
Abstract
As one of the leading causes of death, hematologic malignancies are associated with an ever-increasing incidence, and drug resistance and relapse of patients after treatment represent clinical challenges. Therefore, there are pressing demands to uncover biomarkers to indicate the development, progression, and therapeutic targets for hematologic malignancies. Circular RNAs (circRNAs) are covalently closed circular-single-stranded RNAs whose biosynthesis is regulated by various factors and is widely-expressed and evolutionarily conserved in many organisms and expressed in a tissue−/cell-specific manner. Recent reports have indicated that circRNAs plays an essential role in the progression of hematological malignancies. However, circRNAs are difficult to detect with low abundance using conventional techniques. We need to learn more information about their features to develop new detection methods. Herein, we sought to retrospect the current knowledge about the characteristics of circRNAs and summarized research on circRNAs in hematological malignancies to explore a potential direction.
Similar content being viewed by others


Introduction
Hematologic malignancies, which represent a highly heterogeneous set of blood, bone marrow, and organ-associated diseases, are among the most common neoplasms, with acute leukemia, chronic leukemia, multiple myeloma, lymphoma, and malignant tumors histiocytosis, myeloproliferative disorders to name a few. Moreover, patients plagued by hematological malignancies present with a higher risk for infections and increased relapse rates. Further adding to the plight, acquired drug resistance associated with hematological malignancies poses a clinical challenge. The hard-done work of our peers has illustrated the aberrant expression of various non-coding RNAs (ncRNAs) in the pathogenesis of hematologic malignancies, with several ncRNAs being implicated in the regulation of gene expression at multiple levels, including transcription, translation, and epigenetic modification, thus exerting a myriad of cancer-promoting or cancer-suppressing functions. Meanwhile, ncRNA-based investigations have further reported the involvement of circRNAs in various pathophysiological processes of blood cells, such as blood cell differentiation, proliferation, and apoptosis of blood cells, in addition to participating in the occurrence, development and prognosis of hematological malignancies [1].
Circular RNAs, which contain a covalently closed single-stranded RNA, pertain to a category of ncRNAs and are widely expressed and evolutionarily conserved in a wide array of organisms. The first report about circRNAs was published by Sanger HL et al. in 1976 [2], and wherein circRNA molecules were regarded as a mistake-splicing by-product. In 1979, Hsu MT et al. [3] documented the same structure of RNA molecules located in the cytoplasm of eukaryotic cells and termed them as circRNAs. Nevertheless, limitations of detection technology rendered circRNAs as “transcriptional noise” with little immediate attention for years. However, in 1993, Capel et al. [4] illustrated that circRNA SRY was more highly expressed relative to linear SRY in the testis of mice and capable of protein translation. Thereafter, a flurry of investigations has followed and revealed that circRNAs play a crucial part in numerous physiological processes. Furthermore, extensive studies in regard to the mechanism of circRNA in various cancers are already underway.
CircRNAs are known to exhibit solid potential as a predictive, diagnostic and prognostic biomarker, especially their detectability in liquid biopsy samples, including plasma, saliva, and urine. Nevertheless, research related to the role of circRNAs in hematological malignancies is in its infancy and still has a long way to go. In retrospect, the current study summarizes the current knowledge on the biogenesis, regulation, and function of circRNAs and their clinical potential as biomarkers, therapeutic targets, and collaborative drug targets in hematological tumors.
The biosynthesis of circRNAs
The biosynthesis of circRNAs is inherently dissimilar from the production of mRNAs. Nigro et al. [5] previously documented a disordered RNA transcript, which possessed similar co-splicing sites of DCC (deleted in colorectal cancer) while also containing exons sequences and structures highly-different from DCC. Moreover, the latter research suggested that the by-product might perform a potential function in biological evolution but failed to carry out a further in-depth investigation.
The advent of enhanced technologies, like circRNA microarray and high-through sequencing, has further revealed three biosyntheses mechanisms [6] of circRNAs (Fig. 1): first, back-splicing circularization: Back-splicing is a phenomenon seen in a vast majority of eukaryotes, wherein small nuclear ribonucleoproteins (snRNPs) catalyze the pre-mRNA 5′ donor site to attack 3′ receptor site. Subsequently, a 3′-5′ phosphodiester bond formation ensues, which can be used to connect circRNAs covalently. Second, intron-driven circularization: Circularizable exons are flanked by long introns rich in ALU components to enhance complementary pairing and form repeatedly reverse-complementary secondary structures. Subsequently, the upstream and downstream exons covalently form circRNAs. Interestingly, the report published by Ashwal-Fluss R et al. [7, 8] indicated that flanking introns participated in circRNA generation and the quantity of ALUs exerted a crucial role in circularization efficiency. Third, lariat-driven circularization: The exon splicing donor and receptor combine as a “lariat intermediate,” followed by spliceosome removal of redundant introns. It is worth noting that the accelerated generation of lariat RNAs can be mediated by templates and lariat RNAs, such that lariat RNAs are also capable of generating circRNAs and linear branched-chain RNAs via the debranching enzyme [9].

Biosynthesis of circRNAs. CircRNAs can be directly generated by back-splicing; Intron-driven circularization can be inhibited by RNA helicase DHX9; Lariat-driven circularization generates a lariat structure. Spliceosome can splice to generate circRNAs and lariat-introns RNAs. Lariat-introns RNAs can be degraded, but some lariat-introns RNAs can escape degradation to form ciRNAs
A variety of splicing factors regulate the process of circRNA generation, including cis−/trans-acting splicing regulatory elements [6], which are combined with splicing factors to enhance or inhibit the expression of circRNAs. However, it is worth noting that splicing factors in circRNA generation may confer a different role in linear RNAs. Besides, circRNAs possess the ability to regulate circularization by binding to RNA-binding proteins (RBPs). For instance, circCAMSAP1 in colorectal cancer can directly bind with Epithelial-splicing regulatory protein 1 (ESRP1) to promote circularization of circCAMSAP1 [10]; and QKI can induce normal linear transcripts de novo synthesis and generate circRNAs [11]; whereas, DHX9 can inhibit the formation of ALU’s complementary structures by binding to ALU elements to prevent circRNA production [12].
Moreover, some lariat-introns RNAs can escape from degradation to form circular intronic RNAs (ciRNA). A consensus motif is containing a 7 nt GU-rich element near the 5′ splice site and an 11 nt C-rich element close to the branchpoint in ciRNA-producing introns can help intron lariat escape from the debranching enzyme [13].
Additionally, circRNA biosynthesis can also be affected by some specific physiological conditions, such as immune response [14]. RNase L is activated to rapidly degrade circRNAs with virus infection, which leads to activation of the PRK pathway and leading to a cascade of activation of innate immunity.
The characteristics of circRNA
CircRNAs are widely-expressed and conserved
CircRNAs are expressed in a wide array of organisms, like animals, plants, bacteria, archaea, etc. [4, 15, 16], and sequencing of circRNAs in different organisms has revealed that circRNAs are more conservative than linear RNAs. In addition, higher expression of circRNAs is associated with enhanced conservative properties. For example, circRNAs TTBK2, Ttbk2 and Asator are highly expressed in drosophila’s brains were also found in mammalian brains and exhibit identical sequences [16, 17]. Meanwhile, high conservatism of circRNAs is indicated in exons as reflected in flanking intron sequences of circularization exons, exact back-splicing sites, and expression patterns.
CircRNAs are expressed with exquisite tissue/cell-specificity
CircRNAs are highly expressed in mammalian brain tissues. Like circMfsd6 and circZfp609, some circRNAs present with significantly higher expression relative to their homologous linear RNAs [17]. Further investigation by Rybak-Wolf et al. has revealed that circRNAs are primarily enriched in axons, dendrites, and synapses [17, 18]. The types and expressions of circRNAs are transformed in diverse differentiation stages to exert crucial functions in neuronal differentiation and maintenance. Meanwhile, the study carried out by You et al. has reported that circRNAs are specifically distributed in different subcellular organelles and developmental stages of cells with qRT-PCR and RNA-seq [18].
High stability is a common feature of circRNAs
With advantages such as cyclic structures and lack of free ends, circRNAs are hardly recognized by the exonucleases to resist degradation from the exonuclease exposure. CircRNAs are also known to exhibit stability following treatment with actinomycin D treatment for 48 hours [16]. In addition, circRNAs have a longer half-life, approximately 2.5 times longer than their counterparts linear RNAs [19, 20]. The work of Dongming Liang et al. [21] has previously indicated that damaging RNA splicing mechanism can promote circRNAs to switch to a non-promoter structurally dependent transcription mechanism. These aforementioned evidence indicated that circRNAs maintained high stability and the potential for long-term regulation of cell behaviors (Fig. 2).

Characteristics of circRNAs. A CircRNAs are widely expressed in various organisms; B CircRNAs are expressed in a tissue-specific manner; C. CircRNAs with high stability can resist degradation from the exonuclease
The functions of circRNAs
CircRNAs regulate transcription
Despite the differences in the biosynthesis of circRNAs and linear RNAs, both are catalyzed by conventional splicing signals and canonical splicing system so that circRNAs can serve as an “mRNA trap” to interfere with linear counterparts mRNA competing with the binding site of the spliceosome [22]. Moreover, flanking introns on circularization exons on pre-mRNA can also bind with the spliceosome and influence the fate of mRNA biosynthesis [7].
Moreover, the study carried out by Xiang Li et al. has illustrated that ciRNA ci-ankrd52 maintains R-loop structure to facilitate transcriptional elongation across its producing locus [23].
CircRNAs act as miRNA sponges
After uncovering the presence of miRNA-binding sites, further experimentation has revealed that circRNAs function as “miRNA sponges” and indirectly regulate the expression of mRNAs [24,25,26]. For instance, circRNA MAT2B [25] sponged miR-338-3p to promote glycolysis and malignancy in hepatocellular carcinoma (HCC), while circPAN3 [26] mediated drug resistance in acute myeloid leukemia through binding with miR-153-5p/miR-183-5p.
CircRNAs further possess multiple miRNA response elements of different miRNAs, such as circHIPK3 [27], while there is also evidence to suggest that multiple circRNAs can regulate the same miRNA simultaneously. Subsequently, the miRNAs initially targeted mRNA will bind with circRNAs, and form a circRNA-miRNA-mRNA competing endogenous RNAs (ceRNA) network to regulate mRNA expression. Except for sponges, circRNAs can act as reservoirs to enrich miRNAs and enhance the effect. As an example of the latter, circ-DOCK5 reserve miR-627-3p to repress metastasis in squamous cell carcinoma [28]. Moreover, circRNAs are capable of regulating regulated miRNA biosynthesis via changing the intracellular localization of DICER [29].
CircRNAs interact with proteins
Most circRNAs, which contain multiple RBPs binding sites, were located in the cytoplasm. Herein, circRNAs could work as “decoys” or “scaffolds” to bind to RBPs, isolate RBPs from their target molecules, mediate the intracellular localization of RBPs [30], and thereby regulate RBP biosynthesis, transport, and influence subcellular biological processes [1]. CircRNAs might directly or indirectly crosstalk by transcription, for example, circRHOT1 [31] can recruit TIP60 to NR2F6 promoter to activate NR2F6 to enhance HCC progression. Meanwhile, circFoxo3 can interact with MDM2 and p53 to augment MDM2-induced p53 degradation [32].
Exosomal circRNAs take part in cellular communication
Exosomes are extracellular vesicles with a size range of ~ 40 to 160 nm (average ~ 100 nm) in diameter [33], and serve as an intercellular transit system with pleiotropic functions. In 2015, the study performed by Li et al. [34] has illustrated that circRNAs are enriched and stable in exosomes by RNA-seq. In addition, the previously mentioned study also documented more than 1000 circRNAs in human serum exosomes.
Owing to the stability of circRNAs, exosomal circRNAs can be carried to participate in cellular information transmission and disease progression. For example, serum exosomal circRNA-104484 and circRNA-104670 are significantly enhanced in sepsis [35], and possess the potential as diagnostic markers in sepsis. Exosomal circRNAs can also regulate cancer development; for instance, exosomal circRNA-100338 is capable of regulating angiogenesis and metastasis of HCC, while serum exosomal circRNA-100338 can predict lung metastasis of HCC patients following curative hepatectomy [36]. Similarly, exosomal circRNA-133 of CRC patients can transport normoxic cells to hypoxic cells, regulate the E-cadherin membrane distribution, and promote cancer metastasis via miR-133a/GEF-H1/RhoA axis [37].
CircRNAs regulate epigenetics
Epigenetic regulation is relevant to promote the occurrence and development of multiple diseases in the most abundant type of cells. Currently, numerous data have indicated that N6-methyladenosine modification can modulate the expression, distribution and functions of circRNAs [38]. On the other hand, some circRNAs are previously reported to regulate the expression, functions and proteins interaction of m6A, such as circMAP2K4 sponging miR-139-5p can regulate the expression and activity of YTHDF1 [39]. Meanwhile, circNOTCH1 competitively binds with METTL14 to release NOTCH1 mRNA [40].
CircRNAs translate proteins
Due to a lack of the cap structure and poly adenylate tail, circRNAs were translated by spliceosome activity-dependent and cap-independent methods. The internal ribosome entry site (IRES) [41], located in the 5′ UTR, serves as the main pathway for circRNA translation. IRES exerts its functions by several mechanisms, including analogous RNA structures interacting with 18S rRNA and binding with IRES trans-acting factor ITAF. Losing the cap structure of circRNAs drives IRES activation and recruits IRES factors to initiate circRNA translation. Simultaneously, activated IRES can enhance circRNA translation efficiency upon undergoing stress, such as heat shock, cancer, and hypoxia. IRES has no advantage in overactivity and number in vivo, and thus circRNAs exhibit a lower translation efficiency than linear RNAs, which usually are ten times as much as circRNAs [42]. Nevertheless, circRNA translation can be enhanced in various ways, like m6A modification [43] in eukaryotic circRNAs to help achieve circRNA translation.
It was interesting to explore the discrepancy in translated methods between circRNAs and linear RNAs by studying translated products from circRNAs. A novel translation mechanism previously indicated to produce particular proteins: rolling circle translation [20, 44, 45]. Covalently closed circRNAs are translated when the number of nucleotides contained in the open reading frame (except for the stop codon) are an integer multiple of 3, whereas the termination codons did not take participate in translation (Fig. 3).

Functions of circRNAs. CircRNAs can maintain R-loop structure to facilitate mRNA transcription and indirectly regulate mRNA transcription by recruiting proteins or acting as ceRNA to compete with mRNA. Besides, circRNAs act as a protein scaffold to mediate protein-protein interactions, protein localization, and protein transport. Moreover, circRNAs code proteins and regulate nearby cells by exosome transport
CircRNAs in hematological malignancies
More and more evidence has further classified ncRNAs and associated with hematopoiesis and hematological malignancy initiation, including miRNAs, long non-coding RNAs (lncRNAs), and circRNAs.
CircRNAs in blood cell differentiation
Hematopoietic stem cell (HSC) differentiation is strictly regulated by multiple factors, including transcription factors, ncRNAs (including circRNAs), and other factors. Accumulating reports have shown that an array of circRNAs are expressed explicitly in the continuously hematopoietic process [46,47,48]. CircSPI1 can be up-regulated to inhibit myeloid differentiation of acute myeloid leukemia (AML) cells [49]. Moreover, quantitative detecting of circRNAs in various blood cells by Nicolet et al. [47] illustrated the presence of changes in circRNA type and quantity in addition to the maturation and differentiation of HSCs. CircRNAs are similarly up-regulated in mature erythrocytes and platelets, and Nicolet et al. [46] further indicated their association with translation and housekeeping function exertion in mature erythrocytes. However, there is still little evidence on whether circRNAs can translate proteins in red blood cells, questioning their potential as molecular markers to distinguish differentiation stages and requiring further elaboration.
CircRNAs act as biomarkers in hematological tumors
CircRNAs are regarded as diagnostic and prognostic biomarkers to indicate the development of hematological tumors and therapeutic responses. In 2012, hundreds of circRNAs were documented in AML patients by circRNA microarray and genome-wide microarray analysis [48]. More and more circRNAs have been attested to serve as potential clinical biomarkers. For instance, circAML1, which was transcribed from AML1, which acts as an oncogene of AML, is up-regulated in the bone marrow and peripheral blood [50], while being significantly down-regulated in bone marrow-derived exosomes of essential thrombocythemia patients. Those studies hinted that circAML1 maybe serve as a biomarker in the hematological system [51]. Moreover, high-throughput analysis and bioinformatics analyses have screened and validated differentially expressed circRNAs in AML, underscoring the ability of circ-0004277 as a potential diagnostic marker and therapeutic target for AML [52].
Accumulating evidence further indicates that circRNAs also contribute to discovering the development of disease. For example, circ-VIM was positively correlated with AML progression, wherein circ-VIM served as a pro-oncogenic circRNA to accelerate the progression of AML [53]. Meanwhile, circ-0000190 can serve as an independent risk factor for risk stratification and the prognosis of MM [54], while circ-0004277 [52] was previously associated with risk status and treatment response in AML. Hsa-circ-100352, hsa-circ-104056, and hsa-circ-102817 are differentially expressed and promote the progression of MDS and are related to the survival and prognosis of MDS [55,56,57].
On the other hand, up-regulation of circ-ITCH was was negatively associated with ISS in MM patients and positively correlated with progression-free survival (PFS) and overall survival (OS) [58]. Similarly, circ-VIM was negatively associated with OS and leukemia-free survival (LFS), and reported to serve as an independent poor prognostic factor for OS and LFS in AML patients [59].
CircRNAs regulate the proliferation, apoptosis, cell cycle, invasion and migration of hematological malignancies
Proliferation misregulation has been indicated as one of the critical factors in hematological malignancies. Recent studies demonstrated the correlation between circRNA expression and cell proliferation in hematological malignancies. For instance, circ-0004136 [60] promoted AML cell proliferation by sponging miR-142, while circ-0001947 [61] up-regulated CREBRF to repress AML cell proliferation by inhibiting hsa-miR-329-5p. In addition, circPOLA2 [62] and circ-0121582 [63] were over-expressed in AML and promoted cell proliferation by targeting their miRNAs. Similarly, circMYBL2 in AML serves as a potential therapeutic target and it regulates the level of FLT3 kinase by interacting with PTBP1 to inhibit proliferation [64].
Avoiding apoptosis represents a crucial factor in promoting the hematological malignancies process. Accumulating studies have indicated that circRNAs can regulate cell apoptosis and cell cycle progression during hematological malignancies. For example, both circCDYL [65] and its targeted gene YAP undergo up-regulation to inhibit apoptosis. Moreover, circRNAs can serve as essential factors for the evasion of the death of cells, as circKEL [66] inhibits the apoptosis of AML via sponging miR-335-5p, and up-regulation of circ-0009910 [67, 68] in CML and AML was associated with inhibition of apoptosis by the miR-34a-5p/ULK1 axis. Knockdown of circPTK2 [69] can augment the expression of miR-330-5p and decrease the targeted mRNA FOXM1 to promote apoptosis.
In addition to regulating apoptosis and proliferation, circRNAs can also exert control over cell cycle progression to influence the progression of hematological malignancies process. CircCBFB [70] was previously associated with the cell cycle to facilitate CLL development. Moreover, circ-0002483 [71] could further accelerate the cell cycle period by sponging miR-758-3p.
Meanwhile, circRNAs have been shown to regulate the invasion and migration of cells and further promote the progression of hematological malignancies, such as overexpressing circ-0069767 decreases the invasion and migration capacities of MM cells by sponging miR-636 to regulate the expression of K-RAS [72], whereas circ-0000142 enhances the invasion of MM cells via the circ-0000142/miR-610/AKT3 axis [73], etc.
CircRNAs influence chemo-sensitivity in hematological malignancies
Chemoresistance underlies one of the the key causes of the progression and relapse of hematological malignancies. Some outcomes have suggested that circRNAs can assist in overcoming chemoresistance. For example, circANXA2 [74] was associated with chemoresistance of cytarabine and daunorubicin in AML. Similarly, circPAN3 [26] induced chemoresistance of adriamycin (ADM) via sponging miR-153-5p/miR-183-5p, while bortezomib (BTZ)-circ-0003489 [75] may regulate resistance in MM. Circ-0009910 triggered the activation of autophagy by sponging miR-34a-5p to promote imatinib resistance in chronic myeloid leukemia [68].
Therefore, regulating chemo-sensitivity of hematological malignancies via targeting circRNAs can pave the way for novel clinical therapy and a breakthrough in longer survival time for patients. Table 1
F-circRNAs in hematological tumors
Recently, a novel form of circRNAs--fusion circRNAs (f-circRNAs) was published [57, 88,89,90], f-circRNAs derived from cancer-associated chromosomal translocation were indicated as one of the critical causes of hematological malignancies. F-circRNAs can function as proto-oncogenes, and up-regulation of f-circRNAs promote the proliferation and clonogenicity, and inhibit apoptosis of leukemia cells, whereas silencing of f-circRNAs is associated with reversal of these phenotypes. A comparison of in vitro and in vivo models for the function of f-circRNAs further highlighted that f-circrNAs can directly lead to leukemia occurrence and promote leukemia formation, maintain the progression, and improve the survival ability of leukemia cells. For instance, circAF4 sponges miR-128-3p to promote cell proliferation and inhibit apoptosis in leukemia [91], whereas circBA9.3 derived from BCR-ABL1 can increase tyrosine kinase activity that promotes resistance against resistance TKI therapy [92]. Moreover, f-circRNA generated from chromosomal translocation can produce many proteins that were entirely different from others, in order to mediate leukemia development, progression, prognostic, and chemoresistance [57, 91] Fig. 4.

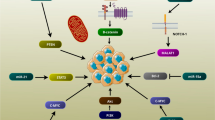
CircRNA in the hematological system. A The expression of circRNAs was positively correlated with cell differentiation during erythrocyte differentiation; B CircRNAs play functions in hematological tumors; C Chromosomal translocation derived f-circRNA, which could translate fusion proteins to promote leukemia
Detecting circRNAs
Accumulating studies have shown that circRNAs only exhibit 1/10000–1/100 of than the expression of their linear counterpart RNAs [93], and because of the same base sequence with counterpart linear RNAs, circRNAs are harder to isolate from total RNA. Accordingly, RNase R is commonly employed to digest the linear RNAs for enriching circRNAs, but it is worth noting that circRNAs can also be digested by long-term RNase R treatment and treated circRNAs should be distinguished from the secondary structure of linear RNAs. For further investigation of circRNAs, high-throughput RNA sequencing can be adopted to detect the differential expression of circRNAs, and bioinformatics (like STAR, circBase) allow the analyses of the functions of passway and the interaction of proteins, etc. Various methods were administrated in the circRNA study. Traditionally used methods in circRNA study are listed as follow [15, 24, 25, 48, 54, 94,95,96] (Table 2, 3 and 4).
Summary
Herein, the current study revealed the function and clinical significance of circRNAs, and partly recognized their roles in cancer. The biosynthesis of circRNAs is regulated by a plethora of factors, which explains the involvement of circRNAs in various physiological processes in an organism. Moreover, the interaction between circRNAs and various molecules serves as a mean for participation in transcription, gene expression, protein interaction, and other mechanisms.
In particular, existing evidence indicates that circRNAs bears great responsibility for hematopoietic stem cell differentiation and development. CircRNAs are further widely and specifically expressed in blood cells, and exhibit stability in mature erythrocytes. Recent investigations have also shown that circRNAs are differentially expressed in multiple hematologic tumors, and accordingly correlated with the occurrence, development, and prognosis of multiple hematologic tumors. Consequently, it would be plausible to suggest that circRNAs possess the ability to serve as novel biomarkers of hematological malignancies.
Furthermore, up-and-coming researches have indicated that circRNAs exert their functions by accumulating high expression in non-nuclear blood cells. Existing studies have expounded on the ability of circRNAs to promote or inhibit the proliferation, migration and invasion, cell cycle, and apoptosis in hematological malignancies. Meanwhile, circRNA-mediated chemoresistance and exosomal circRNA-mediated tumor microenvironment regulation of adjacent cells or tissues also represent potential directions for hematological malignancies treatment. Moreover, circRNAs, which are differently expressed or newly discovered, can also act as appealing biomarkers for the understanding of the development of hematological diseases. Overall, the emergence of f-circRNAs has opened an innovative door for hematological tumors’ diagnosis and therapy.
CircRNAs also exhibit an excellent promising clinical translation in hematological tumors, however, several biological questions and clinical challenges need to be addressed before their translational potential can be realized. For instance, most of the current studies are based on the available approaches for detection and aimed at the function of the “miRNA sponge,” and the precise mechanisms of circRNAs in hematological malignancies remain elusive. Given that the studies focusing on circRNAs have been hardly abundant, future studies should focus on enriching circRNAs, developing sensitive techniques, etc.
Overall, circRNAs represent potential therapeutic targets that exert their crucial influence on the occurrence, development, prognostic, and treatment of hematological tumors, but many of them are yet to be unveiled. Hopefully, more mechanisms of circRNAs could be clarified shortly, and research could be further expanded.
Availability of data and materials
Not applicable.
Abbreviations
- ADM:
-
Adriamycin
- AML:
-
Acute myelocytic leukemia
- BTZ:
-
Bortezomib
- ceRNA net:
-
Competing endogenous RNAs net
- circRNA:
-
Circular RNA
- CLL:
-
Chronic lymphocytic leukemia
- DCC:
-
Deleted in colorectal cancer
- ddPCR:
-
Droplet Digital PCR
- DHX9:
-
DEAH-box helicase 9
- ESRP1:
-
Epithelial-splicing regulatory protein 1
- f-circRNA:
-
Fusion circRNA
- FISH:
-
Fluorescence in situ hybridization
- HSCs:
-
Hematopoietic stem cells
- IRES:
-
Internal ribosome entry site
- ISS:
-
International Staging System
- LFS:
-
Leukemia-free survival
- MDS:
-
Myelodysplastic syndromes
- miRNA:
-
microRNA
- MM:
-
Multiple myeloma
- ncRNA:
-
Non-coding RNA
- OS:
-
Overall survival
- PFS:
-
Progression-free survival
- Pre-mRNA:
-
Heterogeneous nuclear RNA
- QKI:
-
Quaking protein
- qRT-PCR:
-
Quantitative Reverse Transcription-Polymerase Chain Reaction
- RBP:
-
RNA-binding protein
- RCA reaction:
-
Rolling Circle Amplification reaction
- SnRNP:
-
Ribonucleoproteins small nuclear
- UTR:
-
Untranslated regionmultiple myelom
References
Bonizzato A, Gaffo E, Te Kronnie G, Bortoluzzi S. CircRNAs in hematopoiesis and hematological malignancies. Blood Cancer J. 2016;6(10):e483.
Sanger HL, Klotz G, Riesner D, Gross HJ, Kleinschmidt AK. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc Natl Acad Sci U S A. 1976;73(11):3852–6.
Hsu MT, Coca-Prados M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature. 1979;280(5720):339–40.
Capel B, Swain A, Nicolis S, Hacker A, Walter M, Koopman P, et al. Circular transcripts of the testis-determining gene Sry in adult mouse testis. Cell. 1993;73(5):1019–30.
Nigro JM, Cho KR, Fearon ER, Kern SE, Ruppert JM, Oliner JD, et al. Scrambled exons. Cell. 1991;64(3):607–13.
Wang Y, Wang Z. Efficient backsplicing produces translatable circular mRNAs. RNA. 2015;21(2):172–9.
Ashwal-Fluss R, Meyer M, Pamudurti NR, Ivanov A, Bartok O, Hanan M, et al. circRNA biogenesis competes with pre-mRNA splicing. Mol Cell. 2014;56(1):55–66.
Liang D, Wilusz JE. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014;28(20):2233–47. https://doi.org/10.1101/gad.251926.114 Epub 2014 Oct 3.
Carriero S, Damha MJ. Template-mediated synthesis of lariat RNA and DNA. J Org Chem. 2003;68(22):8328–38.
Zhou C, Liu HS, Wang FW, Hu T, Liang ZX, Lan N, et al. circCAMSAP1 promotes tumor growth in colorectal Cancer via the miR-328-5p/E2F1 Axis. Mol Ther. 2020;28(3):914–28.
Conn SJ, Pillman KA, Toubia J, Conn VM, Salmanidis M, Phillips CA, et al. The RNA binding protein quaking regulates formation of circRNAs. Cell. 2015;160(6):1125–34.
Yu J, Xu QG, Wang ZG, Yang Y, Zhang L, Ma JZ, et al. Circular RNA cSMARCA5 inhibits growth and metastasis in hepatocellular carcinoma. J Hepatol. 2018;68(6):1214–27.
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin QF, Xing YH, et al. Circular intronic long noncoding RNAs. Mol Cell. 2013;51(6):792–806. https://doi.org/10.1016/j.molcel.2013.08.017 Epub 2013 Sep 12.
Liu CX, Li X, Nan F, Jiang S, Gao X, Guo SK, et al. Structure and degradation of circular RNAs regulate PKR activation in innate immunity. Cell. 2019;177(4):865–880.e21.
Patop IL, Wüst S, Kadener S. Past, present, and future of circRNAs. EMBO J. 2019;38(16):e100836.
Jeck WR, Sorrentino JA, Wang K, Slevin MK, Burd CE, Liu J, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA. 2013;19(2):141–57.
Rybak-Wolf A, Stottmeister C, Glažar P, Jens M, Pino N, Giusti S, et al. Circular RNAs in the mammalian brain are highly abundant, conserved, and dynamically expressed. Mol Cell. 2015;58(5):870–85.
You X, Vlatkovic I, Babic A, Will T, Epstein I, Tushev G, et al. Neural circular RNAs are derived from synaptic genes and regulated by development and plasticity. Nat Neurosci. 2015;18(4):603–10. https://doi.org/10.1038/nn.3975 Epub 2015 Feb 25.
Enuka Y, Lauriola M, Feldman ME, Sas-Chen A, Ulitsky I, Yarden Y. Circular RNAs are long-lived and display only minimal early alterations in response to a growth factor. Nucleic Acids Res. 2016;44(3):1370–83.
Jeck WR, Sharpless NE. Detecting and characterizing circular RNAs. Nat Biotechnol. 2014;32(5):453–61.
Liang D, Tatomer DC, Luo Z, Wu H, Yang L, Chen LL, et al. The output of protein-coding genes shifts to circular RNAs when the pre-mRNA processing machinery is limiting. Mol Cell. 2017;68(5):940–954.e3.
Starke S, Jost I, Rossbach O, Schneider T, Schreiner S, Hung LH, et al. Exon circularization requires canonical splice signals. Cell Rep. 2015;10(1):103–11.
Li X, Zhang JL, Lei YN, Liu XQ, Xue W, Zhang Y, et al. Linking circular intronic RNA degradation and function in transcription by RNase H1. Sci China Life Sci. 2021;64(11):1795–809. https://doi.org/10.1007/s11427-021-1993-6 Epub 2021 Aug 25.
Cheng X, Zhang L, Zhang K, Zhang G, Hu Y, Sun X, et al. Circular RNA VMA21 protects against intervertebral disc degeneration through targeting miR-200c and X linked inhibitor-of-apoptosis protein. Ann Rheum Dis. 2018;77(5):770–9.
Li Q, Pan X, Zhu D, Deng Z, Jiang R, Wang X. Circular RNA MAT2B promotes glycolysis and malignancy of hepatocellular carcinoma through the miR-338-3p/PKM2 Axis under hypoxic stress. Hepatology. 2019;70(4):1298–316.
Shang J, Chen WM, Wang ZH, Wei TN, Chen ZZ, Wu WB. CircPAN3 mediates drug resistance in acute myeloid leukemia through the miR-153-5p/miR-183-5p-XIAP axis. Exp Hematol. 2019;70:42–54.e3.
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen B, et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat Commun. 2016;7:11215. https://doi.org/10.1038/ncomms11215.
Meng L, Zheng Y, Liu S, Ju Y, Ren S, Sang Y, et al. ZEB1 represses biogenesis of circ-DOCK5 to facilitate metastasis in esophageal squamous cell carcinoma via a positive feedback loop with TGF-β. Cancer Lett. 2021;519:117–29.
Bronisz A, Rooj AK, Krawczyński K, Peruzzi P, Salińska E, Nakano I, et al. The nuclear DICER-circular RNA complex drives the deregulation of the glioblastoma cell microRNAome. Sci Adv. 2020;6(51):eabc0221.
Yang Q, Li F, He AT, Yang BB. Circular RNAs: expression, localization, and therapeutic potentials. Mol Ther. 2021;S1525-0016(21):00018–6.
Wang L, Long H, Zheng Q, Bo X, Xiao X, Li B. Circular RNA circRHOT1 promotes hepatocellular carcinoma progression by initiation of NR2F6 expression. Mol Cancer. 2019;18(1):119.
Du WW, Fang L, Yang W, Wu N, Awan FM, Yang Z, et al. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ. 2017;24(2):357–70.
Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977. https://doi.org/10.1126/science.aau6977.
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J, et al. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. 2015;25(8):981–4. https://doi.org/10.1038/cr.2015.82 Epub 2015 Jul 3.
Tian C, Liu J, Di X, Cong S, Zhao M, Wang K. Exosomal hsa_circRNA_104484 and hsa_circRNA_104670 may serve as potential novel biomarkers and therapeutic targets for sepsis. Sci Rep. 2021;11(1):14141. https://doi.org/10.1038/s41598-021-93246-0.
Huang XY, Huang ZL, Huang J, Xu B, Huang XY, Xu YH, et al. Exosomal circRNA-100338 promotes hepatocellular carcinoma metastasis via enhancing invasiveness and angiogenesis. J Exp Clin Cancer Res. 2020;39(1):20. https://doi.org/10.1186/s13046-020-1529-9.
Yang H, Zhang H, Yang Y, Wang X, Deng T, Liu R, et al. Hypoxia induced exosomal circRNA promotes metastasis of colorectal Cancer via targeting GEF-H1/RhoA axis. Theranostics. 2022;12(6):2635–6. https://doi.org/10.7150/thno.44419.
Xu J, Wan Z, Tang M, Lin Z, Jiang S, Ji L, et al. N6-methyladenosine-modified CircRNA-SORE sustains sorafenib resistance in hepatocellular carcinoma by regulating β-catenin signaling. Mol Cancer. 2020;19(1):163. https://doi.org/10.1186/s12943-020-01281-8.
Chi F, Cao Y, Chen Y. Analysis and validation of circRNA-miRNA network in regulating m6A RNA methylation modulators reveals CircMAP2K4/miR-139-5p/YTHDF1 Axis involving the proliferation of hepatocellular carcinoma. Front Oncol. 2021;11:560506. https://doi.org/10.3389/fonc.2021.560506.
Shen Y, Li C, Zhou L, Huang JA. G protein-coupled oestrogen receptor promotes cell growth of non-small cell lung cancer cells via YAP1/QKI/circNOTCH1/m6A methylated NOTCH1 signalling. J Cell Mol Med. 2021;25(1):284–96. https://doi.org/10.1111/jcmm.15997 Epub 2020 Nov 25.
Legnini I, Di Timoteo G, Rossi F, Morlando M, Briganti F, Sthandier O, et al. Circ-ZNF609 is a circular RNA that can be translated and functions in Myogenesis. Mol Cell. 2017;66(1):22–37.e9.
Pamudurti NR, Bartok O, Jens M, Ashwal-Fluss R, Stottmeister C, Ruhe L, et al. Translation of CircRNAs. Mol Cell. 2017;66(1):9–21.e7.
Yang Y, Fan X, Mao M, Song X, Wu P, Zhang Y, et al. Extensive translation of circular RNAs driven by N6-methyladenosine. Cell Res. 2017;27(5):626–41.
Liu Y, Li Z, Zhang M, Zhou H, Wu X, Zhong J, et al. Rolling-translated EGFR Variants Sustain EGFR Signaling and Promote Glioblastoma Tumorigenicity. Neuro Oncol. 2020;23:noaa279.
Nakamoto K, Abe N, Tsuji G, Kimura Y, Tomoike F, Shimizu Y, et al. Chemically synthesized circular RNAs with phosphoramidate linkages enable rolling circle translation. Chem Commun (Camb). 2020;56(46):6217–20.
Nicolet BP, Engels S, Aglialoro F, van den Akker E, von Lindern M, Wolkers MC. Circular RNA expression in human hematopoietic cells is widespread and cell-type specific. Nucleic Acids Res. 2018;46(16):8168–80.
Salzman J, Gawad C, Wang PL, Lacayo N, Brown PO. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS One. 2012;7(2):e30733.
Dostalova Merkerova M, Krejcik Z, Szikszai K, Kundrat D. Circular RNAs in hematopoiesis with a focus on acute myeloid leukemia and myelodysplastic syndrome. Int J Mol Sci. 2020;21(17):5972.
Wang X, Jin P, Zhang Y, Wang K. CircSPI1 acts as an oncogene in acute myeloid leukemia through antagonizing SPI1 and interacting with microRNAs. Cell Death Dis. 2021;12(4):297.
Wang Q, Yu G, He H, Zheng Z, Li X, Lin R, et al. Differential expression of circular RNAs in bone marrow-derived exosomes from essential thrombocythemia patients. Cell Biol Int. 2021;45(4):869–81. https://doi.org/10.1002/cbin.11534 Epub 2020 Dec 31.
Xu AN, Chen XH, Tan YH, Qi XL, Xu ZF, Zhang LL, et al. Identification of a novel circularized transcript of the AML1 gene. BMB Rep. 2013;46(3):163–8.
Li W, Zhong C, Jiao J, Li P, Cui B, Ji C, et al. Characterization of hsa_circ_0004277 as a new biomarker for acute myeloid leukemia via circular RNA profile and bioinformatics analysis. Int J Mol Sci. 2017;18(3):597.
Yi YY, Yi J, Zhu X, Zhang J, Zhou J, Tang X, et al. Circular RNA of vimentin expression as a valuable predictor for acute myeloid leukemia development and prognosis. J Cell Physiol. 2019;234(4):3711–9.
Feng Y, Zhang L, Wu J, Khadka B, Fang Z, Gu J, et al. CircRNA circ_0000190 inhibits the progression of multiple myeloma through modulating miR-767-5p/MAPK4 pathway. J Exp Clin Cancer Res. 2019;38(1):54.
Zhao X, Yin H, Li N, Zhu Y, Shen W, Qian S, et al. An integrated regulatory network based on comprehensive analysis of mRNA expression, gene methylation and expression of Long noncoding RNAs (lncRNAs) in myelodysplastic syndromes. Front Oncol. 2019;9:200.
Wu WL, Li S, Zhao GJ, Li NY, Wang XQ. Identification of circular RNAs as novel biomarkers and potentially functional competing endogenous RNA network for myelodysplastic syndrome patients. Cancer Sci. 2021;112(5):1888–98.
Guarnerio J, Bezzi M, Jeong JC, Paffenholz SV, Berry K, Naldini MM, et al. Oncogenic role of fusion-circRNAs derived from Cancer-associated chromosomal translocations. Cell. 2016;165(2):289–302.
Zhou H, Zhang J, Chen B, Liu H, Liu X, Sun Z, et al. Potential of circular RNA itchy E3 ubiquitin protein ligase as a biomarker and treatment target for multiple myeloma. Transl Cancer Res. 2020;9(1):335–45. https://doi.org/10.21037/tcr.2019.12.71.
Yi YY, Yi J, Zhu X, Zhang J, Zhou J, Tang X, et al. Circular RNA of vimentin expression as a valuable predictor for acute myeloid leukemia development and prognosis. J Cell Physiol. 2019;234(4):3711–9. https://doi.org/10.1002/jcp.27145 Epub 2018 Aug 28.
Yuan DM, Ma J, Fang WB. Identification of non-coding RNA regulatory networks in pediatric acute myeloid leukemia reveals circ-0004136 could promote cell proliferation by sponging miR-142. Eur Rev Med Pharmacol Sci. 2019;23(21):9251–8.
Han F, Zhong C, Li W, Wang R, Zhang C, Yang X, et al. Hsa_circ_0001947 suppresses acute myeloid leukemia progression via targeting hsa-miR-329-5p/CREBRF axis. Epigenomics. 2020;12(11):935–53.
Li H, Bi K, Feng S, Wang Y, Zhu C. CircRNA circ_POLA2 is upregulated in acute myeloid leukemia (AML) and promotes cell proliferation by suppressing the production of mature miR-34a. Cancer Manag Res. 2021;13:3629–37.
Chen JJ, Lei P, Zhou M. hsa_circ_0121582 inhibits leukemia growth by dampening Wnt/β-catenin signaling. Clin Transl Oncol. 2020;22(12):2293–302.
Sun YM, Wang WT, Zeng ZC, Chen TQ, Han C, Pan Q, et al. circMYBL2, a circRNA from MYBL2, regulates FLT3 translation by recruiting PTBP1 to promote FLT3-ITD AML progression. Blood. 2019;134(18):1533–46.
Chen F, Wang X, Fu S, Wang S, Fu Y, Zhang J, et al. Circular RNA circ-CDYL sponges miR-1180 to elevate yes-associated protein in multiple myeloma. Exp Biol Med (Maywood). 2020;245(11):925–32.
Wu ZJ, Sun Q, Gu DL, Wang LQ, Li JY, Jin H. Expression of circ-KEL in acute myeloid leukemia and its regulatory mechanisms in leukemic cells. Zhonghua Xue Ye Xue Za Zhi. 2021;42(3):230–7 Chinese.
Cao HX, Miao CF, Sang LN, Huang YM, Zhang R, Sun L, et al. Circ_0009910 promotes imatinib resistance through ULK1-induced autophagy by sponging miR-34a-5p in chronic myeloid leukemia. Life Sci. 2020;243:117255.
Wang D, Ming X, Xu J, Xiao Y. Circ_0009910 shuttled by exosomes regulates proliferation, cell cycle and apoptosis of acute myeloid leukemia cells by regulating miR-5195-3p/GRB10 axis. Hematol Oncol. 2021;39(3):390–400.
Yi L, Zhou L, Luo J, Yang Q. Circ-PTK2 promotes the proliferation and suppressed the apoptosis of acute myeloid leukemia cells through targeting miR-330-5p/FOXM1 axis. Blood Cells Mol Dis. 2021;86:102506.
Xia L, Wu L, Bao J, Li Q, Chen X, Xia H, et al. Circular RNA circ-CBFB promotes proliferation and inhibits apoptosis in chronic lymphocytic leukemia through regulating miR-607/FZD3/Wnt/β-catenin pathway. Biochem Biophys Res Commun. 2018;503(1):385–90.
Xiao Y, Ming X, Wu J. Hsa_circ_0002483 regulates miR-758-3p/MYC axis to promote acute myeloid leukemia progression. Hematol Oncol. 2021;39(2):243–53.
Chen F, Wang X, Fu S, Wang S, Fu Y, Liu Z, et al. Effect of the up-regulation of circular RNA Hsa_circ_0069767 derived from C-KIT on the biological behavior of multiple myeloma cells. Cancer Manag Res. 2020;12:11321–31.
Liu F, Wang YL, Wei JM, Huang ZD. Upregulation of circ_0000142 promotes multiple myeloma progression by adsorbing miR-610 and upregulating AKT3 expression. J Biochem. 2021;169(3):327–36.
Ding Y, Dong Y, Lu H, Luo X, Fu J, Xiu B, et al. Circular RNA profile of acute myeloid leukaemia indicates circular RNA annexin A2 as a potential biomarker and therapeutic target for acute myeloid leukaemia. Am J Transl Res. 2020;12(5):1683–99.
Tian FQ, Chen ZR, Zhu W, Tang MQ, Li JH, Zhang XC, et al. Inhibition of hsa_circ_0003489 shifts balance from autophagy to apoptosis and sensitizes multiple myeloma cells to bortezomib via miR-874-3p/HDAC1 axis. J Gene Med. 2021;24:e3329.
Wu Z, Sun H, Liu W, Zhu H, Fu J, Yang C, et al. Circ-RPL15: a plasma circular RNA as novel oncogenic driver to promote progression of chronic lymphocytic leukemia. Leukemia. 2020;34(3):919–23.
Cheng Y, Su Y, Wang S, Liu Y, Jin L, Wan Q, et al. Identification of circRNA-lncRNA-miRNA-mRNA competitive endogenous RNA network as novel prognostic markers for acute myeloid leukemia. Genes (Basel). 2020;11(8):868.
Wu W, Wu Z, Xia Y, Qin S, Li Y, Wu J, et al. Downregulation of circ_0132266 in chronic lymphocytic leukemia promoted cell viability through miR-337-3p/PML axis. Aging (Albany NY). 2019;11(11):3561–73.
Ghetti M, Vannini I, Storlazzi CT, Martinelli G, Simonetti G. Linear and circular PVT1 in hematological malignancies and immune response: two faces of the same coin. Mol Cancer. 2020;19(1):69.
Hu J, Han Q, Gu Y, Ma J, McGrath M, Qiao F, et al. Circular RNA PVT1 expression and its roles in acute lymphoblastic leukemia. Epigenomics. 2018;10(6):723–32.
Zhu Y, Ma X, Zhang H, Wu Y, Kang M, Fang Y, et al. Mechanism of circADD2 as ceRNA in childhood acute lymphoblastic leukemia. Front Cell Dev Biol. 2021;9:639910.
Feng XQ, Nie SM, Huang JX, Li TL, Zhou JJ, Wang W, et al. Circular RNA circHIPK3 serves as a prognostic marker to promote chronic myeloid leukemia progression. Neoplasma. 2020;67(1):171–7.
Liu W, Lei L, Liu X, Ye S. CircRNA_OTUD7A upregulates FOXP1 expression to facilitate the progression of diffuse large B-cell lymphoma via acting as a sponge of miR-431-5p. Genes Genomics. 2021;43(6):653–67.
Shi Y, Ding D, Qu R, Tang Y, Hao S. Non-coding RNAs in diffuse large B-cell lymphoma. Onco Targets Ther. 2020;13:12097–112.
Ding J, Zhang X, Xue J, Fang L, Ban C, Song B, et al. CircNPM1 strengthens Adriamycin resistance in acute myeloid leukemia by mediating the miR-345-5p/FZD5 pathway. Cent Eur J Immunol. 2021;46(2):162–82. https://doi.org/10.5114/ceji.2021.108175 Epub 2021 Aug 6.
Liu J, Du F, Chen C, Li D, Chen Y, Xiao X, et al. CircRNA ITCH increases bortezomib sensitivity through regulating the miR-615-3p/PRKCD axis in multiple myeloma. Life Sci. 2020;262:118506. https://doi.org/10.1016/j.lfs.2020.118506 Epub 2020 Oct 5.
Che H, Ding H, Jia X. circ_0080145 Enhances Imatinib Resistance of Chronic Myeloid Leukemia by Regulating miR-326/PPFIA1 Axis. Cancer Biother Radiopharm. 2020. https://doi.org/10.1089/cbr.2020.3600 Epub ahead of print.
Kristensen LS, Hansen TB, Venø MT, Kjems J. Circular RNAs in cancer: opportunities and challenges in the field. Oncogene. 2018;37(5):555–65.
Wu K, Liao X, Gong Y, He J, Zhou JK, Tan S, et al. Circular RNA F-circSR derived from SLC34A2-ROS1 fusion gene promotes cell migration in non-small cell lung cancer. Mol Cancer. 2019;18(1):98.
Guarnerio J, Bezzi M, Jeong JC, Paffenholz SV, Berry K, Naldini MM, et al. Oncogenic Circular Rnas Arise From Chromosomal Translocations. Cancer Discov. 2016;6(6):OF20. https://doi.org/10.1158/2159-8290.CD-RW2016-068.
Huang W, Fang K, Chen TQ, Zeng ZC, Sun YM, Han C, et al. circRNA circAF4 functions as an oncogene to regulate MLL-AF4 fusion protein expression and inhibit MLL leukemia progression. J Hematol Oncol. 2019;12(1):103.
Pan Y, Lou J, Wang H, An N, Chen H, Zhang Q, et al. CircBA9.3 supports the survival of leukaemic cells by up-regulating c-ABL1 or BCR-ABL1 protein levels. Blood Cells Mol Dis. 2018;73:38–44. https://doi.org/10.1016/j.bcmd.2018.09.002 Epub 2018 Sep 14.
Jadhav SP, Kumari N, Ng L, Tan PF, Yeo-Teh NSL, Goh Y, et al. circASXL1-1 regulates BAP1 deubiquitinase activity in leukemia. Haematologica. 2020;105(7):e343–8.
Jiao J, Duan C, Zheng J, Li D, Li C, Wang Z, et al. Development of a two-in-one integrated assay for the analysis of circRNA-microRNA interactions. Biosens Bioelectron. 2021;178:113032.
Wei Y, Zhang Y, Meng Q, Cui L, Xu C. Hypoxia-induced circular RNA has_circRNA_403658 promotes bladder cancer cell growth through activation of LDHA. Am J Transl Res. 2019;11(11):6838–49.
Li S, Li X, Xue W, Zhang L, Yang LZ, Cao SM, et al. Screening for functional circular RNAs using the CRISPR-Cas13 system. Nat Methods. 2021;18(1):51–9.
Li T, Shao Y, Fu L, Xie Y, Zhu L, Sun W, et al. Plasma circular RNA profiling of patients with gastric cancer and their droplet digital RT-PCR detection. J Mol Med (Berl). 2018;96(1):85–96.
Gao X, Xia X, Li F, Zhang M, Zhou H, Wu X, et al. Circular RNA-encoded oncogenic E-cadherin variant promotes glioblastoma tumorigenicity through activation of EGFR-STAT3 signalling. Nat Cell Biol. 2021;23(3):278–91.
Das A, Rout PK, Gorospe M, Panda AC. Rolling circle cDNA synthesis uncovers circular RNA splice variants. Int J Mol Sci. 2019;20(16):3988.
Boss M, Arenz C. A fast and easy method for specific detection of circular RNA by rolling-circle amplification. Chembiochem. 2020;21(6):793–6.
Li S, Teng S, Xu J, Su G, Zhang Y, Zhao J, et al. Microarray is an efficient tool for circRNA profiling. Brief Bioinform. 2019;20(4):1420–33. https://doi.org/10.1093/bib/bby006.
Glažar P, Papavasileiou P, Rajewsky N. circBase: a database for circular RNAs. RNA. 2014;20(11):1666–70. https://doi.org/10.1261/rna.043687.113 Epub 2014 Sep 18.
Dong R, Ma XK, Li GW, Yang L. CIRCpedia v2: An Updated Database for Comprehensive Circular RNA Annotation and Expression Comparison. Genomics Proteomics Bioinformatics. 2018;16(4):226–33. https://doi.org/10.1016/j.gpb.2018.08.001 Epub 2018 Aug 29.
Li JH, Liu S, Zhou H, Qu LH, Yang JH. starBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014;42(Database issue):D92–7. https://doi.org/10.1093/nar/gkt1248 Epub 2013 Dec 1.
Mokrejs M, Masek T, Vopálensky V, Hlubucek P, Delbos P, Pospísek M. IRESite--a tool for the examination of viral and cellular internal ribosome entry sites. Nucleic Acids Res. 2010;38(Database issue):D131–6. https://doi.org/10.1093/nar/gkp981 Epub 2009 Nov 16.
Dudekula DB, Panda AC, Grammatikakis I, De S, Abdelmohsen K, Gorospe M. CircInteractome: a web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016;13(1):34–42. https://doi.org/10.1080/15476286.2015.1128065.
Xia S, Feng J, Lei L, Hu J, Xia L, Wang J, et al. Comprehensive characterization of tissue-specific circular RNAs in the human and mouse genomes. Brief Bioinform. 2017;18(6):984–92. https://doi.org/10.1093/bib/bbw081.
Mody RJ, Wu YM, Lonigro RJ, Cao X, Roychowdhury S, Vats P, et al. Integrative clinical sequencing in the Management of Refractory or relapsed Cancer in youth. JAMA. 2015;314(9):913–25. https://doi.org/10.1001/jama.2015.10080.
Tang Z, Li X, Zhao J, Qian F, Feng C, Li Y, et al. TRCirc: a resource for transcriptional regulation information of circRNAs. Brief Bioinform. 2019;20(6):2327–33. https://doi.org/10.1093/bib/bby083.
Acknowledgements
Not applicable.
Funding
This study was supported by research grants from the National Natural Science Foundation of China to Sixi Wei (Grant No.81660027, No.81960031), the Science and Technology Project of Guiyang City to Sixi Wei (Grant NO. 20161001021), the Science and Technology Project of Guizhou Province to Sixi Wei (Grant NO. 20185779–70) as well as the Research Funds for Guizhou Provincial Innovative Talents Team for 2019 (Grant No. 5610).
Author information
Authors and Affiliations
Contributions
All authors revised and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
I, Fahua Deng, give my consent for information about myself to be published in Biomarker Research, manuscript number f8d468fc-5e10-450b-9581-0c1055293656 and corresponding author Sixi Wei.
I understand that the information will be published without my name attached, but that full anonymity cannot be guaranteed. I understand that the text and any pictures or videos published in the article will be freely available on the internet and may be seen by the general public. The pictures, videos and text may also appear on other websites or in print, be translated into other languages, or be used for commercial purposes. I have been offered the opportunity to read the manuscript.
Signing this consent form does not remove my rights to privacy.
Name: Fahua Deng.
Date:
Signed:
Author name: Fahua Deng.
Date:
Signed:
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Deng, F., Zhang, C., Lu, T. et al. Roles of circRNAs in hematological malignancies. Biomark Res 10, 50 (2022). https://doi.org/10.1186/s40364-022-00392-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40364-022-00392-2