
Abstract
Background
Culicoides midges have been well-studied in Spain, particularly over the last 20 years, mainly because of their role as vectors of arboviral diseases that affect livestock. Most studies on Culicoides are conducted using suction light traps in farmed environments, but studies employing alternative trapping techniques or focusing on natural habitats are scarce.
Methods
In the present study, we analyze Culicoides captured in 2023 at 476 sites in western Andalusia (southern Spain) using carbon dioxide-baited Biogents (BG)-sentinel traps across different ecosystems.
Results
We collected 3,084 Culicoides midges (3060 females and 24 males) belonging to 23 species, including the new species Culicoides grandifovea sp. nov. and the first record of Culicoides pseudolangeroni for Europe. Both species were described with morphological and molecular methods and detailed data on spatial distribution was also recorded. The new species showed close phylogenetic relations with sequences from an unidentified Culicoides from Morocco (92.6% similarity) and with Culicoides kurensis. Culicoides imicola was the most abundant species (17.4%), followed by Culicoides grandifovea sp. nov. (14.6%) and Culicoides kurensis (11.9%). Interestingly, Culicoides montanus was the only species of the obsoletus and pulicaris species complexes captured, representing the first record of this species in southern Spain. A total of 53 valid Culicoides species have been reported in the area, with 48 already reported in literature records and 5 more added in the present study. Information on the flight period for the most common Culicoides species is also provided.
Conclusions
To the best of our knowledge, our study represents the most comprehensive effort ever done on nonfarmland habitats using carbon-dioxide baited suction traps for collecting Culicoides. Our data suggests that using carbon dioxide traps offers a completely different perspective on Culicoides communities compared with routinely used light traps, including the discovery of previously unrecorded species.
Graphical Abstract

Similar content being viewed by others


Background
Culicoides is a taxonomically diverse genus of tiny hematophagous insects belonging to the family Ceratopogonidae. The number of Culicoides species has increased over the last few years in Europe, particularly those belonging to the subgenus Culicoides, due to the rise of molecular approaches [1,2,3,4,5]. Culicoides are vectors of arboviruses of the Orbivirus genus, such as the African horse sickness virus (AHSV), bluetongue virus (BTV), and Schmallenberg virus (SV) [6,7,8,9]. The recent outbreaks of epizootic hemorrhagic disease (EHD) in Spain, a virus transmitted by Culicoides, which primarily affects cervids and livestock [10, 11], has renewed the interest in this group. Furthermore, Culicoides are also vectors of parasites infecting nonmammal hosts, including the avian malaria-like parasites of the genus Haemoproteus [12].
Due to their minute size, the identification of Culicoides species across Europe has remained challenging. Morphological diagnostic characters that are commonly used for identification are often difficult to observe. Wing spot patterns are of primary importance in species diagnosis [13,14,15]. However, some Culicoides species bear faint spots or lack a defined wing pattern resulting in clear wings without markings, these being routinely grouped as “other Culicoides species” in large faunistic studies [16, 17]. Considering that about 25% of the European Culicoides are faint or unspotted species, the study of Culicoides communities requires mounting specimens in slides. This task requires skills and is laborious, time-consuming, and impracticable when a large number needs to be identified and may be incompatible with the preservation of specimens for pathogen surveillance. Many studies published in Europe only focused on species with wing patterns, which usually correspond to vectors involved in epizootics (subgenus Culicoides and Avaritia) [18,19,20]. However, the identification of nontarget Culicoides fauna should also be undertaken, not only to improve the faunistic inventories, but also for a better characterization of other unknown potential vectors that might arise in future epizootics [21].
Suction light traps, particularly the commercially available Onderstepoort Veterinary Institute (OVI) trap and the ultraviolet (UV)–Center for Disease Control (CDC) downdraft suction trap are the most commonly used traps for the collection of Culicoides [22]. These traps are routinely chosen for their ease of installation, provision of standardized data among studies, and ability to collect a reasonable numbers of vector species when present [22]. However, as shown in other insect groups, using alternative approaches to sample Culicoides may provide new opportunities to collect species attracted to other stimuli [22, 23].
Since information on the composition and distribution of Culicoides species is a prerequisite to understand the epidemiology of Culicoides-borne pathogens, surveillance contributes to the development of effective strategies for disease prevention and control. In Spain, until 2012, 81 Culicoides species were recorded [24], and in the subsequent 12 years, to our knowledge, 5 more species were added [2, 25,26,27]. However, current information of the Culicoides fauna differs between regions, with southern Spain being comparatively understudied in spite that AHSV [28, 29], BTV [30] and EHD [11] outbreaks occurred in the area in 1956–1960 and 2004–2024. This area has been severely affected by West Nile virus outbreaks in recent years [31], and a large effort is being done for the characterization of mosquito communities across the territory. In addition to mosquitoes, Culicoides biting midges are often captured in these traps. Here, we conducted an extensive monitoring of Culicoides in several diverse environments using an alternative sampling method to improve the knowledge of the Culicoides distribution in the area. In addition, we carried out a bibliographic review of the Culicoides species recorded in southern Spain.
Methods
Study area, design, and trapping
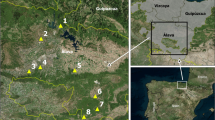
The study was conducted in the provinces of Huelva, Sevilla, Málaga, Córdoba, and Cádiz of the Andalusia region (southern Spain) (Fig. 1). This region is characterized by a Mediterranean climate with mild winters with irregular precipitations and dry, hot, and sunny summers. Some areas experience the hottest temperatures in the country during summer (> 45 °C). In 2023, when this study was conducted, the average year-round temperature was approximately 19 °C, with more than 290 days of sunshine. January was the coldest month and August the hottest.

Map of the 476 sampling sites in the five western provinces of Andalusia (southern Spain). Empty circles: negative sampling sites for the presence of Culicoides. Red circles: sampling sites with captures of Culicoides. Yellow starts: sampling sites with captures of Culicoides pseudolangeroni. Blue triangles: sampling sites with captures of Culicoides grandifovea sp. nov. The map was created using QGIS software (QGIS version 3.32)
Culicoides biting midges were collected using two different surveys. In the first survey (first study hereinafter), a total of 450 sampling sites were sampled three times (spring, summer, and autumn) from April to November 2023. The total area sampled covered 31,500 km2 across the provinces of Sevilla, Cádiz, and Huelva, resulting in an overall trapping effort of approximately 1350 trapping days (Fig. 1). To ensure systematic coverage, the area was divided into grids (5 km × 5 km) with one sampling point selected in each grid. The sampling points and routes were determined at the beginning of the study to design fieldwork tracks that would maximize the number of traps surveyed each day. The order of sampling of these predefined tracks was randomly established to prevent associations between time and land-use geography. The second survey (second study) corresponds to the Junta de Andalusia mosquito survey program, with 26 sampling sites: eight in Sevilla and eight in Cádiz sampled at weekly intervals and six in Córdoba and four in Málaga sampled every 2 weeks, in both cases from 15 June to 23 November 2023 (i.e., 436 trapping days) (Fig. 1).
In all cases, insects were sampled using the commercially available BG-Sentinel model 2 trap (Biogents, Regensburg, Germany). These traps were baited with approximately 1.2 kg of dry ice in a polystyrene box to generate a continuous flow of carbon dioxide at the entrance of the trap, placed on the ground and were operated for 24 h. Traps were set in shaded and sheltered areas to avoid direct sunlight and wind, to increase captures and reduced the risk of vandalism. Samples were preserved in dry ice while transported to the laboratory and subsequently stored at −80 °C.
Morphological identification and molecular analysis
Frozen insects were immediately separated into groups on a chill table (BioQuip, Rancho Dominguez, CA) under the stereomicroscope. Culicoides midges were sorted out by sex and feeding status, and then were separated into species with distinctive wing patterns (i.e., Culicoides jamaicensis, Culicoides newsteadi, Culicoides imicola, Culicoides circumscriptus, etc.) and those with plain wings. Specimens with unspotted wings and/or unknown wing-patterned species were further examined on the basis of other traits such as body size, color, thorax pattern, wing features, and palpi. A subset of 285 distinctive Culicoides specimens was then individually dissected into different body parts (head, thorax, wings, and abdomen), mounted with Hoyer's medium on glass slides using needles (0.5 mm diameter) and dried at room temperature for 7 days. Key diagnostic structures of specimens were examined using a composed optical microscopy, employing a combination of three identification keys [13, 14, 32].
Length of every palpus and flagellar segments, wing length (from basal arculus to wing tip) and wing width (from R2 to vein Cu1), and spermathecae were photographed and measured under the optical microscope (Zeiss, Axioscope, UK) with a digital camera (Axioram 208 model) for the unsubscribed species and the new record Culicoides species (to complete the original descriptions). Measurements of the different parts of the specimens were performed using the Zeiss analyzing software. The area of the sensorial pit in the third palpus was also calculated with the “area calculator tool” in the new Culicoides species identified in this study. Antennal ratio (AR) was calculated as XI–XV antennal segments divided with segments III–X, and the Palpal ratio (PR) was calculated as length of segment III divided with the greatest breadth of the segment III.
The barcoding region of specimens of C. pseudolangeroni (n = 5) and C. grandifovea sp. nov. (n = 6) together with other plain-winged sibling species (specifically Culicoides indistinctus and Culicoides kurensis) (n = 4) were molecularly characterized. The head of each specimen was slide-mounted as previously described whereas the rest of the body was used for molecular analyses. Genomic DNA was extracted from each sample using the Maxwell®16 LEV Blood and Tissue DNA kit following the manufacturer's protocol. A 658 bp fragment of the cytochrome Oxidase Subunit I (COI) gene was amplified and sequenced following Folmer et al. [33]. The presence of amplicons was verified on 1.5% agarose gels. The amplified products were sequenced on both strands using Capillary Electrophoresis Sequencing by UCM (Madrid, Spain), and a consensus sequence was generated using Geneious v.2020.0.3 [34]. Species-level identity was determined with a threshold of > 99% identity score using BLASTn (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Nucleotide sequences generated during this study were deposited in the DNA Data Bank of Japan (DDBJ: https://www.ddbj.nig.ac.jp/index-e.html).
Phylogenetic analyses
A total of 24 Culicoides species (55 sequences) were included in the phylogenetic analysis. A sequence of Atrichopogon levis (GU804122) was used as outgroup. Phylogenetic reconstructions were conducted using the maximum likelihood optimization criterion, employing the GTR + F + I + G4 model as defined by IQ-TREE [35] and model selection was based on Akaike information criterion. The robustness of the resulting ML trees was evaluated using SH-aLRT (Shimodaira–Hasegawa-like approximate likelihood ratio test) and 1000 bootstraps. The tree was visualized using FigTree v1.4.2 (http://tree.bio.ed.ac.uk/software/figtree/). Finally, neighbor–net networks (NNn) were constructed using distance matrices corrected with the Kimura two-parameter model [36].
Literature review
A checklist of Culicoides species reported in southern Spain was compiled through a systematic review following the guidelines outlined by Haddaway et al. [37]. The bibliographic investigation involved using various databases of scientific and public journals such Web of Science, Google Scholar, Scopus, Dialnet, PubMed, Redalyc, SciELO, BioOne, ScienceDirect, ResearchGate, and REDIB to identify articles released from 1900 to 2023. Our research included keywords in English and Spanish of the following combination of terms in the title, abstract and keywords: “Culicoides” OR “jejenes” OR “Ceratopogonidae” AND “Spain” as well as variations combining these keywords with specific regions such as “Andalucía,” “Andalusia,” “Iberia,” “Iberian,” and “Peninsula.” Combinations such as “Culicoides” AND “South Spain” and similar pairs with “jejenes” and “Ceratopogonidae” were also used. Additional articles were obtained from references from the articles reviewed. Duplicated publications and works that do not explicitly mention the Culicoides species or place of capture, were removed from the review. A total of 24 publications were included in the present study.
Results
At least 48 species of Culicoides had been cited in Andalusia in studies published between 1900 and 2023 (Table 1). In our study, we collected 3,084 Culicoides specimens (3060 females and 24 males) in both sampling surveys (n = 2644 in the first one and 440 in the second one), representing 23 valid Culicoides species, four Culicoides variations and five undetermined taxa (Table 2). Our sampling includes, at least, five new records (Table 1–2) for Andalusia, including Culicoides montanus within the Obsoletus group, therefore totalizing 53 valid species for this region. This study includes a new species hereinafter referred to as C. grandifovea sp. nov. and a new record for Europe (C. pseudolangeroni). Culicoides jumineri near bahrainensis was also the first record for Spain, but it is not included as valid species. With these two new records, 88 Culicoides species have been reported in Spain, with about 60% of them present in Andalusia. Our study accounted for 19, 16, 17, 7, and 2 Culicoides species collected in the provinces of Huelva, Cádiz, Sevilla, Córdoba, and Málaga, respectively (Table 1).
Culicoides abundance and species richness
In this study, the most common species was C. imicola (17.4%), followed by C. grandifovea sp. nov. (14.6%), C. kurensis (11.9%), and C. pseudolangeroni (11.7%). Collections of the remaining species were much less abundant (Table 2). Based on the frequency of trapping, C. circumscriptus was recorded as the most widely distributed species (122 times), followed by C. pseudolangeroni (55 times), C. imicola (48 times), and C. grandifovea sp. nov. (14 times). Due to missing, broken, or damaged parts of their bodies, 14.9% of the specimens could not be identified at species level. Out of the 23 collected Culicoides species, eight were plain-winged specimens and 15 were wing patterned species. Out of the 476 sampling sites (approximately 1786 days of trapping effort), 225 times (12.5%) were positive for Culicoides species (Fig. 1), including 118 (6.6%) in spring, 65 (3.6%) in summer, and 42 (2.3%) in autumn.
Flight seasonality of Culicoides species
Culicoides spp. (all species gathered) remained active throughout the entire period of sampling (Fig. 2), with a peak in September and decrease in November. Similar numbers of Culicoides were captured in May, June, October, and November, while the minimum Culicoides numbers were collected in August, coinciding with the hottest month in Andalusia (Fig. 2). Culicoides imicola was more commonly collected in September followed by July, but the peak of C. imicola captures was more notorious in the second survey, where they peaked in autumn (October and November). Culicoides kurensis, the third most common species, followed a similar trend than C. imicola. Moreover, C. grandifovea sp. nov. showed the opposite pattern than C. imicola and C. kurensis. Culicoides grandifovea sp. nov. exhibited moderate peaks from May to August, while was absent in autumn (Supplementary Fig. 1).

Number of Culicoides spp. per trap per night collected in Andalusia (southern Spain) in 476 sampling sites with carbon dioxide baited BG-traps. a 450 localities sampled in three occasions between April and November 2023. b 26 localities sampled at weekly or 2-weeks intervals between June and November 2023. The number above the bar represents the number of trapped individuals in each period
New recorded species
Culicoides grandifovea sp. nov. González, 2024 (Fig. 3).

Habitus of Culicoides grandifovea sp. nov. a General aspect. b Interocular space. c Antennal sensilla coeloconica distribution. d Maxillary palpus. e Sensory pit in the third palpus segment. f Scutum. g Wing pattern. h Spermathecae
ZooBank registration: details of the new species have been submitted to ZooBank. The LSID for the new name Culicoides grandifovea is urn: https://zoobank.org/References/12517686-f069-49b4-9c77-46d1dd617cfc
Type material: holotype (n = 1 female) from Huelva province (2/08/2023; 37.5230120 -6.5379540) (Andalusia) collected by a BG-sentinel 2 trap supplemented with dry ice and deposited in the Entomology collection of the Estación Biológica de Doñana (CODE: 2014.060; EBD-CSIC, Sevilla, Spain) (https://www.csis.es) along with paratypes (n = 3). Specimens stored in ethanol (70%) (1.5 Eppendorf tubes) and slide-mounted specimens (n = 10) also available upon request. https://www.ebd.csic.es/. Coordinates available in Supplementary Table 1.
Habitat: specimens collected in diverse habitats, including mainly Mediterranean scrubs and diverse tree forests such as eucalyptus, pine, olive grove, and Holm oak trees, among others.
Distribution: widely distributed in the provinces of Sevilla, Huelva, and Cádiz (southern Spain) (Fig. 1).
Etymology: based on the large size of the sensory pit of the third palpus segment. Fovea (foʊviə) is Latin for pit. It refers to a pit or depression in the third palpus.
Description of female
Size: medium size species (1.3–1.6 mm) (Fig. 3a).
Head: eyes bare, narrowly separated by a distance of approximately three-fourths or one ocular facet (Fig. 3b). Eyes connected by two fine transverse sutures (superior and central) enclosing the interocular setae. Number of sensilla coeloconica on flagellomeres III–XIV are 3–4/0–1/1/1/1/1/1/1/1/1/1/1/1/2 (Fig. 3c). Flagellomeres III–XIV with means of length of 49.2/32.5/31.6/32.6/32.9/32.4/31.8/32.2/48.2/48.4/50.7/53.1/67.5 µm. Mean AR of 0.97 (0.95–1.03 µm). Palpi are five-segmented with 238.80 ± 20.86 µm in length (Fig. 3d). The third palpus segment is strongly inflated with a single and wide shallow circular sensory pit (occupying almost the front half) of the palpus and is full of sensilla that externally exceed (Fig. 3e). Mean PR of 1.77 (1.65–1.78 µm). Area of sensory pit (third palpus) of 605.20 (68.3) µm2. Distance from posterior pharynx to end of hypopharynx: 215.25 (208–227 µm). Mandible with a mean of 11.75 teeth and maxillae with 15.50 teeth. Further details are provided in Supplementary Table 2.
Thorax: scutum dark brown with ornamentation (shown in fresh specimens). Median part of scutum with two broad light bands along the scutum (Fig. 3f). Color varies depending on light incidence. Halteres pale. Unspotted wings with abundant evenly distributed microtrichia, with a little more abundant microtrichia in the apical zone (R5 area) (Fig. 3g). Mean length of wings is 1087.3 (1039–1128 µm) and width is 502.1 (468–529 µm). Radial cells (R1 and R2) are noticeably dark. Inconspicuous marked pale spots on r–m cross vein and second costal area of wings. Legs brown uniform with tarsal segments lighter (slide-mounted specimens). Slender legs, fore (femur = 393–406 µm length and tibia = 388–409 µm), mid (femur = 392–408 µm length and tibia = 379–408 µm), and hind legs (femur = 306–328 µm length and tibia = 334–369 µm), with first tarsomere two times longer than the second one in the three pairs of legs. Tibial comb in fore legs with four major spurs of similar size along with other spines with smaller size. Spines of tarsomeres absent in fore and hind legs and present in tarsomere I–IV in middle legs.
Abdomen: two fully functional spherical spermathecae highly sclerotized (dark brown color). Spermathecae slightly asymmetric (length versus width: 66.5 ± 2.12 µm × 53.0 ± 4.32 µm and 53.0 ± 7.1 µm × 45.6 ± 6.66 µm) with a short unpigmented neck or without neck (Fig. 3h). Third rudimentary spermatheca vestigial. Sclerotized ring and abdominal sclerites absent.
Differential diagnosis and remarks. This species is similar in size and appearance to other dully colored (plain-wing and half-wing species) medium sized species such as C. kibunensis, C. indistinctus, C. odiatus, and other related species. However, the pale spot on both r–m and second costal spot is more marked in these species compared with C. grandifovea sp. nov. In addition, the large sensory pit of the third maxillary palpus might be observed under 6–8× magnification for ruling out the previous species. The ornamentation of the thorax is also shared with C. indistinctus. Accurate identification requires the elaboration of slide-mounted specimens. Under the compound microscope, C. grandifovea sp. nov. is unique since it combines the three following features: (1) distribution of sensilla coeloconica from III–XIV (variable in IV) (allowing the exclusion of many other plain-wing species), (2) the third palpal segment is moderately swollen with a large circular shallow sensory pit (in C. odiatus and C. indistinctus is different) with certain resemblance to the palpi of C. kurensis, and (3) spermathecae are slightly asymmetric, highly sclerotized (dark brown color), and spherical without neck. Unspotted wings are overall large and showy. Another less relevant feature is the presence of two sutures joining the eyes. Males were not captured, probably because carbon dioxide traps do not attract them.
New record of C. pseudolangeroni
Culicoides pseudolangeroni represents the first record from Europe. Collections of this species (11.8% of the total; 363 females and 1 male from 11 April to 27 October 2023) occurred in the five sampled provinces (Cádiz, Sevilla, Huelva, Málaga, and Córdoba) (Fig. 1). Its body length size is between 0.95–1.15 mm (Supplementary Fig. 2a). Eyes are bare and separated by a distance equal to the diameter of one ommatidial facet (Supplementary Fig. 2b). Thorax is typically brown, unspotted, and covered by visible interspersed setae (Supplementary Fig. 2c). Scutellum is usually yellowish or lighter compared with scutum (Supplementary Fig. 2d). The functional spermathecae are lightly sclerotized with a short neck and ring present (Supplementary Fig. 2e). Antennae, sensilla coeloconica on flagellomeres III–VI and XI–XIV (Supplementary Fig. 2f). Plain wings, with no markings (Supplementary Fig. 2g). Maxillary palpi (third palpus segment) with a single open and shallow sensory pit (Supplementary Fig. 2h). The most characteristic feature of males is the base of the parameters. They bear a highly sclerotized circular-shape structure leading to a pointed protuberance (Supplementary Fig. 2i). Further details are provided in Supplementary Table 2.
Phylogenetic analysis
New COI sequences (> 600 pb) have been deposited for C. grandifovea sp. nov. (LC819641-46), C. pseudolangeroni (LC819647-49, LC819654-55), C. kurensis (LC819650-51), and C. indistinctus (LC819652-53). Intraspecific d values were very low (d = 0.002%) among C. grandifovea sp. nov. sequences. Similarity of C. grandifovea COI sequences with other Culicoides sequences were lower than 92.6%. The more similar sequences corresponded to an unknown Culicoides sp. (MK732284 and MK732286) captured in Morocco and C. kurensis, forming a cluster separate from the rest of the species of the subgenus Oecacta (blue box, Fig. 4). Two out of the six specimens analyzed by barcoding showing lack of sensilla coeloconica in segment IV resulted to be genetically similar to the other four C. grandifovea sp. nov. showing such sensilla.

Maximum likelihood (ML) phylogenetic tree based on 55 COI sequences of Culicoides species. Topological branch support for the ML analysis (aLRT/bootstrap) is reported over specific branches, with values > 75% defining high stability. The sequences of this study (n = 15; four species) are marked in bold
Similarity of C. pseudolangeroni sequences in relation with the closely related species Culicoides langeroni (KJ729987) ranged between 98.06% and 97.25%. According to the phylogenetic tree, C. pseudolangeroni is a single monophyletic group (green box, Fig. 4) with an intraspecific d value (d = 0.001%) for C. pseudolangeroni, albeit clustered alongside C. langeroni. The genetic distance between C. pseudolangeroni and C. langeroni COI sequences ranged between 2% and 3% (Supplementary Table 3).
Discussion
This study provides a comprehensive faunistic catalog of the Culicoides species found in southern Spain, an area historically affected by Culicoides-borne pathogens [29, 59]. The use of nonstandard sampling methods carried out in diverse environments helped in identifying a high diversity of species, including some previously unknown species in this region. Interestingly, using this approach, two of the three most common species recorded in the area resulted to be a new species for science and a new record for Europe. In addition, many of the remaining collected Culicoides species are poorly documented in literature.
Light or carbon dioxide are the most commonly used baits for the collection of Culicoides midges [60, 61]. Due to the strong attraction of UV-light sources, Culicoides are usually collected in high numbers, especially in livestock farms and natural landscapes where wild ruminants are present [60, 62]. Light traps represent a practical and economical trapping system to determine species presence and abundance in an area [63]. However, light is an artificial attraction stimulus that does not mimic or reflect any response to a host. Several studies have indicated carbon dioxide to be an attractant for a number of blood-feeding insects, including Culicoides [64, 65]. However, a limited number of studies have exploited it, as carbon dioxide is relatively expensive, has a short period of operation, and is often considered impractical for routine use in large-scale surveillance programs [22, 63].
Using carbon dioxide traps, we collected a wide range of Culicoides species but in lower numbers compared with light traps, which is in line with other studies [61,62,63, 66]. While we collected a mean of 1.72 Culicoides specimens (per trap per day), 10–100 midges (per trap per day) have been collected in studies using light traps in southern Spain [47, 50]. Regarding the species composition, C. imicola, C. newsteadi, C. pulicaris group, C. circumscriptus, and C. obsoletus group dominated in previous studies using light traps in farms or in presence of livestock in Andalusia [28, 48, 50, 53], which contrasts with our findings. This difference in species composition might be attributed to different factors including variable grade of attraction to carbon dioxide and the habitat sampled, with species such as those of the Obsoletus and Pulicaris groups usually associated with livestock [20, 67, 68]. In addition, the traps used may affect the ratios, abundances, and species richness of Culicoides captured [61, 69,70,71]. Carbon dioxide-baited traps may be useful to capture host-seeking females for epidemiological studies while UV traps might be used to capture blood-fed females for host identification analysis [61, 65, 66, 72]. In fact, only six specimens with blood were collected in our study (0.2%). We showed that carbon dioxide baited traps were successful for the collection of the main Afrotropical vector C. imicola, which is consistent with other studies [64]. However, no collections were made of the widespread species C. obsoletus, probably because this species responds poorly to carbon dioxide-baited traps [66, 73,74,75].
Overall, the Culicoides fauna of southern Spain comprises species with different geographical distributions such as Palearctic, Mediterranean Basin, and Afrotropical species [76]. About 35% of the Culicoides species collected belonged to the so-called group of “plain-wing species” and/or poorly developed wing pattern species, which are predominant in drier and more open habitats [77]. Interestingly, among the recorded species, we found the new species C. grandifovea sp. nov. with a broad distribution in southern Spain and relatively high abundance in the area. The molecular analyses of the barcoding region of this species support a single genetic cluster group, with the closer phylogenetic relations with sequences from unidentified Culicoides from Morocco and with C. kurensis. Culicoides grandifovea sp. nov. displayed a flight activity with a major peak in the hottest and driest month of the year. The fact that this second most abundant species was not recorded previously might indicate its absence in farmland habitats and/or a low attraction to light traps. Also, active trapping conducted in previous years with suction light traps in the region have revealed absence of this species (data not shown). Based on the distribution of sensilla coeloconica and the palpus size, a preference of this species to feed on avian blood might be expected [78]. We also recorded C. pseudolangeroni for the first time in Europe. This species belongs to the C. langeroni species group, together with C. langeroni, Culicoides judae, and Culicoides molotovae [79], which is in line with the phylogenetic tree results. Culicoides pseudolangeroni has been previously found in deserts of Central Asia and North Africa [14, 80,81,82]. In addition, at least three individuals of C. jumineri near bahrainensis were captured in this study. The taxonomy of the Jumineri species group is not yet studied and the taxonomic status of the species remains unclear. Unfortunately, we were unable to recover DNA from these specimens, as they were mounted in slides. A molecular analysis comparing the nucleotide sequences of genes such as COI and/or ITS2 is needed to resolve the issue between C. jumineri s.s. and C. bahrainensis s.s., the latter distributed in Saudi Arabia [83]. In addition, three Culicoides variations (C. haranti variation, C. kurensis variation and C. indistinctus) were recorded. These kinds of variations are frequently recorded in literature [14, 84]. Also, five unknown Culicoides taxa were recorded in low numbers (≤ 2 specimens each one). These species possess sensilla coeloconica variations in antennal flagellomeres and/or other features (atypical pit shape); however, more specimens are necessary to determine if they represent valid species. This material evidences the complexity of the taxonomy of the Culicoides genus.
Regarding BTV, AHSV, and EHD virus disease vectors, C. obsoletus and C. pulicaris group species were less predominant and geographically temporally distributed than C. imicola, but they are frequently reported in southern Spain [45, 50, 53]. It is interesting to note the absence of members of C. pulicaris/C. lupicaris and C. obsoletus groups except two specimens of C. montanus. This can be due to different nonexclusive causes. First, latter species are usually associated with farm environments, and second, it might be possible that C. montanus can be overlooked with C. obsoletus and Culicoides scoticus species. However, they can be easily separated from the other members of the Obsoletus group under the stereo microscope by observing the single deep pit of the palpi. Similar reasons might explain that some species were not recorded in the past by light suction traps, being some of them particularly abundant in our study (C. kurensis, C. grandifovea sp. nov., and C. pseudolangeroni). Ornithophilic species such as C. circumscriptus and C. jamaicensis (previously named C. paolae) commonly captured in the region by different trapping methods [38], have resulted also abundant in our carbon dioxide baited traps.
Finally, our results provide information on the seasonal activity of Culicoides species in the area. Although variable between species, the flight activity of Culicoides spp. (including C. imicola) showed a major peak at the end of spring and another one in September/October. These results agree with previous studies, where C. imicola peaked between August and November with a remarkable variation depending on sites [28, 45, 48, 53]. Abiotic and biotic parameters including climatic variables such as precipitation and temperature may determine the abundance of Culicoides species in the area [28, 48, 50, 52, 85]. This may be especially relevant due to the low rainfall (totally absent in mid-summer) and extremely high temperatures (July–August) in the study area which may impact the development and/or flight activity of most Culicoides species.
Conclusions
Through a comprehensive literature review alongside extensive active trapping, we have expanded the known Culicoides fauna in Spain to 88 valid species, specifically to 53 in southern Spain. Our findings underscore the importance of complementing traditional UV-light traps with alternative trapping methods such as carbon dioxide-baited traps to comprehensively assess Culicoides abundance and distribution. This system allowed us to identify a previously undescribed species of Culicoides despite its widespread distribution and abundance in the area. Future studies should determine the role of these new or poorly documented Culicoides species in the transmission of pathogens of interest in animal and public health.
Availability of data and materials
Data is provided within the manuscript or supplementary information files.
Abbreviations
- UV:
-
Ultraviolet
- AHSV:
-
African horse sickness virus
- BTV:
-
Bluetongue virus
- SV:
-
Schmallenberg virus
- EHD:
-
Epizootic hemorrhagic disease
- OVI:
-
Onderstepoort veterinary institute
- COI:
-
Cytochrome c oxidase subunit I
- AR:
-
Antennal ratio
- PR:
-
Palpal ratio
- LISD:
-
Life science identifiers
References
Nielsen SA, Kristensen M. Delineation of Culicoides species by morphology and barcode exemplified by three new species of the subgenus Culicoides (Diptera: Ceratopogonidae) from Scandinavia. Parasit Vectors. 2015;8:151.
Talavera S, Muñoz-Muñoz F, Verdún M, Pagès N. Morphology and DNA barcoding reveal three species in one: description of Culicoides cryptipulicaris sp. nov. and Culicoides quasipulicaris sp. nov. in the subgenus Culicoides. Med Vet Entomol. 2017;31:178–91.
Pagès N, Muñoz-Muñoz F, Talavera S, Sarto V, Lorca C, Núñez JI. Identification of cryptic species of Culicoides (Diptera: Ceratopogonidae) in the subgenus Culicoides and development of species-specific PCR assays based on barcode regions. Vet Parasitol. 2009;165:298–310.
Sarvašová A, Kočišová A, Candolfi E, Mathieu B. Description of Culicoides (Culicoides) bysta n sp., a new member of the Pulicaris group (Diptera: Ceratopogonidae) from Slovakia. Parasit Vectors. 2017;10:279.
Yildirim A, Dik B, Duzlu O, Onder Z, Ciloglu A, Yetismis G, et al. Genetic diversity of Culicoides species within the Pulicaris complex (Diptera: Ceratopogonidae) in Turkey inferred from mitochondrial COI gene sequences. Acta Trop. 2019;190:380–8.
Mellor PS, Boorman J, Baylis M. Culicoides biting midges: their role as arbovirus vectors. Annu Rev Entomol. 2000;45:307–40.
Carpenter S, Groschup MH, Garros C, Felippe-Bauer ML, Purse BV. Culicoides biting midges, arboviruses and public health in Europe. Antiviral Res. 2013;100:102–13.
Elbers ARW, Meiswinkel R, van Weezep E, van Oldruitenborgh-Oosterbaan MMS, Kooi EA. Schmallenberg virus in Culicoides spp. biting midges, the Netherlands, 2011. Emerg Infect Dis. 2013;19:106–9.
Pagès N, Talavera S, Verdún M, Pujol N, Valle M, Bensaid A, et al. Schmallenberg virus detection in Culicoides biting midges in Spain: First laboratory evidence for highly efficient infection of Culicoides of the Obsoletus complex and Culicoides imicola. Transbound Emerg Dis. 2018;65:e1-6.
Quaglia M, Foxi C, Satta G, Puggioni G, Bechere R, De Ascentis M, et al. Culicoides species responsible for the transmission of Epizootic Haemorrhagic Disease virus (EHDV) serotype 8 in Italy. Vet Ital. 2023;59:83–8.
Jiménez-Cabello L, Utrilla-Trigo S, Lorenzo G, Ortego J, Calvo-Pinilla E. Epizootic hemorrhagic disease virus: current knowledge and emerging perspectives. Microorganisms. 2023;11:1339.
Martínez-de la Puente J, Martínez J, Rivero-De Aguilar J, Herrero J, Merino S. On the specificity of avian blood parasites: revealing specific and generalist relationships between haemosporidians and biting midges. Mol Ecol. 2011;20:3275–87.
González M, Goldarazena A. El género Culicoides en el Pais Vasco: guía práctica para su identificación y control. Gobierno Vasco-Eusko Jaurlaritza: Vitoria-Gasteiz; 2011.
Mathieu B, Cêtre-Sossah C, Garros C, Chavernac D, Balenghien T, Carpenter S, et al. Development and validation of IIKC: an interactive identification key for Culicoides (Diptera: Ceratopogonidae) females from the Western Palaearctic region. Parasit Vectors. 2012;5:137.
Rawlings P. A key, based on wing patterns of biting midges (genus Culicoides Latreille - Diptera: Ceratopogonidae) in the Iberian Peninsula, for use in epidemiological studies. Graellsia. 1996;52:57–71.
Ramilo DW, Diaz S, Pereira da Fonseca I, Delécolle JC, Wilson A, Meireles J, et al. First report of 13 species of Culicoides (Diptera: Ceratopogonidae) in mainland Portugal and Azores by morphological and molecular characterization. PLoS ONE. 2012;7:e34896.
Hristescu D, Bǎrbuceanu F, Dascǎlu L, Nitescu C, Goffredo M, Santilli A, et al. Species composition and relative abundance of the genus Culicoides (Diptera: Ceratopogonidae) in Romania. Parasit Vectors. 2020;13:393.
Goffredo M, Meiswinkel R, Federici V, Di Nicola F, Mancini G, Ippoliti C, et al. The `Culicoides obsoletus group´ in Italy: relative abundance, geographic range, and role as vector for bluetongue virus. Vet Ital. 2016;52:235–41.
Mignotte A, Garros C, Gardès L, Balenghien T, Duhayon M, Rakotoarivony I, et al. The tree that hides the forest: cryptic diversity and phylogenetic relationships in the Palaearctic vector Obsoletus /Scoticus Complex (Diptera: Ceratopogonidae) at the European level. Parasit Vectors. 2020;13:265.
Cuéllar AC, Kjær LJ, Kirkeby C, Skovgard H, Nielsen SA, Stockmarr A, et al. Spatial and temporal variation in the abundance of Culicoides biting midges (Diptera: Ceratopogonidae) in nine European countries. Parasit Vectors. 2018;11:112.
Slama D, Chaker E, Babba H, Slama D. Morphological Keys for the Identification of Tunisian Culicoides Biting Midges (Diptera: Ceratopogonidae). In: Farzana Khan P, editor. The Wonders of Diptera Charactistics, Diversity, and Significance for the World’s Ecosystems. London: IntechOpen; 2022.
McDermott EG, Lysyk TJ. Sampling considerations for adult and immature Culicoides (Diptera: Ceratopogonidae). J Insect Sci. 2020;20:2.
González MA, Goiri F, Prosser SWJ, Cevidanes A, Hernández-Triana LM, Barandika JF, et al. Culicoides species community composition and feeding preferences in two aquatic ecosystems in northern Spain. Parasit Vectors. 2022;15:199.
Alarcón-Elbal PM, Lucientes J. Actualización del catálogo de Culicoides Latreille, 1809 (Diptera, Ceratopogonidae) de España. Graellsia. 2012;68:353–62.
Veiga J, Martínez-de la Puente J, Václav R, Figuerola J, Valera F. Culicoides paolae and C. circumscriptus as potential vectors of avian haemosporidians in an arid ecosystem. Parasit Vectors. 2018. https://doi.org/10.1186/s13071-018-3098-8.
González MA, Bravo-Barriga D, López-Mercadal J, Miranda MÁ, Gutiérrez-López R, Barceló C. First report of Culicoides caucoliberensis in Spain: exploring molecular phylogeny, host-feeding behaviour and avian haemosporidian parasites. Med Vet Entomol. 2023;37:871–7.
Murillo JMS, González M, Díaz MMM, Galán AR. First record of Culicoides paradoxalis Ramilo & Delécolle, 2013 (Diptera, Ceratopogonidae) in Spain. Graellsia. 2015;71:e033.
Ortega MD, Lloyd JE, Holbrook FR. Seasonal and geographical distribution of Culicoides imicola Kieffer (Diptera: Ceratopogonidae) in Southwestern Spain. J Am Mosq Control Assoc. 1997;13:227–32.
Rodriguez M, Hooghuis H, Castaño M. African horse sickness in Spain. Vet Microbiol. 1992;33:129–42.
Rodríguez-Sánchez B, Gortázar C, Ruiz-Fons F, Sánchez-Vizcaíno JM. Bluetongue virus serotypes 1 and 4 in red deer. Spain Emerg Infect Dis. 2010;16:518–20.
Figuerola J, Jiménez-Clavero MÁ, Ruíz-López MJ, Llorente F, Ruiz S, Hoefer A, et al. A one health view of the West Nile virus outbreak in Andalusia (Spain) in 2020. Emerg Microbes Infect. 2022;11:2570–8.
Ramilo D. Phenotypic and genetic characterization of Culicoides (Diptera: Ceratopogonidae) in Portugal and comparison of the effect of pyrethroid insecticides in their control. Doctoral Thesis. University of Lisbon. 2016.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994;3:294–9.
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, et al. Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28:1647–9.
Trifinopoulos J, Nguyen LT, von Haeseler A, Minh BQ. W-IQ-TREE: a fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016;44:W232–5.
Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980;16:111–20.
Haddaway NR, Woodcock P, Macura B, Collins A. Making literature reviews more reliable through application of lessons from systematic reviews. Conserv Biol. 2015;29:1596–605.
Bravo-Barriga D, González MA, Parreira R, Frontera E, Huerta H, Alarcón-Elbal PM. Shedding light on the controversial taxonomic status of Culicoides jamaicensis and Culicoides paolae (Diptera: Ceratopogonidae): an overseas trip among continents. J Med Entomol. 2023;60:944–54.
Strobl G. Chironomidae. Spanische Dipteren (Chironomidae). XI Theil, Wiener Entomologische, XIX, Jahrgang, VI und VII. 169–174
Mellor PS, Boorman JPT, Wilkinson PJ, Martinez-Gómez F. Potential vectors of bluetongue and African horse sickness viruses in Spain. Vet Rec. 1983;112:229–30.
Mellor PS, Jennings DM, Wilkinson PJ, Boorman JP. Culicoides imicola: a bluetongue virus vector in Spain and Portugal. Vet Rec. 1985;116:589–90.
Anónimo. Study on the geographical distribution and seasonal prevalence in Spain during 1990–1991 of different species of the genus Culicoides (Family Ceratopogonidae). Doc científico-técnico restringido, Doc VI/650/92 Minist Agric Pesca y Aliment. Madrid 1992;57 pp.
Ortega MD. Distribution and frequency of the Culicoides spp. (Diptera: Ceratopogonidae) in Andalucía, Spain. 37th Annual Livestock Insect Workers Conference. New Mexico, USA: Santa Fe; 1993.
Ortega MD, Holbrook FR. Presence of Culicoides imicola (Diptera: Ceratopogonidae) in Jaen. Spain J Am Mosq Control Assoc. 1994;10:463.
Ortega MD. 1995 Distribution and seasonal fluctuations of Culicoides spp. in occidental Andalucia, Spain. Masters of Science in Entomology, Wyoming.
Rawlings P, Pro MJ, Pena I, Ortega MD, Capela R. Spatial and seasonal distribution of Culicoides imicola in Iberia in relation to the transmission of African horse sickness virus. Med Vet Entomol. 1997;11:49–57.
Ortega MD, Mellor PR, Rawlings MJP. The seasonal and geographical distribution of Culicoides imicola, C. pulicaris group and C. obsoletus group biting midges in central and southern Spain. Arch Virol. 1998;14:85–91.
Ortega MD. Seasonal distribution and relationship to and precipitation of the most abundant species of Culicoides in five provinces of Andalusia. Spain J Am mosq Control Assoc. 1999;15:391–9.
Estrada R, Carmona VJ, Alarcón-Elbal PM, Miranda MA, Borrás D, Roche ML, et al. Primera cita de Culicoides paolae Boorman, 1996 (Diptera, Ceratopogonidae) para la Península Ibérica y aportaciones sobre su distribución. Boletín Soc Entomológica Aragón. 2011;49:217–21.
Pérez JM, García-Ballester JA, López-Olvera JR, Serrano E. Monitoring bluetongue virus vectors in Andalusia (SW Europe): Culicoides species composition and factors affecting capture rates of the biting midge Culicoides imicola. Parasitol Res. 2012;111:1267–75.
Ferraguti M, Martínez-de la Puente J, Ruiz S, Soriguer R, Figuerola J. On the study of the transmission networks of blood parasites from SW Spain: diversity of avian haemosporidians in the biting midge Culicoides circumscriptus and wild birds. Parasit Vectors. 2013. https://doi.org/10.1186/1756-3305-6-208.
Ducheyne E, Chueca MAM, Lucientes J, Calvete C, Estrada R, Boender GJ, et al. Abundance modelling of invasive and indigenous Culicoides species in Spain. Geospat Health. 2013;8:241–54.
Arenas-Montes AJ. Estudio epidemiológico de lengua azul y enfermedad hemorrágica epizoótica en ecosistemas mediterráneos del sur de España. Doctoral thesis. Spain: University of Cordoba; 2013.
Talavera S, Muñoz-Muñoz F, Durán M, Verdún M, Soler-Membrives A, Oleaga Á, et al. Culicoides species communities associated with wild ruminant ecosystems in Spain: tracking the way to determine potential bridge vectors for arboviruses. PLoS ONE. 2015;10:e0141667.
Martínez-de la Puente J, Navarro J, Ferraguti M, Soriguer R, Figuerola J. First molecular identification of the vertebrate hosts of Culicoides imicola in Europe and a review of its blood-feeding patterns worldwide: implications for the transmission of bluetongue disease and African horse sickness. Med Vet Entomol. 2017;31:333–9.
Pagès N, Muñoz-Muñoz F, Verdún M, Pujol N, Talavera S. First detection of Wolbachia-infected Culicoides (Diptera: Ceratopogonidae) in Europe: Wolbachia and Cardinium infection across Culicoides communities revealed in Spain. Parasit Vectors. 2017;10:582.
Estrada R, Delacour S, Navarro J, Lucientes CJ. Segunda cita de Culicoides (Pontoculicoide) tauricus Gutsevich, 1959 (Diptera, Ceratopogonidae) de la Península Ibérica y primera cita de la provincia de Granada (Andalucía, España). Boletín la SEA. 2019;65:290–2.
Garrido-Bautista J, Martínez-de la Puente J, Ros-Santaella JL, Pintus E, Lopezosa P, Bernardo N, et al. Habitat-dependent Culicoides species composition and abundance in blue tit (Cyanistes caeruleus) nests. Parasitology. 2022;149:1119–28.
Allepuz A, García-Bocanegra I, Napp S, Casal J, Arenas A, Saez M, et al. Monitoring bluetongue disease (BTV-1) epidemic in southern Spain during 2007. Prev Vet Med. 2010;96:263–71.
González M, Alarcón-Elbal PM, Valle-Mora J, Goldarazena A. Comparison of different light sources for trapping Culicoides biting midges, mosquitoes and other dipterans. Vet Parasitol. 2016;226:44–9.
Zhang X, Li J, Gerry AC. Comparison of trap efficiency using suction traps baited with either UV or CO2 for the capture of Culicoides (Diptera: Ceratopogonidae) species in the Southern California desert. United States J Med Entomol. 2023;60:193–201.
Venter GJ, Hermanides KG. Comparison of black and white light for collecting Culicoides imicola and other livestock-associated Culicoides species in South Africa. Vet Parasitol. 2006;142:383–5.
Venter GJ, Boikanyo SNB, Majatladi DM, Morey L. Influence of carbon dioxide on numbers of Culicoides midges collected with suction light traps in South Africa. Med Vet Entomol. 2016;30:117–22.
Logan JG, Cook JI, Mordue Luntz AJ, Kline D. Understanding and exploiting olfaction for the surveillance and control of Culicoides biting midges. In: Takken W, Knols BGJ, editors. Olfaction in vector-host interactions Wageningen Academic Publishers. Netherlands: Wageningen; 2010. p. 217–46.
Nelson RL. Carbon dioxide as an attractant dor Culicoides. J Med Entomol. 1965;2:56–7.
Gerry AC, Sarto I Monteys V, Vidal JOM, Francino O, Mullens BA. Biting rates of Culicoides midges (Diptera: Ceratopogonidae) on sheep in northeastern Spain in relation to midge capture using UV light and carbon dioxide-baited traps. J Med Entomol. 2009;46:615–24.
González M, Baldet T, Delécolle JC, López S, Romón P, Goldarazena A. Monitoring of Culicoides Latreille (Diptera: Ceratopogonidae) after BTV outbreaks, in sheep farms and natural habitats from the Basque Country (Northern Spain). Proc Entomol Soc Washingt. 2013;115:48–69.
Cuéllar AC, Kjær LJ, Baum A, Stockmarr A, Skovgard H, Nielsen SA, et al. Modelling the monthly abundance of Culicoides biting midges in nine European countries using Random Forests machine learning. Parasit Vectors. 2020;13:194.
Becker M, Park JS, Gentry G, Husseneder C, Foil L. Comparison of trapping methods for use in surveys for potential Culicoides vectors of orbiviruses. Parasit Vectors. 2021;14:564.
Carpenter S, Szmaragd C, Barber J, Labuschagne K, Gubbins S, Mellor P. An assessment of Culicoides surveillance techniques in northern Europe: have we underestimated a potential bluetongue virus vector? J Appl Ecol. 2008;45:1237–45.
Viennet E, Garros C, Lancelot R, Allène X, Gardès L, Rakotoarivony I, et al. Assessment of vector/host contact: comparison of animal-baited traps and UV-light/suction trap for collecting Culicoides biting midges (Diptera: Ceratopogonidae), vectors of Orbiviruses. Parasit Vectors. 2011;4:119.
Cohnstaedt LW, Rochon K, Duehl AJ, Anderson JF, Barrera R, Su NY, et al. Arthropod surveillance programs: basic components, strategies, and analysis. Ann Entomol Soc Am. 2012;105:135–49.
González MA. El género Culicoides (Diptera: Ceratopogonidae) en el País Vasco, norte de españa. Doctoral Thesis. University of the UPV. 2014.
Mullens BA, Owen JP, Heft DE, Sobeck RV. Culicoides and other biting flies on the Palos Verdes Peninsula of Southern California, and their possible relationship to equine dermatitis. J Am Mosq Control Assoc. 2005;21:90–5.
Harrup LE, Logan JG, Cook JI, Golding N, Birkett MA, Pickett JA, et al. Collection of Culicoides (Diptera: Ceratopogonidae) using CO2 and enantiomers of 1-octen-3-ol in the United Kingdom. J Med Entomol. 2012;49:112–21.
Bourquia M, Garros C, Rakotoarivony I, Gardès L, Huber K, Boukhari I, et al. Update of the species checklist of Culicoides Latreille, 1809 biting midges (Diptera: Ceratopogonidae) of Morocco. Parasit Vectors. 2019;12:459.
Boorman J. Culicoides (Diptera: Ceratopogonidae) of the Arabian Peninsula with notes on their medical and veterinary importance. Fauna Saudi Arabia. 1989;10:160–224.
Martínez-de la Puente J, Figuerola J, Soriguer R. Fur or feather? feeding preferences of species of Culicoides biting midges in Europe. Trends Parasit. 2015;31:16–22.
Glukhova VM, Braverman Y. Review of the palearctic desert biting midges Culicoides langeroni group, with a description of a new species (Diptera: Ceratopogonidae). J Med Entomol. 1999;36:309–12.
Kremer M, Chaker E, Delécolle JC. Description de Culicoides pseudolangeroni n. sp [Dipt. Ceratopogonidae]. Bull la Société Entomol Fr. 1981;86:291–7.
Slama DC, E, Zrelli S, Mathieu B. Culicoides (Diptera: Ceratopogonidae) Fauna in Central Tunisia. Entomol Ornithol Herpetol Curr Res. 2016;5:2161–983.
Pasteur UL, Messaddeq, N, Lembre C, Kremer M. Reevaluation of the taxonomic status of the Culicoides spp. (Diptera:Ceratopogonidae) from Israel and the eastern Mediterranean and review of their potential medical and veterinary importance. J Am Mosq Control Assoc. 1996;12:437–45.
Kheir SM. Seasonal activity of Culicoides bahrainensis Boorman, 1989 (Diptera: Ceratopogonidae) in Saud Arabia. J King Saud Univ Sci. 2010;22:167–72.
Braverman Y, Messaddeq N, Kremer M. Abnormal features in specimens of eight Culicoides spp. (Diptera: Ceratopogonidae) from the eastern mediterranean area. Isr J Zool. 1993;39:157–66.
Mullen GR, Jones RH, Braverman YNK. Laboratory infections of Culicoides debilipalpis and C. stellifer (Diptera: Ceratopogonidae) with bluetongue virus. Prog Clin Biol Res. 1985;178:239–43.
Acknowledgements
We thank Alvaro Solis, Cintia Vega, Cristina Diaz, Maria del Mar Ándujar, and Juan José Talaverón for helping in the capture and separation of Culicoides. We thank Shirin Taheri for her help in the elaboration of the map and Maria José Ruiz-López for her support in molecular analysis.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This study has been funded by Fundación “La Caixa” through the project ARBOPREVENT (HR22-00123) and Junta de Andalusia. Additional funding derived from project PN2022-2945 from the Organismo Autónomo Parques Nacionales and PLEC2021-007968 project (NEXTHREAT) funded by MCIN/AEI/https://doi.org/10.13039/5011000110333 and supported by EU NextGeneration funds.
Author information
Authors and Affiliations
Contributions
J.F. conceived, designed, and supervised the study and revised the final draft of the manuscript; M.A.G. conceived the study and performed data collection, investigation, and writing—original draft preparation; S.M., J.M.P., V.S.M., and D.B.B. analyzed the data, interpreted the results, and reviewed the manuscript. All authors reviewed and approved the final version for submission.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
González, M.A., Magallanes, S., Bravo-Barriga, D. et al. Sampling of Culicoides with nontraditional methods provides unusual species composition and new records for southern Spain. Parasites Vectors 17, 338 (2024). https://doi.org/10.1186/s13071-024-06414-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06414-2