
Abstract
Background
Aedes aegypti is the main vector of several arthropod-borne viral infections in the tropics profoundly affecting humans, such as dengue fever (DF), West Nile (WN), chikungunya and more recently Zika. Eradication of Aedes still largely depends on insecticides, which is the most cost-effective strategy, and often inefficient due to resistance development in exposed Aedes populations. We here conducted a study of Ae. aegypti resistance towards several insecticides regularly used in the city of Denpasar, Bali, Indonesia.
Methods
Aedes aegypti egg samples were collected with ovitraps and thereafter hatched in the insectary of the Gadjah Mada University. The F0 generation was used for all bioassay-related experiments and knockdown resistance (kdr) assays.
Results
Results clearly showed resistance development of Ae. aegypti against tested insecticides. Mortalities of Ae. aegypti were less than 90% with highest resistance observed against 0.75% permethrin. Mosquitoes from the southern parts of Denpasar presented high level of resistance pattern in comparison to those from the western and northern parts of Denpasar. Kdr analysis of voltage-gated sodium channel (Vgsc) gene showed significant association to S989P and V1016G mutations linked to resistance phenotypes against 0.75% permethrin. Conversely, Ae. aegypti F1534C gene mutation did not result in any significant correlation to resistance development.
Conclusions
Periodically surveillance of insecticide resistances in Ae. aegypti mosquitoes will help local public health authorities to set better goals and allow proper evaluation of on-going mosquito control strategies. Initial detection of insecticide resistance will contribute to conduct proper actions in delaying mosquito resistance development such as insecticide rotation or combination of compounds in order to prolong chemical efficacy in combating Ae. aegypti vectors in Indonesia.
Similar content being viewed by others

Background
Dengue fever (DF) is a serious arthropod-borne arboviral infectious disease, mainly distributed in tropical regions. As reviewed elsewhere [1] endemic dengue virus transmission is reported in the Eastern Mediterranean, American, South-East Asian, Western Pacific and African regions, whereas sporadic local transmission has been reported to occur in Europe and the USA. DF is a major public health concern of Indonesia as one of the principle endemic countries within Southeast Asia. Warm tropical climate, which clearly supports Aedes vector life, is considered one of the main risk factors [2] and influencing further the disease burden and its continuous persistence mainly in equatorial countries. In addition, the mosquito vector, Aedes aegypti, efficiently transmits DF due to its anthropophilic feeding and breeding characteristics [3]. It has been reported that 71,668 DF human cases have so far occurred in Indonesia in 2015 covering 34 provinces, which means that DF has spread now into all national territories [4]. During January-February of 2016, there have been 8,487 cases with 108 mortalities and 11 provinces are categorized of being at a remarkable risk. There is still little data on the origin of Ae. aegypti, which contributes to the DF, spread in Indonesia [5]. Active development of national programs to promote DF elimination has been initiated since many years through community based activities such as vector control strategies and trials on virus vaccination programs. Nonetheless society activities, such as the elimination of Ae. aegypti larvae breeding sites become difficult especially during heavy rainy seasons in Indonesia. To date, vector control is the main tool to reduce viral transmission since antiviral drugs and vaccinations are still in progress despite of intensive research and vaccine development efforts worldwide.
Thus, Ae. aegypti control is still based on the use of large-scale fogging and space-spraying of insecticides, although also [6] Wollbachia-engineered mosquitoes have also been previously used. Alongside, also the use of larvacides containing granule-temephos (ABATE®) in combination with manual-larval inspection by governmental teams are still applied in several tropical geographical regions. Integrated social activities shall be supported by perpetual motivation to close both artificial and natural water containers to prevent mosquito breeding. Nonetheless, the difficulties arise due to opened water media such as flower vases, coconut shells, trash cans cut-wooden basins, which are frequently uncontrolled. As the majority of people are using water reservoirs to bathe, this approach has to be concomittant with changes of social behavior in replacing water reservoirs periodically or by using continuously flowing water systems.
Consequently, the high usage of insecticides has generated dependency of these compounds for regular vector control activities mainly during wet seasons. More importantly, Ae. aegypti resistance to common used insecticides has previously been reported from different countries worldwide such as Colombia [7], Brazil [8, 9], Grand Cayman [10], Thailand [11], India [12], Malaysia [13], Mexico [14, 15] and China [16]. Moreover, mosquito resistance related studies in central regions of Java [17] and Yogyakarta [18], Indonesia, have shown the resistance development to pyrethroids. These studies reported on the association of insecticide resistance with knockdown resistance (kdr) genes on voltage-gated sodium channel (Vgsc) of Aedes mosquitoes. So far, the kdr profiling of Aedes have found at least seven point mutations, which can clearly lead to reduced sensitivity of sodium voltage channels to insecticides [19]. Identification of Ae. aegypti showed that S989P, I1011M/V, V1016G/I, F1269C, F1534C corresponded well to this resistance development [10, 20,21,22,23]. At least two of these mutations, i.e. V1016G /F1534C, and I were responsible for pyrethroid resistance [20, 21]. However, synergistic resistance may also occur by additional point mutations in different locations of Vgsc genes as postulated elsewhere [24].
The most populated islands in Indonesia are Java and Bali with around 60 million of citizens and this although these regions represent less than 7% of the national territory [25]. Population density is influenced by factors such as job opportunities, infrastructures, education facilities, administrative centres and touristic attractions. Disease outbreaks and vector conditions in these areas contribute significantly to the numbers to human DF cases surveillance and national control strategies [5]. Thus, the usage of skin repellents and space-spraying insecticides is much higher in these mostly residential areas. Therefore, the distribution of allele polymorphism in Ae. aegypti in these urban/suburban areas is important to be investigated periodically in order to guarantee proper control programs. Additionally, vast areas of the Indonesian archipelago lead to possible variations throughout tropical areas investigated. We here present genotyping, distributions and variations of Ae. aegypti Vgsc genes in different areas of the city of Denpasar, Bali, Indonesia.
Methods
Mosquito samples
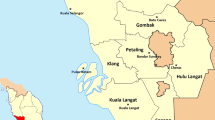
This study was performed in Denpasar, Bali, Indonesia (Fig. 1). Southern Bali area is the most developed part of the island. Ae. aegypti were collected by using 300 artificial ovitraps placed in residential areas of investigated districts. Ovitraps were made of glass with black stain outside. A filter paper was placed in the mouth of the glass. Filled water and filter papers were replaced every week on collection dates. Samples were collected from April to September 2016. The collection sites were randomly selected with particular emphasis on areas with previous reports of human DF cases and in which regular fogging activities occurred. Egg-containing papers were dried at room temperature (RT) and stored in plastic containers thereafter. Eggs were hatched in the insectary of the Department of Parasitology, Faculty of Veterinary Medicine, Gadjah Mada University, Yogyakarta, Indonesia. Larvae were fed with chicken liver (wet and dried) during their development until the pupa stage. Adult Ae. aegypti mosquitoes reared from these collected eggs were then fed with a 10% sugar solution absorbed into cotton balls. Emerged mosquitoes (F0) up to three day-old were used for all insecticide-related experiments.

a Map of the Island of Bali, Indonesia. b Sample sites for Ae. aegypti egg collections within the city of Denpasar
Insecticide susceptibility tests
Insecticide succeptibility tests (IST) were performed according to the World Health Organization (WHO) protocols for anopheline mosquito’s diagnostic doses [26]. The kits and insecticide impregnated papers were supplied by the University Saints Malaysia (Penang, Malaysia). The impregnated papers consisted of 5% malathion, 0.05% deltamethrin, 0.75% permethrin, 0.05% λ-cyhalothrin, 0.1% bendiocarb and 0.15% cyflothrin, respectively. In each assay 150 adult Ae. aegypti mosquitoes from each areas were divided into 6 tubes containing 25 mosquitoes each. Four tubes (4 replicates) served as replicates for 1 insecticide exposure and two tubes were used as controls. The mortality was calculated as a percentage using the formula: total number of dead mosquito/total sample size × 100, as reported elsewhere [26]. Abbott’s formula was not used in this study since control mortalities were always less than 5%. Some of survivors and dead mosquitoes from the bioassays were kept at -20 °C for further molecular resistance analysis. Resistance status of mosquitoes populations were defined according to WHO as < 90% mortality as previously reported [26].
Profiling of domains II and III of the voltage-gated sodium channel gene (Vgsc)
DNA isolation
DNA isolation was performed using PureLink® Genomic DNA Isolation Kit (Invitrogen, Carlsbad, USA) in combination with occasional vortexing using glass beads to ease mosquitoes lysis. Dead and surviving mosquitoes from different collection sites from bioassays were extracted individually.
Partial domain II of sodium voltage-gated channel (Vgsc) amplification
Detection of V1016G/I and S989P mutations were performed by direct sequencing utilizing primers IIP_F and IIS6_R which encompassed domain II segment 6 of Ae. aegypti Vgsc gene as described previously [11]. Domain II amplification was performed by using primer IIP_F dan IIS6_R (Table 1). Domain II PCR consisted of 1 μl of genomic DNA, 1 μl of 10 pmol forward primer (Macrogen, Seoul, Korea), 1 μl of 10 pmol reverse primer, 12.5 μl of DreamTaq Green PCR Master Mix (Thermo Fisher Scientific, Waltham, USA) in 25 μl total reaction volume. Both reactions were performed as follows: 95 °C for 5 min, 35 cycles of 30 s at 95 °C, 45 s at 63 °C, 45 s at 72 °C, and a final elongation step for 5 min at 72 °C. Amplified PCR product was then purified and directly sequenced (Integrated DNA Technologies/1st BASE, Singapore).
F1534C allele-specific PCR
Genotyping of mutant F1534C was performed according to previous publication [11] for allele - specific PCR assay. Primers used for the assay are listed in Table 1. F1534C PCR consisted of 1 μl of 10 pmol primer F1534f, 1 μl of 10 pmol primer C1534r, 0.5 μl of 10 pmol primer CPr, 12.5 μl of DreamTaq Green PCR Master Mix (Thermo Fisher Scientific) in 25 μl total reaction volume. Reactions were performed as follows: 94 °C for 2 min, 35 cycles of 30 s at 94 °C, 30 s at 60 °C, 30 s at 72 °C, and a final elongation step for 2 min at 72 °C. PCR amplification products were loaded onto a 3% agarose gel.
To confirm F1534C genotyping, several Ae. aegypti showing resistance phenotypes were also sequenced (n = 30). Amplification was performed by utilizing primers Ge-IIIS6_F and IIIS6_R [11]. Domain II PCR consisted of 1 μl of genomic DNA, 1 μl of 10 pmol forward primer (Macrogen), 1 μl of 10 pmol primer reverse primer, 12.5 μl of DreamTaq Green PCR Master Mix (Thermo Fisher Scientific) in 25 μl total reaction volume. Reactions were performed as follows: 95 °C for 5 min, 35 cycles of 30 s at 95 °C, 45 s at 63 °C, 45 s at 72 °C, and a final elongation step for 5 min at 72 °C. Amplified PCR product was purified and thereafter directly sequenced (Integrated DNA Technologies/1st BASE, Singapore).
Statistical analysis
Statistical analysis and graphical presentation were carried out using GraphPad Prism® 7.02.
Results
Adult Ae. aegypti resistance development to tested insecticides
A total of 1,890 specimens were tested against various insecticide-impregnated papers according to WHO protocols [26]. All tested Ae. aegypti mosquitoes from urban locations presented resistance levels to insecticides as shown in Fig. 2. The mortality percentages obtained for these tested mosquitoes showed that all mosquito populations collected in Denpasar were resistant to tested insecticides according to WHO criteria [26], i. e. with mortalities < 90%. Mosquitoes from southern Denpasar showed the highest resistance development to malathion 5% which was significantly different from the resistance observed in northern Denpasar (t = 2.81, P = 0.048) but no significant differences were observed when compared to the western parts of Denpasar (t = 2.65, P = 0.057). Resistance development to deltamethrin 0.05% was not significantly different between these urban areas. Moreover, permethrin resistance was quite high with less than 50% mosquito mortalities in all examined areas. Mosquitoes from southern and northern Denpasar showed the highest levels of resistance which differed significantly from that in mosquitoes from western Denpasar (t = 13.4, P ≤ 0.001). Furthermore, mild Ae. aegypti resistance development, with approximately 80–90% mortalities, were also observed against λ-cyhalothrin 0.05%, bendiocarb 0.1% and cyflothrin 0.15%, respectively.

Resistance profiles to different insecticides tested in Ae. aegypti from Denpasar, Bali, Indonesia. The bars are percent mortalities after exposure to insecticides with error bars representing standard deviations
Sequencing and mosquito genotyping the voltage-gated sodium channel gene (Vgsc)
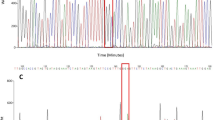
Sequencing of domain II Vgsc gene was analyzed to identify possible S989P and V1016G allele variations within collected mosquito population. Mutations of S989P were found in mosquitoes originating from Denpasar. S989P allele variations were found almost equally in wild type-, heterozygote- and homozygote-specimens. Frequency of P allele was half of resistant phenotypes to permethrin (Table 2). The electropherogram pattern of sequenced domain did not show any separate C encoding prolin pattern in all examined samples. Nonetheless, the presence of T encoding for serin in wild type or heterozygote forms was quite clear. G allele was a dominant allele found in the population with 0.7 frequency of resistance and 0.5 in succeptible groups. Clear electropherograms were always found in the homozygote samples. The homozygote G1016G was the highest allele to be found in the resistance group in comparison to S989P and F1534C (Table 2). Exemplary amplicons representing V1016G and S989P polymorphisms, respectively, were deposited in GenBank (accession nos. KY057038 and KY057037).
Allele-specific PCR of F1534C showed distributions F to C point mutations in the domain III of Vgsc gene. In the resistance (alive) phenotypes, homozygote frequencies of CC were 0.21 and heterozygote frequencies of FC were 0.25. In contrast, succeptible (dead) mosquito samples showed CC 0.13 and FC 0.11 frequencies. Allele containing C was found in heterozygotes and CC allele was relatively rarely found in the population of susceptible samples. Total C frequency was lower when compared to F frequency in Denpasar mosquitoes (Table 2). Sequences of domain III showed F1534C mutations and wild-type F1534F mutations, which were also deposited in GenBank (accession nos. KY078304 and KY078303).
When calculated separately, odd ratio (OR) S989P of samples (n = 110) showed association with resistance to permethrin (Fisher’s exact test, P = 0.0005, OR = 7.67, 95% CI = 2.14–27.49). Further, V1016G (n = 132) also showed significant correlation with resistance to permethrin (Fisher’s exact test, P = 0.014, OR = 2.49, 95% CI = 1.23–5.07). Conversely, F1534C mutation (n = 97) showed no significant association with phenotypes tested for permethrin (P ≥ 0.05). However, when C was present with P and G heterozygotes (Table 3, no. 2) and homozygotes (Table 3, no. 14), all mosquitoes (n = 9) were identified as totally resistant (alive) to permethrin 0.75%. Due to our limited data (results included only 97 clearly identified homozygote and heterozygote patterns with three point mutations), alleles presented in Table 3 only give a tendency in synergistic pattern of resistance. Strong association of resistance showed by PVF/PGF, OR of 15, CI: 1.21–185.21 (Table 3, no. 8), may indicate synergistic resistance by simultaneous presence of P and G.
Discussion
Aedes mosquito control still challenges public health authorities of subtropical and tropical countries worldwide due to mosquito ability to adapt fast to adverse breeding conditions and also by the development of resistance against routinely used insecticides [7, 10, 12, 14, 27, 28]. Moreover, the capability of Aedes spp. to transmit a wide spectrum of relevant arthropod-borne diseases in the subtropics and tropics makes this genus a serious threat to human health. Aedes control through the usage of insecticides is the preferred choice by the Indonesian government to reduce the number of DF. Insecticide fogging is a direct action program performed by Indonesian public health agencies when human DF cases are reported to occur within national territories.
Global movement of humans, international transportations, poor housing conditions, tourism and breeding opportunities of mosquitoes are well-known risk factors to be involved in the epidemiology and transmission of vector-borne diseases, particularly in underdeveloped geographic regions [29]. The above mentioned risk factors increase widespread use of insecticides and consequent resistance development within exposed mosquito populations. Besides, several complex mechanisms also contribute to serious limitations to the long-term usage of insecticides due to efficacy reduction [30]. Resistance indicators of mosquitoes can be analyzed in various ways; however, metabolic resistance-based methodologies are more costly in comparison with gene-targeted resistance-associated assays.
Human DF cases in the city of Denpasar, Bali, reached 6,898 with 38 mortalities recorded from 2014 until mid 2016 according to data records [Bali Health Agency (BHA), Indonesia]. Insecticide fogging with pyrethroid- and organophosphate-based compounds have been performed 818 times covering all regions of Denpasar city in those 3 years, i. e. 255 times in 2014; 310 in 2015; and 253 until mid 2016 (BHA, Indonesia). These data also showed trends of fogging intensities annually in the city of Denpasar. Fogging activities mainly targeted high burden areas based on patients reported from local hospitals. Moreover, urban areas with touristic attractions used mosquitoes repellents at higher frequencies in order to avoid mosquito bites. The commercial brand of repellents based on pyrethroid compounds were used by almost all citizens of Denpasar (BHA, Indonesia).
To our knowledge, we here present for the first time indications of resistence development in Ae. aegypti populations against commonly used insecticides regularly applied in control programs in the city of Denpasar, Bali. The detailed molecular Ae. aegypti phenotype analyses showed high resistance patterns to all insecticides tested. No significant differences from North, West and South Denpasar were detected pointing that insecticide compounds may have been used continuously in almost all areas with the same frequency and/or due to the fact that investigated areas were all from one city district. Noticeably, mosquito repellents such as skin lotions, in-house sprays and coils, are still broadly used by local citizens as well as by international travellers. Thus, mosquito exposure to such various repellent ingredients is continuous and even higher in this touristic tropical island when compared to less tourist-visited Indonesian islands. Our Ae. aegypti resistance results corroborate previous mosquito resistance findings in Java [17, 18], an adjacent island to Bali, which showed similar resistance patterns in Ae. aegypti populations. Although the analyzed area in these studies was much larger than that in our study [17], the resistance patterns were almost the same, thereby suggesting similar usage of insecticide active ingredients in Java. However, multiple insecticide resistance can also be transmitted from dry eggs stuck on containers which are transported from one area to another by humans. Although male or female mating competitiveness of resistant insect species are under investigation [31, 32], it is of concern that with or without insecticide exposure resistance can be accelerated through mating processes when insecticide resistance is initially detected as reported previously elsewhere [29, 30].
Based on analysis pointing on kdr mutations, V1016G haplotypes were most frequently found in comparison with the two other analyzed targets, i. e. F1534C and S989P. This haplotype is commonly found in Ae. aegypti [6, 13, 16, 17, 33] but not in South America where the haplotype V1016I is mainly dominating [34]. Conversely, the haplotype F1534C is much less frequently found in Aedes populations when compared to V1016G and S989P genes. This is in line with previous reports of Java [17, 18], although it is known that F1534C also significantly contributes to type I phyrethroid resistance as published elsewhere [35] and was also found as major allele in Aedes mosquitoes of South America [10].
Sequencing results of domain II revealed that allele carying 2 mutants on S989P and V1016G, respectively, appeared in relatively high numbers of resistant Ae. aegypti mosquitoes. Although the significant contribution on resistance development towards pyrethroids of S989P allele is still under debate, it may play an additive role in Ae. aegypti insecticide resistance development [16, 24, 28], in which the V1016G allele seems to play a pivotal role. Phenotypes in Table 3 may not only be explained by kdr mutations representing insensitivity of target sites towards tested insecticide as metabolic capacities within the genus Aedes might also explain phenotype variations. Consistently, wild-type genotypes with resistant phenotypes have been reported to occur suggesting enhanced metabolic detoxification capacities by catalytic sites of certain mosquito enzymes [36].
Periodical surveillance of insecticide resistance development in mosquitoes will clearly contribute in setting appropriate goals and evaluation process of ongoing control programs by Indonesian governmental authorities. Initial detection of resistance development may contribute to generate proper management/control strategies such as insecticide rotation programs or the combination of different ingredient types of insecticides in order to prolong chemical efficacy in combating these vectors.
Conclusions
Regular surveillance surveys on insecticide resistance development in Ae. aegypti mosquitoes will help local public health authorities to set better goals and allow proper evaluation of ongoing mosquito control strategies. Initial detection of insecticide resistance will also contribute to conduct proper actions in delaying mosquito resistance development such as insecticide rotation or combination of compounds in order to prolong chemical efficacy in combating Ae. aegypti vectors in Indonesia.
Abbreviations
- DF:
-
Dengue fever
- IST:
-
Insecticide succeptibility test
- Kdr :
-
Knockdown resistance
- OR:
-
Odds ratio
- RT:
-
Room temperature
- Vgsc:
-
Voltage-gated sodium channel gene
- WN:
-
West Nile
References
Guzman MG, Gubler DJ, Izquierdo A, Martinez E, Halstead SB. Dengue infection. Nat Rev Dis Primers. 2016;2:16055.
Colon-Gonzalez FJ, Fezzi C, Lake IR, Hunter PR. The effects of weather and climate change on dengue. PLoS Negl Trop Dis. 2013;7:e2503.
WHO. Dengue: guidelines for diagnosis, treatment, prevention and control. Geneva: World Health Organization; 2009.
Ministry of Health Republic of Indonesia. Demam Berdarah Biasanya Mulai Meningkat di Januari. 2015. http://www.depkes.go.id/article/view/15011700003/demam-berdarah-biasanya-mulai-meningkat-di-januari.html. Accessed 29 July 2016.
Ministry of Health Republic of Indonesia. Wilayah KLB DBD Ada di 11 Provinsi. http://www.depkes.go.id/article/print/16030700001/wilayah-klb-dbd-ada-di-11-provinsi.html. Accessed 29 July 2016.
Bull JJ, Turelli M. Wolbachia versus dengue: evolutionary forecasts. Evol Med Public Health. 2013;1:197–207.
Grisales N, Poupardin R, Gomez S, Fonseca-Gonzalez I, Ranson H, Lenhart A. Temephos resistance in Aedes aegypti in Colombia compromises dengue vector control. PLoS Negl Trop Dis. 2013;7:e2438.
Lima EP, Paiva MH, de Araujo AP, da Silva EV, da Silva UM, de Oliveira LN, et al. Insecticide resistance in Aedes aegypti populations from Ceara, Brazil. Parasit Vectors. 2011;4:5.
Linss JG, Brito LP, Garcia GA, Araki AS, Bruno RV, Lima JB, et al. Distribution and dissemination of the Val1016Ile and Phe1534Cys Kdr mutations in Aedes aegypti Brazilian natural populations. Parasit Vectors. 2014;7:25.
Harris AF, Rajatileka S, Ranson H. Pyrethroid resistance in Aedes aegypti from grand Cayman. Am J Trop Med Hyg. 2010;83:277–84.
Yanola J, Somboon P, Walton C, Nachaiwieng W, Somwang P, Prapanthadara LA. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Trop Med Int Health. 2011;16:501–9.
Kushwah RB, Dykes CL, Kapoor N, Adak T, Singh OP. Pyrethroid-resistance and presence of two knockdown resistance (kdr) mutations, F1534C and a novel mutation T1520I, in Indian Aedes aegypti. PLoS Negl Trop Dis. 2015;9:e3332.
Ishak IH, Jaal Z, Ranson H, Wondji CS. Contrasting patterns of insecticide resistance and knockdown resistance (kdr) in the dengue vectors Aedes aegypti and Aedes albopictus from Malaysia. Parasit Vectors. 2015;8:181.
Garcia GP, Flores AE, Fernandez-Salas I, Saavedra-Rodriguez K, Reyes-Solis G, Lozano-Fuentes S, et al. Recent rapid rise of a permethrin knock down resistance allele in Aedes aegypti in Mexico. PLoS Negl Trop Dis. 2009;3:e531.
Saavedra-Rodriguez K, Beaty M, Lozano-Fuentes S, Denham S, Garcia-Rejon J, Reyes-Solis G, et al. Local evolution of pyrethroid resistance offsets gene flow among Aedes aegypti collections in Yucatan State, Mexico. Am J Trop Med Hyg. 2014;92:201–9.
Li CX, Kaufman PE, Xue RD, Zhao MH, Wang G, Yan T, et al. Relationship between insecticide resistance and kdr mutations in the dengue vector Aedes aegypti in Southern China. Parasit Vectors. 2015;8:325.
Sayono S, Hidayati AP, Fahri S, Sumanto D, Dharmana E, Hadisaputro S, et al. Distribution of voltage-gated sodium channel (Nav) alleles among the Aedes aegypti populations in central java province and its association with resistance to pyrethroid insecticides. PLoS One. 2016;11:e0150577.
Wuliandari JR, Lee SF, White VL, Tantowijoyo W, Hoffmann AA, Endersby-Harshman NM. Association between three mutations, F1565C, V1023G and S996P, in the voltage-sensitive sodium channel gene and knockdown resistance in Aedes aegypti from Yogyakarta, Indonesia. Insects. 2015;6:658–85.
Du Y, Nomura Y, Satar G, Hu Z, Nauen R, He SY, et al. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc Natl Acad Sci USA. 2013;110:11785–90.
Brengues C, Hawkes NJ, Chandre F, McCarroll L, Duchon S, Guillet P, et al. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med Vet Entomol. 2003;17:87–94.
Saavedra-Rodriguez K, Urdaneta-Marquez L, Rajatileka S, Moulton M, Flores AE, Fernandez-Salas I, et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol Biol. 2007;16:785–98.
Kawada H, Higa Y, Komagata O, Kasai S, Tomita T, Thi Yen N, et al. Widespread distribution of a newly found point mutation in voltage-gated sodium channel in pyrethroid-resistant Aedes aegypti populations in Vietnam. PLoS Negl Trop Dis. 2009;3:e527.
Srisawat R, Komalamisra N, Apiwathnasorn C, Paeporn P, Roytrakul S, Rongsriyam Y, et al. Field-collected permethrin-resistant Aedes aegypti from central Thailand contain point mutations in the domain IIS6 of the sodium channel gene (KDR). Southeast Asian J Trop Med Public Health. 2012;43:1380–6.
Hirata K, Komagata O, Itokawa K, Yamamoto A, Tomita T, Kasai S. A single crossing-over event in voltage-sensitive Na + channel genes may cause critical failure of dengue mosquito control by insecticides. PLoS Negl Trop Dis. 2014;8:e3085.
Central Bureau of Statistic Republic of Indonesia. Indonesia Population Projection. http://www.bappenas.go.id/files/5413/9148/4109/Proyeksi_Penduduk_Indonesia_2010-2035.pdf.Accessed 29 July 2016.
WHO. Test procedures for insecticide resistance monitoring in malaria vector mosquitoes. Geneva: World Health Organization. 2013.
Brogdon WG, McAllister JC. Insecticide resistance and vector control. J Agromedicine. 2004;9:329–45.
Plernsub S, Saingamsook J, Yanola J, Lumjuan N, Tippawangkosol P, Sukontason K, et al. Additive effect of knockdown resistance mutations, S989P, V1016G and F1534C, in a heterozygous genotype conferring pyrethroid resistance in Aedes aegypti in Thailand. Parasit Vectors. 2016;9:417.
Stoddard ST, Forshey BM, Morrison AC, Paz-Soldan VA, Vazquez-Prokopec GM, Astete H, et al. House-to-house human movement drives dengue virus transmission. Proc Natl Acad Sci USA. 2013;110:994–9.
Nkya TE, Akhouayri I, Kisinza W, David JP. Impact of environment on mosquito response to pyrethroid insecticides: facts, evidences and prospects. Insect Biochem Mol Biol. 2013;43:407–16.
Platt N, Kwiatkowska RM, Irving H, Diabate A, Dabire R, Wondji CS. Target-site resistance mutations (kdr and RDL), but not metabolic resistance, negatively impact male mating competiveness in the malaria vector Anopheles gambiae. Heredity. 2015;115:243–52.
Arnaud L, Haubruge E. Insecticide resistance enhances male reproductive success in a beetle. Evolution. 2002;56:2435–44.
Stenhouse SA, Plernsub S, Yanola J, Lumjuan N, Dantrakool A, Choochote W, et al. Detection of the V1016G mutation in the voltage-gated sodium channel gene of Aedes aegypti (Diptera: Culicidae) by allele-specific PCR assay, and its distribution and effect on deltamethrin resistance in Thailand. Parasit Vectors. 2013;6:253.
Alvarez LC, Ponce G, Saavedra-Rodriguez K, Lopez B, Flores AE. Frequency of V1016I and F1534C mutations in the voltage-gated sodium channel gene in Aedes aegypti in Venezuela. Pest Manag Sci. 2015;71:863–9.
Olanratmanee P, Kittayapong P, Chansang C, Hoffmann AA, Weeks AR, Endersby NM. Population genetic structure of Aedes (Stegomyia) aegypti (L.) at a micro-spatial scale in Thailand: implications for a dengue suppression strategy. PLoS Negl Trop Dis. 2013;7:e1913.
Hemingway J, Hawkes NJ, McCarroll L, Ranson H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem Mol Biol. 2004;34:653–65.
Acknowledgements
We deeply thank the Bali Health Agency (BHA) for providing the fogging intensities of the years 2014–2016. Thanks are also extended to N. M. K. Dewi for managing mosquito samples from the city of Denpasar.
Funding
Experiments of this work were supported and financed by the DIKTI (Direktorat Jenderal Pendidikan Tinggi of Indonesia)-INSINAS research grant No. 226/SP2H/LT/DRPM/III/2016 awarded to PHH.
Availability of data and materials
The data supporting the conclusions of this article are included within the article.
Authors’ contributions
PH, JP, performed the experiments and analyzed the data. AW helped in performing the experiments. PH, CH, AT coordinated the work and wrote the manuscript. CH designed the study. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable. No ethical clearance was issued regarding these experiments as the subject of investigation was performed on Aedes mosquitoes. In addition, no mosquito blood-feedings were provided since exclusively F0 specimens were used for all experiments in this report. Placement of ovitraps on residential houses do not give potential risk and therefore do not require ethical clearance.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Hamid, P.H., Prastowo, J., Widyasari, A. et al. Knockdown resistance (kdr) of the voltage-gated sodium channel gene of Aedes aegypti population in Denpasar, Bali, Indonesia. Parasites Vectors 10, 283 (2017). https://doi.org/10.1186/s13071-017-2215-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-017-2215-4