
Abstract
Background
Studies have demonstrated that transforming growth factor beta-1 (TGF-β1) exhibits oncogenic activity in different types of cancer, including ovarian cancer (OC). However, its regulatory mechanism in OC and whether TGF-β1 is involved in chemosensitivity regulation remains unclear. Thus, the aim of this study was to investigate the role of TGF-β1 in OC.
Methods
The OC cell line SKOV3 was employed, and TGF-β1 overexpression or knockdown vectors were constructed. The cell proliferation of SKOV3 was evaluated with the cell counting kit (CCK8) kit after treatment with different concentrations of cis-platinum. Western blot and protein immunoprecipitation were employed to detect changes in BRCA1 and Smad3 expression and their interactions. Tumor growth in nude mice was evaluated.
Results
The results showed that TGF-β1 knockdown increased chemosensitivity by promoting BRCA1 expression and Smad3 phosphorylation. In vivo studies showed that TGF-β1 knockdown significantly inhibited the growth of tumors, also by upregulating BRCA1 expression and Smad3 phosphorylation.
Conclusion
Taken together, our results suggest that TGF-β1 knockdown inhibits tumor growth and increases chemosensitivity by promotion of BRCA1/Smad3 signaling.
Similar content being viewed by others


Introduction
Ovarian cancer (OC) is the most common and deadly disease among women. The histological divisions include sex cord-stromal, epithelial and germ cell tumors [1,2,3]. Epithelial OC (EOC) is derived from the fallopian tubes or ovary-originating epithelial cells, with more than 90% of total OC occurring postmenopausally [4, 5]. More than 70% of EOCs are diagnosed in the advanced stage due to the difficulty in detection and the absence of effective diagnostic markers, making it the most deadly gynecological malignant tumor [6, 7]. After surgical intervention, cisplatin is the major chemotherapy drug for OC, but patients tend to develop cisplatin resistance in the clinical setting [8, 9]. Therefore, there is a critical need to identify possible targets for therapeutic intervention.
Transforming growth factor beta (TGF-β) plays an important role in promoting cell proliferation and cell cycle regulation in early oncogenesis and normal tissue development [10, 11]. Previous studies have found that TGF-β1 can activate both SMAD2 and SMAD3 [12]. SMAD2 and 3 dimerize, forming the SMAD2/3 complex. Then, the SMAD2/3 complex interacts with SMAD4 and forms a heterohexameric complex. After translocation into the nucleus, the heterohexameric complex transcriptionally regulates cellular process-related target genes such as the induction of chemotherapy resistance [13,14,15]. However, it has been shown that patients who have BRCA1 genetic defects have an increased risk of cancer. Increasing evidence has found that BRCA1 is involved in many cellular processes, including apoptosis, genomic stability, DNA-damage repair and the cell-cycle checkpoint [16]. A previous study found that BRCA1 expression promotes the interaction of Smad3 and Smad4. Additionally, BRCA1 regulates Smad3-mediated TGF-β signaling by interacting with Smad3 [17]. However, it remains unclear whether the TGF-β signal regulates BRCA1 expression.
Thus, in this study we investigated the role of TGF-β1 in OC. We also examined whether dysfunction of TGF-β1 leads to decreased growth and increased chemosensitivity by induction of the BRCA1/SMAD3 pathway.
Materials and methods
Animals and ethics statement
Five to six week old male BALB/C nude mice (18–20 g) were purchased from the SLAC Laboratory Animal Co (Shanghai, China) and housed under 24–26 °C with a 12 h light/dark cycle with free access to food and water provisions. All experiments and animal procedures were approved by the Ethics Committee of Tongji Hospital, Tongji University School of Medicine.
Cells lines and cell culture
SKOV3 cells were purchased from the American Type Culture Collection (ATCC; Manassas, VA, USA). RPMI 1640 (Invitrogen, Carlsbad, CA, USA) was used to culture SKOV3 cells after supplementation with 10% fetal bovine serum (Invitrogen). The cells were cultured at 37 °C in 5% CO2. For the chemotherapy resistance analysis, SKOV3 cells were pretreated with 0, 0.625, 1.25, 2.5, 5 or 10 μg/ml cis-platinum for 48 h before cell proliferation analysis.
Transfection of cells with TGF-β1 overexpression or knockdown vectors
TGF-β1 overexpression vectors or vectors containing siRNA against TGF-β1 were constructed by Wuhan Biobuffer Biotech Service (Wuhan, China). TGF-β1 overexpression vectors were constructed by inserting the TGF-β1 sequence into pCDNA3.1 vectors. Vectors with siRNA against TGF-β1 were constructed by inserting a TGF-β1 silencing sequence into pLenR-GPH vectors. TGF-β1 overexpression or downregulation vectors were then transfected into SKOV3 cells at a final concentration of 50 nM using Lipofectamine 2000 (Invitrogen). After transfection for 48 h, SKOV3 cells were collected for the following experiments.
Cell viability assay
Cell viability was analyzed using the cell counting kit-8 (CCK8; Invitrogen). Briefly, 1 × 104 SKOV3 cells were seeded into a 96-well plate and incubated overnight at the previously described conditions. Then, the medium was discarded and the cells were washed with PBS three times before adding DMEM (90 µl) and CCK8 (10 µl). After incubation for 1.5 h at 37 °C, the optical density (OD) was measured with a microplate reader at 450 nm.
Western blot
Protein from cells or tissues was extracted standardization with a BCA kit (Pierce, Rockford, IL, USA). Then, protein samples (40 μg) were used for sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride (PVDF) membranes. The PVDF membranes were then incubated with the following primary antibodies after blocking in 5% nonfat milk: anti-BRCA1 (1:600), anti-p-Smad3 (1:800), anti-Smad3 (1:800), anti-TGF-β1 (1:1000) and anti-β-actin (1:1000) (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The membranes were then incubated with secondary antibodies (Santa Cruz Biotechnology) for 1 h at room temperature before the chemiluminescence was measured. The Quantity One program (Bio-Rad, Hercules, CA, USA) was used to measure the intensity of the protein bands.
Quantitative real-time PCR (qRT-PCR)
RNA from the cells was isolated using TRIzol reagent (Molecular Research Center, Cincinnati, OH, USA). Then, cDNA was synthesized using the RNA reverse transcription kit (Invitrogen). The Applied Biosystems 7300 qPCR system (Applied Biosystems; Thermo Fisher Scientific) was used for qRT-PCR analysis. The qRT-PCR reaction conditions were as follows: 95 °C for 10 s, 94 °C for 15 s and annealing at 55 °C for 30 s with 40 cycles. The relative expression level of TGF-β1 was calculated using the 2−ΔΔCq method. The PCR primers used were as follows: β-actin: forward 5ʹ- AGCGAGCATCCCCCAAAGTT-3ʹ, reverse 5ʹ-GGGCACGAAGGCTCATCATT-3ʹ; TGF-β1: forward 5ʹ-CAGCAACAATTCCTGGCGATACC-3ʹ, reverse 5ʹ-GCGCTAAGGCGAAAGCCCTCAAT-3ʹ.
Immunoprecipitation
About 1–4 mg of cell lysates from SKOV3 cells were pre-cleared with protein G beads for 30 min at 4 °C and subsequently incubated with protein G beads prebound with antibody for 2–16 h at 4 °C. The beads were washed with 1% NP40 three times before mixing with sample buffer (6×) and were subjected to SDS-PAGE for western blot detection.
Tumor xenograft mouse model
Viable SKOV3 cells [5 × 106; containing TGF-β1 overexpression SKOV3 cells, wildtype (control) SKOV3 cells and siRNA-TGF-β1 SKOV3 cells] were subcutaneously injected into nude mice. Tumor sizes were measured every 3 days after subcutaneous injection for 10 days using a vernier caliper (volume = 1/2 × length × width2). The mice were sacrificed at 40 days after implantation, the tumors were dissected and TGF-β1, BRCA1 and Smad3 expression in tumor tissues was measured.
Immunohistochemical analysis
Tumor tissue samples were fixed in 10% formalin, embedded and cut into 5 μm slices. Then, the tumor sections were stained with an anti-TGF-β1 (Santa Cruz Biotechnology) antibody after dewaxing and rehydration. An Axiophot light microscope (Zeiss, Oberkochen, Germany) was used for TGF-β1 expression analysis.
Statistical analysis
Data are expressed as the mean ± SD. The student’s t-test was used to evaluate the differences between groups using SPSS 16.0 (SPSS Inc., USA). A value of P < 0.05 was considered a statistically significant difference.
Results
Downregulation of TGF-β1 increases chemotherapy sensitivity in ovarian carcinoma cells
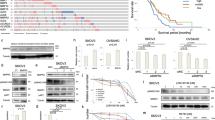
To identify if TGF-β1 plays a role in OC chemotherapy resistance and proliferation, TGF-β1 overexpression or knockdown vectors were constructed. The results showed that the TGF-β1 level was significantly increased after transfection with the TGF-β1 overexpression vector but was downregulated after transfection with the siRNA vector against TGF-β1 compared with the wildtype control at both the mRNA and protein levels in SKOV3 cells (Fig. 1a, b). The SKOV3 cell line was derived from the ascitic fluid of a Caucasian female with an ovarian tumor. After pretreatment with cisplatin for 48 h. Then, CCK8 assays were used to detect the cell viability of SKOV3 cells. The results showed that knockdown TGF-β1 decreased the chemotherapy resistance of SKOV3 cells, and only 0.625 μg/ml cis-platinum led to a 50% loss in cell viability. Conversely, TGF-β1 overexpression decreased the chemotherapy resistance of SKOV3 cells, leading to a tolerance of 2.5 μg/ml cis-platinum (Fig. 1c–e).

Downregulation TGF-β1 increases chemotherapy sensitivity in ovarian carcinoma cells. Western blot (a) and qRT-PCR (b) analysis show the expression of TGF-β1 after transfection with TGF-β1 overexpression or siRNA knockdown vectors. Data are expressed as the mean ± SD. c–e SKOV3 cells were treated with different concentrations of cisplatin for 48 h, and then the cells were collected for CCK8 analysis. Data are expressed as the mean ± SD. ***P < 0.001 versus healthy controls
TGF-β1 knockdown promotes BRCA1/Smad3 signal activation
Increasing evidence has shown that BRCA1 is involved in the regulation of tumor chemotherapy resistance and proliferation [18, 19]. In our study, western blot analysis showed that dysregulation TGF-β1 decreased BRCA1 expression compared with wildtype SKOV3 cells. However, downregulation of TGF-β1 promoted BRCA1 expression (Fig. 2a, b). Further analysis showed that TGF-β1 knockdown promoted Smad3 phosphorylation. On the other hand, TGF-β1 overexpression decreased Smad3 phosphorylation (Fig. 2c, d). Our co-immunoprecipitation data suggested that BRCA1 can interact with Smad3 (Fig. 2e, f). This suggests that TGF-β1 knockdown increases the chemotherapy sensitivity of SKOV3 cells via promotion of BRCA1/Smad3 signaling.

The effect of TGF-β1 on BRCA1/Smad3 signaling. a, b Western blot showing BRCA1 expression. Data are expressed as the mean ± SD. c, d Western blot showing the Smad3 expression and phosphorylated Smad3. Data are expressed as the mean ± SD. e, f Immunoprecipitation analysis of the relationship between Smad3 and BRCA1. ***P < 0.001
TGF-β1 downregulation suppressed tumor growth
The size of SKOV3 tumors in mice was measured and the results showed that knockdown of TGF-β1 suppressed tumor growth when compared with the control group (Fig. 3a, b). TGF-β1 expression in tumors tissues was then analyzed using immunohistochemistry. The results showed that the TGF-β1 level was increased in the TGF-β1 overexpression group but was decreased in the TGF-β1 knockdown group (Fig. 3c, d). Western blot showed that BRCA1 expression and Smad3 phosphorylation were increased in the TGF-β1 knockdown group. However, upregulation TGF-β1 decreased the level of BRCA1 and Smad3 phosphorylation in tumor tissues (Fig. 3e–h).

TGF-β1 downregulation suppressed tumor growth of ovarian cancer in a xenograft model. a Photographs of tumor tissues from different groups at day 40 (n = 5). b Growth curves for tumor volumes in xenografts of nude mice were determined based on the tumor volume measured every 3 days from the tenth day for 40 days (n = 5). ***P < 0.001 versus the control group. ###P < 0.001 versus the TGF-β1 overexpression group. c, d Immunohistochemical analysis showing the expression of TGF-β1 in tumor tissues. Data are expressed as the mean ± SD. e–h Western blot showing the expression of TGF-β1, BRCA1, Smad3 and phosphorylated Smad3. Data are expressed as the mean ± SD. ***P < 0.001
Discussion
Our study reveals a new role and function of TGF-β1 in OC. We show for the first time that knockdown of TGF-β1 suppresses tumor growth and promotes the chemosensitivity of OC cells to cis-platinum, in that we observed a BRCA1 expression increase and the activation of Smad3.
OC is a gynecological disease with a high fatality rate, especially epithelial origin OC (more than 90% of OC cases) [20]. TGF-β1 has been shown to be upregulated in OC and an increasing number of studies has found that in ovarian clear cell carcinomas, the expression of TGF-β1 promotes cancer stem cell properties and the epithelial–mesenchymal transition (EMT) [21]. SMAD transcription factors, which can mediate genetic transcription, are downstream of TGF-β1 [22]. A previous study showed that SMAD3 can be activated by TGF-β1 [23]. Here, we found that TGF-β1 downregulation has effects in addition to the activation of SMAD3. Furthermore, another study found that SMAD3 blocked the phosphorylation of AKT, which promoted the chemosensitivity of hepatocellular carcinoma to cisplatin [24].
In this study, we found that BRCA1 plays an important role in TGF-β1 mediated chemotherapy resistance and tumor growth. We observed that BRCA1 can interact with Smad3 protein, which results in an increase in Smad3-mediated transcriptional activity. Our study also found that BRCA1 mutants suppress the activation of the TGF-β1-responsive reporter or promote Smad3 expression, suggesting that BRCA1 inactivation impairs TGF-β1 signaling [17, 25]. These findings suggest that BRCA1 plays an important role in the link between TGF-β1 and SMAD signaling.
Conclusion
In conclusion, in this study we show that downregulation of TGF-β1 expression inhibits cell proliferation and increases chemosensitivity in OC by activating BRCA1/Smad3 signaling. TGF-β1-mediated BRCA1/Smad3 signaling may therefore be a novel therapeutic and diagnostic option for OC clinical treatment.
Abbreviations
- TGF-β1:
-
transforming growth factor beta-1
- OC:
-
ovarian cancer
- EOC:
-
epithelial OC
- CCK8:
-
cell counting kit-8
- SDS-PAGE:
-
sodium dodecyl sulfate polyacrylamide gel electrophoresis
- PVDF:
-
polyvinylidene difluoride
- qRT-PCR:
-
quantitative real-time PCR
- EMT:
-
epithelial–mesenchymal transition
References
Uma Devi K, Purushotham N, Jayashree N. Management of ovarian cancer in younger women. Rev Recent Clin Trials. 2015;10:263–9.
Chan JK, Cheung MK, Husain A, Teng NN, West D, Whittemore AS, Berek JS, Osann K. Patterns and progress in ovarian cancer over 14 years. Obstet Gynecol. 2006;108:521–8.
Williams AR. Pathological assessment of ovarian cancer. Methods Mol Med. 2001;39:49–60.
Li J, Fadare O, Xiang L, Kong B, Zheng W. Ovarian serous carcinoma: recent concepts on its origin and carcinogenesis. J Hematol Oncol. 2012;5:8.
Jelovac D, Armstrong DK. Recent progress in the diagnosis and treatment of ovarian cancer. CA Cancer J Clin. 2011;61:183–203.
Jayson GC, Kohn EC, Kitchener HC, Ledermann JA. Ovarian cancer. Lancet. 2014;384:1376–88.
Rauh-Hain JA, Krivak TC, Del Carmen MG, Olawaiye AB. Ovarian cancer screening and early detection in the general population. Rev Obstet Gynecol. 2011;4:15–21.
Zhao D, Wang X, Zhang W. GDF15 predict platinum response during first-line chemotherapy and can act as a complementary diagnostic serum biomarker with CA125 in epithelial ovarian cancer. BMC Cancer. 2018;18:328.
Shilnikova K, Piao MJ, Kang KA, Ryu YS, Park JE, Hyun YJ, Zhen AX, Jeong YJ, Jung U, Kim IG, Hyun JW. Shikonin induces mitochondria-mediated apoptosis and attenuates epithelial–mesenchymal transition in cisplatin-resistant human ovarian cancer cells. Oncol Lett. 2018;15:5417–24.
Kanaji N, Yokohira M, Watanabe N, Kadowaki N, Bandoh S. Transforming growth factor-beta produced by non-small cell lung cancer cells contributes to lung fibroblast contractile phenotype. Anticancer Res. 2018;38:2007–14.
Papageorgis P. Complex interplay between aging and cancer: role of TGF-beta signaling. Crit Rev Oncog. 2017;22:313–21.
Cheng JC, Chang HM, Leung PCK. Connective tissue growth factor mediates TGF-β1-induced low-grade serous ovarian tumor cell apoptosis. Oncotarget. 2017;8:85224–33.
Brown JA, Schober M. Cellular quiescence: how TGFβ protects cancer cells from chemotherapy. Mol Cell Oncol. 2018;5:e1413495.
Sferra R, Pompili S, Festuccia C, et al. The possible prognostic role of histone deacetylase and transforming growth factor beta/Smad signaling in high grade gliomas treated by radio-chemotherapy: a preliminary immunohistochemical study. Eur J Histochem. 2017;61:2732.
Sun C, Wang FJ, Zhang HG, Xu XZ, Jia RC, Yao L, Qiao PF. miR-34a mediates oxaliplatin resistance of colorectal cancer cells by inhibiting macroautophagy via transforming growth factor-beta/Smad4 pathway. World J Gastroenterol. 2017;23:1816–27.
Rosen EM, Fan S, Pestell RG, Goldberg ID. BRCA1 gene in breast cancer. J Cell Physiol. 2003;196:19–41.
Li H, Sekine M, Seng S, Avraham S, Avraham HK. BRCA1 interacts with Smad3 and regulates Smad3-mediated TGF-beta signaling during oxidative stress responses. PLoS ONE. 2009;4:e7091.
Capoluongo E, Ellison G, Lopez-Guerrero JA, Penault-Llorca F, Ligtenberg MJL, Banerjee S, Singer C, Friedman E, Markiefka B, Schirmacher P, Büttner R, van Asperen CJ, Ray-Coquard I, Endris V, Kamel-Reid S, Percival N, Bryce J, Röthlisberger B, Soong R, de Castro DG. Guidance statement on BRCA1/2 tumor testing in ovarian cancer patients. Semin Oncol. 2017;44:187–97.
Mir R, Javid J, Al Balawi IA, Alkharsah KR, Hadi MA, Rahman MA, Hamoud E, Al Alawi Y, Al Zahrani ABM, Abu-Duhier FM. A germline mutation in the BRCA1 3′UTR variant predicts susceptibility to breast cancer in a Saudi Arabian population. Asian Pac J Cancer Prev. 2018;19:859–66.
Rojas V, Hirshfield KM, Ganesan S, Rodriguez-Rodriguez L. Molecular characterization of epithelial ovarian cancer: implications for diagnosis and treatment. Int J Mol Sci. 2016;17:2113.
Matsumoto T, Yokoi A, Hashimura M, Oguri Y, Akiya M, Saegusa M. TGF-beta-mediated LEFTY/Akt/GSK-3beta/Snail axis modulates epithelial-mesenchymal transition and cancer stem cell properties in ovarian clear cell carcinomas. Mol Carcinog. 2018;57(8):957–67.
Massague J. TGF-beta signal transduction. Annu Rev Biochem. 1998;67:753–91.
Cheng JC, Auersperg N, Leung PC. TGF-beta induces serous borderline ovarian tumor cell invasion by activating EMT but triggers apoptosis in low-grade serous ovarian carcinoma cells. PLoS ONE. 2012;7:e42436.
Zhou HH, Chen L, Liang HF, Li GZ, Zhang BX, Chen XP. Smad3 sensitizes hepatocelluar carcinoma cells to cisplatin by repressing phosphorylation of AKT. Int J Mol Sci. 2016;17:610.
Dubrovska A, Kanamoto T, Lomnytska M, Heldin CH, Volodko N, Souchelnytskyi S. TGFbeta1/Smad3 counteracts BRCA1-dependent repair of DNA damage. Oncogene. 2005;24:2289–97.
Authors’ contributions
YW and JX generated and analyzed the data. JW and YJ designed the experiments and drafted the manuscript. All authors read and approved the final manuscript.
Acknowledgements
Thanks are due to the Reproductive Medical Center for assistance with the experiments.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Consent for publication
Not applicable.
Ethics approval and consent to participate
All experiments and animal procedures were approved by the Ethics Committee of Tongji Hospital, Tongji University School of Medicine. All surgical procedures were performed under anesthesia and every effort was made to minimize suffering.
Funding
This work was supported by the Project of National Natural Science Fund of China (Grant No. 81302254); Shanghai Key Laboratory of Female Reproductive Endocrine Related Diseases (Grant No. 17DZ2273600).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wang, Y., Xiang, J., Wang, J. et al. Downregulation of TGF-β1 suppressed proliferation and increased chemosensitivity of ovarian cancer cells by promoting BRCA1/Smad3 signaling. Biol Res 51, 58 (2018). https://doi.org/10.1186/s40659-018-0205-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40659-018-0205-4