
Abstract
This article focuses on the pathogenic significance of Helicobacter species naturally colonizing the stomach of dogs, cats and pigs. These gastric “non-Helicobacter (H.) pylori Helicobacter species” (NHPH) are less well-known than the human adapted H. pylori. Helicobacter suis has been associated with gastritis and decreased daily weight gain in pigs. Several studies also attribute a role to this pathogen in the development of hyperkeratosis and ulceration of the non-glandular stratified squamous epithelium of the pars oesophagea of the porcine stomach. The stomach of dogs and cats can be colonized by several Helicobacter species but their pathogenic significance for these animals is probably low. Helicobacter suis as well as several canine and feline gastric Helicobacter species may also infect humans, resulting in gastritis, peptic and duodenal ulcers, and low-grade mucosa-associated lymphoid tissue lymphoma. These agents may be transmitted to humans most likely through direct or indirect contact with dogs, cats and pigs. Additional possible transmission routes include consumption of water and, for H. suis, also consumption of contaminated pork. It has been described that standard H. pylori eradication therapy is usually also effective to eradicate the NHPH in human patients, although acquired antimicrobial resistance may occasionally occur and porcine H. suis strains are intrinsically less susceptible to aminopenicillins than non-human primate H. suis strains and other gastric Helicobacter species. Virulence factors of H. suis and the canine and feline gastric Helicobacter species include urease activity, motility, chemotaxis, adhesins and gamma-glutamyl transpeptidase. These NHPH, however, lack orthologs of cytotoxin-associated gene pathogenicity island and vacuolating cytotoxin A, which are major virulence factors in H. pylori. It can be concluded that besides H. pylori, gastric Helicobacter species associated with dogs, cats and pigs are also clinically relevant in humans. Although recent research has provided better insights regarding pathogenic mechanisms and treatment strategies, a lot remains to be investigated, including true prevalence rates, exact modes of transmission and molecular pathways underlying disease development and progression.
Similar content being viewed by others

1 Introduction
The stomach has long been considered a sterile and hostile environment for bacteria because of the extremely acidic conditions. However, the opposite is true. Among other resident microbes of the stomach, various gastric Helicobacter species are able to colonize the stomach of animals and humans and may cause gastric disease [1,2,3]. Helicobacter (H.) pylori is by far the best known and best studied gastric Helicobacter species with a global prevalence in humans of more than 50%, where this prevalence is generally higher in low-income countries compared to developed countries [4]. Helicobacter pylori infection has been associated with severe gastric disease in humans as it may cause gastritis, peptic ulcer disease, gastric carcinoma and low-grade gastric B cell mucosa-associated lymphoid tissue (MALT) lymphoma [5, 6]. This bacterium was isolated from a human gastric biopsy for the first time in 1982 by Marshall and Warren [7]. Since then, many other gastric Helicobacter species have been identified in human patients’ stomachs, admittedly at a much lower prevalence [8,9,10]. These gastric non-Helicobacter pylori Helicobacter species (NHPH) naturally reside in animal hosts, including dogs, cats and pigs. To date, the Helicobacter genus includes 48 distinct, validly published species, among which 17 gastric and 31 enterohepatic NHPH [11, 12]. In recent years, increasingly more insights have been gained regarding the pathogenic significance and zoonotic potential of the hitherto identified gastric NHPH, however, a lot remains to be investigated.
This review aims to provide an overview of the existing evidence concerning the pathogenic significance of dog-, cat- and pig-associated gastric NHPH in their respective animal hosts and in human patients.
2 Taxonomy of gastric non-Helicobacter pylori Helicobacter species
Only recently a consensus has been reached on the complex nomenclature of the gastric NHPH, a still expanding group of helicobacters. First, the name “Gastrospirillum hominis” was proposed in 1989 after the first discovery of Gram-negative, long, helical-shaped bacteria, morphologically distinct from the curve-shaped H. pylori, residing in the gastric mucosa of patients with gastrointestinal symptoms [13]. Based on 16S rRNA gene sequencing, it was discovered that these bacteria actually belonged to the Helicobacter genus and so, the NHPH were renamed to “H. heilmannii”, after Konrad Heilmann who described for the first time a large case study of NHPH infections in humans [9, 14, 15]. Further genetic analysis showed significant differences in the 16S rRNA gene nucleotide sequence among different species within this group of gastric NHPH, which led to a subclassification into “H. heilmannii” type 1 and “H. heilmannii” type 2 [9, 16, 17].
In 1990, another “Gastrospirillum” species was discovered residing in the gastric mucosa of pigs, named “Gastrospirillum suis” [18, 19]. Through sequencing of the 16 rRNA gene, fluorescent in situ hybridization and electron microscopy studies, it was proven that “Gastrospirillum suis” belonged to the Helicobacter genus and sequencing of the urease A and B (ureAB) genes proved that it was identical to “H. heilmannii” type 1. Therefore, the name “Candidatus Helicobacter suis” was proposed and the bacterium was designated a new taxon [17, 20, 21]. The term “Candidatus” indicates that, at that time, the agent could not yet be isolated and cultivated in vitro [22]. Acquirement of the first in vitro isolates of this bacterium in 2006 led to its characterization as H. suis [17].
“Helicobacter heilmannii” type 2 on the other hand represents several dog- and cat-associated gastric NHPH which have been proven to be genetically distinct upon isolation and characterization. These include H. felis [23], H. bizzozeronii [24], H. salomonis [25], H. cynogastricus [26], H. baculiformis [27], H. heilmannii sensu stricto (H. heilmannii s.s.) [21, 28] and H. ailurogastricus [29].
To avoid confusion concerning the term H. heilmannii, it has been proposed to use the terms H. heilmannii sensu lato (H. heilmannii s.l.) and H. heilmannii s.s. [30]. The term H. heilmannii s.l. is then used to refer to all gastric NHPH, which is useful when only results of histopathology or electron microscopy or crude taxonomic data are available, whereas the term H. heilmannii s.s. or another specific species name of a gastric NHPH refers to a single fully characterized NHPH.
Sequencing of the 16S rRNA or 23S rRNA genes allows differentiation between the pig-associated H. suis and the group of dog- and cat-associated gastric NHPH [16]. However, it does not allow to differentiate between the different dog- and cat-associated gastric NHPH [26, 27]. To achieve this anyway, sequencing of the (partial) ureAB genes is the preferred method since the sequences of these genes are available for all gastric NHPH [9, 21]. Moreover, sequencing of the ureAB genes does not allow differentiation between H. heilmannii s.s. and its closest relative H. ailurogastricus [29, 31]. These two species can be distinguished through genome sequencing based approaches [29] or sequencing of the IceA gene and LpsA gene [32]. Whole cell protein profiling is another method to distinguish the gastric NHPH, however this method requires pure in vitro cultures [30]. More recently, MALDI-TOF mass spectrometry has been shown to allow differentiation between gastric Helicobacter species cultured in vitro [33]. Given the fastidious nature of the gastric NHPH, in vitro isolation and cultivation is complicated, as is characterization of potential new species detected in human and animal stomachs [9, 30]. An overview of the dog-, cat- and pig-associated gastric NHPH and their corresponding phenotypic characteristics is presented in Table 1.
3 Pathogenic significance, prevalence and transmission of Helicobacter suis in pigs
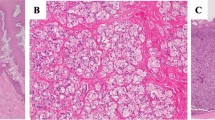
Helicobacter suis infection in pigs has been associated with decreased daily weight gain [34], gastritis [35,36,37] and ulceration of the non-glandular part of the stomach, the pars oesophagea [9, 38,39,40,41], therefore impacting animal health, welfare and production. Images of lesions associated with ulcer formation at the level of the pars oesophagea of the porcine stomach are presented in Figure 1. Helicobacter suis colonizes the fundic and pyloric gland zone of the stomach. During the more acute phase of the infection, this may result in decreased gastric acid secretion in this distal, glandular part of the stomach, hereby affecting the composition of the microbiota of the pars oesophagea. An important factor in this microbiota composition shift was discovered to be related with an increased colonization by Fusobacterium (F.) gastrosuis [42]. Production of epithelial cell death inducing metabolites by F. gastrosuis potentially plays a role in porcine gastric ulceration [43]. During the more chronic phase of infection, secretion of excessive amounts of gastric acid in the distal part of the stomach is induced, which may lead to increased contact of the pars oesophagea with hydrochloric acid. As this part of the porcine stomach is not protected by mucus, hyperkeratosis, erosion and ulcer formation is likely to occur [9, 44]. To confirm this hypothesis, experimental studies involving pigs infected with H. suis and F. gastrosuis should be performed in order to find out which putative virulence factors are expressed and secreted and whether these are functional [43]. Furthermore, the etiology of ulceration of the pars oesophagea is most probably multifactorial and therefore, mere H. suis infection of the porcine stomach may not be sufficient to cause gastric ulcers [35]. Several predisposing factors associated with development of ulceration of the porcine stomach have been described, including nutritional, housing, management and environmental factors. Finely ground pelleted feed and disruptions in feed delivery are the two most important ones. Increased risk may also result from hot weather and outbreaks of acute infectious diseases [45]. Recently, Nunes et al. were able to detect H. suis DNA in the gastric mucosa of a wild boar, of which the stomach presented with macroscopic signs of antral inflammation [46].

Ulcer formation at the level of the pars oesophagea in slaughter pigs. A normal mucosa; the pars oesophagea is delineated by a black dotted line, B severe hyperkeratosis, C hyperkeratosis with few erosions (erosions are indicated by black arrows), D hyperkeratosis with several erosions (erosions are indicated by black arrows), E ulceration.
Similar to what has been clearly demonstrated for H. pylori, some H. suis strains might be more pathogenic than others [47]. To this end, multilocus sequence typing (MLST) of H. suis strains has been executed, but a possible relationship between MLST type and virulence has not been demonstrated yet [48].
There are clear indications that H. suis originates from non-human primates, as it was discovered to colonize the stomach of rhesus and cynomolgus monkeys and a host jump from macaques to pigs would have happened between 100,000 and 15,000 years ago [8]. In general, host jumps like these are known to lead to an increase in disease severity, while coevolution between a bacterium and its natural host generally results in less severe pathogenicity. This accounts for H. suis too. Infections with H. suis are usually asymptomatic in non-human primates and cause gastric disease in pigs. Furthermore, pig domestication has significantly impacted the spread of H. suis within the pig population following the host jump [1].
Several independent studies conducted in different countries report different prevalence rates of H. suis colonizing the gastric mucosa of pigs at slaughter age, varying from about 10 to 60% or more, mainly depending on the geographic area, the diagnostic tools used and the management and hygiene standards applied [35,36,37,38, 41, 44]. The prevalence is found to be increasing with the pig’s age, being 2% in suckling piglets and 90% in adult pigs [49].
The low prevalence in suckling piglets is probably due to anti-microbial effects of the sow’s milk through protective antibodies and other antibacterial factors [50]. As this disappears after weaning, the prevalence of H. suis infections increases rapidly as regrouping of piglets favors the spread of the few H. suis infections already present before weaning to noninfected pigs [9]. Furthermore, the proportion of Lactobacillus populations in the upper gastrointestinal tract microbiota decreases upon weaning, possibly resulting in a higher risk of developing a H. suis infection. The decrease in the proportion of Lactobacillus populations attenuates the natural resistance against infection or colonization by pathogens. This is due to decreased competition for nutrients and epithelial binding sites, less production of antimicrobial factors such as lactic acid and bacteriocins and an increased pH in the gastrointestinal tract, creating a less hostile environment for bacteria [51, 52]. Therefore, the potential of Lactobacillus strains as probiotic bacteria that protect the gastric environment has been suggested, since they are abundantly present in porcine stomachs without ulceration of the pars oesophagea [52].
Since H. suis DNA has been detected in saliva but not in feces of naturally and experimentally infected pigs, transmission is suggested to occur via the oral-oral route or the gastric-oral route [9]. In one study Helicobacter DNA was detected by a genus specific PCR assay in saliva and fecal samples from pigs with gastric ulceration. All these samples reacted negative in a H. suis specific PCR assay, whereas H. suis DNA was frequently detected in gastric samples. As the researchers suggest, these findings may indicate that DNA from Helicobacter species other than H. suis was detected in saliva and feces from these pigs [53]. These might be enterohepatic Helicobacter species, since in previous studies several enterohepatic species, including H. bilis and H. trogontum, have been detected in and isolated from porcine fecal samples [54,55,56].
4 Pathogenic significance, prevalence and transmission of dog- and cat-associated gastric non-Helicobacter pylori Helicobacter species in their natural hosts
The clinical significance of most gastric NHPH infections in cats and dogs remains controversial or yet to be explored. No conclusive evidence exists on the association between clinical signs, including chronic vomiting and inappetence, and the presence of gastric NHPH [57]. Concerning the more recently discovered NHPH H. baculiformis, H. ailurogastricus and H. cynogastricus, the clinical significance in their respective animal hosts as well as in humans remains unknown [26, 27, 29].
After investigating the clinical impact of the presence of gastric NHPH in the stomachs of cats showing clinical signs of gastrointestinal disease, Bridgeford et al. hypothesized that gastric NHPH infections in cats may be associated with feline gastric MALT lymphoma [58]. Scanziani et al. detected lymphoid follicular hyperplasia with mild pangastric mononuclear inflammation and eosinophilic infiltrates in cats naturally infected with H. felis [59]. Besides, correlations have been found between the degree of colonization by NHPH of the feline gastric mucosa and the extent of lymphocyte and plasma cell infiltration, the number of lymphoid follicles and the occurrence of fibrosis [60]. However, in cats diagnosed with H. heilmannii s.s. infection and infection with other, unclassified NHPH only minimal mononuclear inflammation was detected, as was also the case in uninfected cats. Indeed, different species and strains of NHPH may impact the severity of gastritis in cats, as well as the species and strain of the host, and the duration of infection. Furthermore, it has been suggested that induction of gastric inflammation may require factors other than Helicobacter infection alone [59]. In a more recent cohort study including 56 cats with and without chronic gastritis, gastric Helicobacter species specified as H. heilmannii s.s. were present in 28 cases (50%), with no evidence of significant associations with severity of gastric disease [61]. This supports evidence from another study in which H. heilmannii s.s. infection was frequently present in pet cats (38 of 49; 78%), although no significant correlation between the presence of infection and the degree of gastritis was found [62]. Similarly, Norris et al. found that in a cohort of healthy pet cats, infection with a gastric Helicobacter species closely related to H. felis was frequently present, with no evidence of significant gastritis in any case [63].
Several research groups did not find any correlation between NHPH infection and (the severity of) gastric lesions in dogs [60, 64,65,66,67,68]. However, Kubota-Aizawa et al. reported that infection of dogs with gastric NHPH, particularly H. heilmannii s.s., may be associated with more severe gastritis compared to dogs negative for NHPH. Furthermore, they suggested that gastric NHPH infection in dogs may be associated with a higher frequency of chronic diarrhea compared to dogs not infected with NHPH [69]. Experimental infection with H. felis in gnotobiotic Beagle dogs has been associated with histopathologic changes including lymphofollicular gastritis, with lymphoid nodules being most abundantly present in the fundus and body of the stomach [70]. Simpson et al. reported opposite findings after experimental infection with H. felis in specific-pathogen-free, Helicobacter-free Beagle dogs [71]. Taking into account the low density of colonization and limited duration of infection, they reported that H. felis is most likely not a gastric pathogen in dogs as they found no association between H. felis infection and the severity of gastritis in dogs. Mild gastric inflammation and lymphoid follicles were observed in both infected and uninfected dogs and the gastric secretory axis was not different in infected compared to uninfected dogs. In another study, Simpson et al. showed that the gastric function in dogs with a naturally acquired NHPH infection was not hampered and antimicrobial and antisecretory therapy administration caused suppression of the infection rather than eradication of the NHPH [72]. Again, several factors may underlie these contradictory findings, including differences in virulence of different species and strains of NHPH, the strain of the host, and the duration of infection.
As stated earlier, coevolution of a bacterium and its natural host generally leads to decreased pathogenicity. Dog- and cat-associated gastric NHPH coevolved with their host far before domestication of either cats or dogs. Therefore it is hypothesized that these NHPH cause mainly asymptomatic infections in cats or dogs and domestication has had little impact on the spread of these bacteria within the population of cats and dogs [1]. Furthermore, co-infections with multiple gastric NHPH species are commonly detected in dogs and cats. In that respect, dog- and cat-associated gastric NHPH can be regarded as sympatric (“i.e. two bacterial species sharing the same niche and host who frequently encounter one another”). Hence, extensive genetic exchange has been detected among these gastric NHPH, with this interspecies admixture being most evident in H. heilmannii s.s. and H. bizzozeronii. Also, the cat-associated H. baculiformis and dog-associated H. cynogastricus have been found to share a significant amount of DNA with H. felis and have therefore been defined as hybrids [1].
The prevalence of gastric NHPH in dogs and cats is found to be high, irrespective of clinical signs. The prevalence of gastric NHPH in clinically healthy dogs ranges between 67 and 86% and between 61 to 100% in dogs presented for investigation of chronic vomiting [9, 60, 64, 67, 68, 71, 73,74,75,76]. In naturally infected dogs, H. bizzozeronii was reported to be the most prevalent gastric NHPH [67, 77, 78]. Gastric NHPH in cats were detected in 41% to 100% of clinically healthy cats and were slightly more prevalent in cats presented for investigation of chronic vomiting [9, 59, 60, 75, 76, 79]. In cats, H. felis and H. heilmannii s.s. have been found to be the predominant NHPH infecting the gastric mucosa [61, 75].
Gastric NHPH have already been detected in the saliva and feces of dogs and cats, suggesting that transmission could happen via saliva and feces [64, 65, 80]. On the other hand, Craven et al. claimed that transmission via saliva is unlikely, based on the low frequency of NHPH detection in the saliva of dogs, compared to the frequent detection of NHPH in the gastric mucosa [81]. The exact mode of transmission still remains unclear.
5 Zoonotic potential of gastric non-Helicobacter pylori Helicobacter species
It has been shown that H. suis, H. felis, H. bizzozeronii, H. salomonis and H. heilmannii s.s. have zoonotic potential and are potentially clinically relevant in humans [1]. The zoonotic potential of the other dog- and cat-associated gastric NHPH, H. ailurogastricus, H. baculiformis and H. cynogastricus, has not yet been established.
Prevalence rates of gastric NHPH infections in human patients undergoing a gastroscopy have been reported to vary between 0.2% and 6%, depending on the geographical area and the detection methods used [9, 82]. Populations where animals and humans live closely together and sanitary conditions are poor could be prone to an increased NHPH prevalence rate [64]. PCR-based studies investigating human gastric biopsy samples with histological evidence of presence of NHPH showed that H. suis had the highest prevalence (14 to 37%), followed by H. salomonis (21%), H. felis (15%), H. heilmannii s.s. (8%) and H. bizzozeronii (4%) [3, 75, 83]. These data predate the description of H. ailurogastricus and therefore no distinction between H. heilmannii s.s. and its closest relative, H. ailurogastricus, was made in these studies [29]. Co-infections with more than one NHPH or with H. pylori and NHPH have also been reported [75, 83,84,85,86].
Due to the very fastidious growth characteristics of gastric NHPH, their focal colonization pattern in the human stomach and the limited amount of gastric tissue that is usually available when a biopsy sample is taken from human patients, only very few gastric NHPH isolates from humans have been obtained worldwide. The first H. bizzozeronii strain was isolated from a human patient in Denmark in 2001 [87]. Later, this species was also isolated from a Finnish patient presenting with severe dyspeptic symptoms, including nausea and vomiting, and chronic active gastritis [88]. In Germany, H. felis was isolated in 2012 from the gastric mucosa of a 14-year-old girl presenting with persistent epigastric pain and episodes of vomiting without diarrhea, accompanied with mild chronic gastritis [89]. Helicobacter felis has also been isolated from a patient presenting with acute abdominal pain, nausea and vomiting of brown fluid, symptoms that appeared to be related to severe acute gastritis [90]. Only recently, in 2020, Rimbara et al. succeeded in obtaining pure cultures of H. suis strains from three human patients with gastric diseases, including gastric MALT lymphoma [47, 91]. Isolation and cultivation of other gastric NHPH from human patients with gastric diseases has not yet been described.
Transmission of gastric NHPH to humans is generally suggested to happen through direct or indirect contact with dogs, cats or pigs, and potentially via consumption of contaminated water [9, 83, 92,93,94,95,96,97,98,99]. For H. suis, an additional means of transmission may include consumption of contaminated pork [100]. These means of direct and indirect transmission are further validated by reports of molecular analyses proving the great genetic similarity of gastric NHPH strains present in humans and these animal hosts [8, 47, 61, 93, 94]. Furthermore, since laboratory mice are easily colonized by most gastric NHPH, the role of wild mice as a vector for the transmission of gastric NHPH should also be considered. Nunes et al. raise awareness for the potential role of wild boars and other wild animal species as a reservoir for H. suis and possible source of infection for humans [46, 101]. Although, several earlier reports revealed that H. suis is absent in the majority of wild boar populations, which is in accordance with the fact that the host jump of H. suis from non-human primates to pigs occurred rather recently [8]. Therefore, this route of transmission needs further investigation. To date, no cases of human-to-human transmission have been reported [8].
Infection with gastric NHPH in humans has been associated with chronic gastritis, peptic and duodenal ulcer disease and low-grade MALT lymphoma of the stomach [7, 9, 82, 102,103,104,105,106]. Occasionally, cases of acute gastric mucosal lesions have also been reported [90, 107]. Since the risk of developing gastric MALT lymphoma is higher with NHPH than with H. pylori [108] the association between gastric NHPH and gastric MALT lymphoma has been most clearly described in literature. In addition, many studies have been performed on eradication of gastric NHPH as a potential cure for MALT lymphoma. On the other hand, the severity of lesions associated with chronic gastritis caused by gastric NHPH has been reported to be lower compared to H. pylori [108, 109].
Several research groups have recently investigated the clinical relevance of the potentially zoonotic gastric NHPH in large cohorts of patients. Among 236 Japanese gastric disease patients who tested negative for H. pylori based on a rapid urease test, 49 (20.8%) tested positive for NHPH using PCR and immunohistochemical methods. Helicobacter suis was detected in 20 patients, H. heilmannii s.s./H. ailurogastricus in 7 patients and the other 22 species remained unidentified. Gastric diseases within this group of patients included nodular gastritis, gastric MALT lymphoma, chronic gastritis and gastroduodenal ulcers, with no significant association between the Helicobacter species detected and the types of disease diagnosed. Standard eradication therapy was administered to 45 of these NHPH positive patients. This led to successful eradication of the infectious agent in all cases based on PCR results and improvement of endoscopic findings in most cases, except for some MALT lymphoma cases [82]. Furthermore, out of 3847 Japanese patients who underwent biopsy of the gastric mucosa between 2010 and 2020, 50 patients (1.30%) were diagnosed with NHPH gastritis based on microscopic findings after Giemsa staining. PCR analysis detected mainly H. suis (26/28) (2/28 were H. heilmannii s.s./H. ailurogastricus) and pathology was described as “crack-like mucosa” and nodular gastritis in the gastric antrum and regular arrangement of collecting venules in the gastric corpus [110]. In Turkey, a study including 110 children with gastric complaints revealed the presence of H. suis infection in two cases and H. heilmannii s.s./H. ailurogastricus in one case by performing PCR analysis. The former presented with active chronic gastritis without macroscopic lesions, while the latter presented with hyperemia without presence of gastric inflammation. Also, a mixed infection with H. suis and H. pylori was detected in one patient who presented with gastric ulceration in combination with moderate, active chronic gastritis [111]. Similar studies in western hospitals are currently lacking, but such studies are needed to establish the clinical relevance of these potentially important zoonotic NHPH and potential eradication programs.
Also of clinical interest, H. pylori infection has been described to play a role in the development and progression of central nervous system (CNS) disorders being linked, for example, to an increased risk of all-cause dementia [112] and to the onset of idiopathic parkinsonism (IP) and its motor symptoms [113,114,115]. Recent findings also revealed possible associations between H. suis infection and CNS disorders, including IP and Alzheimer’s disease (AD). In gastric biopsies from IP patients, a significantly higher presence of H. suis DNA (27%) was detected compared to control patients without evidence of clinical IP symptoms (2%) [10]. In addition, a blood sample from a patient affected by IP and AD tested positive for H. suis DNA. Following eradication therapy, both gastric and neurological symptoms improved substantially [10]. Recently, H. suis infection in IP patients was also associated with higher mortality [116]. It is not yet clear how a gastric H. suis infection might impact the CNS [114]. One hypothesis starts from gastric inflammation induced by H. suis. This is associated with loss of the gastrointestinal barrier function, causing a leaky gut with leakage of TLR4 ligands (and other molecules) into the blood. This might affect the blood-cerebrospinal fluid barrier and induce microgliosis, resulting in cognitive decline. Another pathway assumes a possible relationship with production of gamma-glutamyl transpeptidase and urease by H. suis which might result in a depletion of glutamine, a precursor for several neurotransmitters, and continuous slightly higher levels of ammonia, respectively. Changes in the gastrointestinal microbiome associated with a H. suis infection, might also impact brain homeostasis. Similar to how H. suis has already been associated with changes of the microbiome of the pars oesophagea of the porcine stomach, gastric NHPH have indeed been associated with shifts in structure and function of the gastric microbiome in human patients too [117]. The exact relevance in the pathogenesis of gastric diseases and the development of CNS disorders remains to be further investigated.
The efficacy of administration of standard H. pylori eradication therapy, typically consisting of a proton pump inhibitor and 2 or 3 antibiotics, in order to eradicate the gastric NHPH and achieve remission of the associated gastric pathology, is currently being investigated [99, 118,119,120,121]. In a patient diagnosed with H. suis infection and chronic gastritis, 10 days of standard triple therapy (esomeprazole, metronidazole and amoxicillin) led to H. suis eradication and regression of the gastric mucosal lesion upon esophagogastroduodenoscopy (EGD) three months post therapy. Also, there was no recurrence of similar lesions upon EGD within two years after administration of the eradication therapy [118]. Another case report demonstrated that, three months post standard triple eradication therapy in a patient with asymptomatic H. suis gastric MALT lymphoma, the bacterium was no longer detected, the erosive mucosal lesions improved, small lymphocyte infiltration decreased and lymphoepithelial lesions had disappeared. However, lymphoid nodules were still present. After nine months, no lymphoid infiltrations could be detected and complete remission was achieved [119]. Furthermore, a complete response rate to eradication therapy was achieved in 4 cases of H. suis positive gastric MALT lymphoma with nodular gastritis-like appearance, where both the infection and gastric disease were eradicated. Standard triple eradication therapy in a H. heilmannii s.s. infected patient diagnosed with multiple gastric ulcers also led to ulcer improvement and H. heilmannii s.s. eradication [99]. Other research groups reported similar results [103, 110, 122, 123] and confirmed earlier animal studies [124]. These findings suggest a definite causal relationship between gastric NHPH infection and the development of gastric disease [125].
Antimicrobial susceptibility tests have revealed that acquired antimicrobial resistance occasionally occurs in several gastric NHPH, including H. suis isolates from pigs and macaques [126], feline H. heilmannii s.s. isolates [31] and feline H. felis isolates [127]. The minimum inhibitory concentration (MIC) of amoxicillin for a H. suis isolate obtained from a human patient (16 mg/L) was approximately 10 times higher than that of three other isolates from humans (1 mg/L) which might be due to several mutations in the penicillin binding proteins PBP2 and FtsI (PBP3) [47]. Presence of acquired resistance in some NHPH isolates might complicate treatment as already described for H. pylori. Determinations of MIC indicated that porcine H. suis strains are intrinsically less sensitive to aminopenicillins than non-human primate H. suis strains and other gastric Helicobacter species. Helicobacter suis strains also seem to be intrinsically less susceptible to metronidazole than H. pylori strains. Although aminopenicillins and metronidazole are often used for treatment of human patients infected with gastric Helicobacter species, the possible therapeutic significance of these findings is not clear [128].
6 Rodent models in research for gastric non-Helicobacter pylori Helicobacter species
Rodent models (particularly mice and gerbils) are frequently used to understand the basic principles of gastric NHPH infection as well as for translational research. This ranges from patho- and immunogenicity studies to the search for potential curative and preventive therapies. For instance, experimental infection in mice with a H. suis strain originally isolated from pigs has been suggested to elicit a predominant Th2/Th17 response [129, 130], where a Th2 response may be associated with B cell accumulation leading to the development of gastric MALT lymphoma [131]. In another study, inoculation of mice and Mongolian gerbils with H. suis strains isolated from stomachs of non-human primates and pigs has been shown to induce a Th17 response and elevated CXCL13 levels accompanied by severe gastric inflammation. The immune response evoked was shown to depend on the experimental host rather than the original host to which the strain belongs [132]. Upregulation of CXCL13 levels has also been observed in H. suis infected pigs [44, 133]. Since CXCL13 is a chemokine which attracts B cells, this pathway is hypothesized to be involved in the development of gastric MALT lymphoma associated with gastric NHPH infection. Furthermore, mice experimentally infected with H. felis have long been used as a model for human H. pylori infection [134].
In order to screen for potential antibiotic and other treatment regimens, several mouse models have been developed [124, 135,136,137]. Guided by the mindset that administration of certain Lactobacillus strains might lead to reduced gastric NHPH colonization, administration of the Lacobacillus gasseri strain SBT2055 in a C57BL/6 J mouse model infected with H. suis, effectively suppressed the progression of gastric MALT lymphoma [138]. Vaccination as a mode of prophylactic immunization against NHPH, including H. felis and H. suis, has been tested in mouse models as well, but no formulation has provided complete protection yet. Major side effects, including post-vaccination gastritis and increased mortality rates, have been reported and have hampered this research domain [139,140,141]. However, promising results were obtained in a study which proved inhibition of the development of MALT lymphomas after immunization based on H. felis whole cell sonicate in a mouse model challenged with H. felis [142]. Completely unexpectedly, it was recently discovered that experimental infection with H. suis in a 6-OHDA Parkinson’s disease mouse model had (partial) neuroprotective effects on Parkinson’s disease pathology, mediated by gene expression changes in brain tissue related to reduction of oxidative stress [143].
Although rodent models are indispensable in gaining novel insights into pathogenicity and possible cure, significance in the relevant host, and thus translatability, must always be taken into account.
7 Virulence mechanisms of gastric non-Helicobacter pylori Helicobacter species
An overview of the virulence factors and their roles in pathogenesis is presented in De Witte et al. [56]. The most important virulence factors of gastric NHPH are briefly summarized below.
Gastric helicobacters as a group share several virulence mechanisms, although some virulence factors are Helicobacter species- and even strain-dependent. They are mainly involved in colonization, induction of gastric pathologies and host immune evasion.
Gastric helicobacters are capable to survive the extremely acidic conditions of the stomach lumen and, at the same time, they can penetrate the outer mucus layer covering the gastric mucosa in order to reach their ideal niche which has an external pH of 5 to 6.
Characteristically, all gastric helicobacters produce the enzyme urease, containing subunits UreA and UreB which are assembled into a catalytically active, nickel-containing dodecamer via the actions of the accessory proteins UreE, UreF, UreG and UreH [144, 145]. An inner membrane protein, UreI, forms a proton-gated urea channel in order to upregulate urease activity under acidic conditions and allow rapid entry of urea into the cytoplasm [146]. Urease hydrolyzes urea into carbon dioxide and ammonia, hereby neutralizing the gastric acid and creating a pH neutral microenvironment surrounding the bacterium [9]. Urease activity and UreI have both been proven to be essential in the pathogenesis of H. pylori, as colonization in animal models of infection is hampered in case either one of these is lacking [146,147,148].
In order to allow the gastric helicobacters to penetrate the outer mucus layer and move towards the gastric mucosal surface where they naturally reside, motility of the bacteria is essential. To this end, gastric NHPH make use of their helical shape and the presence of bipolar flagella [9, 149, 150]. The ideal niche for NHPH is located deep in the gastric pits, as opposed to H. pylori, which is usually detected at the surface epithelium [9]. Furthermore, gastric helicobacters possess genes encoding proteins involved in chemotactic behavior. As such they are able to sense the surrounding pH and move away from low pH regions towards more neutral regions [9]. Helicobacter pylori has been found to prefer the colonization of injured epithelium as it may possess high amounts of favorable nutrients [151] and sense the amount of bacteria in close proximity in order to spread out and avoid shortage of resources [152].
Adherence and binding to epithelial cells is an important step in the colonization of the gastric mucosa by helicobacters. To this end, several mucins and glycolipids expressed on the apical surface of gastric mucosal epithelial cells play a crucial role. Mucins are long and filamentous glycoproteins and therefore, an ideal first point of interaction between the host tissue and helicobacters [153, 154]. In BALB/c mice experimentally infected with H. heilmannii s.s., expression of the secreted mucin Muc6 and the transmembrane mucin Muc13 was upregulated in the first 9 weeks post-infection [155]. A positive correlation was found between the number of H. heilmannii s.s. bacteria and the increased expression of Muc6 in the antrum and Muc13 in the fundus, suggesting the importance of these mucins in Helicobacter colonization. Upregulation of Muc13 expression remained until 52 weeks post-infection, confirming earlier suggestions about its potential involvement in promoting the transition from chronic inflammation to cancer [156]. Mucins, such as the cell surface mucin MUC1 to which H. pylori is able to bind, have also been shown to serve as a releasable decoy in order to inhibit colonization by mucin-binding bacteria, and have been shown to create steric hindrance in order to avoid adhesion of non-mucin binding bacteria to the cell surface [153]. Also, H. suis infection has been shown to lead to a decrease in glycan structures to which H. suis is able to bind in the pig gastric mucus niche, a decrease in the H. suis-binding ability of human mucins, and to alter the porcine and human mucin composition in a way that the growth-inhibiting effect of mucins gives way to a growth-enhancing effect. This is thought to lead to a more stable and inhabitable niche for H. suis, likely supporting long-term colonization and outcome of infection [157]. Furthermore, H. suis has been shown to bind to both human and pig mucins and glycolipids via two modes of adhesion. One mode of adhesion is to glycans terminating with Galβ3GlcNAc (eg. lactotetraosylceramide present in pig stomach) at both neutral and acidic pH and the other is to negatively charged structures at acidic pH [158]. Since the glycosylation profile of gastric mucosa is dependent on the animal species, glycan receptors involved in the binding by different Helicobacter species have been found to differ, as part of host adaptation. Essentially, different gastric NHPH dispose of different adhesion mechanisms mediated by proteins with alternative receptor specificity. In the canine gastric mucosa, NHPH have been suggested to possibly bind via terminal α-GalNAc and type 2 Lewis antigens, and H. heilmannii s.s. has been shown to adhere the most [159]. The significance of these findings in the physiopathology of NHPH infection in dogs remains to be further investigated.
The genome of gastric helicobacters encodes a large set of outer membrane proteins (OMPs) involved in colonization and virulence. According to comparative genomic studies, gastric NHPH lack all known adhesins discovered in H. pylori so far [29, 160,161,162]. However, they share few homologs of the Hor and Hom family [160, 162] and in H. heilmannii s.s., HofE and HofF have been identified as adhesins with an important role in IL-1β induced gastric MUC13 expression [163]. Proteomic and phylogenetic analysis of the OMPs of gastric NHPH identified the Hof proteins as a Helicobacter-specific OMP family, found widespread amongst all gastric Helicobacter species, with a putative function in adhesion (Family 33) [162]. This study also discovered a Helicobacter-specific SfpA/LpxR OMP (Family 36) important in immune evasion, possibly via lipopolysaccharide modulation, and a Helicobacter-specific vacuolating cytotoxin A (VacA)-like cytotoxin family with a function in colonization capacity (Family X3) in gastric helicobacters. Presence of an outer membrane factor (OMF) (Family 42) and Imp/OstA (Family 14), both important in antimicrobial resistance, TonB-dependent OMPs with a function in metal and vitamin-uptake (Family 3 and Family X2), and an outer membrane phospholipase (OMPLA) (Family 38) involved in colonization capacity have also been identified in most gastric Helicobacter species. So, it is known that Helicobacter-, species- and even site-specific members of OMPs/OMP families exist, but their significance in virulence and colonization remains to be further explored [162].
Regarding virulence factors associated with induction of gastric pathologies, both cytotoxin-associated gene pathogenicity island (cagPAI) and VacA, which are two major cytotoxic virulence factors in H. pylori [5], are not present in any of the gastric NHPH discussed in this review.
Gamma-glutamyltranspeptidase (GGT) is an epithelial cell death inducing virulence factor present in each gastric Helicobacter species in a highly conserved manner. The metabolic function of GGT for gastric NHPH is to break down extracellular glutathione (GSH) and glutamine in order to produce glutamate, which is incorporated intracellularly into the tricarboxylic acid cycle. This glutamate-generating metabolism is essential for survival and growth of gastric helicobacters [164]. Gamma-glutamyltranspeptidase as a virulence factor has been identified as a direct causal factor for Helicobacter-induced gastric disorders. Unraveling of the underlying molecular mechanism has shown that degradation of extracellular reduced GSH by GGT generates GSH degradation products which increase the extracellular concentrations of reactive oxygen species (ROS) in a cell-independent manner, resulting in oxidative cell damage on different levels, including cellular lipid peroxidation. This oxidative stress induced cell damage ultimately leads to different types of cell death of gastric epithelial cells, with apoptosis in case of low extracellular ROS concentrations and oncosis/necrosis in case of high extracellular ROS concentrations [165].
Several other possible virulence factors have been determined to be encoded for in the genome of gastric NHPH based on genome sequence analyses of H. suis [160], H. felis [166], H. bizzozeronii [161] and H. heilmannii s.s. [167], but their exact significance in the development of gastric pathologies associated with NHPH infections remains unclear.
In addition to aforementioned OMPs and other virulence factors involved in host immune evasion for persistence of infection, gastric helicobacters also possess the ability to influence dendritic cell maturation and cytokine production. In pigs, induction of semi-maturation of monocyte-derived dendritic cells by H. suis has been observed, eliciting expansion of regulatory T cells [133]. Via this regulatory T cell response, a tolerogenic phenotype is induced, thereby stimulating chronic colonization [133, 168]. Since H. pylori urease has been found to bind CD47, thereby impeding the folding and transport of the MHCII molecules [169], and since this is a highly conserved virulence factor among gastric helicobacters, urease is hypothesized to play a role in host immune evasion. Furthermore, GGT and L-asparaginase II have been found to inhibit T cell proliferation, further promoting the tolerogenic phenotype [170, 171].
In H. ailurogastricus, several homologs encoding H. pylori virulence and colonization factors previously identified in H. heilmannii could not be retrieved and it was found to have a lower binding capacity for gastric epithelial cells in vitro. It is therefore suggested to be a low-virulence gastric NHPH [29].
Besides disparities in virulence among the gastric NHPH, strain-specific differences have also been identified. Rimbara et al. observed differences in the progression of gastric mucosal metaplasia in mice inoculated with different H. suis strains isolated from human patients, indicating variation in virulence between H. suis isolates, as has frequently been reported for H. pylori [47]. In the accessory genome of a more virulent H. suis strain, many genes were detected which were absent in a less virulent strain, including type 4 secretion system-associated genes located in plasticity zones. Further studies are necessary to unravel the significance of this finding.
8 Conclusions
Besides H. pylori, several other gastric helicobacters appear to be clinically relevant in humans. At least four dog-, cat- and pig-associated gastric NHPH have been detected in humans and have been proven to be a causal factor in gastric disease. Clinical and preclinical research has provided evidence regarding pathogenic mechanisms and possible treatment strategies. However, a lot remains to be investigated, including true prevalence rates, exact modes of transmission and other zoonotic aspects, and molecular pathways underlying disease development and progression. Further elucidation of these research questions will require large patient cohort studies incorporating standardized detection methods. Also, the optimization of in vitro isolation methods for these extremely fastidious gastric NHPH could be of great value for a better understanding of the pathogenic mechanisms and developing potential treatment and prevention strategies. To date, 8 gastric NHPH which are commonly present in dogs, cats and pigs have been fully characterized. The clinical significance of H. suis in pigs has been established, whereas the clinical significance of cat- and dog-associated gastric NHPH in their natural host remains controversial or yet to be explored. However, evidence regarding coevolution suggests that this may be low.
References
Smet A, Yahara K, Rossi M, Tay A, Backert S, Armin E, Fox JG, Flahou B, Ducatelle R, Haesebrouck F, Corander J (2018) Macroevolution of gastric Helicobacter species unveils interspecies admixture and time of divergence. ISME J 12:2518–2531
Yang I, Nell S, Suerbaum S (2013) Survival in hostile territory: the microbiota of the stomach. FEMS Microbiol Rev 37:736–761
Bento-Miranda M, Figueiredo C (2014) Helicobacter heilmannii sensu lato: an overview of the infection in humans. World J Gastroenterol 20:17779–17787
Hooi JKY, Lai WY, Ng WK, Suen MMY, Underwood FE, Tanyingoh D, Malfertheiner P, Graham DY, Wong VWS, Wu JCY, Chan FKL, Sung JJY, Kaplan GG, Ng SC (2017) Global prevalence of Helicobacter pylori infection: systematic review and meta-analysis. Gastroenterology 153:420–429
Kusters JG, Van Vliet AHM, Kuipers EJ (2006) Pathogenesis of Helicobacter pylori Infection. Clin Microbiol Rev 19:449–490
Dunn BE, Cohen H, Blaser MJ (1997) Helicobacter pylori. Clin Microbiol Rev 10:720–741
Marshall BJ, Warren JR (1984) Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 323:1311–1315
Flahou B, Rossi M, Bakker J, Langermans JAM, Heuvelman E, Solnick JV, Martin ME, O’Rourke J, Ngoan LD, Hoa NX, Nakamura M, Øverby A, Matsui H, Ota H, Matsumoto T, Foss DL, Kopta LA, Omotosho O, Franciosini MP, Casagrande Proietti P, Guo A, Liu H, Borilova G, Bracarense AP, Lindén SK, De Bruyckere S, Zhang G, De Witte C, Smet A, Pasmans F, Ducatelle R, Corander J, Haesebrouck F (2018) Evidence for a primate origin of zoonotic Helicobacter suis colonizing domesticated pigs. ISME J 12:77–86
Haesebrouck F, Pasmans F, Flahou B, Chiers K, Baele M, Meyns T, Decostere A, Ducatelle R (2009) Gastric helicobacters in domestic animals and nonhuman primates and their significance for human health. Clin Microbiol Rev 22:202–223
Blaecher C, Smet A, Flahou B, Pasmans F, Ducatelle R, Taylor D, Weller C, Bjarnason I, Charlett A, Lawson AJ, Dobbs RJ, Dobbs SM, Haesebrouck F (2013) Significantly higher frequency of Helicobacter suis in patients with idiopathic parkinsonism than in control patients. Aliment Pharmacol Ther 38:1347–1353
Parte AC (2018) LPSN - List of prokaryotic names with standing in nomenclature (Bacterio.net), 20 years on. Int J Syst Evol Microbiol 68:1825–1829
Genus: Helicobacter. https://lpsn.dsmz.de/genus/helicobacter. Accessed 17 Aug 2021
McNulty CAM, Dent JC, Curry A, Uff JS, Ford GA, Gear MWL, Wilkinson SP (1989) New spiral bacterium in gastric mucosa. J Clin Pathol 42:585–591
Heilmann KL, Borchard F (1991) Gastritis due to spiral shaped bacteria other than Helicobacter pylori: Clinical, histological, and ultrastructural findings. Gut 32:137–140
Solnick JV, O’Rourke J, Lee A, Paster BJ, Dewhirst FE, Tompkins LS (1993) An uncultured gastric spiral organism is a newly identified Helicobacter in humans. J Infect Dis 168:379–385
Dewhirst FE, Shen Z, Scimeca MS, Stokes LN, Boumenna T, Chen T, Paster BJ, Fox JG (2005) Discordant 16S and 23S rRNA gene phylogenies for the genus Helicobacter: implications for phylogenetic inference and systematics. J Bacteriol 187:6106–6118
Baele M, Decostere A, Vandamme P, Ceelen L, Hellemans A, Mast J, Chiers K, Ducatelle R, Haesebrouck F (2008) Isolation and characterization of Helicobacter suis sp. nov. from pig stomachs. Int J Syst Evol Microbiol 58:1350–1358
Mendes EN, Queiroz DM, Rocha GA, Moura SB, Leite VHR, Fonseca MEF (1990) Ultrastructure of a spiral micro-organism from pig gastric mucosa ('Gastrospirillum suis’). J Med Microbiol 33:61–66
Queiroz DMM, Rocha GA, Mendes EN, Lage AP, Carvalho ACT, Barbosa AJA (1990) A spiral microorganism in the stomach of pigs. Vet Microbiol 24:199–204
De Groote D, Van Doorn LJ, Ducatelle R, Verschuuren A, Haesebrouck F, Quint WGV, Jalava K, Vandamme P (1999) “Candidatus Helicobacter suis”, a gastric Helicobacter from pigs, and its phylogenetic relatedness to other gastrospirilla. Int J Syst Bacteriol 49:1769–1777
O’Rourke JL, Solnick JV, Neilan BA, Seidel K, Hayter R, Hansen LM, Lee A (2004) Description of “Candidatus Helicobacter heilmannii” based on DNA sequence analysis of 16S rRNA and urease genes. Int J Syst Evol Microbiol 54:2203–2211
Dewhirst FE, Fox JG, On SLW (2000) Recommended minimal standards for describing new species of the genus Helicobacter. Int J Syst Evol Microbiol 50:2231–2237
Paster BJ, Lee A, Fox JG, Dewhirst FE, Tordoff LA, Fraser GJ, O’Rourke JL, Taylor NS, Ferrero R (1991) Phylogeny of Helicobacter felis sp. nov., Helicobacter mustelae, and related bacteria. Int J Syst Bacteriol 41:31–38
Hänninen ML, Happonen I, Saari S, Jalava K (1996) Culture and characteristics of Helicobacter bizzozeronii, a new canine gastric Helicobacter sp. Int J Syst Bacteriol 46:160–166
Jalava K, Kaartinen M, Utriainen M, Happonen I, Hänninen ML (1997) Helicobacter salomonis sp. nov., a canine gastric helicobacter sp. related to Helicobacter felis and Helicobacter bizzozeronii. Int J Syst Bacteriol 47:975–982
Van den Bulck K, Decostere A, Baele M, Vandamme P, Mast J, Ducatelle R, Haesebrouck F (2006) Helicobacter cynogastricus sp. nov., isolated from the canine gastric mucosa. Int J Syst Evol Microbiol 56:1559–1564
Baele M, Decostere A, Vandamme P, Van den Bulck K, Gruntar I, Mehle J, Mast J, Ducatelle R, Haesebrouck F (2008) Helicobacter baculiformis sp. nov., isolated from feline stomach mucosa. Int J Syst Evol Microbiol 58:357–364
Smet A, Flahou B, D’Herde K, Vandamme P, Cleenwerck I, Ducatelle R, Pasmans F, Haesebrouck F (2012) Helicobacter heilmannii sp. nov., isolated from feline gastric mucosa. Int J Syst Evol Microbiol 62:299–306
Joosten M, Lindén S, Rossi M, Tay ACY, Skoog E, Padra M, Peters F, Perkins T, Vandamme P, Van Nieuwerburgh F, D’Herde K, Van den Broeck W, Flahou B, Deforce D, Ducatelle R, Marshall B, Haesebrouck F, Smet A, Vann Nieuwerburgh F, D’Herde K, Van den Broeck W, Flahou B, Deforce D, Ducatelle R, Marshall B, Haesebrouck F, Smet A (2016) Divergence between the highly virulent zoonotic pathogen Helicobacter heilmannii and its closest relative, the low-virulence “Helicobacter ailurogastricus” sp. nov. Infect Immun 84:293–306
Haesebrouck F, Pasmans F, Flahou B, Smet A, Vandamme P, Ducatelle R (2011) Non-Helicobacter pylori Helicobacter species in the human gastric mucosa: a proposal to introduce the terms H. heilmannii sensu lato and sensu stricto. Helicobacter 16:339–340
Matos R, De Witte C, Smet A, Berlamont H, De Bruyckere S, Amorim I, Gärtner F, Haesebrouck F (2020) Antimicrobial susceptibility pattern of Helicobacter heilmannii and Helicobacter ailurogastricus isolates. Microorganisms 8:957
Berlamont H, Joosten M, Ducatelle R, Haesebrouck F, Smet A (2017) Presence of gastric Helicobacter spp. in feces and saliva from dogs and cats. Vlaams Diergeneeskd Tijdschr 86:73–78
Berlamont H, De Witte C, De Bruyckere S, Fox JG, Backert S, Smet A, Boyen F, Haesebrouck F (2021) Differentiation of gastric Helicobacter species using MALDI-TOF mass spectrometry. Pathogens 10:366
De Bruyne E, Flahou B, Chiers K, Meyns T, Kumar S, Vermoote M, Pasmans F, Millet S, Dewulf J, Haesebrouck F, Ducatelle R (2012) An experimental Helicobacter suis infection causes gastritis and reduced daily weight gain in pigs. Vet Microbiol 160:449–454
Hellemans A, Chiers K, Decostere A, De Bock M, Haesebrouck F, Ducatelle R (2007) Experimental infection of pigs with ‘Candidatus Helicobacter suis’. Vet Res Commun 31:385–395
Mendes EN, Queiroz DMM, Rocha GA, Nogueira AMMF, Carvalho ACT, Lage AP, Barbosa AJA (1991) Histopathological study of porcine gastric mucosa with and without a spiral bacterium ('Gastrospirillum suis’). J Med Microbiol 35:345–348
Grasso GM, Ripabelli G, Sammarco ML, Ruberto A, Iannitto G (1996) Prevalence of Helicobacter-like organisms in porcine gastric mucosa: a study of swine slaughtered in Italy. Comp Immunol Microbiol Infect Dis 19:213–217
Queiroz DMDM, Rocha GA, Mendes EN, De Moura SB, De Oliveira AMR, Miranda D (1996) Association between Helicobacter and gastric ulcer disease of the pars esophagea in swine. Gastroenterology 111:19–27
Roosendaal R, Vos JH, Roumen T, Van Vugt R, Cattoli G, Bart A, Klaasen HLBM, Kuipers EJ, Vandenbroucke-Grauls CMJE, Kusters JG (2000) Slaughter pigs are commonly infected by closely related but distinct gastric ulcerative lesion-inducing Gastrospirilla. J Clin Microbiol 38:2661–2664
Nurgaliev FM (2020) Detection of Helicobacter suis bacteria in pigs of different age groups. Vet Sci Today 4:266–271
Barbosa AJ, Silva JC, Nogueira AM, Paulino Júnior E, Miranda CR (1995) Higher incidence of Gastrospirillum sp. in swine with gastric ulcer of the pars oesophagea. Vet Pathol 32:134–139
De Witte C, Flahou B, Ducatelle R, Smet A, De Bruyne E, Cnockaert M, Taminiau B, Daube G, Vandamme P, Haesebrouck F (2017) Detection, isolation and characterization of Fusobacterium gastrosuis sp. nov. colonizing the stomach of pigs. Syst Appl Microbiol 40:42–50
De Witte C, Demeyere K, De Bruyckere S, Taminiau B, Daube G, Ducatelle R, Meyer E, Haesebrouck F (2019) Characterization of the non-glandular gastric region microbiota in Helicobacter suis-infected versus non-infected pigs identifies a potential role for Fusobacterium gastrosuis in gastric ulceration. Vet Res 50:39
De Witte C, Devriendt B, Flahou B, Bosschem I, Ducatelle R, Smet A, Haesebrouck F (2017) Helicobacter suis induces changes in gastric inflammation and acid secretion markers in pigs of different ages. Vet Res 48:34
Friendship RM (2004) Gastric ulceration in swine. J Swine Heal Prod 12:34–35
Nunes FC, Mateus TL, Teixeira S, Barradas P, de Witte C, Haesebrouck F, Amorim I, Gärtner F (2021) Presence of Helicobacter pylori and H. suis DNA in free-range wild boars. Animals 11:1269
Rimbara E, Suzuki M, Matsui H, Nakamura M, Morimoto M, Sasakawa C, Masuda H, Nomura S, Osaki T, Nagata N, Shibayama K, Tokunaga K (2021) Isolation and characterization of Helicobacter suis from human stomach. Proc Natl Acad Sci USA 118:e2026337118
Liang J, Ducatelle R, Pasmans F, Smet A, Haesebrouck F, Flahou B (2013) Multilocus sequence typing of the porcine and human gastric Pathogen Helicobacter suis. J Clin Microbiol 51:920–926
Hellemans A, Chiers K, Maes D, De Bock M, Decostere A, Haesebrouck F, Ducatelle R (2007) Prevalence of “Candidatus Helicobacter suis” in pigs of different ages. Vet Rec 161:189–192
Gustafsson A, Hultberg A, Sjöström R, Kacskovics I, Breimer ME, Borén T, Hammarström L, Holgersson J (2006) Carbohydrate-dependent inhibition of Helicobacter pylori colonization using porcine milk. Glycobiology 16:1–10
Su Y, Yao W, Perez-Gutierrez ON, Smidt H, Zhu WY (2008) Changes in abundance of Lactobacillus spp. and Streptococcus suis in the stomach, jejunum and ileum of piglets after weaning. FEMS Microbiol Ecol 66:546–555
Almeida LR, Costa PS, Nascimento AMA, de Reis M, P, Barros KO, Alvim LB, Nunes ÁC, Queiroz DMM, Rocha GA, Nicoli JR, de Moura SB, (2018) Porcine stomachs with and without gastric ulcer differ in Lactobacillus load and strain characteristics. Can J Microbiol 64:493–499
Proietti PC, Bietta A, Brachelente C, Lepri E, Davidson I, Franciosini MP (2010) Detection of Helicobacter spp. in gastric, fecal and saliva samples from swine affected by gastric ulceration. J Vet Sci 11:221–225
Inglis GD, McConville M, De Jong A (2006) Atypical Helicobacter canadensis strains associated with swine. Appl Environ Microbiol 72:4464–4471
Hänninen ML, Kärenlampi RI, Koort JMK, Mikkonen T, Björkroth KJ (2005) Extension of the species Helicobacter bilis to include the reference strains of Helicobacter sp. flexispira taxa 2, 3 and 8 and Finnish canine and feline flexispira strains. Int J Syst Evol Microbiol 55:891–898
De Witte C, Berlamont H, Haesebrouck F (2023) Helicobacter. In: Prescott JF, MacInnes JI, Van Immerseel F, et al (eds) Pathogenesis of Bacterial Infections in Animals, 5th edition. Willey-Blackwell, in press
Solnick JV, Schauer DB (2001) Emergence of diverse Helicobacter species in the pathogenesis of gastric and enterohepatic diseases. Clin Microbiol Rev 14:59–97
Bridgeford EC, Marini RP, Feng Y, Parry NMA, Rickman B, Fox JG (2008) Gastric Helicobacter species as a cause of feline gastric lymphoma: a viable hypothesis. Vet Immunol Immunopathol 123:106–113
Scanziani E, Simpson KW, Monestiroli S, Soldati S, Strauss-Ayali D, Del Piero F (2001) Histological and immunohistochemical detection of different Helicobacter species in the gastric mucosa of cats. J Vet Diagn Invest 13:3–12
Hermanns W, Kregel K, Breuer W, Lechner J (1995) Helicobacter-like organisms: histopathological examination of gastric biopsies from dogs and cats. J Comp Pathol 112:307–318
Kubota-Aizawa S, Ohno K, Kanemoto H, Nakashima K, Fukushima K, Uchida K, Chambers JK, Goto-Koshino Y, Mimuro H, Watanabe T, Sekizaki T, Tsujimoto H (2017) Epidemiological study on feline gastric Helicobacter spp. in Japan. J Vet Med Sci 79:876–880
Neiger R, Dieterich C, Burnens A, Waldvogel A, Corthésy-Theulaz I, Halter F, Lauterburg B, Schmassmann A (1998) Detection and prevalence of Helicobacter infection in pet cats. J Clin Microbiol 36:634–637
Norris CR, Marks SL, Eaton KA, Torabian SZ, Munn RJ, Solnick JV (1999) Healthy cats are commonly colonized with “Helicobacter heilmannii” that is associated with minimal gastritis. J Clin Microbiol 37:189–194
Guerra Segundo DD, Mello CBE, Cargnelutti JF, Flores MM, Pedrotti LF, Antunes BN, Milech V, Velasquez OG, Martins LR, Pinto Filho STL (2021) Evidence of Helicobacter spp. in saliva and gastric mucosa of domestic dogs in the central region of Rio Grande do Sul. Brazil. Vet Med Int 2021:8857231
Ekman E, Fredriksson M, Trowald-Wigh G (2013) Helicobacter spp. in the saliva, stomach, duodenum and faeces of colony dogs. Vet J 195:127–129
Polanco R, Salazar V, Reyes N, García-Amado MA, Michelangeli F, Contreras M (2011) Alta prevalencia de ADN de los helicobacteres no-H. pylori en la mucosa gástrica de perros domésticos venezolanos y sus alteraciones histopatológicas. Rev Inst Med Trop Sao Paulo 53:207–212
Wiinberg B, Spohr A, Dietz HH, Egelund T, Greiter-Wilke A, McDonough SP, Olsen J, Priestnall S, Chang YF, Simpson KW (2005) Quantitative analysis of inflammatory and immune responses in dogs with gastritis and their relationship to Helicobacter spp. infection. J Vet Intern Med 19:4–14
Amorim I, Smet A, Alves O, Teixeira S, Saraiva AL, Taulescu M, Reis C, Haesebrouck F, Gärtner F (2015) Presence and significance of Helicobacter spp. in the gastric mucosa of Portuguese dogs. Gut Pathog 7:12
Kubota-Aizawa S, Ohno K, Fukushima K, Kanemoto H, Nakashima K, Uchida K, Chambers JK, Goto-Koshino Y, Watanabe T, Sekizaki T, Mimuro H, Tsujimoto H (2017) Epidemiological study of gastric Helicobacter spp. in dogs with gastrointestinal disease in Japan and diversity of Helicobacter heilmannii sensu stricto. Vet J 225:56–62
Lee A, Krakowka S, Fox JG, Otto G, Eaton KA, Murphy JC (1992) Role of Helicobacter felis in chronic canine gastritis. Vet Pathol 29:487–494
Simpson KW, McDonough PL, Strauss-Ayali D, Chang YF, Harpending P, Valentine BA (1999) Helicobacter felis infection in dogs: effect on gastric structure and function. Vet Pathol 36:237–248
Simpson KW, Strauss-Ayali D, McDonough PL, Chang YF, Valentine BA (1999) Gastric function in dogs with naturally acquired gastric Helicobacter spp. infection. J Vet Intern Med 13:507–515
Eaton KA, Dewhirst FE, Paster BJ, Tzellas N, Coleman BE, Paola J, Sherding R (1996) Prevalence and varieties of Helicobacter species in dogs from random sources and pet dogs: animal and public health implications. J Clin Microbiol 34:3165–3170
Okubo BM, Ricci-Azevedo R, Zobiole NN, Buccini DF, Moreno SE (2017) Prevalence of Helicobacter spp. in dogs from Campo Grande-MS. Cienc Anim Bras 18:e17286
Van Den Bulck K, Decostere A, Baele M, Driessen A, Debongnie JC, Burette A, Stolte M, Ducatelle R, Haesebrouck F (2005) Identification of non-Helicobacter pylori spiral organisms in gastric samples from humans, dogs, and cats. J Clin Microbiol 43:2256–2260
Neiger R, Simpson KW (2000) Helicobacter infection in dogs and cats: facts and fiction. J Vet Intern Med 14:125–133
Jalava K, On SLW, Vandamme PAR, Happonen I, Sukura A, Hänninen ML (1998) Isolation and identification of Helicobacter spp. from canine and feline gastric mucosa. Appl Environ Microbiol 64:3998–4006
Neiger R, Tschudi ME, Burnens A, Göke B, Schmassmann A (1999) Diagnosis and identification of gastric Helicobacter species by polymerase chain reaction in dogs. Microb Ecol Health Dis 11:234–240
Lecoindre P, Chevallier M, Peyrol S, Boude M, Ferrero RL, Labigne A (2000) Gastric helicobacters in cats. J Feline Med Surg 2:19–27
Jankowski M, Spuzak J, Kubiak K, Glińska-Suchocka K, Biernat M (2016) Detection of Helicobacter spp. in the saliva of dogs with gastritis. Pol J Vet Sci 19:133–140
Craven M, Recordati C, Gualdi V, Pengo G, Luini M, Scanziani E, Simpson KW (2011) Evaluation of the Helicobacteraceae in the oral cavity of dogs. Am J Vet Res 72:1476–1481
Nakamura M, Øverby A, Michimae H, Matsui H, Takahashi S, Mabe K, Shimoyama T, Sasaki M, Terao S, Kamada T, Yanaka A, Iwamoto J, Tanabe S, Tari A, Nasu S, Suzuki H, Yamagata Murayama S (2020) PCR analysis and specific immunohistochemistry revealing a high prevalence of non-Helicobacter pylori helicobacters in Helicobacter pylori-negative gastric disease patients in Japan: high susceptibility to an Hp eradication regimen. Helicobacter 25:e12700
De Groote D, Van Doorn LJ, Van Den Bulck K, Vandamme P, Vieth M, Stolte M, Debongnie JC, Burette A, Haesebrouck F, Ducatelle R (2005) Detection of Non-pylori Helicobacter species in “Helicobacter heilmannii”-infected humans. Helicobacter 10:398–406
Liu J, He L, Haesebrouck F, Gong Y, Flahou B, Cao Q, Zhang J (2015) Prevalence of coinfection with gastric non-Helicobacter pylori Helicobacter (NHPH) species in Helicobacter pylori-infected patients suffering from gastric disease in Beijing, China. Helicobacter 20:284–290
Trebesius K, Adler K, Vieth M, Stolte M, Haas R (2001) Specific detection and prevalence of Helicobacter heilmannii-like organisms in the human gastric mucosa by fluorescent in situ hybridization and partial 16S ribosomal DNA sequencing. J Clin Microbiol 39:1510–1516
Yakoob J, Abbas Z, Khan R, Naz S, Ahmad Z, Islam M, Awan S, Jafri F, Jafri W (2012) Prevalence of non Helicobacter pylori species in patients presenting with dyspepsia. BMC Gastroenterol 12:3
Jalava K, On SLW, Harrington CS, Andersen LP, Hänninen ML, Vandamme P (2001) A cultured strain of “Helicobacter heilmannii”, a human gastric pathogen, identified as H. bizzozeronii: evidence for zoonotic potential of Helicobacter. Emerg Infect Dis 7:1036–1038
Kivistö R, Linros J, Rossi M, Rautelin H, Hänninen ML (2010) Characterization of multiple Helicobacter bizzozeronii isolates from a finnish patient with severe dyspeptic symptoms and chronic active gastritis. Helicobacter 15:58–66
Wüppenhorst N, von Loewenich F, Hobmaier B, Vetter-Knoll M, Mohadjer S, Kist M (2012) Culture of a gastric non-Helicobacter pylori Helicobacter from the stomach of a 14-year-old girl. Helicobacter 18:1–5
Ghysen K, Smet A, Denorme P, Vanneste G, Haesebrouck F, Van Moerkercke W (2018) An atypical presentation of an acute gastric Helicobacter felis infection. Acta Gastroenterol Belg 81:436–438
Rimbara E, Suzuki M, Matsui H, Nakamura M, Kobayashi H, Mori S, Shibayama K (2020) Complete genome sequence of Helicobacter suis strain SNTW101c, originally isolated from a patient with nodular gastritis. Microbiol Resour Announc 9:e01340-e1419
Stolte M, Wellens E, Bethke B, Ritter M, Eidt H (1994) Helicobacter heilmannii (formerly Gastrospirillum hominis) gastritis: an infection transmitted by animals? Scand J Gastroenterol 29:1061–1064
Dieterich C, Wiesel P, Neiger R, Blum A, Corthésy-Theulaz I (1998) Presence of multiple “Helicobacter heilmannii” strains in an individual suffering from ulcers and in his two cats. J Clin Microbiol 36:1366–1370
van Loon S, Bart A, den Hertog E, Nikkels P, Houwen R, De Schryver J, Oudshoorn J (2003) Helicobacter heilmannii gastritis caused by cat to child transmission. J Pediatr Gastroenterol Nutr 36:407–409
Thomson MA, Greer R, Cleghorn GJ, Storey P (1994) Canine-human transmission of Gastrospirillum hominis. Lancet 343:1605–1607
Meining A, Kroher G, Stolte M (2009) Animal reservoirs in the transmission of Helicobacter heilmannii: results of a questionnaire-based study. Scand J Gastroenterol 33:795–798
Joosten M, Flahou B, Meyns T, Smet A, Arts J, De Cooman L, Pasmans F, Ducatelle R, Haesebrouck F (2013) Case report: Helicobacter suis infection in a pig veterinarian. Helicobacter 18:392–396
De Cooman L, Houf K, Smet A, Flahou B, Ducatelle R, De Bruyne E, Pasmans F, Haesebrouck F (2014) Presence of Helicobacter suis on pork carcasses. Int J Food Microbiol 187:73–76
Matsumoto T, Kawakubo M, Akamatsu T, Koide N, Ogiwara N, Kubota S, Sugano M, Kawakami Y, Katsuyama T, Ota H (2014) Helicobacter heilmannii sensu stricto-related gastric ulcers: a case report. World J Gastroenterol 20:3376–3382
De Cooman L, Flahou B, Houf K, Smet A, Ducatelle R, Pasmans F, Haesebrouck F (2013) Survival of Helicobacter suis bacteria in retail pig meat. Int J Food Microbiol 166:164–167
Nunes FC, Mateus TL, Teixeira S, Barradas PF, Gärtner F, Haesebrouck F, Amorim I (2021) Molecular detection of human pathogenic gastric Helicobacter Species in wild rabbits (Oryctolagus cuniculus) and wild quails (Coturnix coturnix). Zoonotic Dis 1:42–50
Debongnie J, Donnay M, Mairesse J, Lamy V, Dekoninck X, Ramdani B (1998) Gastric ulcers and Helicobacter heilmannii. Eur J Gastroenterol Hepatol 10:251–254
Morgner A, Lehn N, Andersen LP, Thiede C, Bennedsen M, Trebesius K, Neubauer B, Neubauer A, Stolte M, Bayerdörffer E (2000) Helicobacter heilmannii–associated primary gastric low-grade MALT lymphoma: complete remission after curing the infection. Gastroenterology 118:821–828
Okiyama Y, Matsuzawa K, Hidaka E, Sano K, Akamatsu T, Ota H (2005) Helicobacter heilmannii infection: clinical, endoscopic and histopathological features in Japanese patients. Pathol Int 55:398–404
O’Rourke JL, Dixon MF, Jack A, Enno A, Lee A (2004) Gastric B-cell mucosa-associated lymphoid tissue (MALT) lymphoma in an animal model of “Helicobacter heilmannii” infection. J Pathol 203:896–903
Øverby A, Murayama SY, Michimae H, Suzuki H, Suzuki M, Serizawa H, Tamura R, Nakamura S, Takahashi S, Nakamura M (2017) Prevalence of gastric Non-Helicobacter pylori-helicobacters in Japanese Patients with gastric disease. Digestion 95:61–66
Tsukadaira T, Hayashi S, Ota H, Kobayashi N, Sekiguchi Y, Kodaira H, Matsumoto T, Horiuchi K, Negishi T, Kurahashi M (2021) Acute gastric mucosal lesions caused by acute infection of non-Helicobacter pylori Helicobacter: a case report. Helicobacter 26:e12814
Stolte M, Kroher G, Meining A, Morgner A, Bayerdörffer E, Bethke B (2009) A comparison of Helicobacter pylori and H. heilmannii gastritis: a matched control study involving 404 patients. Scand J Gastroenterol 32:28–33
Joo M, Ji EK, Sun HC, Kim H, Chi JG, Kim KA, Jeon HY, June SL, Moon YS, Kim KM (2007) Helicobacter heilmannii-associated gastritis: clinicopathologic findings and comparison with Helicobacter pylori-associated gastritis. J Korean Med Sci 22:63–69
Tsukadaira T, Hayashi S, Ota H, Kobayashi N, Sekiguchi Y, Kodaira H, Matsumoto T, Horiuchi K, Negishi T, Kurahashi M (2021) Prevalence, clinical features, and esophagogastroduodenoscopy (EGD) findings of non-Helicobacter pylori Helicobacter infection: a study of 50 cases at a single facility in Japan. Helicobacter 26:e12811
Bahadori A, De Witte C, Agin M, De Bruyckere S, Smet A, Tümgör G, Güven Gökmen T, Haesebrouck F, Köksal F (2018) Presence of gastric Helicobacter species in children suffering from gastric disorders in Southern Turkey. Helicobacter 23:e12511
Liu NY, Sun JH, Jiang XF, Li H (2021) Helicobacter pylori infection and risk for developing dementia: an evidence-based meta-analysis of case-control and cohort studies. Aging (Albany NY) 13:22571–22587
Dobbs SM, Dobbs RJ, Weller C, Charlett A (2000) Link between Helicobacter pylori infection and idiopathic parkinsonism. Med Hypotheses 55:93–98
Gorlé N, Bauwens E, Haesebrouck F, Smet A, Vandenbroucke RE (2021) Helicobacter and the potential role in neurological disorders: there is more than Helicobacter pylori. Front Immunol 11:584165
Shen X, Yang H, Wu Y, Zhang D, Jiang H (2017) Meta-analysis: association of Helicobacter pylori infection with Parkinson’s diseases. Helicobacter 22:e12398
Augustin AD, Savio A, Nevel A, Ellis RJ, Weller C, Taylor D, Tucker RM, Ibrahim MAA, Bjarnason I, Dobbs SM, Dobbs RJ, Charlett A (2019) Helicobacter suis is associated with mortality in Parkinson’s disease. Front Med 6:188
Peng X, Zhou L, Gong Y, Song Z, He L, Lin S, Zhang J (2017) Non-pylori helicobacters (NHPHs) induce shifts in gastric microbiota in Helicobacter pylori-infected patients. Front Microbiol 8:1038
Nakagawa S, Shimoyama T, Nakamura M, Chiba D, Kikuchi H, Sawaya M, Chinda D, Mikami T, Fukuda S (2018) The resolution of Helicobacter suis-associated gastric lesions after eradication therapy. Intern Med 57:203–207
Okamura T, Iwaya Y, Yokosawa S, Suga T, Arakura N, Matsumoto T, Ogiwara N, Higuchi K, Ota H, Tanaka E (2013) A case of Helicobacter heilmannii-associated primary gastric mucosa-associated lymphoid tissue lymphoma achieving complete remission after eradication. Clin J Gastroenterol 6:38–45
Stolte M, Eidt S (1993) Healing gastric MALT lymphomas by eradicating H pylori? Lancet 342:571–575
Wotherspoon AC, Doglioni C, Diss TC, Pan L, Moschini A, de Boni M, Isaacson PG (1993) Regression of primary low-grade B-cell gastric lymphoma of mucosa-associated lymphoid tissue type after eradication of Helicobacter pylori. Lancet 342:575–577
Masaki S, Takigawa H, Kunisho S, Kimoto A, Yuge R, Tanaka S, Kitadai Y (2020) Association between Helicobacter suis infection and the pathogenesis of gastric mucosa-associated lymphoid tissue lymphoma. Hiroshima J Med Sci 69:33–37
Shiratori S, Mabe K, Yoshii S, Takakuwa Y, Sato M, Nakamura M, Kudo T, Kato M, Asaka M, Sakamoto N (2016) Two cases of chronic gastritis with non-Helicobacter pylori Helicobacter infection. Intern Med 55:1865–1869
Hellemans A, Decostere A, Haesebrouck F, Ducatelle R (2005) Evaluation of antibiotic treatment against “Candidatus Helicobacter suis” in a mouse model. Antimicrob Agents Chemother 49:4530–4535
Takigawa H, Masaki S, Naito T, Yuge R, Urabe Y, Tanaka S, Sentani K, Matsuo T, Matsuo K, Chayama K, Kitadai Y (2019) Helicobacter suis infection is associated with nodular gastritis-like appearance of gastric mucosa-associated lymphoid tissue lymphoma. Cancer Med 8:4370–4379
Berlamont H, Smet A, De Bruyckere S, Boyen F, Ducatelle R, Haesebrouck F, De Witte C (2019) Antimicrobial susceptibility pattern of Helicobacter suis isolates from pigs and macaques. Vet Microbiol 239:108459
Van den Bulck K, Decostere A, Gruntar I, Baele M, Krt B, Ducatelle R, Haesebrouck F (2005) In vitro antimicrobial susceptibility testing of Helicobacter felis, H. bizzozeronii, and H. salomonis. Antimicrob Agents Chemother 49:2997–3000
Vermoote M, Pasmans F, Flahou B, Van Deun K, Ducatelle R, Haesebrouck F (2011) Antimicrobial susceptibility pattern of Helicobacter suis strains. Vet Microbiol 153:339–342
Flahou B, Haesebrouck F, Pasmans F, D’Herde K, Driessen A, van Deun K, Smet A, Duchateau L, Chiers K, Ducatelle R (2010) Helicobacter suis causes severe gastric pathology in mouse and mongolian gerbil models of human gastric disease. PLoS One 5:e14083
Flahou B, Van DK, Pasmans F, Smet A, Volf J, Rychlik I, Ducatelle R, Haesebrouck F (2012) The local immune response of mice after Helicobacter suis infection: strain differences and distinction with Helicobacter pylori. Vet Res 43:75
Greiner A, Knörr C, Qin Y, Sebald W, Schimpl A, Banchereau J, Müller-Hermelink HK (1997) Low-grade B cell lymphomas of mucosa-associated lymphoid tissue (MALT-type) require CD40-mediated signaling and Th2-type cytokines for in vitro growth and differentiation. Am J Pathol 150:1583–1593
Bosschem I, Flahou B, Bakker J, Heuvelman E, Langermans JAM, De Bruyne E, Joosten M, Smet A, Ducatelle R, Haesebrouck F (2017) Comparative virulence of in vitro-cultured primate- and pig-associated Helicobacter suis strains in a BALB/c mouse and a Mongolian gerbil model. Helicobacter 22:e12349
Bosschem I, Flahou B, Van Deun K, De Koker S, Volf J, Smet A, Ducatelle R, Devriendt B, Haesebrouck F (2017) Species-specific immunity to Helicobacter suis. Helicobacter 22:e12375
Péré-Védrenne C, Flahou B, Loke MF, Ménard A, Vadivelu J (2017) Other helicobacters, gastric and gut microbiota. Helicobacter 22:e12407
Lemos LMS, Miyajima F, Castilho GRC, Martins DTO, Pritchard DM, Burkitt MD (2017) Hexane extracts of Calophyllum brasiliense inhibit the development of gastric preneoplasia in Helicobacter felis infected INS-Gas mice. Front Pharmacol 8:92
Kim D-J, Kang M-J, Choi J-A, Na D-S, Kim J-B, Na C-S, Park J-H (2016) Anti-Helicobacter pylori activity of crude N-acetylneuraminic acid isolated from glycomacropeptide of whey. Lab Anim Res 32:99–104
Yang L, Bin Tian Z, Yu YN, Zhang CP, Li XY, Mao T, Jing X, Zhao WJ, Ding XL, Yang RM, Zhang SQ (2017) Saccharomyces boulardii administration can inhibit the formation of gastric lymphoid follicles induced by Helicobacter suis infection. Pathog Dis 75:ftx006
Matsui H, Takahashi T, Øverby A, Murayama SY, Yoshida H, Yamamoto Y, Nishiyama K, Seto Y, Takahashi T, Mukai T, Nakamura M (2015) Mouse models for assessing the protective efficacy of Lactobacillus gasseri SBT2055 against Helicobacter suis infection associated with the development of gastric mucosa-associated lymphoid tissue lymphoma. Helicobacter 20:291–298
Bosschem I, Bayry J, De Bruyne E, Van Deun K, Smet A, Vercauteren G, Ducatelle R, Haesebrouck F, Flahou B (2015) Effect of different adjuvants on protection and side-effects induced by Helicobacter suis whole-cell lysate vaccination. PLoS One 10:e0131364
Flahou B, Hellemans A, Meyns T, Duchateau L, Chiers K, Baele M, Pasmans F, Haesebrouck F, Ducatelle R (2009) Protective immunization with homologous and heterologous antigens against Helicobacter suis challenge in a mouse model. Vaccine 27:1416–1421
Sutton P, Danon SJ, Walker M, Thompson LJ, Wilson J, Kosaka T, Lee A (2001) Post-immunisation gastritis and Helicobacter infection in the mouse: a long term study. Gut 49:467–473
Sutton P, O’Rourke J, Wilson J, Dixon MF, Lee A (2004) Immunisation against Helicobacter felis infection protects against the development of gastric MALT lymphoma. Vaccine 22:2541–2546
Berlamont H, Bruggeman A, Bauwens E, Vandendriessche C, Clarebout E, Xie J, De Bruyckere S, Van Imschoot G, Van Wonterghem E, Ducatelle R, Santens P, Smet A, Haesebrouck F, Vandenbroucke RE (2021) Gastric Helicobacter suis infection partially protects against neurotoxicity in a 6-OHDA Parkinson’s disease mouse model. Int J Mol Sci 22:11328
Labigne A, Cussac V, Courcoux P (1991) Shuttle cloning and nucleotide sequences of Helicobacter pylori genes responsible for urease activity. J Bacteriol 173:1920–1931
Mobley HLT, Garner RM, Bauerfeind P (1995) Helicobacter pylori nickel-transport gene nixA: synthesis of catalytically active urease in Escherichia coli independent of growth conditions. Mol Microbiol 16:97–109
Skouloubris S, Thiberge J, Labigne A, De Reuse H (1998) The Helicobacter pylori UreI protein is not involved in urease activity but is essential for bacterial survival in vivo. Infect Immun 66:4517–4521
Eaton K, Krakowka S (1994) Effect of gastric pH on urease-dependent colonization of gnotobiotic piglets by Helicobacter pylori. Infect Immun 62:3604–3607
Eaton K, Gilbert J, Joyce E, Wanken A, Thevenot T, Baker P, Plaut A, Wright A (2002) In vivo complementation of ureB restores the ability of Helicobacter pylori to colonize. Infect Immun 70:771–778
Sycuro LK, Wyckoff TJ, Biboy J, Born P, Pincus Z, Vollmer W, Salama NR (2012) Multiple peptidoglycan modification networks modulate Helicobacter pylori’s cell shape, motility, and colonization potential. PLoS Pathog 8:e1002603
Constantino MA, Jabbarzadeh M, Fu HC, Shen Z, Fox JG, Haesebrouck F, Linden SK, Bansil R (2018) Bipolar lophotrichous Helicobacter suis combine extended and wrapped flagella bundles to exhibit multiple modes of motility. Sci Rep 8:1–15
Aihara E, Closson C, Matthis AL, Schumacher MA, Engevik AC, Zavros Y, Ottemann KM, Montrose MH (2014) Motility and chemotaxis mediate the preferential colonization of gastric injury sites by Helicobacter pylori. PLoS Pathog 10:e1004275
Bassler BL (1999) How bacteria talk to each other: regulation of gene expression by quorum sensing. Curr Opin Microbiol 2:582–587
Lindén SK, Sheng YH, Every AL, Miles KM, Skoog EC, Florin THJ, Sutton P, McGuckin MA (2009) MUC1 limits Helicobacter pylori infection both by steric hindrance and by acting as a releasable decoy. PLoS Pathog 5:e1000617
Matos R, Amorim I, Magalhães A, Haesebrouck F, Gärtner F, Reis CA (2021) Adhesion of Helicobacter species to the human gastric mucosa: a deep look into glycans role. Front Mol Biosci 8:656439
Liu C, Smet A, Blaecher C, Flahou B, Ducatelle R, Linden S, Haesebrouck F (2014) Gastric De Novo Muc13 expression and spasmolytic polypeptide-expressing metaplasia during Helicobacter heilmannii infection. Infect Immun 82:3227–3239
Shimamura T, Ito H, Shibahara J, Watanabe A, Hippo Y, Taniguchi H, Chen Y, Kashima T, Ohtomo T, Tanioka F, Iwanari H, Kodama T, Kazui T, Sugimura H, Fukayama M, Aburatani H (2005) Overexpression of MUC13 is associated with intestinal-type gastric cancer. Cancer Sci 96:265–273
Padra M, Adamczyk B, Flahou B, Erhardsson M, Chahal G, Smet A, Jin C, Thorell A, Ducatelle R, Haesebrouck F, Karlsson NG, Lindén SK (2019) Helicobacter suis infection alters glycosylation and decreases the pathogen growth inhibiting effect and binding avidity of gastric mucins. Mucosal Immunol 12:784–794
Padra M, Adamczyk B, Benktander J, Flahou B, Skoog EC, Padra JT, Smet A, Jin C, Ducatelle R, Samuelsson T, Haesebrouck F, Karlsson NG, Teneberg S, Lindén SK (2018) Helicobacter suis binding to carbohydrates on human and porcine gastric mucins and glycolipids occurs via two modes. Virulence 9:898–918
Amorim I, Freitas DP, Magalhães A, Faria F, Lopes C, Faustino AM, Smet A, Haesebrouck F, Reis CA, Gärtner F (2014) A comparison of Helicobacter pylori and non-Helicobacter pylori Helicobacter spp. binding to canine gastric mucosa with defined gastric glycophenotype. Helicobacter 19:249–259
Vermoote M, Vandekerckhove TT, Flahou B, Pasmans F, Smet A, De Groote D, Van Criekinge W, Ducatelle R, Haesebrouck F (2011) Genome sequence of Helicobacter suis supports its role in gastric pathology. Vet Res 42:51
Schott T, Kondadi PK, Hänninen ML, Rossi M (2011) Comparative genomics of Helicobacter pylori and the human-derived Helicobacter bizzozeronii CIII-1 strain reveal the molecular basis of the zoonotic nature of non-pylori gastric Helicobacter infections in humans. BMC Genomics 12:534
Bauwens E, Joosten M, Taganna J, Rossi M, Debraekeleer A, Tay A, Peters F, Backert S, Fox J, Ducatelle R, Remaut H, Haesebrouck F, Smet A (2018) In silico proteomic and phylogenetic analysis of the outer membrane protein repertoire of gastric Helicobacter species. Sci Rep 8:15453
Cheng L, Rossi M, Lindén S, Padra M, Blaecher C, Bauwens E, Joosten M, Flahou B, Van den Broeck W, Ducatelle R, Haesebrouck F, Smet A (2016) The Helicobacter heilmannii hofE and hofF genes are essential for colonization of the gastric mucosa and play a role in IL-1β-induced gastric MUC13 expression. Helicobacter 21:504–522
De Bruyne E, Ducatelle R, Foss D, Sanchez M, Joosten M, Zhang G, Smet A, Pasmans F, Haesebrouck F, Flahou B (2016) Oral glutathione supplementation drastically reduces Helicobacter-induced gastric pathologies. Sci Rep 6:20169
Flahou B, Haesebrouck F, Chiers K, Van Deun K, De Smet L, Devreese B, Vandenberghe I, Favoreel H, Smet A, Pasmans F, D’Herde K, Ducatelle R (2011) Gastric epithelial cell death caused by Helicobacter suis and Helicobacter pylori γ-glutamyl transpeptidase is mainly glutathione degradation-dependent. Cell Microbiol 13:1933–1955
Arnold IC, Zigova Z, Holden M, Lawley TD, Rad R, Dougan G, Falkow S, Bentley SD, Müller A (2011) Comparative whole genome sequence analysis of the carcinogenic bacterial model pathogen Helicobacter felis. Genome Biol Evol 3:302–308
Smet A, Van NF, Ledesma J, Flahou B, Deforce D, Ducatelle R, Haesebrouck F (2013) Genome sequence of Helicobacter heilmannii sensu stricto ASB1 Isolated from the gastric mucosa of a kitten with severe gastritis. Genome Announc 1:e00033-e112
Käbisch R, Semper RP, Wüstner S, Gerhard M, Mejías-Luque R (2016) Helicobacter pylori γ-glutamyltranspeptidase induces tolerogenic human dendritic cells by activation of glutamate receptors. J Immunol 196:4246–4252
Beswick EJ, Pinchuk IV, Minch K, Suarez G, Sierra JC, Yamaoka Y, Reyes VE (2006) The Helicobacter pylori urease B subunit binds to CD74 on gastric epithelial cells and induces NF-kappaB activation and interleukin-8 production. Infect Immun 74:1148–1155
Zhang G, Ducatelle R, Pasmans F, D’Herde K, Huang L, Smet A, Haesebrouck F, Flahou B (2013) Effects of Helicobacter suis γ-glutamyl transpeptidase on lymphocytes: modulation by glutamine and glutathione supplementation and outer membrane vesicles as a putative delivery route of the enzyme. PLoS One 8:e77966
Shibayama K, Takeuchi H, Wachino J, ichi, Mori S, Arakawa Y, (2011) Biochemical and pathophysiological characterization of Helicobacter pylori asparaginase. Microbiol Immunol 55:408–417
Acknowledgements
Research on Helicobacter was supported by the Research Fund of Ghent University, Belgium and the Portuguese Foundation for Science and Technology.
Author information
Authors and Affiliations
Contributions
All of the authors meet the ICMJE authorship criteria with the following contributions specified: ET conducted most of the literature search and contributed a major portion to the writing of the manuscript. FH designed the literature review, contributed in writing the manuscript and supervised the paper. KC, IA, FG, DM and CVS reviewed and supervised the paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Taillieu, E., Chiers, K., Amorim, I. et al. Gastric Helicobacter species associated with dogs, cats and pigs: significance for public and animal health. Vet Res 53, 42 (2022). https://doi.org/10.1186/s13567-022-01059-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-022-01059-4