
Abstract
Background
We report compelling evidence suggesting a predation event of a pop-up satellite archival tagged anadromous Dolly Varden (Salvelinus malma) by a marine mammal during summer in the Beaufort Sea based on abrupt changes in temperature and vertical movements. This observation provides insight on predator avoidance behaviour by Dolly Varden and the predator’s feeding frequency while the tag was ingested. Based on published distribution and ecology information, we presumed the predator was a beluga whale (Delphinapterus leucas). Supplemental satellite telemetry data from previously tagged Dolly Varden and beluga whales were used to determine the extent of spatial and vertical overlap between species in the area where predation occurred.
Results
Prior to the predation event, depths and temperatures occupied by the tagged Dolly Varden averaged 1.1 m and 3.1 °C, respectively. On July 7, 2020, depths remained shallow apart from a sudden dive to 12.5 m (16:45 UTC) followed by a precipitous increase in temperature from 4.4 to 27.1 °C (16:52 UTC) suggesting predation by an endotherm. Subsequent readings indicated the endotherm had a resting stomach temperature of 36.1 °C. Including the predation event, eight separate feeding events were inferred during the 20-h period the tag was ingested (before presumed regurgitation) based on subsequent declines in stomach temperatures (mean decline to 31.1 °C) that took an average of 24.1 min to return to resting temperature. The predator occupied mainly shallow depths (mean = 2.3 m), overlapping with tagged belugas that spent 76.9% of their time occupying waters ≤ 2.5 m when frequenting the area occupied by tagged Dolly Varden in the Canadian Beaufort Sea in July. Back-calculation based on tag drift and mean displacement by tagged belugas indicated the predation likely occurred west of the Mackenzie Delta.
Conclusion
Our findings provide new information on both anti-predator behaviour by, and marine predators of, Dolly Varden in the Beaufort Sea. We provide the first estimate of feeding frequency and stomach temperature recovery in a presumed wild beluga, and evidence for shallow foraging behaviour by belugas. Elucidating the likely predator and exploring the extent of overlap between Dolly Varden and beluga whales contributes towards knowledge on the trophic interactions in the Beaufort Sea.
Similar content being viewed by others
Background
Biologging and telemetry technologies have enabled the characterization of animal movement, behaviour, habitat use, and trophic ecology [1,2,3]. The application of these technologies in polar marine ecosystems are especially valuable given the challenges of tracking animal movements and their interspecific interactions in regions where direct observations can be limited. The Arctic is experiencing significant rates of change in the marine environment [4], underscoring the need to explore multiple approaches for understanding energy flow and trophic interactions to improve the management and conservation of Arctic marine ecosystems. Elucidating food web relationships and the energetic transfer within ecosystems requires identifying and defining predator–prey relationships. Characterizing these relationships can inform processes that drive trophic cascades, population dynamics, and species distribution [5, 6].
Defining predation using biologging and telemetry instrumentation has informed trophic interactions over space and time that are difficult to obtain with more traditional dietary methods, such as stomach content or biochemical tracer analyses [7, 8]. Advancements in telemetry that enable the collection of acceleration force, light level, pressure (depth), and temperature data can be used to qualitatively or quantitatively infer the predation of an externally or internally tagged individual based on changes in movement behaviour, light levels, sudden increase in temperature (i.e. consumed by endotherm), and temporal dissociation between time-series depth and temperature profiles while the tag remains in the digestive tract of the predator (i.e. consumed by ectotherm) [3, 9,10,11]. Predation of archival tagged fishes by marine mammals, such as whales, provides novel information on sources of natural mortality, predator species, and animal behaviours that are not commonly documented (see [11,12,13]).
Here we report on compelling evidence to suggest a predation event of an archival tagged anadromous adult Dolly Varden (Salvelinus malma) by a marine mammal in the eastern Beaufort Sea based on abrupt changes in temperature and vertical movement patterns. Dolly Varden is an iteroparous salmonid that can undertake seasonal migrations between freshwater (spawning and overwintering) and marine (feeding) habitats, and are important for Indigenous subsistence fisheries in the Alaskan and western Canadian Arctic [14, 15]. Piscivorous marine mammal predators that occur in this region are beluga whales (Delphinapterus leucas) [16, 17], ringed seals (Phoca hispida) [18], bearded seals (Erignathus barbatus) [19], and on rare occasions killer whales (Orcinus orca) [20]. Harbour porpoise (Phocoena phocoena) [21] and walrus (Odobenus rosmarus) [22] have been observed in exceptionally rare occasions.
We sought to identify the most likely predator of the tagged Dolly Varden using published spatial distribution and prevalence, internal body temperatures, diet, feeding mechanisms, and diving behaviours of known marine mammals in the eastern Beaufort Sea region. Our objectives were to use archived data prior to and during the predation event to (1) demonstrate predator avoidance behaviour by Dolly Varden and (2) characterize predator feeding behaviour, respectively. After concluding the predator was most likely a beluga whale, we used available satellite telemetry data from belugas and Dolly Varden inhabiting the eastern Beaufort Sea to (3) confirm whether the predator exhibited similar vertical behaviour to tagged belugas and (4) estimate the area where predation occurred and the extent of spatial and vertical overlap between these species in July. Elucidating the likely predator and exploring the extent of overlap between Dolly Varden and beluga whales contributes towards knowledge on habitat use and trophic interactions in the eastern Beaufort Sea. This is particularly relevant given the listing of Dolly Varden as ‘Special Concern’ under Species at Risk legislation in Canada, the nearby marine protected area (Tarium Niryutait Marine Protected Area) in place to conserve beluga whales and their prey, and because of the paucity of documentation of anadromous salmonid predation by marine mammals in the Canadian Arctic.
Materials and methods
Dolly Varden capture and tagging
Anadromous Dolly Varden were captured as part of ongoing fisheries-independent assessment studies in mid-September 2019 in the Firth River (68.64313° N; 140.98715° W), Yukon Territory, Canada (Fig. 1). A long seine net was deployed in shallow pools near perennial groundwater springs occupied by Dolly Varden using the techniques described by Sandstrom et al. [23]. The Dolly Varden of interest was a current-year spawning male with a fork length of 723 mm (estimated weight = ~ 3.75 kg based on length–weight regression of Dolly Varden captured along the Canadian Beaufort Sea coast; Gallagher et al. unpublished observations) that had a pop-up satellite archival tag (PSAT) (miniPAT; Wildlife Computers, USA) (https://wildlifecomputers.com/our-tags/pop-up-satellite-tags-fish/minipat/) attached using the methods described by Courtney et al. [24]. The tag was intended to remain tethered to the fish for 10 months until the scheduled release date of July 15, 2020. Depth (± 0.5 m) and temperature data (± 0.1 °C) were sampled at a 1 s interval and programmed to transmit as time-series subsampled at a 7.5 min interval (with associated TRange and DRange error parameters, respectively, which are a function of the range of temperatures and depths, respectively, encoded in a 6 h message). Depth and temperature data recorded by the tags are measured within 25 ms of each other, and the time constant of the thermistor is < 1 s (Matthew Rutishauser, Wildlife Computers, personal communication). Therefore, the tag precisely records and transmits data on the external environment encountered at a given time. Tags also transmitted the maximum and minimum depth and temperature (sampled at 1 s intervals) for each 6 h summary period while reporting locations in the Beaufort Sea were calculated through transmissions to ARGOS satellites after pop-up. Light level logging on the PSAT was disabled given light-based geolocation of fishes at high latitudes is currently unreliable [25]. The mean ± SD depth and temperature occupied by the tagged Dolly Varden in the Beaufort Sea prior to predation were calculated using raw time-series of data, while ranges were determined using SeriesRange data logged by the tag. Additionally, PSAT data on the marine habitat use by 13 Dolly Varden (same deployment approach and tagging method as the fish from Firth River) from two other Canadian populations east of the Firth River (Big Fish River: 68.3025° N, 136.3476° W; and Rat River: 67.7558° N; 136.2935° W) collected between 2015 and 2018 [26] were used to delineate the geographic extent of the eastern Beaufort Sea that could potentially be inhabited by Dolly Varden during summer based on the locations where tags reported to satellites in mid-July. Furthermore, PSAT data from these fishes were used to help characterize depths typically occupied by Dolly Varden in the Beaufort Sea.

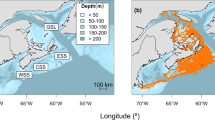
Map of study area in the eastern Beaufort Sea illustrating the location where an anadromous Dolly Varden was satellite archival tagged in September, 2019, the area where the tagged fish was presumably eaten by a beluga whale on July 7, 2020, the location where the tag was presumably regurgitated by the beluga whale, and the location where the tag reported to satellites on July 15, 2020. Locations are shown where beluga whales were satellite archival tagged in summer 2018 and 2019, and whose depth and spatial habitat use were assessed in the Dolly Varden spatial polygon (delineated based on previously satellite archival tagged Dolly Varden from Canadian populations (Big Fish R. and Rat R.) that reported in mid-July in the Beaufort Sea (2015–2018)). Sea surface temperatures (SST) (average between July 3 and 10, 2020) and sea-ice conditions (July 8, 2020) of the Beaufort Sea near the approximate date of predation are shown along with the start locations of the detailed dive profiles illustrated in Fig. 4 from tagged belugas within the spatial polygon
Beluga whale capture and tagging
Adult male belugas were tagged with satellite-linked time-depth recorders (TDRs) from Hendrickson Island (Fig. 1; 69.48° N; 133.61° W), Northwest Territories, Canada in July of 2018 and 2019 [27]. Tags were mounted on the dorsal ridge of belugas that were live captured using a net encirclement method [28] and remotely deployed on free-ranging belugas. Tags included SPLASH10-F-238, SPLASH10-F-321, MiniPAT (Wildlife Computers Ltd., Redmond, WA), and CTD-SRDL tags (Sea Mammal Research Unit—SMRU, University of St. Andrews). Tags from 17 belugas provided Fastloc® GPS and/or Argos location data, and 13 of the tags transmitted TDR data as time-series of depths at 75 s intervals.
Inferring the predator
We searched published literature using Web of Science (Clarivate Analytics (USA)) to deduce the most likely predator based on physiological and behavioural data provided by the ingested tag along with other information on diet, feeding ecology, depth occupancy, and prevalence in the eastern Beaufort Sea. Harbour porpoise and walrus were omitted given the exceptionally rare instances both species have been observed in the eastern Beaufort Sea, although we retained killer whales given their increasing prevalence in the western Arctic [29]. We compared resting stomach/body temperature among the four candidate predators (beluga whale, killer whale, ringed seal, and bearded seal). Furthermore, we searched for records of Dolly Varden or other salmonid predation among each predator, as well as their diet preference and feeding kinematics. We examined whether the candidate predators are known to forage at depths typically occupied by Dolly Varden (upper 2.5 m; [26]), and if they make dives to the maximum depths recorded by the ingested tag (48 m). The amassed information was placed in a summary table and we deduced the likely species based on conformity to these criteria.
Estimating the location of the predation event
Estimating the location of the predation event required the consideration of two components: (1) tag drift following expulsion from the predator (removal of the tag from the predator either by regurgitation or defecation) and (2) the distance travelled by the predator for the time between ingestion (initial consumption of the tag by the predator) and expulsion (20 h). Based on depth and temperature readings, the tag was expelled by the predator roughly 8.3 days before data transmissions to overhead Argos satellites on the programmed reporting date (July 15, 2020). Therefore, to infer where the tag was expelled by the predator, the estimated distance travelled by the tag while drifting on the ocean surface following expulsion (i.e. premature tag drift) was subtracted from the location where it reported to satellites. Furthermore, the estimated area of predation could only include depth contours ≥ 12.5 m given this was the final depth recorded by the Dolly Varden immediately prior to predation. The vector (bearing and speed) of tag drift was estimated from the first 48 h of tag transmission. While this analysis assumed that tag-reporting drift was similar to premature tag drift and did not provide an accurate location, it did provide an approximation of the last known location of the predator.
Assuming the predator was a beluga (see ‘predator identification’ in ‘Results’), we estimated the average distance travelled by tagged beluga whales in a 20 h timespan (i.e. the duration over which the tag remained in the gastrointestinal tract of the predator). Fastloc® GPS and Argos location data from tagged belugas were filtered and correlated random walk models were fit, with locations predicted at 15 min intervals [27]. To identify comparable beluga displacement distance in an area known to be occupied by Dolly Varden, we analysed beluga telemetry data from 29th June through July in 2018 and 2019 that were in the geographic vicinity occupied in previous years by PSAT tagged Dolly Varden in the Canadian Beaufort Sea (bounded by 70.7550° N, 143.5860° W, 132.8793° W, and the shoreline [26], henceforth referred as the ‘spatial polygon’) (Fig. 1). The ‘geosphere’ v1.5-10 [30] package in R v3.6.0 [31], with the ‘distVincenityEllipsoid’ function, was used to calculate the distance between all pairs of beluga locations in the spatial polygon at times t and t + 20 h. The average displacement distance was calculated and subtracted from the approximate location where the tag was expelled to estimate the general area where predation occurred.
Beluga dive behaviour in the region occupied by Dolly Varden
To identify the proportion of time tagged belugas whales spent at depths typically occupied by Dolly Varden inside and outside the spatial polygon, we calculated the proportion of beluga TDR data that recorded depths ≤ 2.5 m in the spatial polygon during the entire month of July in 2018 and 2019, and compared it to the TDR data from outside the spatial polygon during July. To allow for error in the transmitted depth data, we subtracted the depth resolution from the depth; hence, the calculations here give maximal estimates of the time spent in the surface 2.5 m. A paired t-test was used to calculate whether the proportion of time by tagged belugas occupying the surface 2.5 m differed inside and outside the spatial polygon. Dive profiles from tagged belugas occupying the spatial polygon during July were plotted to provide higher-resolution examples of dive behaviour of belugas that reached depths close to those reported by the ingested tag (maximum 48 m).
Characterizing feeding frequency
Declines in marine mammal stomach temperature can indicate ingestion of prey or water [32]; however, it has been suggested that ingestion of water may be unlikely in Arctic cetaceans as they can obtain sufficient water from prey, and heating ingested water would be associated with high energetic costs [33]. Therefore, we consider any recorded drop in temperature while the tag was ingested to indicate a subsequent feeding event by the predator. As the temperature data were collected at 7.5 min intervals, we note that the declines and stabilizations in temperature could have occurred within 7 min 29 s prior to the times reported; therefore, the durations of stomach temperature recovery have an error of ± 7 min 29 s.
Results
Dolly Varden vertical behaviour and predation event
After overwintering in the Firth River, the tagged Dolly Varden entered the Beaufort Sea on approximately June 22, 2020 based on a dramatic drop in water temperature, suggesting a transition from freshwater to marine environments (see [24]). Depth and temperature (mean ± SD) occupied while feeding at sea was 1.1 ± 1.3 m (range = 0.5–20 m) and 3.1 ± 1.8 °C (range = − 1.3 to 6.7 °C) consistent with findings from other PSAT tagged Dolly Varden in the Beaufort Sea [26, 34]. Depths occupied by the tagged Dolly Varden during the several hours prior to predation ranged between 0.5 and 5 m (mean = 1.36 m) apart from the last depth recorded immediately prior to ingestion (12.5 m at 16:45 UTC, Fig. 2b). On July 7, 2020 (16:52 UTC) the temperature increased precipitously from 4.4 to 27.1 °C and increased to 34 °C approximately 30 min later, indicating ingestion by an endotherm.

a Temperature and b depth recorded by a pop-up satellite archival tag attached to a Dolly Varden prior to, during, and after predation by a presumed beluga whale in the Beaufort Sea. Numbers on top of panels refer to timing of temperature changes, specified in Table 1. Note that temperature and depth error are based on the TRange and DRange, respectively, values reported in the time-series files, which are a function of the range of temperature and depths encoded in a 6 h message
While inside the predator the tag recorded an overall maximum temperature of 36.2 °C with maximum temperatures of 36.1 °C recorded during 75% of the 6 h summary periods. Stomach temperature stabilized at 36.1 °C after initial tag ingestion and subsequent temperature drops, and was therefore considered to be the predator’s resting stomach temperature. After the initial tag ingestion there were seven subsequent declines in temperature (Table 1, Fig. 2a). The mean stomach temperature following a decline was 31.1 °C (mean temperature decrease = 5.0 °C); however, these declines were highly variable with a maximum temperature decrease to 13.8 °C (drop of 22.3 °C, event 10, Table 1, Fig. 2a) and a minimum decrease to 35.7 °C (drop of only 0.4 °C, event 12, although the 7.5 min sample interval may have underestimated the magnitude of this decrease). The durations for stomach temperature recovery were also variable. After the initial tag ingestion event, it took 2.75 h for the predator's stomach temperature to stabilize (Table 1); however, subsequent temperature drops during this period (events 3 and 4, Fig. 2a) indicate that additional ingestion events took place during recovery. For the remaining temperature drops (events 6, 8, 10, 12, and 14, Table 1, Fig. 2a) the recovery duration was between 7.5 min and 37.5 min (± 7 min 29 s) (mean = 24.1 min). On July 8, 2020 (12:45 UTC) temperature immediately decreased to 4.7 °C (Fig. 2a) and depth became constant at 0.5 m (i.e. tag presumably floating at the surface), suggesting tag expulsion after a 20 h duration in the gastrointestinal tract of the predator (Fig. 2b).
Ultimately, it appears there were seven ingestion events during the period when the tag was in the predator’s stomach (eight, including the predation event of the tagged fish) (Table 1). The 7.5 min interval depth time-series data suggested that the predator was primarily making shallow dives (mean ± SD = 2.3 ± 2.1 m, Fig. 2b); however, maximum depths of 41, 48, and 45 m were reported during three of the 6 h summary periods (depth information was missing for one 6 h summary period); hence, the predator made deeper dives than the time-series data suggest (Fig. 2b) and we cannot be certain at which depths the later predation events occurred.
Predator identification: comparing prevalence, body temperature, dive behaviour, and feeding kinematics among Beaufort Sea marine mammals
Beluga whales, killer whales, ringed seals, and bearded seals are all known to occur along the nearshore (coastal) and offshore Beaufort shelf that are occupied by Dolly Varden (up to 69–152 km offshore; [26, 34]) in summer (Table 2), and this region has been identified as a hotspot for Arctic cetaceans and pinnipeds [35]. The most prevalent among the four predators in the eastern Beaufort Sea are beluga whales [36,37,38] and ringed seals [39, 40]. There have been extensive telemetry campaigns of belugas and ringed seals in the region relative to other potential predators. Bearded seals have been detected year-round along the shelf of the Alaskan Beaufort Sea by passive acoustic monitoring [41], although acoustic activity tends to decrease between June and July, which could be due to changes in behaviour or movement offshore following seasonal sea ice retreat [42]. Killer whales have only been observed occasionally in this region (31 documented observations between 1940 and 2009; [20]), although there are signs of increasing presence [29].
All four predator species considered here are known to dive to ≥ 48 m (Table 2). Beluga whales in this region are thought to principally undertake foraging during deeper dives [27, 43]. However, without direct observation or use of ancillary data, inferences as to dive function are based on occupancy time at depth or dive profile characteristics from tagged individuals, which may not always be representative of various behaviours [44, 45]. Elsewhere in their range, beluga whales have been observed feeding at the surface (e.g. [46]) and undertaking shallow foraging-type dives (square U-shaped, mean maximum depth = 13.35 m; [47]). Ringed seals in this region also tend to make fewer shallow than deep dives [48], although repetitive diving to < 25 m suggests shallower foraging also occurs [49]. Bearded seals make both shallow and deep dives; however, they principally feed on the benthos [42, 50, 51]. Both mammal-eating and fish-eating killer whales can undertake shallow foraging behaviour [52, 53].
While inferably in the stomach of a predator, the 36.1 °C resting stomach temperature was similar to documented body temperatures of the marine mammals known to occur in this region, including beluga whales (34.9–35.9 °C, Table 2) and killer whales (36.0–38.0 °C), yet lower than body temperatures of seals (Table 2). However, previous studies were based on few individuals or limited to certain age classes, and did not always provide the range of temperatures recorded. Therefore, temperature alone cannot be used to deduce candidate predators. Additionally, resting stomach temperatures may vary between individuals, as shown in a study that deployed stomach temperature pills (STPs) in narwhals [33], a close relative of belugas and the only other extant member of the Monodontidae (mean resting stomach temperature = 35.5 °C, range = 34.5–36.2 °C).
While diet information is limited for these marine mammal species, Dolly Varden has been detected in small amounts (e.g. < 1% of enumerated diet items) in the stomach contents of beluga whales from the nearby Alaskan Chukchi Sea and Bristol Bay [16, 54]. Closely related Arctic char (S. alpinus) has been observed in Beaufort Sea beluga stomachs [55] and similar-sized fish (range = 549–738 mm) have been documented in beluga stomachs [16]. Available published literature of ringed and bearded seal diets has not found Dolly Varden present (e.g. [19]). It is noted that a small number of Dolly Varden captured during annual sampling at spawning and overwintering habitats in fall among various rivers in Canada have injuries from predators (prevalence from 2015 to 2019 is < 2% for Dolly Varden > 700 mm; Gallagher unpublished observation) with some appearing consistent with attempted capture by presumed seals (e.g. elongated wound or healed scar from assumed claw and bite marks) [56] (Fig. 3). While they can both consume fish, ringed seals are characterized as ‘pierce feeders’ (use a combination of biting and suction to opportunistically catch prey) [57], and bearded seals are principally characterized as suction feeders [57] with a small gape (2.7 ± 0.85 cm; [58]). Furthermore, it is assumed that given their gape size, seals would be unlikely to completely ingest a whole salmonid [11] and likely avoid swallowing a pop-up satellite archival tag [59]. Together, the small gape size and lack of observed predation on larger prey items, such as Dolly Varden, suggests neither phocid species was the predator of interest in this study.

Examples of anadromous Dolly Varden (approximate fork length between 450 and 550 mm) captured while spawning in freshwater that had an injury along the ventral surface between pelvic and anal fins presumably as a result of a predation attempt by a ringed seal in the Beaufort Sea
Belugas consume their prey whole using either ram or suction feeding, during both of which they have exhibited a large gape (10.02 ± 0.87 cm and 6.3 ± 0.4 cm, respectively; [60]) during experimental trials where they were fed prey smaller than the Dolly Varden consumed in this study (mean length of herring = 242 ± 28 mm). Beluga gape is presumed to increase beyond this size given they are known to feed on large salmonids over 549 mm in length based on stomach contents from the Cook Inlet, Alaska [16]. Killer whales can consume whole fish [61], but have also been observed tearing large salmonids apart at the surface [62]. There have been unconfirmed suggestions that killer whales in the Pacific Arctic may consume fish [63]. However, evidence from group size [20], injuries to landed bowhead whales [64], and bowhead whale carcasses [65] provide strong support for killer whales observed in the Beaufort Sea being mammal-eating transients. Given their low occurrence and likely specialization as marine mammal predators in the Beaufort Sea, a killer whale appears less likely as the predator of the tagged Dolly Varden. However, killer whales meet the diving and feeding criteria (Table 2) and cannot be discounted based on the lack of knowledge of this predator in the study region. In contrast, the extensive use of the area by belugas, feasibility of feeding kinematics, and past observations of Dolly Varden and similar shaped/size fish in their diet supports the high likelihood of the beluga whale being the predator in question, and we consider this interaction further to better characterize the predation event.
Location and diving behaviour of tagged belugas
A total of 17 tagged belugas recorded 83.9 days of data while in the spatial polygon delineating documented Dolly Varden marine habitat in July (Table 3). Beluga whales tagged in 2018 and 2019 recorded an average of 4.9 days (SD = 2.7 days) in the polygon during July (Table 3; Fig. 1). Belugas recorded a higher proportion of time in the upper 2.5 m of the water column while in the polygon relative to all other regions occupied during July (mean ± SD = 76.9% ± 14.3% vs. 42.0% ± 11.2%, p < 0.05, Table 3). Beluga dive profiles within the polygon revealed that dives targeted the mid-water column as well as the seafloor (Fig. 4a, b). Dives to 48 m were often short (6.25 min, Fig. 4c), which may have been missed given depths of the consumed tag were collecting at a 7.5 min interval.

Vertical behaviour of two representative tagged beluga whales over 20 h periods with depths recorded at 75 s intervals within an area (see ‘spatial polygon’ in Fig. 1) used by Canadian anadromous Dolly Varden near the estimated location of the ingested tagged Dolly Varden: a RD2019#18 on 16th July 2019 between 69.06374° N 137.0342° W and 69.5230° N 137.5928° W, and b RD2019#20 on 17th July 2019 between 69.83131° N 137.0729° W and 70.15336° N 137.3574° W, with the arrow denoting the time period (c) when two dives to 48 m completed in 8.75 min and 6.25 min, respectively. Seafloor depth estimated from correlated random walk-modelled locations and the International Bathymetric Chart of the Arctic Ocean (IBCAO) version 3.0 [101], which has a spatial resolution of 500 m
Location of predation event
Based on estimated tag drift (i.e. tag floating on the surface prior to transmitting first location), the estimated location of the predation event was approximately 390 km southeast of the reporting location in the vicinity of Shingle Point, Yukon, near the Mackenzie Delta (Fig. 1). The grand mean displacement distance over a 20 h period for belugas that were detected ≥ 3 days in this region (i.e. spatial polygon) was 46.7 km (Table 3). The depth of 12.5 m reported by the Dolly Varden immediately prior to ingestion also limits the location of the predation event to areas with a seafloor depth ≥ 12.5 m.
This estimated area of predation is corroborated by pop-up location of other satellite tagged Dolly Varden in Canada [26]. Additionally, tag-reported seasurface (< 0.5 m) temperature of the day prior to inferred predation was ~ 5 °C. Satellite-derived sea surface temperature estimates for July (MODIS 8-day average daytime 4 km SST V2019.0) (accessed from https://podaac-opendap.jpl.nasa.gov/opendap/allData/modis/L3/aqua/11um/v2019.0/4km/8day/) provided additional support for the estimated location of predation, and further suggest that the true location of the predation event is likely within 200 km of the Mackenzie Delta and south of the 5 °C isoline (Fig. 1), similar to our back-calculated estimate. Furthermore, the likelihood of this region being the area of predation is supported by the concurrence between the bathymetry (seafloor depths < 50 m) and the maximum depths reached by the predator while the tag was ingested (41–48 m, Fig. 2b).
Discussion
After examining the published literature on predator species prevalence in the eastern Beaufort Sea, body temperature data, vertical and foraging behaviour, diet records, and feeding kinematics, we posit that the predator of the tagged Dolly Varden was a beluga whale from the Eastern Beaufort Sea (EBS) population. This is the first documentation of a trophic interaction between Dolly Varden and presumed beluga whale using biologging technology. EBS belugas not only inhabit the Mackenzie Shelf in the southeastern Beaufort Sea west of the Mackenzie Delta in early July [37, 38, 66], where predation likely occurred, but tagged individuals also demonstrated similar diving behaviours and vertical distribution to that of the predator of the tagged Dolly Varden. Interestingly, the area where the presumed predation event occurred (i.e. near Shingle Point) is a common feeding area for Dolly Varden [67].
Predator avoidance behaviour
Behaviours to reduce the risk of mortality can be influenced, among others, by predator density and perceived risk in a particular habitat, where a prey’s responses to a predator can result in energetic or reproductive costs that influence fitness [68, 69]. We hypothesize the beluga whale prompted the tagged Dolly Varden to dive to greater depths (12.5 m) very quickly in an attempt to escape. Dive depths > 12 m are rare for Dolly Varden in the Beaufort Sea, as these depths accounted for < 0.001% of observations in Courtney et al. [34]. Performing rapid deep dives to avoid predators has been described in salmonids from the genus Oncorhynchus [70], although our study is the first description of this behaviour for adult Dolly Varden or any anadromous salmonid in the Arctic prior to ingestion by a marine predator. Presumably, other behaviours, such as evasive manoeuvering and increased swimming speeds, that have been documented for Oncorhynchus spp. [70] would have also been performed by the tagged Dolly Varden. The 7.5 min sample interval of the PSAT time-series data in this study limits fine-scale and detailed examination of the vertical manoeuvers performed by the Dolly Varden immediately prior to predation. While deep-diving behaviour by Dolly Varden may not be solely associated with predator avoidance (e.g. it can also be related to olfactory orientation, bioenergetic gain, foraging opportunities; see summary for anadromous S. alpinus by Mulder et al. [71]), our observation is useful for re-examining datasets of archival tagged Dolly Varden to elucidate instances of potential predator avoidance or presumed tag loss. It is possible that the PSAT could have made the Dolly Varden more vulnerable to predation given the drag caused by the external tag that may have affected the manoeuverability of the fish and its ability to evade the predator [72]. Furthermore, the tag could have increased acoustic detectability of the fish given the additional surface area of the tag. Belugas have a highly directional sonar beam and can detect prey that are smaller than the Dolly Varden in our study at distances of up to 300 m [73]. Therefore, it is unlikely that the attached PSAT caused significant increase in the Dolly Varden’s detectability to belugas.
Insights into presumed beluga feeding
Measuring timing and periodicity of the feeding events of a species are relevant for elucidating trophic ecology and have been estimated using technology that measures stomach temperatures in various endothermic marine predators where it is assumed that rapid declines in temperature indicate ingestion of water or ectothermic prey [32, 74]. The predator in this study took a total of 2 h 45 min (± 14 min 58 s) (Table 1, Fig. 2) to recover its stomach temperature after ingesting an approximately 3.75 kg Dolly Varden, which is considerably longer than studies on captive seals for equivalent meal sizes [32]. This discrepancy may be explained somewhat by captive studies typically feeding marine mammals a larger quantity of smaller-sized fish rather than a single large item, and at a warmer temperature (mean = 17.5 ± 0.2 °C; [32]) than the Dolly Varden in the present study (3.1 ± 1.8 °C). This may have an effect on recovery time when meal sizes are large [75]. Previous studies have shown that time for stomach temperature to recover after an ingestion event is positively correlated with meal size [32, 75]. For example, Kuhn and Costa [32] demonstrated that captive northern elephant seals (Mirounga angustirostris) and California sea lions (Zalophus californianus) had mean recovery times of 87.2 min and 95.2 min, respectively, when fed 3 kg of fish compared to 61.7 min and 55.2 min, respectively, when fed 0.5 kg of fish.
Longer recovery to resting stomach temperature may also be explained by the ingestion events indicated by the two subsequent declines in temperature prior to stomach temperature stabilization (events 3 and 4, Table 1, Fig. 2a). After the initial ingestion, seven temperature declines were recorded, including the declines in temperature between the initial tag ingestion and the first temperature stabilization (events 3 and 4). The mean decline in temperature (5.0 °C) corresponds well with the magnitude of temperature declines assumed to represent ingestion events in narwhals [33]. Recovery times were longer in our study (mean = 24.1 min) than in narwhals (mean = 10 min), which could represent consumption of larger prey; however, it is important to note the low sampling interval of temperature measurements (7.5 min) in our study may under- or overestimate recovery time. If all declines in temperature over the 20 h period including the initial ingestion event (n = 8) represent ingestion of prey, this extrapolates to 9.6 predation events over a 24 h period, which is comparable to the 9.9 (SD = ± 4.2) predation events per 24 h estimated for narwhals [33]. However, this is likely an underestimate of the number of prey items ingested. A study on bottlenose dolphins (Tursiops truncatus) that used STPs recording temperature every 15 s found that temperature drops frequently occurred during stomach temperature recovery, indicating that multiple prey items are taken during feeding bouts [76, 77]. This could have been the case in event 14 (Table 1, Fig. 2), when rate of stomach temperature increase slowed during recovery. As the presumed beluga made at least three dives to between 41 and 48 m during the 20 h period, the tag was in its gastrointestinal tract, and experienced smaller drops in stomach temperature and shorter recovery times than after the initial ingestion event, it is probable that foraging on prey located at deeper depths and of smaller size than Dolly Varden may have occurred. For example, feeding on epibenthic invertebrates, such as decapods, represented 60% of the invertebrate stomach contents [16] and approximately 1% of a total diet of EBS belugas based on biotracers [17]. Other benthic fish consumed by belugas are also known to occur in the nearshore and shelf regions of the southeastern Beaufort Sea [55, 78]. This is supported by the tagged beluga dive profiles showing frequent benthic diving in this region (Fig. 4). Belugas principally occupy this region of the Beaufort Sea during July [36, 38]; thus, our estimation of feeding rate cannot be extrapolated to later in the summer or fall when belugas are distributed in deeper offshore habitats [36, 43].
Information on digestion timing by the presumed beluga whale, which is currently an unknown physiological parameter for this species in its natural habitat in the Arctic, would have been obtained if the ingested archival tag passed through the entire digestive system. The only study to date that has measured digestion rates in beluga whales was conducted by feeding captive animals a 6 g gelatine capsule of red dye [79], which took an average of 4.5 h to pass through the gastrointestinal tract (estimated lengths of seven times the total body length [80]). To our knowledge rates of digestion of solid food have not been measured in belugas. Heide-Jørgensen et al. [33] inserted STPs of two different sizes (30 g, 4 × 2 cm; and 60 g, 8 × 2 cm) into the stomachs of narwhals and the retention time within the stomach ranged from 10 h to 17.3 days. Heide-Jørgensen et al. [33] suggested that the shorter deployments may have been regurgitated, but the longest deployments (the longest of which was using the smaller STP size) may have passed through the gastrointestinal tract. The MiniPAT deployed in the present study was larger than the STPs in the narwhal study, with dimensions 12.4 × 3.8 cm, a weight of 60 g, and was attached by a 15 cm fishing line with two plastic plates (1 × 5 × 2 cm) [24]. Given the size of the tag and the duration the tag was inside the predator (20 h), it is most likely that the tag was regurgitated.
Potential importance of Dolly Varden as prey to beluga whales and interspecific spatial and vertical overlap
Long-term beluga harvest monitoring programmes in the Mackenzie estuarine coastal area [81] have provided samples to study diet through stomach contents and biotracers. Characterizing beluga diet in the eastern Beaufort Sea has been challenging given the findings of mainly empty stomachs in harvested whales (> 94% reported empty; [82]) along with the over- and under-representation of hard and soft tissues, respectively. The issues surrounding stomach content analysis have supported the use of biotracers such as fatty acids and stable isotopes to characterize diet (e.g. [83, 84]). Dietary assessments using biotracers require the biotracer data for both the predator and prey, and to date there has not been a beluga diet evaluation that included Dolly Varden fatty acids (e.g. [17, 55]).
In accounts of beluga stomach contents, the presence of Dolly Varden has been rare (< 1% in Bristol Bay, Alaska; [16]). Despite this, Dolly Varden are a lipid-rich source of prey (mean 12.3% lipid wet weight of anadromous non-spawning Dolly Varden > 600 mm estimated using a Distell Model 692 Fish Fatmeter (Distell Inc., West Lothian, Scotland); see [85]; Gallagher unpublished observation) that would be sought by opportunistic beluga whales given their preferred prey range from 5 to 8% lipid wet weight [17, 86]. Tagged belugas spent an average of 4.9 days (maximum = 10.15 days, Table 3) in the spatial polygon, and significantly more time at the depths typical of Dolly Varden in the spatial polygon relative to all other regions occupied during July, supporting the hypothesis that EBS belugas may be foraging at these depths while also presumably foraging during the deeper dives at other locations identified for this population [27, 43]. Furthermore, while belugas typically move offshore in late July, some individuals tagged in 2018 and 2019 returned to the Mackenzie estuary regions on one or more occasions (Loseto et al. unpublished observation). The sensory advantage that belugas have over their prey in the turbid waters of the estuary [73] and the presence of Dolly Varden at this time could indicate exploitation of a profitable food source in this region that is energetically favourable due to its shallower nature relative to deeper offshore prey [27].
Our findings underscore that Dolly Varden appear to be vulnerable to predation by belugas in the eastern Beaufort Sea as tagged beluga whales spent 76.9% of the time at depths primarily occupied by Dolly Varden when in proximity to the Yukon Territory coast/ Mackenzie Delta, which suggests an important degree of habitat overlap between species. However, further research is needed to elucidate how the overlap engenders risk of predation, including the role of environmental conditions (e.g. spatial distribution of sea ice, which is known to affect the spatial distribution of both species in the Beaufort Sea), particularly given that Dolly Varden has not been documented in stomach contents of EBS belugas. Further studies on beluga movements and the energetic costs of dive types (considering depth, duration, etc.) in relation to prey energetics (e.g. lipid content, size, schooling fish) are required to better explore predator–prey dynamics and the relationship between belugas and Dolly Varden.
Conclusions
We demonstrate a useful application of archival tag data to document behaviours expressed by both prey and predator in the Arctic as a result of predation. The findings of this interesting evidence of a trophic interaction in the eastern Beaufort Sea provides new information on both marine predators of and anti-predator behaviour by Dolly Varden in this region. Identifying sources of natural mortality for Dolly Varden in the marine environment is important for population assessment and management given the dearth of information on this topic. Furthermore, we provide the first estimate of feeding frequency and stomach temperature recovery duration in a wild beluga, and evidence for shallow foraging behaviour by belugas from the eastern Beaufort Sea population. Additional research is required to determine the importance of Dolly Varden to the diet of beluga whales in the Beaufort Sea, and to resolve the extent to which Dolly Varden are targeted by belugas that forage in the upper depths of the water column in the vicinity of the Alaska/ Yukon North Slope.
Availability of data and materials
The datasets used and/or analysed for this study are available from the corresponding and senior authors upon reasonable request.
References
Hussey NE, Kessel ST, Aarestrup K, Cooke SJ, Cowley PD, Fisk AT, et al. Aquatic animal telemetry: a panoramic window into the underwater world. Science. 2015;348:1255642. https://doi.org/10.1126/science.1255642.
Alfyard EA, Webber D, Del Papa J, Leadley T, Kessel ST, Colborne SF, et al. Evaluation of an acoustic telemetry transmitter designed to identify predation events. Methods Ecol Evol. 2017;8:1063–71. https://doi.org/10.1111/2041-210X.12726.
Tolentino ER, Howey RP, Howey LA, Jordan LKB, Grubbs RD, Brooks A, et al. Was my science project eaten? A novel approach to validate consumption of marine biologging instruments. Anim Biotelem. 2017;5:3. https://doi.org/10.1186/s40317-016-0117-4.
Landrum L, Holland MM. Extremes become routine in an emerging new Arctic. Nat Clim Change. 2020;10:1108–15. https://doi.org/10.1038/s41558-020-0892-z.
Schmitz OJ, Beckerman AP, O’Brien KM. Behaviorally mediated trophic cascades: effects of predation risk on food web interactions. Ecology. 1997;78:1388–99. https://doi.org/10.1890/0012-9658(1997)078[1388:BMTCEO]2.0.CO;2.
Brose U. Body-mass constraints on foraging behaviour determine population and food-web dynamics. Funct Ecol. 2010;24:28–34. https://doi.org/10.1111/j.1365-2435.2009.01618.x.
Goulet P, Guinet C, Swift R, Madsen PT, Johnson M. A miniature biomimetic sonar and movement tag to study the biotic environment and predator-prey interactions in aquatic animals. Deep Sea Res Part I Oceanog Res Pap. 2019;148:1–11. https://doi.org/10.1016/j.dsr.2019.04.007.
Hammerschlag N. Quantifying shark predation effects on prey: dietary data limitations and study approaches. Endang Species Res. 2019;38:147–51. https://doi.org/10.3354/esr00950.
Viviant M, Trites AW, Rosen DAS, Monestiez P, Guinet C. Prey capture attempts can be detected in Steller sea lions and other marine predators using accelerometers. Polar Biol. 2010;33:713–9. https://doi.org/10.1007/s00300-009-0750-y.
Cosgrove R, Arregui I, Arrizabalaga H, Goni N, Neilson JD. Predation of pop-up satellite archival tagged albacore (Thunnus alalunga). Fish Res. 2015;162:45–52. https://doi.org/10.1016/j.fishres.2014.09.003.
Strøm JF, Rikardsen AH, Campana SE, Righton D, Carr J, Aarestrup, et al. Ocean predation and mortality of adult Atlantic salmon. Sci Rep. 2019;9:7890. https://doi.org/10.1038/s41598-019-44041-5.
Wahlberg M, Westerberg H, Aarestrup K, Feunteun E, Gargan P, Righton D. Evidence of marine mammal predation of the European eel (Anguilla anguilla L.) on its marine migration. Deep Sea Res Part I Oceanog Res Pap. 2014;86:32–8. https://doi.org/10.1016/j.dsr.2014.01.003.
Seitz AC, Courtney MB, Evans MD, Manishin K. Pop-up satellite archival tags reveal evidence of intense predation on large immature Chinook salmon (Oncorhynchus tshawytscha) in the North Pacific Ocean. Can J Fish Aquat Sci. 2019;76:1608–15. https://doi.org/10.1139/cjfas-2018-0490.
Pedersen S, Linn A. Kaktovik 2000–2002 subsistence fishery harvest assessment. U.S. Fish and Wildlife Service, Office of Subsistence Management. 2005. 58 p. Final report for FIS Study 01-101, Fairbanks, Alaska.
DFO, FJMC, GRRB, PCA (Department of Fisheries and Oceans Canada, Fisheries Joint Management Committee, Gwich’in Renewable Resources Board, and Parks Canada Agency). Integrated fisheries management plan for Dolly Varden (Salvelinus malma malma) of the Gwich’in Settlement Area and Inuvialuit Settlement Region, Northwest Territories and Yukon North Slope. Volume 1: The Plan—2019 Update. 2019. 52 p. https://www.dfo-mpo.gc.ca/fisheries-peches/ifmp-gmp/dolly-varden/2019/index-eng.html.
Quakenbush LT, Suydam RS, Bryan AL, Lowry LF, Frost KJ, Mahoney BA. Diet of beluga whales, Delphinapterus leucas, in Alaska from stomach contents, March–November. Mar Fish Rev. 2015. https://doi.org/10.7755/MFR.77.1.7.
Choy ES, Giraldo C, Rosenberg B, Roth JD, Ehrman AD, Majewski A, et al. Variation in the diet of beluga whales in response to changes in prey availability: insights on changes in the Beaufort Sea ecosystem. Mar Ecol Prog Ser. 2020;647:195–210. https://doi.org/10.3354/meps13413.
Smith TG, Harwood LA. Observations of neonate ringed seals, Phoca hispida, after early break-up of the sea ice in Prince Albert Sound, N.T. Canada, spring 1998. Polar Biol. 2001;24:215–9. https://doi.org/10.1007/s003000000198.
Crawford J, Quakenbush LT, Citta JJ. A comparison of ringed and bearded seal diet, condition and productivity between historical (1975–1984) and recent (2003–2012) periods in the Alaskan Bering and Chukchi seas. Prog Oceang. 2015;136:133–50. https://doi.org/10.1016/j.pocean.2015.05.011.
Higdon JW, Byers T, Brown L, Ferguson S. Observations of killer whales (Orcinus orca) in the Canadian Beaufort Sea. Polar Rec. 2013;49:350. https://doi.org/10.1017/S0032247412000356.
van Bree PJH, Sergeant DE, Hoek W. A harbour porpoise, Phocoena phocoena (Linnaeus, 1758), from the Mackenzie River delta, Northwest Territories, Canada (Notes on Cetacea, Delphinoidea VII). Bearfortia. 1977;26:99–105.
Harington CR. Extralimital occurrences of walruses in the Canadian Arctic. J Mammal. 1966;47:506–13. https://doi.org/10.2307/1377692.
Sandstrom S, Harwood L, Howland K. Status of anadromous Dolly Varden charr (Salvelinus malma) of the Rat River, Northwest Territories, as assessed through mark-recapture and live-sampling at the spawning and overwintering site (1995–2007). Can Tech Rep Fish Aquat Sci. 2009. 68 p. Report No. 2842. http://waves-vagues.dfo-mpo.gc.ca/Library/338722.pdf.
Courtney MB, Scanlon BS, Rikardsen AH, Seitz AC. Marine behavior and dispersal of an important subsistence fish in Arctic Alaska, the Dolly Varden. Environ Biol Fish. 2016;99:209–22. https://doi.org/10.1007/s10641-015-0468-3.
Chittenden CM, Ådlandsvik B, Pedersen OP, Righton D, Rikardsen AH. Testing a model to track fish migrations in polar regions using pop-up satellite archival tags. Fish Oceanogr. 2013;22:1–13. https://doi.org/10.1111/fog.12000.
Gallagher CP, Courtney MB, Seitz AC, Lea EV, Howland KL. Ocean-entry timing and marine habitat-use of Canadian Dolly Varden: dispersal among conservation, hydrocarbon exploration, and shipping areas in the Beaufort Sea. Estuar Coast Shelf Sci. 2021. https://doi.org/10.1016/j.ecss.2021.107609.
Storrie L, Hussey NE, MacPhee SA, O’Corry-Crowe G, Iacozza J, Barber DG, Nunes A, Loseto LL. Habitat-dependent dive characteristics of male beluga whales from the Eastern Beaufort Sea population tracked over an annual cycle. Front Mar Sci. In Press. https://doi.org/10.3389/fmars.2021.715412.
Orr JR, Joe R, Evic D. Capturing and handling of white whales (Delphinapterus leucas) in the Canadian Arctic for instrumentation and release. Arctic. 2001;54:299–304. https://doi.org/10.14430/arctic789.
Stafford KM. Increasing detections of killer whales (Orcinus orca), in the Pacific Arctic. Mar Mamm Sci. 2018;35:696–706. https://doi.org/10.1111/mms.12551.
Hijmans RJ, Williams E, Vennes C. Geosphere: spherical trigonometry. R package version 1.5–10. 2019. https://cran.r-project.org/web/packages/geosphere/index.html.
R Core Team. R: a language and environment for statistical computing. Vienna, Austria; 2019. https://www.R-project.org/.
Kuhn CE, Costa DP. Identifying and quantifying prey consumption using stomach temperature change in pinnipeds. J Exp Biol. 2006;209:4524–32. https://doi.org/10.1242/jeb.02530.
Heide-Jørgensen MP, Nielsen NH, Hansen RG, Blackwell SB. Stomach temperature of narwhals (Monodon monoceros) during feeding events. Anim Biotelem. 2014;2:9. https://doi.org/10.1186/2050-3385-2-9.
Courtney MB, Scanlon B, Brown RJ, Rikardsen AH, Gallagher CP, Seitz AC. Offshore ocean dispersal of adult Dolly Varden Salvelinus malma in the Beaufort Sea. Polar Biol. 2018;41:817–25. https://doi.org/10.1007/s00300-017-2246-5.
Yurkowski DJ, Auger-Methe M, Mallory ML, Wong SNP, Gilchrist G, Derocher AE, et al. Abundance and species diversity hotspots of tracked marine predators across the North American Arctic. Biodivers Res. 2018;25:328–45. https://doi.org/10.1111/ddi.12860.
Harwood LA, Innes S, Norton P, Kingsley MCS. Distribution and abundance of beluga whales in the Mackenzie Estuary, southeast Beaufort Sea and west Amundsen Gulf during late July 1992. Can J Fish Aquat Sci. 1996;53:2262–73. https://doi.org/10.1139/f96-180.
Allen BM, Angliss RP. Alaska marine mammal stock assessments, 2012. U.S. Department of Commerce, NOAA Tech Memo NMFS-AFSC. 2013. 282 p. Report No. 245. https://repository.library.noaa.gov/view/noaa/4379.
Hauser DDW, Laidre KL, Suydam RS, Richard PR. Population-specific home ranges and migration timing of Pacific Arctic beluga whales (Delphinapterus leucas). Polar Biol. 2014;37:1171–83. https://doi.org/10.1007/s00300-014-1510-1.
Harwood LA, Stirling I. Distribution of ringed seals in the southeastern Beaufort Sea during late summer. Can J Zool. 1992;70:891–900. https://doi.org/10.1139/z92-127.
Citta JJ, Lowry LF, Quakenbush LT, Kelly BP, Fishbach AS, London JM, et al. A multi-species synthesis of satellite telemetry data in the Pacific Arctic (1987–2015): overlap of marine mammal distributions and core use areas. Deep Sea Res Part II Top Stud Oceanogr. 2018;152:132–53. https://doi.org/10.1016/j.dsr2.2018.02.006.
MacIntyre KQ, Stafford KM, Berchok CL, Boveng PL. Year-round acoustic detection of bearded seals (Erignathus barbatus) in the Beaufort Sea relative to changing environmental conditions, 2008–2010. Polar Biol. 2013;36:1161–73. https://doi.org/10.1007/s00300-013-1337-1.
Olnes J, Crawford J, Citta JJ, Druckenmiller ML, Von Duyke AL, Quakenbush L. Movement, diving, and haul-out behaviors of juvenile bearded seals in the Bering, Chukchi and Beaufort seas, 2014–2018. Polar Biol. 2020;34:1307–20. https://doi.org/10.1007/s00300-020-02710-6.
Hauser DDW, Laidre KL, Parker-Stretter SL, Horne JK, Suydam RS, Richard PR. Regional dive behaviour of Pacific Arctic beluga whales Delphinapterus leucas and possible associations with prey. Mar Ecol Prog Ser. 2015;541:245–64. https://doi.org/10.3354/meps11530.
Baechler J, Beck CA, Bowen WD. Dive shapes reveal temporal changes in the foraging behaviour of different age and sex classes of harbor seals (Phoca vitulina). Can J Zool. 2002;80:1569–77. https://doi.org/10.1139/z02-150.
Heerah K, Hindell M, Guinet C, Charrassin JB. A new method to quantify within dive foraging behaviour in marine predators. PLoS ONE. 2014;9:e99329. https://doi.org/10.1371/journal.pone.0099329.
Watts PD, Draper BA. Note on the behaviour of beluga whales feeding on capelin. Arct Alp Res. 1986;18:439–439.
Lefebvre SL, Lesage V, Michaud R, Humphries MM. Classifying and combining herd surface activities and individual dive profiles to identify summer behaviours of beluga (Delphinapterus leucas) from the St. Lawrence Estuary, Canada. Can J Zool. 2018;96:393–410. https://doi.org/10.1139/cjz-2017-0015.
Harwood LA, Smith TG, Auld JC, Melling H, Yurkowski DJ. Seasonal movements and diving of ringed seals, Pusa hispida, in the Western Canadian Arctic, 1999–2001 and 2010–11. Arctic. 2015;66:193–209. https://doi.org/10.14430/arctic4479.
Von Duyke AL, Douglas DC, Herreman JK, Crawford JA. Ringed seal (Pusa hispida) seasonal movements, diving, and haul-out behavior in the Beaufort, Chukchi, and Bering Seas (2011–2017). Ecol Evol. 2019;10:5595–616. https://doi.org/10.1002/ece3.6302.
Young BG, Loseto LL, Ferguson SH. Diet differences among age classes of Arctic seals: evidence from stable isotope and mercury biomarkers. Polar Biol. 2010;33:153–62. https://doi.org/10.1007/s00300-009-0693-3.
Hamilton CD, Kovacs KM, Lydersen C. Individual variability in diving, movement and activity patterns of adult bearded seals in Svalbard, Norway. Sci Rep. 2018;8:16988. https://doi.org/10.1038/s41598-018-35306-6.
Similä T, Ugarte F. Surface and underwater observations of cooperatively feeding killer whales in Northern Norway. Can J Zool. 1993;71:1494–9. https://doi.org/10.1139/z93-210.
Miller PJO, Shapiro AD, Deecke VB. The diving behaviour of mammal-eating killer whales (Orcinus orca): variations with ecological not physiological factors. Can J Zool. 2010;88:1103–12. https://doi.org/10.1139/Z10-080.
Seaman GA, Lowry LF, Frost KJ. Foods of Belukha whales (Delphinapterus leucas) in western Alaska. Cetology. 1982;44:1–19.
Loseto LL, Brewster JD, Ostertag SK, Snow K, MacPhee SA, McNicholl DG, Choy ES, Giraldo C, Hornby CA. Diet and feeding observations from an unusual beluga harvest in 2014 near Ulukhaktok, Northwest Territories, Canada. Arct Sci. 2018;4:421–31. https://doi.org/10.1139/as-2017-0046.
Bugaev AV, Shevlyakov EA. Wounding of Pacific salmon by predators in gillnet catches in the Russian economic zone in 2004. N Pac Anadr Fish Comm Bull. 2007;4:145–54. https://npafc.org/wp-content/uploads/Bulletins/Bulletin-4/145-154Bugaev.pdf
Kienle SS, Berta A. The better to eat you with: the comparative feeding morphology of phocid seals (Pinnipedia, Phocidae). J Anat. 2016;228:396–413. https://doi.org/10.1111/joa.12410.
Marshall CD, Kovacs KM, Lydersen C. Feeding kinematics, suction and hydraulic jetting capabilities in bearded seals (Erignathus barbatus). J Exp Biol. 2008;211:699–709. https://doi.org/10.1242/jeb.009852.
Lacroix GL. Large pelagic predators could jeopardize the recovery of endangered Atlantic salmon. Can J Fish Aquat Sci. 2014;71:343–50. https://doi.org/10.1139/cjfas-2013-0458.
Kane EA, Marshall CD. Comparative feeding kinematics and performance of odontocetes: belugas, Pacific white-sided dolphins and long-finned pilot whales. J Exp Biol. 2009;212:3939–50. https://doi.org/10.1242/jeb.034686.
Macleod CD, Reidenberg JS, Weller M, Santos MB, Herman J, Goold J, Pierce GJ. Breaking symmetry: the marine environment, prey size, and the evolution of asymmetry in cetacean skulls. Anat Rec. 2007;290:539–45. https://doi.org/10.1002/ar.20539.
Vester H, Hammerschmidt K. First record of killer whales (Orcinus orca) feeding on Atlantic salmon (Salmo salar) in northern Norway suggest a multi-prey feeding type. Mar Biodivers Rec. 2013;6:e9. https://doi.org/10.1017/S1755267212001030.
George JC, Suydam R. Observations of killer whale (Orcinus orca) predation in the northeastern Chukchi and western Beaufort Seas. Mar Mamm Sci. 1998;14:330–2. https://doi.org/10.1111/j.1748-7692.1998.tb00722.x.
George JC, Sheffield G, Reed DJ, Tudor B, Stimmelmayr R, Person BT, et al. Frequency of injuries from line entanglements, killer whales, and ship strikes on Bering-Chukchi-Beaufort seas bowhead whales. Arctic. 2017;70:37–46. https://doi.org/10.14430/arctic4631.
Willoughby AL, Ferguson MC, Stimmelmayr R, Clarke JT, Brower AA. Bowhead whale (Balaena mysticetus) and killer whale (Orcinus orca) co-occurrence in the U.S. Pacific Arctic, 2009–2018: evidence from bowhead whale carcasses. Polar Biol. 2020;43:1669–79. https://doi.org/10.1007/s00300-020-02734-y.
Fraker MA, Gordon CD, McDonald JW, Ford JKB, Cambers G. White whale (Delphinapterus leucas) distribution and abundance and the relationship to physical and chemical characteristics of the Mackenzie estuary. Fish Mar Serve Tech Rep. 1979. 56 p. Report No. 863. https://waves-vagues.dfo-mpo.gc.ca/Library/20122.pdf.
Gallagher CP, Howland KL, Bajno R, Sandstrom SJ, Reist JD. Population abundance, biological characteristics, and contribution to coastal mixed-stock fisheries of Dolly Varden (Salvelinus malma malma) from the Babbage River: 2010–2014. DFO Can Sci Advis Sec Res Doc. 2018. 105 p. Report No. 2018/029. https://www.dfo-mpo.gc.ca/csas-sccs/Publications/ResDocs-DocRech/2018/2018_029-eng.html.
Lima SL, Dill LM. Behavioral decisions made under the risk of predation: a review and prospectus. Can J Zool. 1990;68:619–40. https://doi.org/10.1139/z90-092.
Godin JJ. Evading predators. In: Godin JJ, editor. Behavioural ecology of teleost fishes. Oxford: Oxford University Press; 1997. p. 190–229.
Wright BM, Ford JKB, Ellis GM, Deecke VB, Shapiro AD, Battaile BC, Trites AW. Fine-scale foraging movements by fish-eating killer whales (Orcinus orca) relate to the vertical distributions and escape responses of salmonid prey (Oncorhynchus spp.). Mov Ecol. 2017;5:3. https://doi.org/10.1186/s40462-017-0094-0.
Mulder IM, Morris CJ, Dempson JB, Fleming IA, Power M. Marine temperature and depth use by anadromous Arctic char correlates to body size and diel period. Can J Fish Aquat Sci. 2020;77:882–93. https://doi.org/10.1139/cjfas-2019-0097.
Jepsen N, Thorstad EB, Havn T, Lucas MC. The use of external electronic tags on fish: an evaluation of tag retention and tagging effects. Anim Biotelem. 2015;3:1–23. https://doi.org/10.1186/s40317-015-0086-z.
Zahn MJ, Laidre KL, Stilz P, Rasmussen MH, Koblitz JC. Vertical sonar beam width and scanning behaviour of wild belugas (Delphinapterus leucas) in West Greenland. PLoS ONE. 2021;16:e0257054.
Austin D, Bowen W, McMillan J, Boness D. Stomach temperature telemetry reveals temporal patterns of foraging success in a free-ranging marine mammal. J Anim Ecol. 2006;75:408–20. https://doi.org/10.1111/j.1365-2656.2006.01057.x.
Bekkby T, Bjørge AJ. Variation in stomach temperature as indicator of meal size in harbour seal, Phoca vitulina. Mar Mamm Sci. 1998;14:627–37. https://doi.org/10.1111/j.1748-7692.1998.tb00750.x.
Shippee S, Townsend F, Deckert F, Knowles K. Monitoring swimming, diving, and forestomach temperature changes on instrumented wild dolphins to determine post-release foraging effort and success. Final technical report. Ft. Pierce (FL): Harbor Branch Oceanographic Institution; 2006. 46 p. Contract No. PWD2003-11.
Wells RS, McHugh KA, Douglas DC, Shippee S, McCabe EB, Barros NB, et al. Evaluation of potential protective factors against metabolic syndrome in bottlenose dolphins: feeding activity patterns of dolphins in Sarasota Bay, Florida. Front Endocrinol. 2013;4:139. https://doi.org/10.3389/fendo.2013.00139.
Majewski AR, Atchison S, MacPhee S, Eert J, Niemi A, Michel C, Reist JD. Marine fish community structure and habitat associations on the Canadian Beaufort shelf and slope. Deep-Sea Res Pt. 2017;121:169–82. https://doi.org/10.1016/j.dsr.2017.01.009.
Kastelein RA, Ford J, Berghour E, Wiepkema PR, van Boxsel M. Food consumption and reproduction of belugas (Delphinapterus leucas) in human care. Aquat Mamm. 1994;20:81–97. https://www.aquaticmammalsjournal.org/share/AquaticMammalsIssueArchives/1994/Aquatic_Mammals_20_2/20.2Kastelein.pdf.
Kleinberg SE, Yablokov AV, Bel’kovich BM, Tarasevich MN. Beluga (Delphinapterus leucas): investigation of the species. (Translated from Russian) Israel Program for Scientific Translations, Jerusalem; 1969. 376 pp.
Harwood LA, Kingsley MCS, Pokiak F. Monitoring beluga harvests in the Mackenzie Delta and near Paulatuk NT, Canada: harvest efficiency and trend, size and sex of landed whales, and reproduction, 1970–2009. Can Man Rep Fish and Aquat Sci. 2015. 32 p. Report No. 3059. http://publications.gc.ca/collections/collection_2021/mpo-dfo/fs70-5/Fs70-5-2020-075-eng.pdf.
Harwood LA, Zhu X, Angasuk L, Emaghok, L, Ferguson S, Gruben C, et al. Research, monitoring and hunter knowledge in support of the 2017 assessment of the eastern Beaufort Sea beluga stock. DFO Can Sci Advis Sec Res Doc. 2020. 48 p. Report No 2020/075. https://www.dfo-mpo.gc.ca/csas-sccs/Publications/ResDocs-DocRech/2020/2020_075-eng.pdf.
Budge SM, Cooper MH, Iverson SJ. Demonstration of the deposition and modification of dietary fatty acids in pinniped blubber using radiolabelled precursors. Physiol Bichem Zool. 2004;77:682687. https://doi.org/10.1086/420945.
Goetsch D, Conners MG, Budge SM, Mitani Y, Walker WA, Bromaghin JF, et al. Energy-rich mesopelagic fishes revealed as a critical prey resources for deep-diving predator using quantitative fatty acid signature analyses. Front Mar Sci. 2018;5:430. https://doi.org/10.3389/fmars.2018.00430.
Crossin GT, Hinch SG. A nonlethal, rapid method for assessing the somatic energy content of migrating adult Pacific salmon. Trans Am Fish Soc. 2005;134:184–91. https://doi.org/10.1577/FT04-076.1.
Hop H, Tonn WM, Welch HE. Bioenergetics of Arctic cod (Boreogadus saida) at low temperatures. Can J Fish Auqat Sci. 1997;54:1772–84. https://doi.org/10.1139/f97-086.
Hornby CA, Hoover C, Iacozza J, Barber DG, Loseto LL. Spring conditions and habitat use of beluga whales (Delphinapterus leucas) during arrival to the Mackenzie River estuary. Polar Biol. 2016;39:2319–34. https://doi.org/10.1007/s00300-016-1899-9.
Richard PR, Martin AR, Orr JR. 1997. Study of summer and fall movements and dive behaviour of Beaufort Sea belugas, using satellite telemetry: 1992–1995. Environmental Studies Research Funds Report. 1997. 26 p. Report No. 134. http://publications.gc.ca/collections/collection_2016/one-neb/NE22-4-134-eng.pdf.
Kasting NW, Adderley SAL, Safford T, Hewlett KG. Thermoregulation in beluga (Delphinapterus leucas) and killer (Orcinus orca) whales. Physiol Biochem Zool. 1989;1989(62):687–701. https://doi.org/10.1086/physzool.62.3.30157921.
Katsumata E, Furuta C, Katsumata H, Watanabe G, Taya K. Basal body temperature method for detecting ovarian cycle in the Captive Beluga (Delphinapterus leucas). J Reprod Dev. 2006;52:59–63. https://doi.org/10.1262/jrd.17066.
Melero M, Rodríguez-Prieto V, Rubio-García A, García-Párraga D, Sánchez-Vizcaíno JM. Thermal reference points as an index for monitoring body temperature in marine mammals. BMC Res Notes. 2015;8:411. https://doi.org/10.1186/s13104-015-1383-6.
Whittow G, Hampton I, Matsuura D, Ohata C, Smith R, Allen J. Body temperature of three species of whales. J Mamm. 1974;55:653–6. https://doi.org/10.2307/1379555.
Ford JKB, Ellis GM. Selective foraging by fish-eating killer whales Orcinus orca in British Columbia. Mar Ecol Progr Ser. 2006;316:185–99. https://doi.org/10.3354/meps316185.
Smith TG, Hamill MO, Taugbøl G. A review of the developmental, behavioural and physiological adaptations of the ringed seal, Phoca hispida, to life in the Arctic winter. Arctic. 1991;44:95–175. https://doi.org/10.14430/arctic1528.
Sinisalo T, Valtonen ET, Helle E, Jones RI. Combining stable isotope and intestinal parasite information to evaluate dietary differences between individual ringed seals (Phoca hispida botnica). Can J Zool. 2006;84:823–31. https://doi.org/10.1139/z06-067.
Oksansen SM, Niemi M, Ahola MP, Kunnasranta M. Identifying foraging habitats of Baltic ringed seals using movement data. Mov Ecol. 2015;3:33. https://doi.org/10.1186/s40462-015-0058-1.
Stirling I, Archibald WR, DeMaster D. Distribution and abundance of seals in the eastern Beaufort Sea. J Fish Res Bd Can. 1977;34:976–88. https://doi.org/10.1139/f77-150.
Irving L, Krog J. Body temperatures of Arctic and subarctic birds and mammals. J Appl Physiol. 1954;1954(6):667–80. https://doi.org/10.1152/jappl.1954.6.11.667.
Pikharev, GA. Some data on the feeding of the far eastern bearded seal. Izvestia TINRO. 1941;20:101–20. (Translated from Russian) Coyle, K, University of Alaska, Fairbanks.
Nikolaev AM, Skalkin VA. On the food of true seals of the eastern coast of Sakhalin. Environmental assessment of the Alaskan Continental Shelf. Annual reports of Principal Investigators for the year ending March 1977. Volume 1. Receptors—Mammals. U.S. Department of Commerce, NOAA, Environmental Research Laboratories and U.S. Department of the Interior, Bureau of Land Management, Boulder, CO; 1975. p. 384–8.
Jakobsson M, Mayer LA, Coakley B, Dowdeswell JA, Forbes S, Fridman B, et al. The international bathymetric chart of the Arctic Ocean (IBCAO) version 3.0. Geophys Res Lett. 2012;39:L12609. https://doi.org/10.1029/2012GL052219.
Acknowledgements
For Dolly Varden telemetry the authors acknowledge the support provided by Fisheries and Oceans Canada’s Species at Risk Program, Fisheries Joint Management Committee, Parks Canada Agency, Aklavik Hunters and Trappers Committee, and the West Side Working Group. The authors thank Frank Dillon (Aklavik), Jay Frandsen (Parks Canada), and Matt Garnett (Parks Canada) for assistance with sampling Dolly Varden and Great Slave Helicopters (Connor Gould) and Aklak Air for providing flights. For the Eastern Beaufort Sea (EBS) beluga telemetry programme, they acknowledge their partners the Inuvialuit Game Council and Fisheries Joint Management Committee, as well as members of the Tagging Advisory Group. They thank members of the field crew for their work in data collection, advice on project delivery, and discussions on beluga ecology: Nigel Hussey (University of Windsor), Greg O’Corry-Crowe (Florida Atlantic University), John Noksana Sr., Joseph Felix Jr., Raymond Ettagiak, James Pokiak and Mikkel Panaktalok (Tuktoyaktuk), Lawrence Kaglik and Linley Day (Inuvik), Dennis Arey and Dwayne Benoit (Aklavik), Patrick Akhiatak (Ulukhaktok), Norman Anikina (Sachs Harbour), Émilie Couture and Benjamin Lamglait (Granby Zoo/Canadian Wildlife Health Cooperative), Greg Elias (Aurora Research Institute), and Jeremy Hansen (Fisheries and Oceans Canada, Inuvik). They thank Laura Murray (Fisheries and Oceans Canada), Emily Way-Nee (Joint Secretariat), Jimmy Kalinek (Only Way Outfitting), and James Keevik for key logistical support for beluga telemetry. They thank the three anonymous reviewers for their constructive feedback in helping to improve the manuscript.
Funding
Financial and in-kind support for Dolly Varden telemetry was provided by Fisheries and Oceans Canada (Species at Risk Program, Fisheries Management, and Science), Inuvialuit Final Agreement implementation funds (Fisheries Joint Management Committee), Parks Canada Agency, and Department of Natural Resources Canada’s Polar Continental Shelf Program. Funding for the beluga telemetry programme was provided by Crown-Indigenous Relations and Northern Affairs Canada (Beaufort Regional Strategic Environmental Assessment), Fisheries and Oceans Canada (Intergovernmental Strategy, Strategic Program for Ecosystem-based Research and Advice, Arctic Science Fund, National Conservation Plan), Inuvialuit Final Agreement implementation funds (Fisheries Joint Management Committee), Tarium Niryuitait Marine Protected Area funds and Anguniaqvia niqiqyuam Marine Protected Area funds (Fisheries Joint Management Committee), and Natural Resources Canada’s Polar Continental Shelf Program.
Author information
Authors and Affiliations
Contributions
CPG conceived the Dolly Varden telemetry study; CPG and KLH secured funding to support Dolly Varden telemetry; LL and SM led programme implementation and secured funding and partnerships for the beluga telemetry programmes; CPG and EVL conducted fieldwork for Dolly Varden telemetry; LS, SM, and LL conducted fieldwork for beluga telemetry; CPG, LS, LL, and MBC conceptualized the manuscript and led the writing; LS and MBC performed data analyses and interpretation, and mapping. All authors reviewed and edited the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All capture and tagging methodology used for Dolly Varden and beluga whales were approved under Fisheries and Oceans Canada’s License to Fish for Scientific Purposes (S-19/20-3022-YK for Dolly Varden and S-18/19-3020-YK, S-19/20-3020-YK for beluga whale) and the Freshwater Institute’s Animal Care Committee (FWI-ACC-2019-33 for Dolly Varden and FWI-ACC-2018-24, FWI-ACC-2019-29 for beluga whale). For Dolly Varden telemetry we received approval from the Aklavik Hunters and Trappers Committee, and the West Side Working Group, while approval was provided by all Hunters and Trappers Committees in the Inuvialuit Settlement Region for beluga tagging telemetry.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gallagher, C.P., Storrie, L., Courtney, M.B. et al. Predation of archival tagged Dolly Varden, Salvelinus malma, reveals predator avoidance behaviour and tracks feeding events by presumed beluga whale, Delphinapterus leucas, in the Beaufort Sea. Anim Biotelemetry 9, 48 (2021). https://doi.org/10.1186/s40317-021-00271-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40317-021-00271-x