenium) and fungicides (Maven, Montoro, and Decent) which selected from 12 treatments according to their important effects on infected barley plants. Bio-arc, Tilt, Rush up, Curve, and Amisto treatments were left out because of their lower efficiency of some treatments and similarity of the active ingredients of some other of them.
Disease severity (%) was significantlty decreased as a result of the selected treatments, except eugenol which showed less reduction than the control. Disease symptoms were suppressed and electrolyte leakage % was reduced significantly due to all treatments than the control treatment. As a consequence of treatments, endogenous reactive oxygen species (ROS) such as superoxide (O2−) was significantly elevated early after the inoculation, consequently, later on catalase (CAT), peroxidase (POX), and polyphenol oxidase (PPO) activities were increased significantly than the control. Elevated levels of O2− early after inoculation could play essential role in killing or suppressing the fungus and inhibiting disease symptoms as well as stimulated enzyme activities. Interestingly, the treatments were effective so that the yield characters (1000 K.W. and grain yield/plo) were increased significantly than the control treatment. These results indicated that the biological commercial product of Trichoderma asperilium 34 and non-traditional biological compounds in this research study are so effective and could be used as an alternative to fungicides.
Similar content being viewed by others

Background
Barley is one of the most important cereal crops in the world. It is used for animal feed and human food (Malcolmson et al. 2005). Barley net blotch caused by Drechslera teres f. sp. teres is one of the most important diseases of barley in the world under suitable conditions (Tekauz, 2003). Symptoms can increase quickly; causing great grain yield loss under severs conditions. Net blotch is a wide-spread disease in Middle and Northern Egypt, whereas not noticed in Upper Egypt (El-Nashar, 1983). It can cause yield loss in some susceptible cultivars such as Beka, Holker, HB-52, and HB-120 (Teshome et al. 2008). The combination of pyraclostrobin + epoxiconazole fungicides was the most effective treatment for controlling the disease and improving the yield (Stepanović et al. 2016). The pyraclostrobin, propiconazole and the propiconazole mixed with iprodione were the effective treatment for controlling the disease. Similarly, azoxystrobin, trifloxystrobin, difenoconazole, and epoxiconazole also were effective in controlling the disease (Jayasena et al. 2002). Triazole fungicides such as epoxiconazole and propiconazole exhibited great efficacy in controlling barley net blotch disease than other fungicides (Moustafa, 2017). Trichoderma viride showed maximum inhibition percentage to the mycelial growth of Bipolaris sorokiniana (Singh et al. 2018). The selenium element protected the cell membrane (Ferrarese et al. (2012). It is a co-factor of glutathione peroxidase and neutralizes tissues oxidative damage (Sors et al. 2005). The mineral (chemical) and nano (biological) selenium treatments had a significant positive effect on yield characters as well as yield quantity and quality (Shedeed et al. 2018). Both soil and foliar selenium applications had a positive effect in increasing the selenium content in some plants without negatively effect on the N, P, K, Ca, Mg, Fe, Mn, Cu, and Zn contents (Wanga et al. 2013). Bio Arc (Bacillus megaterium) was effective in controlling Alternaria solani the causal of tomato early blight disease (Moustafa et al. 2018). B. cinerea were treated with eugenol showed sever suppression. Eugenol increased the concentration of potassium ion and cellular materials in the medium (Wang et al. 2010). Hyphae treated with eugenol showed morphological alterations observed with the light and scanning electron microscopy in which appeared such as cytoplasmic coagulation, vacuolation, and hyphal shriveling. Eugenol generates hydrogen peroxide (H2O2) and increased free Ca2+ in the cytoplasm of pathogen (Wang et al. 2010). The antifungal activity of eugenol is due to membrane binding and permeability alteration, leading to disruption of the plasma membrane (Polanco et al. 2014). Foliar spray with potassium silicate, sodium molybdate, combination of potassium silicate and sodium molybdate, with or without the fungicide azoxystrobin (Azox), could reduce anthracnose symptoms caused by Colletotrichum lindemuthianum and increase yield (Menzies et al. 1992). Applications of potassium silicate in dicotyledonous plants reduced the number of colonies of Sphaerotheca fuliginea and reduced angular leaf spot severity on common bean compared to the untreated control (Rodrigues et al. 2010). It covered the cuticle of vine leaves and prevented germination and the penetration of ascospores of Uncinula necator (Bowen et al. 1992). Infection of plants led to increase reactive oxygen species (ROS) which stimulate oxidative stress in plants. Normally, upregulation of antioxidant defense systems seems to be a general response to oxidative stress (Halliwell and Gutteridge, 1999). H2O2 and O2− are the most important ROS correlated with oxidative stress (Hafez et al. 2012). Endogenous ROS were stimulated significantly early after natural infection consequently, activities of catalase (CAT), peroxidase (POX), and poly phenol oxidase (PPO) were increased significantly than the control (Hafez et al. 2014a). Elevated levels of ROS early after infection could play a pivotal role in fungal suppressing and inhibiting the severity of disease symptoms as well as immunizing plants (Hafez et al. 2012).
The present research aimed to develop non-traditional alternative and biological control compounds against D. teres compared to commercial fungicides.
Materials and methods
Source of samples
Samples of diseased barley leaves showing typical net blotch symptoms of Drechslera teres were collected during the course of this study from different governorates of Egypt.
Isolation of the causal organism
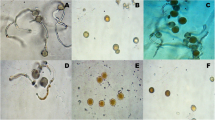
Small pieces of infected leaves were surface sterilized with 95% ethyl alcohol for 10 s and 10% sodium hypochlorite solution (5.25% NaClO) for 90 s, then rinsed in sterile distilled water and blotted between dry sterilized filter papers to remove excessed water. The sterilized pieces were aseptically transferred into Petri plates containing 2% water agar medium and incubated at 20 ± 2 °C. Pure cultures of identified D. teres were obtained from EPCRS Excellence Center, Faculty of Agriculture, Kafrelsheikh University, Egypt. It obtained by hyphal tip method (Brown, 1924). Hyphal tip, identified using a compound microscope, was transferred onto PDA medium. Barley cv. Giza 2000, highly susceptible to D. teres was used in pathological studies of different isolates.
Inoculum preparation
Mycelial fragments from 10 days old cultures of D. teres grown on PDA plates were obtained by adding approximately 20 ml of distilled water to each plate and homogenized with blender for 5 min. The density of fragment suspension was adjusted to contain 105/ml (El-Nashar, 1990).
Glasshouse experiment
Under glasshouse at Barley Disease Department, Sakha Agricultural Station, Agricultural Research Center, Egypt, barley plants (cv. Giza 2000) were inoculated, 10 days after sowing according to El-Nashar (1990). The leaves were washed well by water and swabbed with absorbent cotton to facilitate inoculation of the leaves. Barley plants were inoculated by spraying the fragment suspension using hand atomizer until the leaves were covered by an even deposit fine droplet. Control plants were sprayed by sterilized water. The inoculated plants maintained in a relative humidity of about 100% at 22 ± 2 °C for 48 h. After 48 h, the plants maintained at 22 ± 2 °C. Infection responses on the plants were scored 14 days after inoculation (El-Nashar, 1990).
Seedling stage test
Pots (7 cm) each pot containing barley 10 plants (cv. Giza 2000) were used. Ten-day-old seedlings were treated twice; the first was pre-24 h and the second 48 h post-inoculation. Three replicates were used for each treatment. Untreated plants were used as a control. After 14 days, the disease assessment was assessed visually by evaluating necrosis using a 1–10 scale (Tekauz, 1985) and data manipulated to the percentages of disease control relative to the untreated control for each treatment using the following formula:
The same experiment was repeated as additional experiment to assure the result.
Adult stage test under field conditions
Field experiments were carried out at Sakha Experimental Station during 2017/2018 and 2018/2019 growing seasons in randomized complete block design with 3 replications, using Giza 2000 susceptible barley cultivar and each plot (measured 3 × 3.5 m). Under natural infection, the disease severity was recorded pre-treatments, spraying with treatments were carried out at booting and ear emergence stages in growth stages 40 and 59 (Zadoks et al. 1974). The disease severity was recorded according to Eyal et al. (1987). The efficacy of the treatments was determined according to the equation adopted by Rewal and Jhooty (1985) as follows:
Where C = % infection in the control. T = % infection in the treatment.
Net blotch rating under glasshouse-grown barley
Net-spot blotch reaction was recorded according to Tekauz, (1985) scale of 1–10 was used for net blotch (Fig. 1), as follows:
-
1 =
Minute pinpoint or fleck-type lesions, without visible chlorosis,
-
2 =
Minute pinpoint or small (1–2 mm long) necrotic lesion, slight chlorosis on the leaf blade, and less than 20% leaf tip necrosis (R-MR),
-
3 =
Small to large (1-2 mm or larger) necrotic lesion, slight chlorosis on the leaf blade, and 21–30% leaf tip necrosis (MR),
-
4 =
Small to large necrotic lesion, slight to moderate chlorosis on the leaf blade, and about 31–40% leaf tip necrosis (MR-MS),
-
5 =
Small to large necrotic lesion, slight to moderate chlorosis on the leaf blade, and about 41–50% leaf tip necrosis (MR-MS),
-
6 =
Coalescing lesion, moderate chlorosis on the leaf blade, and 51–60% leaf necrosis (MR-MS),
-
7 =
Coalescing lesion, moderate to severe chlorosis, and 61–70% leaf necrosis (MS),
-
8 =
Coalescing lesion, sever chlorosis, and 71–80% leaf necrosis (MS-S),
-
9 =
Coalescing lesion, more necrosis than chlorosis, and 81–90% leaf necrosis (S) and
-
10 =
Coalescing lesions, more necrosis than chlorosis, and less than 10% green area visible on the leaf blade (VS). Necrosis on the leaf blade (R).

A numerical scale used for visual net blotch assessment on barley plants (Tekauz, 1985)
Net blotch rating under field-grown barley
Net blotch rating under field-grown barley Net blotch reaction under field conditions was assessed according to double-digit scale 00–99. The first digit gives the relative height rate of the disease (Saari and Prescott, 1975), where 0 = free from infection (0) 1 = resistant: a few isolated lesions on only the lowest leaves (R). 2 = resistant: scattered lesions on the second set of leaves (R). 3 = resistant: light infection of lower third of plant (R, MR). 4 = moderately resistant: moderate infection of lower leaves (MR). 5 = moderately susceptible: severe infection of lower leaves. Moderate to light infection (MR, MS). 6 = moderately susceptible: severe infection on lower third of plant. Moderate on middle leaves (MS). 7 = susceptible: lesions. Severe on lower and middle leaves (MS, S). 8 = susceptible: lesions severe on lower and middle leaves. Moderate to severe infection of upper third of plant, flag leaf infected in amounts more than a trace (S). 9 = highly susceptible: severe infection on all leaves; spike also infected to some degree. The second digit shows the disease severity as a percentage of leaf area affected in terms of 0–9, where 0 = 0%, 1 = 10%, 2 = 20%, and so on (Eyal et al. 1987).
Treatments
Chemical, biocides and non-traditional compounds (Table 1) were used. Tilt 25% EC, Decent 32.5% SC, Curve 25% EC, Amisto 25% SC, Rush up 12.5 SC, Montoro 30% EC, Maven 18.7% WG, Silicate-Potassium, and T34 12% WP were obtained from Shoura Chemical Company as well as Bio ARC 6% WP was produced by Kafr El-Zayat Chemical Company, while Eugenol active substance (99.0%) extracted from clove plants in which used in this study was brought from Faculty of Pharmacy, Cairo University, Egypt. Nano-selenium was prepared biologically in Soil Microbiology Department, Soil, Water and Environment Research Institute (SWERI), according to Eszenyi et al. (2011). The size of selenium nanoparticles and other properties were measured using TEM and X-ray in Nanotechnology Laboratory in Agriculture Research Center, Giza.
Seven treatments were selected according to their important effects on infected barley plants as follow:
Electrolyte leakage percentage
Determination of electrolyte leakage percentage (EL %) was done according to Whitlow et al. (1992) in 7 treatments at 48 hai. One gram of leaf discs (1 cm diameter) of barley leaves, where they placed in 25 ml deionized water (Milli-Q 50, USA). Flasks were shaking for 20 h at ambient temperature to facilitate electrolyte leakage from injured tissues. Initial electrical conductivity determinations were recorded for each vial using an Acromet AR20 electrical conductivity meter (Fisher Scientific, Chicago, IL, USA). Flasks were then immersed in a hot water bath (Fisher Isotemp, Indiana, PA) at 80 °C for 1 h to induce cell rupture. The vials were again placed on the Innova 2100 platform shaker for 20 h at 21 °C and final conductivity was measured for each flask. EL % for each sample was calculated as initial conductivity/final conductivity × 100.
Detection of superoxide
Superoxide (O2−) was visualized as a purple coloration of nitro blue tetrazolium (NBT) in treatments and measured at 48 hai. Barley leaves were vacuum infiltrated with 10 mM potassium phosphate buffer (pH 7.8) containing 0.1 w/v % NBT (Sigma–Aldrich, Germany) as described by Ádám et al. (1989). NBT-treated samples were incubated under daylight for 20 min and subsequently cleared in 0.15 w/v % trichloroacetic acid in ethanol: chloroform 4:1 v/v for 1 day (Hückelhoven et al. 1999). Cleared samples were washed with water and placed in 50% glycerol prior to evaluation. Discoloration of leaf discs resulted from NBT staining was quantified using a ChemiImager 4000 digital imaging system (Alpha Innotech Corp., San Leandro, USA). This test was repeated three times.
Activity of antioxidant enzymes
A half g leaf material was grinding at 0–4 °C in 3 ml of 50 mM TRIS buffer (pH 7.8), 1 mM EDTA-Na2 and 7.5% polyvinylpyrrolidone at 48, 72, and 96 hai. The homogenates were centrifuged (12,000 rpm, 20 min, 4 °C), and the total soluble enzyme activities were measured spectrophotometrically in the supernatant (Hafez et al. 2012). All measurements were carried out at 25 °C, using the model 6105, UV-Vis spectrophotometer (Jenway, UK). The enzyme activities were tested three times.
Catalase (CAT), polyphenol oxidase (PPO), and peroxidase (POX) activities were detected spectrophotometrically according to Aebi (1984), Malik and Singh (1980) and Hammerschmidt et al. (1982), respectively. CAT activity was determined by the changes in the absorbance at 495 nm were recorded every 30 s intervals for 3 min. Enzyme activity was expressed as the increase in absorbance min−1 g−1 fresh weight. PPO activity was directly determined of the crude enzyme extract according to a typical procedure proposed by the changes in absorbance at 470 nm were recorded every 30 s intervals for 3 min. POX enzyme activity was expressed as the increase in absorbance min−1 g−1 fresh weight.
Yield assessments
At maturity stage (no green tissues are present), plants of each treatment were harvested and the following parameters were determined: (1) grain yield per plot (G.y/plot), (2) weight of thousand kernels (1000 K.W.). The increase over control in yield components was estimated according to the equation adopted by Ibrahim et al. (2003), in which increase over control % = yh/yd – 1 × 100.
Statistical analysis
Two experiments were conducted in randomized complete plot design (RCPD) with three replicates for each treatment. Data represent the mean ± SD. Student’s t test was used to determine whether significant difference (P < 0.05) existed among mean values according to O’Mahony (1986). Data collected from each experiment was analyzed statistically by Duncan multiple range test (DMRT) (Gomez and Gomez 1984). The analysis was performed by the software Assist-Statistical Attendance (Silva and Azevedo, 2006).
Results and discussion
Effect of treatments on disease assessment (%) and disease symptoms under greenhouse conditions
The efficacy of 7 chemical fungicides (at the recommended dose), 2 Biocides and 3 organic materials on disease assessment were evaluated. These 12 treatments were sprayed on barley plants (10 days old) twice, the first was applied at 24 h before inoculation and the second was at 48 h post-inoculation.
Data in Table 2 indicate that the 12 chemical and biocides pyraclostrobin + dimethomorph (Maven), epoxiconazole (Rush up), propiconazole + difenoconazole (Montoro), azoxystrobin + difenoconazole (Decent), azoxystrobin (Amisto), difenoconazole (Curve), propiconazole (Tilt), Trichoderma asperellum strain (T34), (nano-selenium), and (silicate-potassium) prevented the disease development whether applied before or after inoculation. The efficacy was 100% relative to the untreated control. On the other hand, the Biocide, Bacillus megaterium (Bio Arc), and Natural material (Eugenol) were effective by 44.4 and 22.2%, respectively, when applied before inoculation, while the efficacy in diseased control was 33.3 and 11.1%, respectively, when applied after inoculation Table 2 and Fig. 2. Jordan and Best (1981) found that propiconazole, prochloraz and fenpropimorph were the most effective of nineteen fungicides in glasshouse testes against net blotch of barley. Mondrup and Welling (1984) also found that triadimefon (Bayleton 25 WP) had a very slight effect on net-spot blotch while propiconazole (Tilt 250 EC) reduced it by 80–90%. These results agree with our findings.

Effect of biological, chemical fungicides and some non-traditional compounds on disease incidence at Seedling stage. Numbers, 1: T34, 2: Bio Arc, 3: Maven, 4: Eugenol, 5: Silicate-potassium, 6: Nano-Selenium, 7: Montoro, 8: Rush up, 9: Decent, 10: Curve, 11: Tilt and 12: Amisto. A=24 h before inoculation, B= 48 h after inoculation
ROS level and action of enzymes in D. teres infected barley plants
Seven treatments were selected from the 12 treatments in this study. All treatments were capable of increase early after inoculation with 48 hai. the levels of endogenous ROS, mainly superoxide (O2−) in barley plants (Fig. 3). The purple discoloration and spots in all treatments were the indicators of O2− intensity or high levels than the control in which O2− levels were significantly reduced thus, purple discoloration was not seen (Fig. 3). Discoloration of leaf discs caused by NBT staining was visualized using the naked eye. Activities of catalase (CAT), peroxidase (POX) and polyphenol oxidase (PPO) were significantly increased in barley leaves treated with fungicides, Biological commercial product and the non-rational treatments compared to control treatments (Fig. 4). The increased activities of CAT, POX, and PPO enzymes were found after inoculation with 48, 72, and 96 h in the greenhouse.

Purple discoloration of superoxide on barley leaves early after inoculation with Drechslera teres 48 hai. Cont.: barley leaves inoculated only with D. teres. T34: commercial product og T. asperilium. Eugenol: active substance from cloves. K. Silicate: potassium silicate, nano-selenium: nano-selenium, Maven, Montoro, and Decent: fungicides

Antioxidant enzyme activities in barley leaves 48, 72, and 96 hai with D. teres. CAT: catalase, POX: peroxidase, PPO: polyphenol oxidase enzymes. Cont.: barley leaves inoculated only with D. teres. T34: commercial product of T. asperilium. Eugenol: active substance from cloves. K. silicate: potassium silicate, nano-selenium: nano-selenium, Maven, Montoro, and Decent: fungicides
As a result of early stimulation of ROS, enzyme activities were increased after inoculation; therefore, disease severity (%) and disease symptoms were suppressed (Fig. 4); this may be due to the inhibiting or the killing activity of ROS to the fungal pathogen early after infection consequently, there was no chance for the pathogen to grow or propagate (Hafez et al. 2014b). The results are supported by the non-host resistant phenomena. In the non-host/pathogen combination, when wheat infected with barley powdery mildew, superoxide, and hydrogen peroxide were accumulated early after inoculation, this early induction of ROS may be the cause of non-host resistant (Hafez et al. 2009). This result supports the idea that when ROS applied exogenously or any compound producing ROS or inducing ROS at an early time after inoculation, pathogens could be killed or significantly inhibited (Hafez and El-Baghday, 2013). Data of the present study indicated that the high levels of ROS after the inoculation encouraged, as a result, increasing the antioxidant enzyme activities therefore, protected plants against disease infection. Several studies indicated that salicylic acid (SA) and functional analogs like BTH and 2,6-dichloroisonicotinic acid (INA) can cause the ROS accumulation through the mitochondrial electron transport inhibition (Norman et al. 2004) or antioxidant enzymes (Bayoumi and Hafez, 2006). It was suggested that H2O2 induces SA accumulation and SA enhances the accumulation of H2O2 (Van Camp et al. 1998). As a result of H2O2 and O2− accumulation, microbursts may generate intensify and spread the H2O2 signal which is required for oxidative cell death and the establishment of systemic acquired resistance (SAR). It was also found an early burst of ROS and transient inhibition of antioxidant defense 1–2 days after the inoculation of Xanthi-nc leaves with TMV, followed by a massive induction of antioxidants (Fodor et al. 2001).
Electrolyte leakage
Electrolyte leakage (EL) is an indicator of the cell membrane permeability (Whitlow et al. 1992). Barley plants inoculated with D. teres and treated with fungicides as well as non-rational treatments and biological commercial products exhibited the highest significant decrease in electrolyte leakage compared to control which showed a significant increase of the membrane permeability (Fig. 5). K. Silicate, Nano-selenium, Montoro, and Maven were the best treatments which can protect cell membranes followed by Eugenol, T34, and Decent compared with control (Fig. 5). Similarly, chemical compounds and biotic or abiotic stresses could alter the resistance or susceptibility of plants to infection through their effects on membrane permeability (Hafez et al. 2014b). High-temperature stress could induce susceptibility in maize through its effect on membrane permeability as measured by increased electrolyte leakage (Garraway et al. 1989). This might result in the loss of host cells constituents which may be used by the invading pathogen as a source of nutrients. The results indicated that these treatments protected cell membranes of barley during the pathogen attack, while the cell membrane of the control treatment was affected by the pathogen infection and lost its constituents. The present results are in agreement with those obtained by (Hafez et al. 2014b).

Electrolyte leakage in barley leaves after inoculation with Drechslera teres 48 hai. Cont.: barley leaves inoculated only with D. teres. T34: commercial product of T. asperilium. Eugenol: active substance from cloves. K. Silicate: potassium silicate, nano-selenium: nano-selenium, Maven, Montoro, and Decent: fungicides
Effect of treatments on yield components
The effect of 12 treatments (traditional and non-traditional compounds) on net blotch severity and yield components (1000 K.W.) and grain weight/plot) were evaluated under natural infection at Sakha Experimental Station during two successive seasons (2017/2018 and 2018/2019).
Data in Table 3 also showed that the difference in disease severity and efficacy between one or two sprays of each fungicide was very high. The effectiveness of 12 tested traditional and non-traditional compounds against net blotch during the two seasons, could be arranged in descending manner as follows: Decent, Maven Montoro, Amisto, Curve, Rush up, Tilt, T34, Silicate-potassium, Nano-Selenium, Bio Arc, and Eugenol while efficacy of treatments indicated that the best treatments were Maven, Decent and Montoro in chemical compounds. In this respect, Jordan and Best (1981) obtained partial control of a severe net blotch infection and a 23% yield increase on winter barley with a single spray of propiconazole (Tilt) at flag leaf emergence. Locke et al. (1981) meanwhile reported that the best fungicide control of net blotch was achieved with the systemic material propiconazole and prochloraz. They also determined the best time of application of both fungicides to be during the period between flag leaf emergence and full ear emergence (growth stages 37–58).
They expected returns for yield increased by 15-20% and exceeded the costs of the spray application. Six fungicides in vivo were tested to evaluate their efficiency in controlling net blotch (EL-Nashar, 1990). It was found that the propiconazole performance as two sprays to either pot or field growth barley was effective for controlling net blotch disease in barley (Chin et al. 2001). A combination of pyraclostrobin + epoxiconazole fungicide was the most effective treatment for controlling the net blotch disease and improving yield (Stepanović et al. 2016). In this respect, Jayasena et al. (2002) evaluated 10 fungicides for control of spot-type net blotch of barley caused by D. teres f. sp. maculata at 3 locations during 1999 and 2000. It was found that under moderate disease severity, yield losses ranged from 17 to 19% depending on location and under high-disease severity, yield losses reached 32%. Pyraclostrobin, propiconazole, and combination of propiconazole with iprodione were effective in controlling the disease, improving yield and grain quality. These results are in agreement with our findings. On the other hand, in bio-fungicides and non-traditional compounds, T34 and silicate-potassium were the best treatments when applied one and two sprays. Polanco et al. (2014) determined whether foliar sprays of potassium silicate, sodium molybdate, or a combination of both, with or without the fungicide azoxystrobin could reduce anthracnose symptoms and increase yield. Foliar application increased silica concentration in bean leaves (Rodrigues et al. 2010). The highest silicate-potassium rate reduced angular leaf spot severity on common bean by 42 and 30% and defoliation by 17 and 33% at pH 5.5 and 10.5, respectively, than the untreated control. Yield also increased by 30 and 43% as rates increased from 0 to 60 g/L with pH 5.5 and 10.5, respectively (Rodrigues et al. 2010). It covered the cuticle of vine leaves and prevented both germination and the penetration of ascospores of Uncinula necator; in contrast, the fungus grew without being impeded on the leaf areas that were not covered. Trichoderma viride exhibited maximum inhibition percentage (73.07 and 72.30%) on mycelial growth of Bipolaris sorokiniana (Singh et al. 2018). Trichoderma strains produce enzymes such as chitinases and β-1,3-glucanases that cause hydrolyze the structural compounds of fungal cell walls, chitin, and β-glucan (Qualhato et al. 2013). The effect of treatments on 1000 K.W. showed a significant difference between the treatments and the control (Table 4). The 1000 K.W. were 55.15, 54.91, and 54.77 g when Decent, Maven, and Montoro were applied as one spray followed by Rush up 53.17 g, Amisto 52.91 g, Curve 51.2 g, Tilt 50.70 g, T34 50.67 g, silicate-potassium 50.61 g, nano-selenium 48.7 g, Bio arc 48.47 g, and Eugenol 46.25 g compared to the control 45.21 g. While in the two sprays the 1000 K.W. to the treatments Decent, Maven, Montoro, Rush up, Amisto, Curve, Tilt, T34, silicate-potassium, nano-selenium, Bio Arc, and eugenol reached to 57.15 g, 56.30 g, 56.10 g, 54.67, 53.01,52.83, 52.63, 52.31,51.35,50.17, and 48.2 g, respectively, when were applied compared to the control which, the 1000 K.W. reached to 45.21 g. The increase over control of 1000 K.W. were observed in one and two sprays, which the increase over control reached 26.41% in Decent, 24.5 and 24.08% in Maven and Montoro, followed by Rush up, Amisto, Curve, Tilt, T34, silicate-potassium, nano-selenium, and Bio Arc. The lowest increase was observed at 6.61% in eugenol. Similar results observed in the 2018/2019 season, which the treatments were effective on increase the 1000 K.W. and increase over control compared to the control, when the treatments were applied as two sprays (Table 4). The mineral-Se and nano-Se selenium spraying treatments had a significant effect on all yield characters such as plant height, number of leaves, number of branches, fresh weight of leaves and stems, dry weight of leaves and stems, chlorophyll (SPAD), and yield quantity and quality, which represented in pod weight, plant yield, and total yield of pea plant (Shedeed et al. 2018). Foliar spray with selenium “nano or mineral-Se” increased vegetative growth, yield, and quality. Moreover, the best use of selenium as a foliar spray was the nano type because it is more safe and friendly in environmental applications compared to the chemical type. Wanga et al. (2013) demonstrated that both soil and foliar Se applications had a positive effect in increasing the Se content in some plants without a negative effect on the micro and macro elements contents. Furthermore, foliar application is effective than soil application. Triazoles fungicides showed greater efficacy in the control net blotch of barley than other fungicides (Moustafa, 2017). B. megaterium was the most effective one (67.8%) followed by Bio Zeid (43%) in control tomato early blight disease (Moustafa et al. 2018). Wang et al. (2010) showed that Botrytis cinerea fungi exposed with eugenol showed strong propidium iodide fluorescence in the cytosol. Eugenol improved the rate of potassium ion and cellular materials in the medium. Fungal exposed to eugenol revealed morphological alterations in hyphae, such as cytoplasmic coagulation, vacuolation, and hyphal shriveling. Eugenol induced H2O2 and increased free Ca2+ in the cytoplasm.
Decent, Maven, and Montoro fungicides were the best in increasing the grain yield/plot reached to 5.27, 5.23, and 5.23 kg, respectively, while the control reached to 4.19 kg (Table 5). The increase over control reached to 25.77, 24.82, and 24.82% when the treatments, Decent, Maven, and Montoro were applied as two sprays, while the Eugenol occupied the last rank, which the grain yield/plot was 4.38 kg and the increase over control 4.53%.
In the 2018/2019 growing season, the same results were observed, which the best treatments Decent, Maven and Montoro were effective on the grain yield/plot 5.24, 5.22, and 5.21 kg than the control 4.12 kg. The increase over control was observed in Decent (27.18%), followed by Maven and Montoro, which the increase over control was 26.69 and 26.45%, respectively.
Conclusion
The results indicated that the non-rational compounds, biological commercial products, and tested fungicides treatments suppressed the fungal growth in correlation with increased ROS level mainly superoxide early, decreasing the electrolyte leakage and perhaps direct toxic effect of some treatments, as a result, the antioxidants were stimulated accordingly. Therefore, it can be suggested as an alternative to fungicides.
Availability of data and materials
All data and materials are available.
Abbreviations
- CAT:
-
Catalase
- Cv.:
-
Cultivar
- dai:
-
Days after infection
- EL:
-
Electrolyte leakage
- hai:
-
Hours after infection
- K.W:
-
Kernel weight
- MR:
-
Moderately resistant
- MS:
-
Moderately susceptible
- O2 . :
-
Superoxide
- PDA:
-
Potato dextrose agar
- Pox:
-
Peroxidase
- PPO:
-
Polyphenol oxidase
- R:
-
Resistant
- ROS:
-
Reactive oxygen species
- S:
-
Susceptible
References
Ádám A, Farkas T, Somlyai G, Hevesi M, Király Z (1989) Consequence of O2·-generation during a bacterially induced hypersensitive reaction in tobacco: deterioration of membrane lipids. Physiol Mol Plant Pathol 34:13–26
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
Bayoumi YA, Hafez YM (2006) Effect of organic fertilizers combined with benzo (1,2,3) thiadiazole-7-carbothioic acid S-methyl ester (BTH) on the cucumber powdery mildew and the yield production. Acta Biolo Szeged 50:131–136
Bowen PJ, Menzies J, Ehret D (1992) Soluble silicon sprays inhibit powdery mildew development on grape leaves. J Am Soc Hortic Sci 117:906–912
Brown W (1924) Two mycological methods. 11. A method of isolating single strains of fungi by cutting a hyphal tip. Ann-Bot 38:404
Chin KM, Chavaillaz D, Kaesbohrer M, Staub T, Felsenstein FG (2001) Characterizing resistance risk of Erysiphe graminis f.sp tritici to strobilurins. Crop Prot 20:87–96
El-Nashar FK (1983) Net blotch disease of barley caused by Drechslera teres (Sacc.) Shoem. M.Sc. Thesis, Faculty of Agricultur Cairo University, p 103
El-Nashar FK (1990) Further studies on net blotch of barley. Ph. D, Faculty of Agricultur Cairo University, p 112
Eszenyi P, Sztrik A, Babka B, Prokisch J (2011) Elemental, nano-sized (100-500 nm) selenium production by probiotic lactic acid bacteria. Internat J of Biosci, Biochemi and Bioinform 1(2):148–152
Eyal Z, Sharen AL, Prescott JM, Van Ginkel M (1987) The septoria diseases of wheat: concepts and methods of disease management. DF CIMMYT, Mexico
Ferrarese A, Saurestani MN, Quattrini E, Schiavi M, Ferrante A (2012) Biofortification of spinach plants applying selenium in the nutrient solution of floating system. J Agric & Environ. Sci 76:127–136
Fodor J, Hideg E, Kecskes A, Kiraly Z (2001) In vivo detection of tobacco mosaic virus-induced local and systemic oxidative burst by electron paramagnetic resonance spectroscopy. Plant Cell Physiol 42:775–779
Garraway MO, Akhtar M, Wokoma ECW (1989) Effect of high temperature stress on peroxidase activity and electrolyte leakage in relation to sporulation of Bipolaris maydis race T. Phytopathol 79:800–805
Gomez KA, Gomez AA (1984) Statistical procedures foragricultural research, 2nd edn. International Rice Research Institute, College, Laguna, p 680
Hafez YM, Bacso R, Király Z, Kunstler A, Király L (2012) Up-regulation of antioxidants in tobacco by low concentrations of H2O2 suppresses necrotic disease symptoms. Phytopathol 102:848–856
Hafez YM, El-Baghdady NA (2013) Role of reactive oxygen species in suppression of barley powdery mildew fungus, Blumeria graminis f.sp. hordei with benzothiadiazole and riboflavin. Egyp J Biol Pest Control 23(1):125–132
Hafez YM, Király Z, Manninger K (2009) Hydrogen peroxide has a key role in resistance to leaf rust (Puccinia triticina) in several Egyptian and other wheat cultivars. Cer Res Commun 37:161–164
Hafez YM, Mourad RY, Mansour M, Abdelaal Kh AA (2014b) Impact of non- traditional compounds and fungicides on physiological and biochemical characters of barely Infected with Blumeria graminis f. sp hordei under field conditions. Egypt J Biol Pest Control 24(1):445–453
Hafez YM, Soliman NK, Saber MM, Imbabi IA, Abd-Elaziz AS (2014a) Induced resistance against Puccinia triticina the causal agent of wheat leaf rust by chemical inducers. Egypt J Biol Pest Control 24(1):173–181
Halliwell B, Gutteridge JMC (1999) Free Radicals in Biology and Medecine. Oxford University Press Inc, New York, pp 246–350
Hammerschmidt R, Nuckles EM, Kuć J (1982) Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol Plant Pathol 20(1):73–82
Hückelhoven R, Fodor J, Preis C, Kogel KH (1999) Hypersensitive cell death and papilla formation in barley attacked by the powdery mildew fungus are associated with hydrogen peroxide but not with salicylic acid accumulation. Plant Physiol 119:1251–1260
Ibrahim AS, Khafagi YS, Ghanim AM, EL-Abbasi IH (2003) Integrated management of powdery mildew on cantaloupe. Egypt J of Applied Sci 18:521–531
Jayasena KW, Loughman R, Majewski J (2002) Evaluation of fungicides in control of spot-type net blotch on barley. Crop Prot 21:63–69
Jordan VWL, Best GR (1981) Evaluation of fungicide treatments for control of barley net blotch caused by pyrenophora teres proceeding 1981. British Crop Protection Conference- Pest and Disease 1:249–258
Locke T, Evans EJ, Dobson SC, Morgan W (1981) A review of factors influencing net blotch of barley and current information on fungicidal control. Proceeding 1981 British Crop Protection Conference- Pest and Disease 1:283–290
Malcolmson L, Nowkirk R, Carson G (2005) Expanding opportunities for barley food and feed through product innovation. Feed and quality; 18th National American Barley Research Workshop 4th Canadian Barley symposium, pp 2–4
Malik CP and Singh MB (1980) In: Plant Emymology and Histoenzymology. Kalyani Publishers. Indian and printed in Navin. Shanndara. Delhi; pp. 54–56.
Menzies JP, Bowen DL, Glass ADM (1992) Foliar applications of potassium silicate reduce severity of powdery mildew on cucumber, muskmelon, and zucchini squash. J Am Soc Hortic Sci 117:902–905
Mondrup A, Welling B (1984) Net-spot blotch (Drchslera teres) assessments in some barley varieties. Tidsskr Planteavl 88:203–211
Moustafa MSH, ALkolaly AMA, El-Dakar HAM (2018) Impact of Bioagents and organic acids on Alternaria solani, the causal organism of tomato early blight disease and their side effect assessment. Glob J of Advan Res. 5(7):234–242
Moustafa NA (2017) Use of Linear Correlation between in Vitro And in Vivo Tests for Evaluating Efficiency of Some Fungicides in Controlling Net Blotch of Barley (Hordeum Vulgare). J Agric Chem and Biotechn Mansoura Univ 8(2):35–45
Norman C, Howell KA, Millar AH, Whelan JM, Day DA (2004) Salicylic acid is an uncoupler and inhibitor of mitochondrial electron transport. Plant Physiol 134:492–501
O’Mahony M (1986) Sensory Evaluation of Food: Statistical Methods and Procedures. CRC Press, p 487
Polanco LR, Rodrigues FA, Moreira EN, Duarte HSS, Cacique IS, Valente LA, Vieira RF, Paula Júnior TJ, Vale FXR (2014) Management of anthracnose in common bean by foliar sprays of potassium silicate, sodium molybdate, and fungicide. Plant Dis 98:84–89
Qualhato FT, Lopes FAC, Steindorff AS, Brando RS, Jesuino RSA, Ulhoa CJ (2013) Mycoparasitism studies of Trichoderma species against three phytopathogenic fungi: evaluation of antagonism and hydrolytic enzyme production. Biotechnol Lett 35:1461–1468
Rewal HS, Jhooty JS (1985) Differential response of wheat varieties to systemic fungicides applied to Ustilago tritici (Pres.). Rust Indian J Agric Sci 55(8):548–549
Rodrigues FA, Duarte HSS, Rezende DC, Wordell Filho JA, Korndörfer GH, Zambolim L (2010) Foliar spray of potassium silicate on the control of angular leaf spot on beans. J Plant Nutr 33:2082–2093
Saari EE, Prescott JM (1975) A scale for appraising the foliar intensity of wheat disease. Plant Dis Reporter 59:377–380
Shedeed SI, Fawzy ZF, El-Bassiony AM (2018) Nano and mineral selenium foliar application effect on pea plants (Pisum sativum L.). Biosci res 15(2):645–654
Silva FS, Azevedo CAV (2006) A new version of the assistat-statistical assistance software. In: World congress on computers in agriculture, 4, Orlando-FL-USA: Anais. Orlando: American Society of Agricultural and Biological Engineers, p 393–396
Singh D, Pande SK, Yadav KJK, Kumar S (2018) Bioefficacy of Trichoderma spp. against Bipolaris sorokiniana Causing Spot Blotch Disease of Wheat and Barley. In J Curr Microbiol App Sci 7(3):2322–2327
Sors TG, Ellis DR, Salt DE (2005) Selenium uptake, translocation, assimilation and metabolic fate in plants. Photosynthesis Res 86:373–389
Stepanović M, Rekanović E, Milijašević-Marčić S, Potočnik I, Todorović B, Stepanović J (2016) Field efficacy of different fungicide mixtures in control of net blotch on barley. Pestic Phytomed (Belgrade) 31(1-2):51–57
Tekauz A (1985) A numerical scale to classify reactions of barley to Pyrenophora teres. Canad J Plant Pathol 7:181–183
Tekauz A (2003) Spot blotch caused by Bipolaris sorskiniana: an emerging problem in barley in western Canada. In: proceedings of 8 th International Congress of plant Pathology Christchurch, New Zealand, p 103
Teshome G, Chemeda F, Geremew B (2008) Effects of Net Blotch (Pyrenophora teres) on Malt Barley Yield and Grain Quality at Holeta Central Ethiopia. East Africa J of Sci. 2(2):150–158
Van Camp W, Van Montagu M, Inzé D (1998) H2O2 and NO: redox signals in disease resistance. Trends Plant Sci 3:330–334
Wang C, Zhang J, Chen H, Fan Y, and Shi Z (2010) Antifungal activity of eugenol against Botrytis cinereal. Tropical Plant Pathology, 35 (3):137–143.
Wanga J, Wanga Z, Maoa H, Zhaoa H, Huanga D (2013) Increasing Se concentration in maize grain with soil- or foliar-applied selenite on the Loess Plateau in China. Field Crops Research 150:83–90
Whitlow TH, Bassuk NL, Ranney TG, Reichert DL (1992) An improved method for using electrolyte leakage to assess membrane competence in plant tissues. Plant Physiol 98:198–205
Zadoks JC, Chang TT, Konzak CF (1974) A decimal code for the growth stage of Cereals. Weed Research 14:415–421
Acknowledgements
The laboratory research was conducted at Plant Pathology and Biotechnology Laboratory, (ISO/17025 Accredited) and EPCRS Excellence Centre (Certified according to ISO/9001, ISO/14001 and OHSAS/18001), Department of Agricultural Botany, Faculty of Agriculture, Kafr-Elsheikh University, Kafr-Elsheikh, Egypt.
Funding
Funding is by EPCRS Excellence Center and authors.
Author information
Authors and Affiliations
Contributions
All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethics committee approved the research article and all authors agree (consent) to participate in this research article.
Consent for publication
All authors participated in the work and consent for publication.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Hafez, Y., Abdelfatah, A., El-Nashar, F. et al. Management of barley net blotch using Trichoderma asperellum (T34), eugenol, non-traditional compounds and fungicides. Egypt J Biol Pest Control 29, 88 (2019). https://doi.org/10.1186/s41938-019-0179-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-019-0179-8