
Abstract
Background
High-throughput technologies have the potential to identify non-invasive biomarkers of liver pathology and improve our understanding of basic mechanisms of liver injury and repair. A metabolite profiling approach was employed to determine associations between alterations in serum metabolites and liver histology in patients with chronic hepatitis C virus (HCV) infection.
Methods
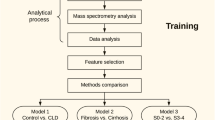
Sera from 45 non-diabetic patients with chronic HCV were quantitatively analyzed using 1H-NMR spectroscopy. A metabolite profile of advanced fibrosis (METAVIR F3-4) was established using orthogonal partial least squares discriminant analysis modeling and validated using seven-fold cross-validation and permutation testing. Bioprofiles of moderate to severe steatosis (≥33 %) and necroinflammation (METAVIR A2-3) were also derived. The classification accuracy of these profiles was determined using areas under the receiver operator curves (AUROCSs) measuring against liver biopsy as the gold standard.
Results
In total 63 spectral features were profiled, of which a highly significant subset of 21 metabolites were associated with advanced fibrosis (variable importance score >1 in multivariate modeling; R2 = 0.673 and Q2 = 0.285). For the identification of F3–4 fibrosis, the metabolite bioprofile had an AUROC of 0.86 (95 % CI 0.74–0.97). The AUROCs for the bioprofiles for moderate to severe steatosis were 0.87 (95 % CI 0.76–0.97) and for grade A2–3 inflammation were 0.73 (0.57–0.89).
Conclusion
This proof-of-principle study demonstrates the utility of a metabolomics profiling approach to non-invasively identify biomarkers of liver fibrosis, steatosis and inflammation in patients with chronic HCV. Future cohorts are necessary to validate these findings.
Similar content being viewed by others

Background
An estimated 185 million people worldwide are chronically infected with the hepatitis C virus (HCV) [1]. Chronic HCV carriers are at risk of progressive liver fibrosis, and up to 30 % can develop cirrhosis after decades of infection with significant risk of morbidity and mortality. Hepatic fibrosis occurs in the setting of liver injury from immune mediated chronic inflammation and hepatocyte cell turnover. This can ultimately lead to cirrhosis, manifested by architectural distortion of the liver and nodule formation [2]. Accumulation of the extracellular matrix occurs following metabolic and synthetic impairment of hepatocytes.
Recently the HCV infection treatment landscape has changed with the advent of new oral directly acting antiviral agents which offer >90 % successful treatment response rates in patients with HCV genotype 1 infection. Successful treatment is defined as a sustained virological response, SVR or undetectable HCV RNA by sensitive PCR assay 3 months after the end of anti-HCV treatment [3, 4]. The achievement of an SVR has been shown to correlate with reduced risk of liver disease progression and even reversal of liver fibrosis development. Thus, the currently approved antiviral therapy is recommended in all patients with advanced fibrosis or cirrhosis who have the highest risk of HCV-related complications such as liver failure and hepatocellular carcinoma. However, treatment is costly and not widely available, especially in resource-limited countries. Thus accurate fibrosis staging, especially the diagnosis of cirrhosis, is needed in clinical practice to determine the urgency and need for anti-HCV antiviral therapy [5, 6].
The current gold standard for the diagnosis HCV-related fibrosis and cirrhosis is liver biopsy. However biopsy is limited due to risk, invasive nature, technical issues (i.e., sample size, sampling error) and patient willingness. A number of complementary non-invasive tools have been developed and validated to diagnose and monitor liver fibrosis progression. The “physical approach” is characterized by the measurement of liver stiffness by diagnostic or ultrasound imaging techniques (i.e., Acoustic Radiation Force Impulse ultrasound imaging, and transient elastography; by FibroScan [7–11]. The “biological approach” focuses on serum biomarkers as well as developing profiles using high-throughput technologies such as genomics, transcriptomics, glycomics and metabolomics. Serum biomarkers include inexpensive tests such as the AST-to-Platelet Index (APRI), as well as the commercially developed FibroTest). Shaheen et al. reported that APRI under 0.5 could exclude significant fibrosis with 80 % accuracy [12]. It was however limited in accurately staging fibrosis in about 65 % of the patients in the study. Poynard et al. assessed the performance of Fibrotest and reported and area under the receiver operating characteristic curve of 0.80 (95 % confidence interval 0.78–0.82) for the identification of significant fibrosis (Stage 2–4) [13]. These various tests may also be limited by sensitivity, specificity, as well as cost and availability. Consequently, there remains a need for complementary non-invasive biomarkers to assist in treatment decisions and to monitor the progression or resolution of liver fibrosis.
Metabolomics as a high-throughput platform offers to capture the metabolic alterations that are reflective of the disease process. For example, we have successfully characterized gastrointestinal tumor characteristics using serum metabolic profiles [14, 15]. Since the liver is involved in important metabolic processes, pathological process such as hepatic fibrosis manifests as metabolic alterations that can assist in understanding the disease processes. For example, in a recent study, Jimenez et al. were able to characterize serum metabolites using 1H-NMR metabolomics in cirrhotic patients at risk of developing hepatic encephalopathy [16]. Recently, Baniasadi et al. were able to discriminate between cirrhotic HCV patients who were at high risk for developing hepatocellular carcinoma (HCC), from those who had already progressed to HCC using blood metabolite measurements [17]. Gao et al. were able to distinguish healthy controls when compared with serum samples from patients with liver cirrhosis and HCC [18], while HCV patients pre-therapy have been shown to have elevated levels of tryptophan in patients with SVR compared to those who were non-responders to antiviral therapy [19].
Despite this work in clinical populations, there is limited information specifically regarding the association of liver histology with clinically accessible metabolic profiles from blood or urine in HCV populations, the exception being a study examining HIV/HCV co-infection [20]. In the current study, patients with chronic HCV infection (>80 % genotype 1 infection) with varying degrees of fibrosis (F0–4), necroinflammation (A1–3) and steatosis were evaluated. We hypothesized that a metabolomics approach would be able to stratify HCV related liver histological disease progression by analysis of patient sera samples using a quantitative NMR metabolomics profiling approach. Using multivariate statistical analysis tools, we could identify non-invasive metabolite biomarkers that are capable to discriminate between various stages of fibrosis, necroinflammation and steatosis.
Methods
Study design
The study included patients chronically infected with the HCV seen at the Calgary Liver Unit, Viral Hepatitis Clinic, Department of Gastroenterology and Hepatology, University of Calgary. Study inclusion criteria included (1) adult patients (≥18 years of age,) (2) presence of chronic HCV infection (positive anti-HCV antibodies and HCV RNA in serum), (3) biopsy diagnosis of fibrosis and necroinflammation, according to METAVIR classification for HCV [21], (4) macrovesicular steatosis assessment by histopathology and graded as none (0–5 %), mild (5–32 %), moderate (33–65 %) and severe (≥66 %) steatosis. The exclusion criteria included: (1) obvious cirrhosis based on radiological investigations (e.g. nodular liver, splenomegaly), or clinical features of decompensated liver disease (e.g. jaundice, encephalopathy, ascites), (2) conditions that may alter the accuracy of biomarkers of fibrosis including extrahepatic obstruction, immunosuppression (e.g. due to concomitant HIV infection, medication), pregnancy, and systemic inflammatory conditions (e.g. sepsis, inflammatory bowel disease) [22], (3) excessive alcohol consumption defined as ≥40 g/d for men and ≥20 g/d for women in patients with HCV, (4) antiviral therapy for HCV within the previous 6 months. The study approval was obtained from the University of Calgary Conjoint Health Research Ethics Board (CHREB) and all study participants provided signed informed consent for study enrolment according to the Declaration of Helsinki. Relevant C = clinical and demographic information collected included, gender, age, HCV genotype, serum alanine-aminotransferase (ALT) levels, and body mass index (BMI).
Peripheral blood samples were collected approximately 24 h after liver biopsy was performed, after a 12-h fast. The isolated serum were immediately transferred to cryovials and stored at −80 °C for subsequent metabolomics analysis. In total 45 study participants with chronic HCV infection were enrolled, for sera spectra analysis. Participants were divided into three groups based on liver histopathology characteristics of fibrosis, necroinflammation and steatosis (Table 1).
Liver tissue sampling and histopathology evaluation
Percutaneous liver biopsy was performed under local anesthesia with an ultrasound guidance via the right coastal approach [23]. An 18 gauge (width 1.2 mm, cutting depth 2.2 cm) spring loaded, cutting needle (Bard®, Monopty® Percutaneous biopsy instrument; C.R. Bard-Inc., Billerica, MA) was used. If a sample less than 2.0 cm long was obtained after the first pass of the needle, an additional pass was performed. Following biopsy, patients were observed according to a standardized, 4 h protocol to observe for immediate complications [23]. The liver biopsy tissue was fixed, paraffin-embedded and stained with hematoxylin and eosin and Massaon’s trichome. All biopsies were examined by two hepatopathologists blinded to the clinical data. The size of biopsy, number of portal triads and fragmentation were recorded to account for the effect of these factors on the accuracy of the biomarkers under study [24, 25]. Liver fibrosis was staged from 0 to 4 according to the METAVIR classification [21] and NALFD score [22]. Histopathological assessment of fibrosis and necroinflammation were done using the METAVIR grading system. These classifications have excellent intra- and inter-observer agreement for liver fibrosis (kappa values >0.80) [21, 22].
The METAVIR scoring system assesses fibrosis in chronic HCV patients in accordance to a 5-stage classification, as previously published. Based on the percentage of involved hepatocytes, steatosis were divided into 4 groups as none (0–5 %), mild (5–32 %), moderate (33–65 %) and severe (≥66 %) steatosis.
1H NMR spectroscopy
1H NMR spectroscopy was performed using a protocol previously described [26]. For NMR analysis, serum samples were thawed on ice. 250 μl of serum sample were filtered through a prewashed Nanasep 3 K Omega Filter Eppendorf to remove high molecular weight (>3 kDa) compounds (e.g. large proteins, lipid complexes etc.). The filtrate was then centrifuged and buffered to a pH of 7.0 for analysis. Regular one-dimensional proton NMR spectra were obtained using a 600-MHz Bruker Ultrashield NMR spectrometer (Bruker Biospin, Milton, Canada). The spectra were acquired using a standard NOESY 1D pulse sequence that had good water suppression characteristics and is commonly used for metabolite profiling of serum samples [27]. Relaxation delay of 1 s was used; t1 was set to 4 μs and a 100 ms mixing time was employed. Initial samples for each batch were shimmed to ensure half-height line width of <1.1 Hz for the dimethyly-silapentane-sulphonate peak, calibrated to 0.0 ppm. Spectra were acquired with 1024 scans, then zero filled and Fourier transformed to 128 k data points using the Chenomx NMRSuite processor. Additional 2-dimensional NMR experiments were performed for the purpose of confirming chemical shift assignments, including homonuclear total correlation spectroscopy (2D 1H-1H TOCSY) and heteronuclear single quantum coherence spectroscopy (2D 1H-13C HSQC), using standard Bruker pulse programs.
Data processing for statistical analysis
Raw data from 1H NMR was processed and profiled using ChenomX NMR Suite software 4.6 (Cheonomx Inc., Edmonton, Canada) to a library of 63 compounds. 1H-NMR spectral data was evaluated using the strategy of “targeted profiling” [28]. This allowed quantification of metabolite concentrations in the serum samples. The data was subsequently log transformed, centered and unit variance scaled. Data analysis was done using multi-variate statistical analytical software SIMCA- P + (V12, Umetrics AB, Umea, Sweden). Firstly, a principal component analysis (PCA) was performed to detect any group separation based on variation in NMR signals. PCA was also performed to check the unsupervised segregation of the metabolome.
Orthogonal projection least squares-discriminant analysis (OPLS-DA) was done which allowed us to differentiate between the variables concerning the different stages of fibrosis. Model variance and predictive ability was assessed using R2 and Q2 values. The R2X and R2Y values were representative of the explained variation in the X and Y matrices respectively. The Q2Y value was indicative of the predictability of the model generated. Model significance was assessed using a cross-validated ANOVA based on seven-fold cross validation. Variables were selected according to the VIP (variables importance in projection) values, which were reflective of the correlation of the metabolites towards different response. Model performance was assessed using AUROC (area under the receiver operator curve).
Pathway analysis
Preliminary metabolite function was assigned using the Human Metabolome Database [29]. Dataset of metabolites that were differentially abundant in the NMR analysis were uploaded into the program. This was done independently for each comparison. Metabolite Set Enrichment Analysis (MSEA) [30] was used to elucidate the various biochemical pathways involved in HCV fibrosis, necroinflammatory disease and steatosis.
Results
Patients
In total 45 study participants with chronic HCV infection were enrolled, for sera spectra analysis. Participants were divided into three groups based on liver histopathology characteristics of fibrosis, necroinflammation and steatosis (Table 1). These included participants who had fibrosis and necroinflammation (classified according to the METAVIR score [21] and patients with moderate to severe steatosis. One-third of the patients were scored with F3–4 fibrosis; 13 and 42 % had moderate to severe steatosis and inflammation, respectively.
Metabolomic analysis
Metabolite bioprofile of advanced HCV fibrosis (F3–4)
For 1H NMR spectral profiling, a total of 63 features were profiled. The results indicated that 21 metabolites were differentially abundant associated with advanced fibrosis (variable importance score >1 in multivariate modeling) (Table 2). The 1H NMR spectra of two patient’s sera, with F4 (cirrhosis) and F1 stage of fibrosis are represented in Fig. 1a. It illustrates the relative decrease in the concentration of creatine in patients with cirrhosis (F4) in comparison with patients with F1 fibrosis, supporting the results indicated in the coefficient plot (Fig. 1b). The sera spectra of patients with F0-2 and F3-4 were well discriminated with the OPLS-DA model (R2 = 0.673) (Fig. 1c). The predictive ability of the model was measured by sevenfold cross validation (Q2 = 0.285), and cross-validated ANOVAl indicates a significant model (p = 0.008). Figure 1e is reflective of biochemical pathways over-represented in advanced fibrosis, with metabolites shown in Fig. 1d being relatively decreased as a function of increasing fibrosis, while those listed in Fig. 1f have been relatively increased.

Metabolite bioprofiling facilitates discrimination of advanced HCV fibrosis (F3–4). a Illustrates the relative decrease in concentration of creatine in HCV carrier with F4 fibrosis (cirrhosis) in comparison with a HCV carrier with Stage 1 (F1) liver fibrosis in the NMR spectra. b Represents the coefficient plot from the OPLS-DA analysis showing differences in serum metabolite concentration in the patients with HCV fibrosis (F3–4). C Shows OPLS-DA score plots of serum samples from HCV patients with fibrosis. Each data point is representative of the complete metabolite measurement from one HCV patient: blue square Stage 0–2, red square Stage 3–4. The x and y-axis represent the latent variable 1 and orthogonal component 2 respectively. R2 is the explained variance; Q2 is the predictive ability of the model; Model significance was assessed using a cross-validated ANOVA based on seven-fold cross validation (R2 = 0.673, Q2 = 0.285, p = 0.008). Biochemical pathways involved in HCV advanced fibrosis (F3–4). d Illustrates the metabolites that have been relatively decreased in the fibrosis group. e Indicates the biochemical pathways involved in fibrosis model; higher intensity with a higher association with the fibrotic group. f Represents the metabolites which have been relatively increased in the fibrotic group
Metabolite bioprofiles of necroinflammation (A2–3) and steatosis (>33 %)
Based on 1H-NMR profiling, patients with necroinflammatory activity classified as grade A2 and A3 (moderate to severe activity) could be partially distinguished from patients graded as A1 (minimal activity). The sera of patients with necroinflammation (A2–3) were distinguished from the rest of the necroinflammatory samples, based on differences in 17 spectral features (Table 2) in an OPLS-DA model with a variation of R2 = 0.405, Q2 = 0.102 and was weakly significant CV-ANOVA p = 0.10 (Fig. 2b). Of these metabolites that were identified, eight were shared with the fibrosis model (Table 3). The coefficients plot representing the differential abundance for each of the metabolites is illustrated in Fig. 2a.

Metabolite bioprofiling facilitates discrimination of HCV necroinflammation (A2–3). a Represents the coefficient plot from the OPLS-DA analysis showing differences in serum metabolite concentration in the patients with necroinflammatory disease (A2–3). b Shows OPLS-DA score plots of serum samples from HCV patients with necroinflammation. Each data point is representative of the complete metabolite measurement from one HCV patient: blue square A1, red square A2–3. The t[1] value represent the score of each sample in principal component 1. R2 is the explained variance; Q2 is the predictive ability of the model; Model significance was assessed using a cross-validated ANOVA based on seven-fold cross validation (R2 = 0.405, Q2 = 0.102, p = 0.10)
Study participants were divided into 4 categories based on degree of liver steatosis (<5, 5–32, 33–65 and ≥66 %). A distinction in-group with ≥33 % was observed in OPLS-DA with a variation of R2 Y = 0.67, Q2Y = 0.166 and weakly significant with a CV-ANOVA = 0.11 (Fig. 3b). Among the sixteen metabolites that were found to correspond to the patient sera samples with a steatosis of ≥33 %, four of those metabolites were common with the fibrosis model whereas seven were shared with the necroinflammation model (Table 3). Metabolites derived from the model and their respective differential abundance are illustrated in Fig. 3a.

Metabolite bioprofiling facilitates discrimination of HCV patients with steatosis (≥33 %). a Represents the coefficient plot from the OPLS-DA analysis showing differences in serum metabolite concentration in the patients with steatosis. b Shows OPLS-DA score plots of serum samples from HCV patients with steatosis. Each data point is representative of the complete metabolite measurement from one HCV patient: red triangle <5 %, green triangle 5–32 %, blue triangle 33–65 %, and black triangle ≥66 %. The x and y-axis represent the latent variable 1 and orthogonal component 2 respectively. R2 is the explained variance; Q2 is the predictive ability of the model; Model significance was assessed using a cross-validated ANOVA based on sevenfold cross validation (R2 = 0.67, Q2 = 0.16, p = 0.11)
Clinical applicability of the fibrosis, necroinflammation and steatosis model
As an assessment of the possible applicability of the metabolites from each pathophysiological model, the relative overlap between metabolites from each model was assessed (Fig. 4). Of the total set of metabolites deemed to be discriminating in each case, three metabolites were shared (methylsuccinate, creatine, and adenosine). In addition, asparagine was shared between the fibrosis and steatosis models. A listing of the overlapping metabolites is provided in Fig. 4b. Notably, a large subset of metabolites were unique to each model; twelve metabolites were discriminatory only in the fibrosis testing. This result is encouraging as it suggests that clinical applicability is truly based on metabolic differences.

Overall significant metabolites from models of fibrosis, necroinflammation and steatosis. a Venn diagram illustrating degree of overlap between metabolites changed with highest significance (VIP > 1) in each model. b Summary of metabolites by overlapping group
We further tested the predictive performance of discriminant model between sera of patients with stages F0–2 and F3–4, A0–1 and A2–3 and steatosis (≥33 %) respectively. This was done by constructing seven models with one-seventh of the data excluded from each of the seven models, with each sample excluded once. This method provided the predictive ability of the model.
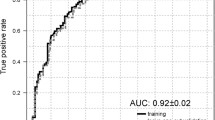
For the identification of F3–4 fibrosis, the metabolite bioprofile had an AUROC of 0.86 (95 % CI 0.74–0.97) as seen in Fig. 5. Metabolite bioprofiling facilitated the discrimination of advanced HCV fibrosis (F3-4) using a cross-validation cut off >1.39 (sensitivity 80 %, specificity 83 %, PPV 71 % and NPV 89 %) with an overall accuracy of 82 % (Fig. 2b). Similarly, the necroinflammation model, A2–3 yielded a metabolite bioprofile with an AUROC of 0.73 (95 % CI 0.57–0.89) (Fig. 5c). The steatosis model (≥33 %) produced a metabolite bioprofile with an AUROC of 0.87 (95 % CI 0.76–0.97) (Fig. 5d).

Clinical applicability of the advanced fibrosis (F3–4), necroinflammation and the steatotic model based on the receiver operator curve. a For the advanced fibrosis model, the AUROC is 0.86 (95 % CI 0.74–0.97). b Accuracy of the model based on the cross-validated score and the fibrosis stage. Accuracy of the model is 82 % with a sensitivity, specificity, positive predictive value and negative predictive value of 80, 83, 71 and 89 % respectively. c Shows the receiver operator characteristic (ROC) curve for the necroinflammation model with an AUROC of 0.73 (95 % CI 0.57–0.89). d Receiver operator characteristic (ROC) curve for the steatotic model with an AUROC of 0.87 (95 % CI 0.76–0.97)
Pathway analysis
In the fibrosis model, metabolites including amino acids, nucleic acids, and short chain fatty acids indicative of protein synthesis and catabolism as well as nitrogen metabolism were identified (Table 2; Fig. 1d–f; Additional file 1: Table S1). Alterations in metabolites were not gender-related and little or no model variance could be explained by other patient characteristics (e.g. age, body mass index). Asparagine, histidine and methionine are involved in protein biosynthetic pathways with asparagine was relatively decreased while histidine and methionine were relatively elevated with increasing fibrosis. Methionine is coupled to betaine, glycine and serine metabolism. Additionally asparagine is also involved in the ammonia-recycling pathway with glycine and glutamine. Glutamine is a member of pathways such as urea cycle, glutamate and pyrimidine metabolism.
Similar analysis of necroinflammatory disease metabolites indicated a relative up regulation of six amino acids, three dicarboxylic acids and one tricarboxylic acid (Table 2, Additional file 1: Figure S1). Pathway analysis suggests only protein biosynthesis to be impacted by this model (Additional file 1: Table S2).
On the other hand, the hepatic steatotic model (steatosis ≥33 %) included six amino acids, four dicarboxylic acids and two hydroxy acids (Table 2) over-representing several pathways related to energy and nitrogen metabolism, primarily ammonia and alanine metabolism, as well as the glucose alanine cycle (Additional file 1: Table S3, Figure S2).
It is worth noting that the pathway analysis of hepatic steatosis and necroinflammation included metabolites involved in both convergent and divergent sets of metabolic pathways. For example, in patients with steatosis several by-products of fat metabolism (e.g. acetone and 3-hydroxybutarate) and glycolysis (e.g. pyruvate and lactate) were up regulated (Fig. 3a). These compounds were not associated with fibrosis. At the same time, it is clear that nitrogen metabolism is central to all processes examined here.
Discussion
The presence of concomitant liver fibrosis, necroinflammatory activity as well as steatosis can only be reliably confirmed by liver histological analysis. However, liver biopsy is invasive and has risk of complications [31]. In addition, liver biopsy may be limited by the size of the specimen obtained as well as sampling, intraobserver, and interobserver variability [32]. Although liver biopsy is considered the gold standard for the evaluation of the grade of necroinflammation and stage of fibrosis [4, 33, 34] many guidelines also state that transient elastography, in combination with noninvasive biomarkers (i.e., FIB-4, FibroTest), is highly useful to evaluate fibrosis in patients with chronic HCV. We propose that metabolomics is ideal to study the disease induced changes caused by any pathological process as it enables us to describe the full complement of metabolites present in in biofluids as well as tissues [3]. These metabolites represent the final downstream product of all transcriptional and translational processes within a cell. Alterations due to a disease process result in changes in groups of metabolites, which represents a pattern unique to the disease process. This metabolomic profile can be studied in detail to understand the histological changes in the tissue, which eventually leads to disease progression. For example, Cassol et al. studied the metabolomic changes in patient population with HCV and HIV co-infection. It was noted that there was a direct relationship between elevation in plasma bile acids and non-invasive markers of liver fibrosis [20]. Patients who had recorded high scores on the FIB-4 and APRI scale also had increased levels of plasma GCA and TCA. Similarly, our analysis is reflective of increased bile acid synthesis, with glycine being down regulated in the advanced fibrosis model (Fig. 1b). This is illustrated in the cholesterol conjugation pathway, where the enzyme BA-CoA:amino acid N-acyltransferase (BAT), amidates BA-CoA with glycine to form tertiary bile acids such as GCA and TCA [35]. Interestingly, O-acylcarnitine, which is a marker of beta oxidation of long chain fatty acid, was elevated in the necroinflammation (A2–3) group (Additional file 1: Figure S1), a finding that was supported by Cassol et al. [20], who noted a positive correlation between elevated bile acid levels and acylcarnitines.
Saito et al. [19] investigated the metabolomic response in HCV patients undergoing therapy in relation to their viral load. It was noted that patients with SVR had an elevated tryptophan levels before the initiation of treatment. Although our study did not compare pre- and post-treatment changes in metabolites, an increase in the level of tryptophan was noted in both in the advanced necroinflammation (A2–3) and steatosis group (≥33 %) analyses. On histological examination, the advanced steatosis groups tended to have advanced inflammation characteristics as well explaining the overlap. Zhang et al. were able to identify twenty distinct urinary metabolites contributing to liver disease progression [36]. Thirteen of the identified metabolites were significantly increased whereas seven were decreased. Eleven of these urinary metabolites were common to the metabolite findings in this study and included glycine, citrate, arginine, betaine, histidine, leucine, aspartic acid, succinic acid, carnitine, valine and tyrosine. In our study, citrate was differentially increased in the necroinflammation (A2–3) model and was involved in the citric acid pathway in the steatosis (≥33 %) group. Histidine was upregulated in the fibrosis (F3–4), necroinflammation (A2–3) and steatosis (≥33 %) group. Carnitine, a urinary metabolite that was decreased in the patients with disease progression, was upregulated in the necroinflammation (A2–3) and steatosis (≥33 %) group. In the necroinflammation model, it was present in the form of O-acetylcarnitine participating in the glucose-alanine oxidation of branched chain fatty acids. However, in the steatosis (≥33 %) group, it was involved in gluconeogenesis, glutamate and tryptophan metabolism. Tyrosine a urinary metabolite that was decreased in patients with disease progression was found to be up regulated in the fibrosis (F3–4) and necroinflammation (A2–3) group. In these models, it was involved in protein biosynthesis, catecholamine metabolism and phenylalanine and tyrosine metabolism pathways.
We can interpret the analysis to suggest that liver being a metabolically complex organ is representative of unique metabolomic alterations reflective of the disease process. Fibrosis, necroinflammation and steatosis of the liver are separate histological entities; the results of this pilot study suggest that we can uniquely identify metabolites that are distinctive of the three histological conditions. At the same time, certain metabolites are shared amongst three histopathological conditions, such as those in involved in urea and nitrogen metabolism (asparagine and histidine) that can be attributed to their involvement in the common metabolite pathways of amino acid, fatty acid and carbohydrate metabolism (Additional file 1: Figure S3).
Using 1H-NMR, we have demonstrated that serum metabolomic profiles of patients with chronic HCV differ in accordance to their histopathological state. However, larger studies will be needed to comprehensively validate the metabolic alterations associated with the histological changes in chronic HCV patients. Factors that have limited our study include a small sample size and wide gender difference in patient groups. Information regarding non-anti-viral drug use was not available. Despite these limitations, our study was stringent with respect to the standardized timing of sample collection and dietary intake (i.e., serum after a 12-h fast) as well as correlation with liver histological analysis. Although it can be argued that the metabolome is influenced by genetic variations and environmental factors, studies have indicated that a distinct metabolite signature is conserved in disease with a common etiological factor and pathophysiology [37].
Conclusion
In this paper, we have described a proof-of-principle study that demonstrates that metabolomic profile of the chronic HCV carriers varies in accordance with the liver histopathology. Our study is able to demonstrate different metabolic profiles in association with clearcut pathology changes. It emphasizes the significance of metabolomic profiling as a promising technology for the non-invasive assessment of HCV-related liver histology. However, further experiments in the form of large-scale validation studies are required to develop our understanding of the histopathological contributions of the diseased liver site and the host, on the alterations in the metabolomic pathways, in addition to confirming the metabolomic profiles observed.
Abbreviations
- HCV:
-
hepatitis C virus
- AUROC:
-
area under the receiver operator curve
- 1H-NMR:
-
proton-nuclear magnetic resonance
- SVR:
-
sustained virological response
- APRI:
-
AST to platelet ratio
- HCC:
-
hepatocellulat carcinoma
- HIV:
-
human immunodeficiency virus
- F:
-
fibrosis
- A:
-
necroinflammation
- CHREB:
-
conjoint health research ethics board
- ALT:
-
alanine-aminotransferase
- AST:
-
aspartate-aminotransferase
- BMI:
-
body mass index
- NAFLD:
-
non-alcoholic fatty liver disease
- PCA:
-
principal component analysis
- OPLSDA:
-
orthogonol partial least squared discriminant analysis
- ANOVA:
-
analysis of variance
- VIP:
-
variables importance in projection
- HMDB:
-
human metabolome database
- PPV:
-
positive predictive value
- NPV:
-
negative predictive value
- FIB-4:
-
fibrosis-4
- GCA:
-
glycolic acid
- TCA:
-
taurocholic acid
References
Guidelines for the Screening (2014) Care and treatment of persons with hepatitis C infection. World Health Organization, Geneva
Saffioti F, Pinzani M (2015) Pathogenesis and evolution of liver fibrosis: cirrhosis or cirrhoses? Springer, Cham, pp 3–12
Myers RP, Shah H, Burak KW, Cooper C, Feld JJ (2015) An update on the management of chronic hepatitis C: 2015 consensus guidelines from the canadian association for the study of the liver. Can J Gastroenterol Hepatol. 29(1):19–34
AASLD/IDSA HCV Guidance Panel (2015) Hepatitis C guidance: AASLD-IDSA recommendations for testing, managing, and treating adults infected with hepatitis C virus. Hepatology 62(3):932–954
Myers RP, Fong A, Shaheen AAM (2008) Utilization rates, complications and costs of percutaneous liver biopsy: a population-based study including 4275 biopsies. Liver Int 28(5):705–712
Rockey DC, Caldwell SH, Goodman ZD, Nelson RC, Smith AD (2009) Liver biopsy. Hepatology 49:1017–1044
Pang JXQ, Zimmer S, Niu S, Crotty P, Tracey J, Pradhan F et al (2014) Liver stiffness by transient elastography predicts liver-related complications and mortality in patients with chronic liver disease. PLoS One 9(4):e95776
Sebastiani G, Ghali P, Wong P, Klein MB, Deschenes M, Myers RP (2014) Physicians’ practices for diagnosing liver fibrosis in chronic liver diseases: a nationwide, Canadian survey. Can J Gastroenterol Hepatol 28(1):23
Castera L, Pinzani M (2010) Non-invasive assessment of liver fibrosis: are we ready? Lancet 375(9724):1419–1420
Myers RP, Elkashab M, Ma M, Crotty P, Pomier-Layrargues G (2010) Transient elastography for the noninvasive assessment of liver fibrosis: a multicentre Canadian study. Can J Gastroenterol 24(11):661–670
Sadler MD, Crotty P, Fatovich L, Wilson S, Rabin HR, Myers RP (2015) Noninvasive methods, including transient elastography, for the detection of liver disease in adults with cystic fibrosis. Can J Gastroenterol Hepatol 29(3):139–144
Shaheen AAM, Myers RP (2007) Diagnostic accuracy of the aspartate aminotransferase-to-platelet ratio index for the prediction of hepatitis C-related fibrosis: a systematic review. Hepatology 46(3):912–921
Poynard T, Imbert-Bismut F, Munteanu M, Messous D, Myers RP, Thabut D, Ratziu V, Mercadier A, Benhamou Y, Hainque B (2004) Overview of the diagnostic value of biochemical markers of liver fibrosis (FibroTest, HCV FibroSure) and necrosis (ActiTest) in patients with chronic hepatitis C. Comp Hepatol 3:8
Bathe OF, Shaykhutdinov R, Kopciuk K, Weljie AM, McKay A, Sutherland FR et al (2011) Feasibility of identifying pancreatic cancer based on serum metabolomics. Cancer Epidemiol Biomarkers Prev. 20(1):140–147
Farshidfar F, Weljie AM, Kopciuk K, Buie WD, Maclean A, Dixon E et al (2012) Serum metabolomic profile as a means to distinguish stage of colorectal cancer. Genome Med. 4(5):42
Jiménez B, Montoliu C, MacIntyre DA, Serra MA, Wassel A, Jover M et al (2010) Serum metabolic signature of minimal hepatic encephalopathy by 1H-nuclear magnetic resonance. J Proteome Res 9(10):5180–5187
Baniasadi H, Gowda GAN, Gu H, Zeng A, Zhuang S, Skill N et al (2013) Targeted metabolic profiling of hepatocellular carcinoma and hepatitis C using LC-MS/MS. Electrophoresis 34(19):2910–2917
Gao H, Lu Q, Liu X, Cong H, Zhao L, Wang H et al (2009) Application of 1H NMR-based metabonomics in the study of metabolic profiling of human hepatocellular carcinoma and liver cirrhosis. Cancer Sci 100(4):782–785
Saito T, Sugimoto M, Igarashi K, Saito K, Shao L, Katsumi T et al (2013) Dynamics of serum metabolites in patients with chronic hepatitis C receiving pegylated interferon plus ribavirin: A metabolomics analysis. Metabolism. 62(11):1577–1586
Cassol E, Misra V, Holman A, Kamat A, Morgello S, Gabuzda D (2013) Plasma metabolomics identifies lipid abnormalities linked to markers of inflammation, microbial translocation, and hepatic function in HIV patients receiving protease inhibitors. BMC Infect Dis 13(1):203
The French METAVIR Cooperative Study Group, Bedossa P (1994) Intraobserver and interobserver variations in liver biopsy interpretation in patients with chronic hepatitis C. Hepatology 20(1):15–20
Poynard T, Munteanu M, Imbert-Bismut F, Charlotte F, Thabut D, Le Calvez S et al (2004) Prospective analysis of discordant results between biochemical markers and biopsy in patients with chronic hepatitis C. Clin Chem 50(8):1344–1355
McGill DBD (2001) Liver biopsy: when, how, by whom, and where? Curr Gastroenterol Rep 3(1):19–23
Poynard TT, McHutchison JJ, Manns MM, Myers RPR, Albrecht JJ (2003) Biochemical surrogate markers of liver fibrosis and activity in a randomized trial of peginterferon alfa-2b and ribavirin. Hepatology 38(2):481–492
Kleiner DED, Brunt EME, Van Natta MM, Behling CC, Contos MJM, Cummings OWO et al (2005) Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 41(6):1313–1321
Weljie AM, Dowlatabadi R, Miller BJ, Vogel HJ, Jirik FR (2007) An inflammatory arthritis-associated metabolite biomarker pattern revealed by 1H NMR spectroscopy. J Proteome Res 6(9):3456–3464
Nicholson JK, Lindon JC, Holmes E (1999) “Metabonomics”: understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 29(11):1181–1189
Weljie AM, Newton J, Mercier P, Carlson E, Slupsky CM (2006) Targeted profiling: quantitative analysis of 1H NMR metabolomics data. Anal Chem 78(13):4430–4442
Wishart DS, Knox C, Guo AC, Eisner R, Young N, Gautam B et al (2009) HMDB: a knowledgebase for the human metabolome. Nucleic Acids Res 37(Database):D603–D610
Xia J, Wishart DS (2010) MSEA: a web-based tool to identify biologically meaningful patterns in quantitative metabolomic data. Nucleic Acids Res 38(Web Server):W71–W77
Eisenberg E, Konopniki M, Veitsman E, Kramskay R, Gaitini D, Baruch Y (2003) Prevalence and characteristics of pain induced by percutaneous liver biopsy. Anesth Analg 96(5):1392–1396
Regev A, Berho M, Jeffers LJ, Milikowski C, Molina EG, Pyrsopoulos NT et al (2002) Sampling error and intraobserver variation in liver biopsy in patients with chronic HCV infection. Am J Gastroenterol 97(10):2614–2618
European Association for Study of Liver (2015) EASL recommendations on treatment of hepatitis C 2015. J Hepatol 63(1):199–236
Spratlin JL, Serkova NJ, Eckhardt SG (2009) Clinical applications of metabolomics in oncology: a review. Clin Cancer Res 15(2):431–440
Thomas C, Pellicciari R, Pruzanski M, Auwerx J, Schoonjans K (2008) Targeting bile-acid signalling for metabolic diseases. Nat Rev Drug Discov 7(8):678–693
Zhang A-H, Sun H, Han Y, Yan G-L, Yuan Y, Song G-C et al (2013) Ultraperformance liquid chromatography-mass spectrometry based comprehensive metabolomics combined with pattern recognition and network analysis methods for characterization of metabolites and metabolic pathways from biological data sets. Anal Chem 85(15):7606–7612
Griffin JL, Shockcor JP (2004) Metabolic profiles of cancer cells. Nat Rev Cancer 4(7):551–561
Authors’ contributions
AMW, CSC, HJV and RPM conceived the study, participated in its design and coordination and helped draft the manuscript. ZG was involved in the pathological analysis of the acquired samples. PMC and PZ were involved in acquisition and organization of patient samples. AAZMS contributed to the initial analysis of the data set. MOS participated in data analysis, interpretation and writing of the manuscript. AMW has contributed to critically revising the manuscript and supervised the project for MOS. All authors read and approved the manuscript.
Competing interests
The authors declare that they have no competing interests.
Authors’ information
MOS is a Clinical Fellow at the Department of Molecular and Laboratory Medicine, McMaster University in Hamilton, Ontario. He received his Internal Medicine training at the McMaster University. He is currently doing his Masters in Clinical Biomarkers at University of Calgary, Calgary, Alberta.
RPM is Associate Professor of Medicine and Director of the Viral Hepatitis Clinic at the University of Calgary. He received his Internal Medicine training at the University of Western Ontario, Gastroenterology fellowship at the University of Calgary, and Hepatology research fellowship at the Université de Paris VI in Paris, France. He completed his M.Sc. in Epidemiology at the University of Calgary. RPM is the past Chair of the Education Committee of the Canadian Association for the Study of the Liver (CASL) and a member of the National Education Advisory Committee of the Canadian Liver Foundation. In 2012, he was awarded a Queen Elizabeth II Diamond Jubilee Medal for his contributions to research and patient care in liver disease.
CSC is a Assistant Professor of Medicine. She received her Internal Medicine training from Memorial University of Newfoundlan, Gastroenterology at the University of Calgary and Hepatology fellowship at University of California, San Francisco. She is the recipient of the 2012 CIHR New Investigator Award for Biomedical/Clinical Research and has received operating grant bridge funding from CIHR (2010), and junior faculty awards including the 2010 AASLD Jan Albrecht Commitment to Clinical and Translational Research Award and the 2011 American Gastroenterology Association Research Scholar Award. In 2012, she received an infrastructure grant from the Canadian Foundation for Innovation and Alberta Advanced Education Technology to develop a translational health research clinical database and a Biobank.
ZHG is currently the Pathologist-in-Chief at the McGill University Health Center which includes six university-affiliated hospitals and the Chair for the Department of Pathology at McGill University which includes 8 university-affiliated hospitals. He obtained his Medical degree from Qingdao Medical College, Master degree from Harbin Medical University, and Ph.D. degree from Peking Union Medical College. He received his post-doctoral fellowship training at the Johns Hopkins Hospital, pathology residency training at Dalhousie University, and subspecialty pathology fellowship at the University of Chicago.
AAMS is affiliated with the Liver Unit, Division of Gastroenterology and Hepatology, Department of Medicine, University of Calgary, Calgary, Alberta, Canada.
PMC RN is the Research Manager for the Liver Unit at the University of Calgary.
PZ is senior research associate at the Ling Research Group, University of Calgary.
HJV is Professor of Biochemistry at the University of Calgary. He has been the academic lead of the academic Metabolomics Research Centre since its inception in 2006. His research interests include the application of protein chemistry and molecular biology methods, used in combination with high-resolution NMR spectroscopy. HJV holds the Armstrong Chair in Molecular Cancer Research.
AMW is the Research Assistant Professor of Pharmacology and leads the Weljie Lab at the Department of Systems Pharmacology and Translational Therapeutics at the University of Pennsylvania.
Ethics approval and consent to participate
As mentioned, the study approval was obtained from the University of Calgary Conjoint Health Research Ethics Board (CHREB) and all study participants provided signed informed consent for study enrolment according to the Declaration of Helsinki. Relevant C = clinical and demographic information collected included, gender, age, HCV genotype, serum alanine-aminotransferase (ALT) levels, and body mass index (BMI).
Funding
This research was supported in part by the Canadian Institute of Health Research (CSC and RPM), Canadian Liver Foundation (RPM), Alberta Heritage Foundation for Medical Research (HJV), Alberta Innovates-Health Solution (HJV) and American Gastroenterological Association (CSC).
Author information
Authors and Affiliations
Corresponding authors
Additional information
M. Omair Sarfaraz and Robert P. Myers contributed equally to this work
Additional file
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sarfaraz, M.O., Myers, R.P., Coffin, C.S. et al. A quantitative metabolomics profiling approach for the noninvasive assessment of liver histology in patients with chronic hepatitis C. Clin Trans Med 5, 33 (2016). https://doi.org/10.1186/s40169-016-0109-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40169-016-0109-2