
Abstract
Background
Pancreatic ductal adenocarcinoma (PDAC) is one of the deadliest tumors worldwide, with extremely aggressive and complicated biology. Krüppel-like factors (KLFs) encode a series of transcriptional regulatory proteins and play crucial roles in a variety of processes, including tumor cell differentiation and proliferation. However, the potential biological functions and possible pathways of KLFs in the progression of PDAC remain elusive.
Methods
We systematically evaluated the transcriptional variations and expression patterns of KLFs in pancreatic cancer from the UCSC Xena. Based on difference analysis, the non-negative matrix factorization (NMF) algorithm was utilized to identify the immune characteristics and clinical significance of two different subtypes. The multivariate Cox regression was used to construct the risk model and then explore the differences in tumor immune microenvironment (TIME) and drug sensitivity between high and low groups. Through single-cell RNA sequencing (scRNA-seq) analysis, we screened KLF6 and further investigated its biological functions in pancreatic cancer and pan-cancer.
Results
The KLFs exhibited differential expression and mutations in the transcriptomic profile of PDAC. According to the expression of KLFs, patients were classified into two distinct subtypes, each exhibiting significant differences in prognosis and TIME. Moreover, the KLF signature was developed using univariate Cox and Lasso regression, which proved to be a reliable and effective prognostic model. Furthermore, the KLF_Score was closely associated with immune infiltration, response to immunotherapy, and drug sensitivity and we screened small molecule compounds targeting prognostic genes separately. Through scRNA-seq analysis, KLF6 was selected to further demonstrate its role in the malignance of PC in vitro. Finally, pan-cancer analysis emphasized the biological significance of KLF6 in multiple types of tumors and its clinical utility in assessing cancer prognosis.
Conclusion
This study elucidated the pivotal role of KLF family genes in the malignant development of PC through comprehensive analysis and revealed that KLF6 would be a novel diagnostic biomolecule marker and potential therapeutic target for PDAC.
Similar content being viewed by others

Introduction
Pancreatic ductal adenocarcinoma (PDAC) is a common malignant cancer of the digestive tract and has ranked as one of the most lethal tumors [1]. Due to the rapid progression and ease of recurrence or metastasis of the tumor, as well as the absence of early diagnostic biomarkers, the 5-year survival rate for PC patients is less than 10% [2]. Currently, chemotherapy and surgery offer limited efficacy, especially in advanced cases. Thus, to improve the clinical outcomes of patients, identifying sensitive biomarkers for the early diagnosis and effective therapy of PDAC is urgently needed.
The Krüppel-like factors (KLFs) family contains 17 transcription factors, each with three conserved Cys2/His2 zinc finger domains in their C-terminal region [3]. They specifically bind to promoter and enhancer regions, regulating cell activities such as proliferation, apoptosis, differentiation, and embryonic development [4]. Multiple members of KLFs play key roles in mediating numerous signaling pathways to regulate the biological process [5]. Recently, increasing evidence has shown that dysregulated expression of KLFs is an important factor in the progression of several malignant tumors [6]. They could generally act as suppressors or oncogenes in different tumors. In primary hepatocellular carcinoma, loss of KLF4 promotes oncogenic TGF-β signaling by activating Samd7 to facilitate the malignant progression [7]. Additionally, KLF4 also inhibits the ERK/ JNK/ NF-κB signaling pathway in non-small cell lung cancer [8]. Conversely, KLF5 regulates cell cycle-related genes, promoting the cell cycle and cancer progression, including pancreatic, gastric, and breast cancers [9,10,11].
Tumor microenvironment (TME) is associated with the malignant progression of tumors, which are filled with various immune cells and stromal components [12]. KLFs also participate in regulating tumor immune microenvironment (TIME). It’s reported that KLF2 enhances B-cell differentiation and function by enhancing the expression of CD62L, CXCR7, and β7-harmonized protein [13]. KLF5 promotes the transcription of COX2 and increases the synthesis of PGE2, which binds to EP2 and EP4 receptors on the surface of CD8 + T-cells thereby inhibiting the function of CD8 + T-cells and shaping a suppressive immune microenvironment [14]. In summary, KLFs are crucial for the progression of tumors and the modulation of the immune microenvironment. However, the role of KLFs in PDAC progression remains clearly unelucidated.
In our study, we classified patients with PDAC into two subtypes based on differentially expressed KLFs. The immune microenvironmental differences between subtypes and the prognostic significance were assessed. Using univariate Cox regression and least absolute shrinkage and selection operator (LASSO)-Cox regression analysis, we developed a reliable risk score model and further investigated the associations between immune infiltration, KLF signature, immune checkpoints, and drug sensitivity. Also, the small-molecule drugs were screened by targeting the prognostic genes. Through single-cell RNA sequencing (scRNA-seq), we screened for KLF6 and explored its biological role in pancreatic cancer cells. Furthermore, we explored the biological and clinical relevance of KLF6 across different cancer types. Overall, we clarified the expression and prognostic significance of the KLF family in pancreatic cancer and provided new insights for future treatment.
Materials and methods
Data preparation and processing
The traditional RNA sequencing (Bulk RNA-seq) of PDAC patients and corresponding clinical characteristics and copy number variation (CNV) data were obtained from the UCSC Xena (https://xenabrowser.net/datapages/). The validation cohort GSE79668 was downloaded from the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/) [15]. The scRNA-seq GSE154778 was also gained from the GEO database, consisting of 10 primary tumor tissues and 6 metastatic lesions [16]. We investigated the mutation frequency and patterns of KLF family members using the cBioPortal database (https://www.cbioportal.org/).
Identification of differentially expressed genes and NMF algorithm analysis
We performed “DESeq2” R package to find differentially expressed genes (DEGs) of KLFs in PDAC. |log2Fold change (FC)|> 1 and p < 0.05 were considered as the filter criteria. According to the expression of differentially expressed KLFs, we utilized the “NMF” R package to explore molecular subtypes of patients with PDAC. The optimal number of clusters (k) was selected based on the cophenetic, dispersion, and silhouette metrics. Eventually, patients were divided into the most significant different molecular clusters when k = 2. The survival curve of two clusters was calculated by the “survival” R package. The “pheatmap” R package was used to visualize the differential expression KLFs and clinicopathological characteristics of the two subtypes.
Gene set variation analysis (GSVA) and Gene set enrichment analysis (GSEA)
The “clusterProfiler” and “GSVA” R packages were applied for functional enrichment analysis. The Kyoto Encyclopedia of Genes and Genomes (KEGG) database was used for GSVA analysis which regarded adjusted p < 0.05 as statistically significant enrichment. For the analysis of gene set enrichment, GSEA 4.2.3 was employed. Hallmark gene sets were collected from the MSigDB [17]. A p < 0.05 and |normalized enrichment score (NES) |> 1.5 were regarded to indicate significant differences between the two distinct subgroups.
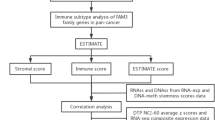
Immune landscape analysis
To compare the TIME between the different subgroups, we performed the CIBERSORT algorithm to calculate the scores of immune cells in 22 types, which derived the proportion of immune cell infiltration. Additionally, the stromal score, immune score, and estimate score were computed by the "estimate" R package. To predict the immunotherapy response in two subsets, the expression of programmed death-ligand 1 (PD-L1 or CD274), programmed death-1 (PD-1or CD279), and cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) were calculated.
Construction of the risk score model
Differentially expressed KLFs underwent univariate Cox regression to determine their prognostic relevance, and genes with p < 0.05 were selected (Table S1). The "glmnet" R package and the least absolute shrinkage and selection operator (LASSO) regression were used in conjunction to prevent overfitting. The multivariate Cox regression was subsequently utilized to assess the remaining genes, generating a risk score for each patient with PDAC (Table S2). The KLF_score formula was as follows: KLF_score = Σ (Expi * coefi). Patients were separated into high- and low-risk groups based on median values. Prognostic significance and discrimination of the risk score model were evaluated using the "survival" and "timeROC" R packages for Kaplan–Meier (K-M) survival analysis and time-dependent receiver operating characteristic (ROC) curve, respectively. Principal component analysis (PCA) was performed using the R tool "plotly". The TCGA Pancreatic Cancer (PAAD) dataset from UCSC Xena was utilized as the training set to develop the prognostic model, while GSE79668 served as an independent validation set to assess the accuracy and predictive performance of the model.
TIME analysis and prediction of immunotherapy response
A correlation analysis was used to explore the relationship between immune cell abundance and KLF_score. Additionally, immune processes were examined based on KLF_score, using single-sample gene set enrichment analysis (ssGSEA). Additionally, the effectiveness of immunotherapy was evaluated by calculating the expression of the immune checkpoint-related genes, including BTLA, BTNL2, CD160, CD200R1, CD244, CD27, CD274, CD276, CD28, CD40, CD40LG, CD44, CD48, CD80, CTLA4, HHLA2, ICOS, IDO1, IDO2, LAG3, LGALS9, NRP1, PDCD1, TIGIT, TMGD2, TNFRSF14, TNFRSF18, TNFRSF9, TNFSF14, TNFSF18, TNFSF4 and VTCN1. The Tumor Immune Dysfunction and Exclusion (TIDE), dysfunction, and tumor mutation burden (TMB) scores were used to predict immunotherapy response high TMB and low TIDE and dysfunction scores indicated a greater response to immunotherapy. The correlation between TMB and KLF_score and the potential of KLF_score to predict the response to immunotherapy were examined.
Drug sensitivity and screening
We employed the "oncoPredict" R package to assess the therapeutic efficacy variations of targeted therapy between the low- and high-risk categories. The Genomics of Drug Sensitivity in Cancer (GDSC) database was used to find data on drug sensitivity [18]. Additionally, we used Autodock for molecular docking to explore the interactions of small-molecule drugs with prognostic genes. We retrieved the list of drugs interacting with prognostic genes from the Comparative Toxicogenomics Database (CTD), and the structures of the small molecules from the PubChem database [19, 20]. Relevant biological macromolecular structures encoded by the prognostic genes were retrieved from the Uniport database. The molecule that interacted most strongly with the molecular target having the lowest binding energy was identified through automatic standard docking. PyMol was used for visualization.
Establishment and validation of the predictive nomogram
Clinical parameters (age, gender, etc.) and KLF_score underwent univariate Cox and multivariate Cox regression to build an enhanced prediction model. Both univariate and multivariate Cox regressions yielded independent prognostic factors with p < 0.05. We used the "regplot" R package to create an interactive nomogram that predicted the 1-, 3-, and 5-year overall survival. Calibration curves gauged the precision of the prediction, while K-M survival analysis and time-dependent ROC curves assessed relevance and discrimination for the 1-, 3-, and 5-year overall survival. Model accuracy was confirmed by the validation dataset GSE79668.
ScRNA-seq analysis
GSE154778 was performed using the Seurat (v4.1.1) in R. The primary processes in the analysis pipeline include object construction, data normalization, data downscaling, clustering, and marker gene identification. Cell-type annotation utilized a standard Seurat process, referencing a prior study and the CellMarker database [21].
Pan-cancer analysis
To further investigate the biological importance of KLF6 in various cancers, we assessed its differential expression in pan-cancer. We also analyzed its clinical prognostic value and its association with TMB and microsatellite instability (MSI). Additionally, the co-expression analysis was performed to investigate the association between KLF6 and immunostimulators and immunoinhibitors.
Clinical samples collection
20 pairs of samples from patients with PDAC were collected from Ruijin Hospital affiliated to School of Medicine, Shanghai Jiaotong University. The procedure was approved by the Medical Ethics Committee of the hospital. Before collecting the samples, we obtained informed consent from each patient. Each sample contained a tumor and adjacent normal tissue. All patients had histopathologic confirmation of PDAC and had never received chemotherapy. Samples were used for RNA extraction.
Cell culture and transfection
Pancreatic cancer cell lines (PATU-8988, PANC-1, hTERT-HPNE, ASPC-1, and CAPAN-1) were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). They were cultured in RMPI 1640 with 10% fetal bovine serum and 1% Penicillin streptomycin. Short hairpin RNAs (shRNA) targeting KLF6 were mixed with HilyMax (Dojindo) for 15 min, and the mixture was added to the supernatant to achieve transient transfection. All cells were cultured at 37 °C and 95% humidity, in a 5% CO2 atmosphere. The protein expression level was assessed 3 days later.
Western blotting
Cells were lysed using RIPA buffer (Abclonal, Wuhan, China) containing 1% protease and phosphatase inhibitors (MCN Biotech). The mixture was set on ice for 10 min and heated to 100 °C for 30 min. The obtained proteins were separated using 10% sodium dodecyl-sulfate polyacrylamide gel electrophoresis, transferred to polyvinylidene fluoride membranes, and blocked. The membranes were incubated overnight at 4 °C with primary antibodies, including anti-KLF6 (abclonal, A10011) and anti-β-Actin (abclonal, AC004) antibodies. The next day, the membranes were incubated with a secondary antibody. An enhanced chemiluminescence (ECL) detection system was used for the visualization of the proteins.
Cell proliferation assay
Cell activity was measured using Cell Counting Kit-8 (CCK-8) (Meilunbio, China), according to the manufacturer’s instructions. Transfected cells (2000/well) were seeded in 96-well plates. Prior to the assay, 90 μL complete RMPI 1640 and 10 μL CCK-8 were added to each well and incubated at 37 °C for 2 h. Absorbance at 450 nm was measured, representing cell activity.
For the colony formation assay, 1000 tumor cells or transfected cells were cultured in 6-well plates for 10 days. On day 10, the cell supernatant was discarded, and cells were stained with a crystal violet staining solution and subsequently imaged.
Patient-derived organoid (PDO) construction
According to the instruction of the human Tumor Dissociation Kit (Miltenyi), organoids were extracted and digested from tumor tissues of PDAC patients who had not received chemotherapy at Ruijin Hospital. Organoids were seeded into the Matrigel (Corning) and then cultured in OmaStem® Pan-cancer Advanced (OmaStem). After transfecting the plasmids, organoids were digested by TrypLETM Express (ThermoFisher) and seeded in 96-well plates. Thereafter, representative images were obtained every 5 days, and proliferative activity was assayed by CellTiter-Glo® 3D Cell Viability Assay (Promega).
Transwell assay
For migration, treated and control cells were suspended in 200 μl serum-free DMEM in the upper chamber with 2 × 104 cells per well. As for invasion, the upper portion of plates was covered with Matrigel (Corning, NY, USA). In addition,700 μl complete RMPI 1640 in the lower chamber was available. After 48 h, cells passing through membranes were fixed with 4% PFA stained with 1% crystal violet and counted under the microscope.
RNA extraction and Quantitative Real-Time PCR (qRT-PCR)
Total RNA was extracted using TRIzol reagent (Invitrogen, USA). HiScript Reverse Transcriptase (Vazyme, China) was used to conduct reverse transcription. SYBRGreen PCR Kit (Vazyme, China) was used for RT-PCR, with β-actin as an internal control. The following primer pairs were used:
KLF6: GGCAACAGACCTGCCTAGAG (Forward Sequence).
KLF6: CTCCCGAGCCAGAATGATTTT (Reverse Sequence).
β-Actin: GTCATTCCAAATATGAGATGCGT (Forward Sequence).
β-Actin: GCTATCACCTCCCCTGTGTG (Reverse Sequence).
Immunohistochemistry (IHC)
The tissues underwent paraffin embedding, formalin fixation, and sectioning onto slides. After that, the conventional streptavidin–biotin-peroxidase complex method was used for IHC staining. The slides were deparaffinized and rehydrated. Subsequently, they underwent antigen retrieval, inactivation, primary and secondary antibody incubation, DAB staining, and sealing. Ultimately, representative images were taken under a microscope, and the area and intensity of staining were used to calculate IHC scores.
Statistical analysis
All statistical tests and analyses were conducted using GraphPad Prism 7 and R (version 4.1.1). Clustering analysis was performed using the "NMF" R package. Survival analysis employed the Kaplan–Meier (KM) method and log-rank tests from the "survival" R package. The prognosis was assessed using the area under the ROC curve (AUC) via the "timeROC" R program. Correlation analysis was performed using the Spearman correlation test. The Student’s two-tailed unpaired t-test and one-way ANOVA were used for statistical analyses. Statistical significance was set as P < 0.05.
Results
Identification of differently expressed KLFs in PDAC
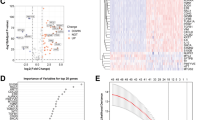
To elucidate the transcriptome profiles of the KLF family in pancreatic cancer, we accessed mRNA expression data from the UCSC Xena database. Using | log2FC |> 1 and p-value < 0.05 as criteria for differential expression, we identified significant expression differences among most KLFs except for KLF11, KLF12, and KLF16 (Fig. 1A, B). Spearman analysis revealed strong correlations within the KLF family, with the strongest correlation being detected between KLF3 and KLF5, which indicated potential synergistic interactions among KLF family members (Fig. 1C). Next, to explore the genetic variation of KLFs in PDAC, we evaluated CNV and somatic mutation. As shown in Fig. 1D, some KLFs, including KLF7, KLF10, KLF5, KLF3, KLF2, and KLF17 exhibited increased CNV, while KLF6, KLF14, KLF9, KLF4, KLF1, KLF15, KLF13, and KLF8 displayed decreased CNV. Notably, we found higher mutation frequencies in KLF10, KLF5, KLF4, KLF7, KLF8, KLF3, KLF6, KLF14, KLF15, and KLF17 among patients with PDAC (Fig. 1E). The above results illustrated the robust connections between the transcriptomic landscape and the mRNA expression level of KLFs.

Assessment of KLF family expression and genetic variations in PDAC. A, B The volcano gram and box plots showed differentially expressed KLFs in tumor tissues and adjacent normal tissues C Expression correlation analysis of KLFs D The CNV frequency of KLFs in PDAC. The frequency of the changes was indicated by the height of the bars. The green dot denoted CNV deletions (loss), whereas pink indicated CNV amplification (gain). E Mutation frequency and type of KLFs. (*p < 0.05; **p < 0.01; ***p < 0.001; ns no significance)
NMF clustering identifying the molecular subtypes of KLFs
Based on the differentially expressed KLFs, NMF clustering was performed and the optimal number of clusters was 2 (Fig. S1A,B). Hence, we separated patients into two clusters (Fig. 2A). It could be observed that the prognosis of patients in cluster 1 was worse than that of cluster 2 patients following the survival curve (Fig. 2B). PCA analysis indicated consistent and stable subtypes in the distribution of cluster 1 and cluster 2 (Fig. 2C). The heatmap illustrated the associations between KLF expression patterns within the clusters and their corresponding clinicopathological features (Fig. 2D). To demonstrate the potential underlying biological pathways, GSVA and GSEA were performed. As shown in Fig. 2E, cluster 1 was notably enriched in KEGG_RNA_POLYMERASE, KEGG_PROTEASOME, and KEGG_DNA_REPLICATION, while cluster 2 was markedly enriched in immune-related pathways, such as KEGG_CHEMOKINE_SIGNALING_PATHWAY and KEGG_T_CELL_RECEPTOR_SIGNALING_PATHWAY. Furthermore, GSEA indicated that cluster 1 was influenced by HALLMARK_GLYCOLYSIS, HALLMARK_HYPOXIA, HALLMARK_P53_PATHWAY, and other pathways (Fig. 2F–K). These findings revealed a distinct pattern, indicating that patients in cluster 1 exhibited heightened activation of tumor-related pathways, whereas patients in cluster 2 demonstrated greater activation of immune-related pathways. Consequently, it was deduced that cluster 1 displayed a more pronounced pro-tumorigenic activity, whereas cluster 2 exhibited enhanced immune activity. This disparity in pathway activation plausibly accounted for the divergent prognoses observed between these two groups.

Classifying pancreatic cancer patients based on differentially expressed KLFs. A The non-negative matrix decomposition algorithm classified patients into two distinct subgroups (C1 and C2). B Kaplan–Meier plots compared the overall survival of the two subgroups (log-rank p = 0.043). CThe two subgroups were clearly distinguished on PCA analysis. DThe heatmap displayed the expression of KLFs in clusters 1 and 2, as well as the associations between clinicopathological characteristics and different subtypes. EThe enrichment analysis between cluster 1 and cluster 2. Higher pathway richness was indicated by a yellow pathway, whereas lower pathway enrichment was indicated by a blue pathway. Statistical significance is defined as an adjusted p < 0.05. F–K GSEA revealed the enriched pathways in patients with different subtypes
TIME analysis of molecular subtypes
As a crucial factor in mediating tumor malignant progression, TIME is implicated in tumor metastasis, immune escape, recurrence, and other processe [22]. Therefore, we hypothesized that there were distinct immunological characteristics between the two subtypes. To confirm this hypothesis, immune infiltration was assessed, and the results revealed that significant enrichment of B cells naïve, T cells CD8, and T cells CD4 memory resting in cluster 2 (Fig. 3A). Furthermore, higher immune score, stromal score, and ESTIMATE score were represented in cluster 2, implying patients in cluster 2 had a more robust immune response and a lower purity of tumor cells in TIME (Fig. 3B–D). Given the pivotal role of the immune checkpoint in tumor immunotherapy, we found that PD-1 and CTLA-4 had significantly higher levels of expression in cluster 2 compared to cluster 1 (Fig. 3E–G). In short, these findings revealed that patients in cluster 2 exhibited immune activation traits, while patients in cluster 1 displayed immunosuppressive features, which suggested that the KLF pattern could better guide patient subtyping and immunotherapy in PDAC.

Immune landscape of patients with PDAC in different subgroups. A The CIBERSORT algorithm calculated the abundance of each immune infiltrating cell in cluster 1 and cluster 2. B–D ESTIMATE algorithm evaluated stromal score, immune score, and ESTIMATE score. E–G Expression levels of immune checkpoints in cluster 1 and cluster 2. (*p < 0.05; **p < 0.01; ***p < 0.001; ns no significance)
Construction and validation of the prognostic model
To further explore the relationship between KLFs and the prognosis of patients with pancreatic cancer, univariate Cox regression was performed, and we identified 4 significantly prognostic genes (KLF3, KLF4, KLF5, KLF6) with p < 0.05 (Fig. 4A). Furthermore, we utilized the LASSO-Cox regression to establish a model (including KLF3, KLF5, and KLF6) named the KLF_score model (Fig. 4B, C). The model assigned each patient with PDAC a risk score using the following formula: (0.129498515* expression of KLF3 + 0.27195101* expression of KLF5 + 0.246696073* expression of KLF6). Patients were classified into high- and low-risk groups based on their median risk score. In both the training and validation sets, patients with high-risk scores had significantly shorter overall survival (Fig. 4D, G). Time-dependent ROC curves demonstrated the model's robustness in the training cohort (5-year AUC, 0.722; 3-year AUC, 0.697; 1-year AUC, 0.613; Fig. 4E) and validation cohort (5-year AUC, 0.938; 3-year AUC, 0.765; 1-year AUC, 0.694; Fig. 4H). PCA and t-SNE effectively differentiated the two risk subgroups (Fig. 4F, I), indicating a clear separation of patients into distinct parts (low- or high-risk) with high discriminatory power.

Establishment and verification of KLF signature. A Forest plot demonstrated survival-associated KLFs screened by univariate Cox regression analysis. B LASSO regression was performed to obtain the prognostic genes. C Optimal lambda (λ) was selected based on the cross-validation error curve. D Kaplan–Meier survival analysis in the training cohort. E The time-dependent ROC curve in the training cohort. F PCA analysis in the training cohort. G Survival analysis in the validation cohort. H The time-dependent ROC curve in the validation cohort. I PCA analysis in the validation cohort
The biological contribution of KLF_score
To better understand the biological functions of the two risk groups, GSVA was performed. As shown in Fig S2A, the high-risk group exhibited significant activation of tumor-associated pathways, including the p53 pathway and phospholipid metabolism, while these pathways were less expressed in the low-risk group. Moreover, GSEA confirmed the activation of tumor-associated pathways in the high-risk group (Fig. S2B-G). These results collectively indicated an intimate link between KLF_score and tumor metabolism and proliferation. A higher score indicated heightened tumor activity, which could better explain the worse prognosis observed in the high-risk group.
Immune characteristics in relation to KLF_score
A mesenchymal space of immune cells exists within pancreatic cancer, including immune cells, extracellular matrix, fibroblasts, and growth factors, which together constitute a suppressive TIME [23]. TIME plays an important role in tumor progression and chemoresistance in PDAC [24]. Thus, we explored potential differences in the TIME across risk groups. Through correlation analysis and assessment of immune processes, we found an inverse relationship between KLF_score and immune cell abundance, suggesting that a lower KLF_score presented a higher immune activity and more activated immune functions (Fig. 5A–E). In other words, the patients in the high-risk group were suffering from an immunosuppressive status. Additionally, the expressions of immune checkpoints were examined, and the results revealed that CD44, TNFRSF14, HHLA2, and LGALS9 were highly expressed in the high-risk group (Fig. 5F). Moreover, as seen in Fig. 5G–J, the patients in the high-risk group displayed lower TIDE and dysfunction scores and a higher TMB score, which indicated they would be better applied immunotherapy to release immunosuppression and restore immune activity.

Correlation analysis between KLF_score and tumor immune microenvironment. A–D Correlation of risk score and immune cells. E The boxplot showed the different immune-related functions in the high-risk group and low-risk group. F The expression of immune checkpoints between high-risk group and low-risk group. G–I TIDE score, dysfunction score, and TMB score of high- and low-risk groups. J Correlation of risk score and TMB. (*p < 0.05; **p < 0.01; ***p < 0.001; ns no significance)
Drug sensitivity prediction and small-molecule drug screening
Previous study has shown that patients eventually obtained longer survival time when treated by combining chemotherapy with immunotherapy or small molecule targeted drugs [25]. To better guide clinical treatment, we used “OncoPredict” to predict the drug sensitivity of PDAC patients and observed the marked difference between the high-risk and low-risk groups. Patients with low KLF_score were sensitive to gefitinib, bosutinib, cytarabine, nilotinib, pazopanib, sunitinib, and axitinib, while the high-risk group responded better to bleomycin and paclitaxel, which highlighted the relevance of KLF_score to drug sensitivity (Fig. S3A-L). Additionally, we screened the CTD and employed molecular docking and a pharmacological toxicology study to identify three small-molecule drugs targeting prognostic genes. As seen in Fig. 6A, estradiol exhibited a strong binding energy of -7.6 (kcal/mol) with KLF3, potentially suppressing its transcription. Vorinostat was better at lowering KLF6 mRNA expression levels and presented an optimal binding energy of -6.5 (kcal/mol) (Fig. 6B). Tamibarotene exhibited a high binding energy of -8.4 (kcal/mol), indicating potential in inhibiting KLF5 mRNA expression (Fig. 6C).

Small molecule drug screening. A–C 3D structure drawings of molecular docking for KLF3-Estradiol, KLF5-Tamibarotene and KLF6-Vorinostat
Construction of a nomogram and evaluation of the prognostic model
Clinical characteristics are inseparably related to the patient’s prognosis [26]. To determine whether the risk score and other clinical parameters can independently predict pancreatic cancer prognosis, we conducted univariate and multifactor Cox regression and uncovered that age, N stage, and risk score are independent prognostic factors (Fig. 7A). According to the above findings, an interactive nomogram was performed for predicting 1-, 3-, and 5-year overall survival (Fig. 7B). As shown in Fig. 7C, D, the survival of high- and low-risk groups displayed significant differences, with AUC values of 0.616, 0.758, and 0.639 at 1, 3, and 5 years, respectively. Calibration curves confirmed the accuracy and reliability of the model (Fig. 7E). In the validation set, the K-M curve also indicated significantly improved overall survival in the low-risk group, aligning with the training set result (Fig. 7F). Consistent ROC and calibration curves further confirmed the reliability and stability of the prediction model (Fig. 7G, H).

Establishment and evaluation of survival prediction nomogram. A Multivariate Cox regression analysis between the risk score and clinical characteristics. B A nomogram was established to predict 1-, 3-, and 5-year overall survival based on risk score and clinical features. C The comparison of overall survival between high- and low-risk groups in K-M plots. D Time-dependent ROC curves for predicting 1-, 3- and 5-year overall survival according to KLF_score. E Calibration curves implied the accuracy of 1-, 3-, and 5-year survival. F–H The results of K-M plots, ROC curves, and calibration curves in the validation cohort
ScRNA-seq dataset analysis of key genes
To further determine the crucial gene of KLFs in the PDAC, we obtained the GSE154778 dataset from GEO and conducted a single-cell analysis. Cell grouping and annotation were performed in a descending manner based on t-SNE (Fig. 8A). As described in Fig. 8B, C, KLF6 exhibited significantly higher expression levels than KLF3 and KLF5 in epithelial tumor cells (ETCs). Moreover, it could be observed that KLF6 was highly expressed in other immune cells, suggesting it may affect PDAC progression in multiple aspects.

ScRNA-seq analysis for screening key genes. A The result of t-SNE dimensionality reduction and cell clusters annotation. B–C The expression and distribution of KLF3, KLF5, and KLF6 in different cell types by Featureplot and ridge map visualization
Biological functions of KLF6 in PDAC
To validate the role of KLF6 in the progression of PDAC, we collected 20 paired tumor tissues and non-tumor tissues from Ruijin Hospital to examine the expression of KLF6. RT-qPCR analysis revealed that the expression level of KLF6 was higher in tumor tissues and the IHC was further verified that KLF6 was infiltrated more in PDAC (Fig. 9A–C). In pancreatic cancer cells, KLF6 was expressed higher in PATU-8988 and PANC-1 cell lines compared to others (Fig. 9D). Next, the pro-tumorigenic functions of KLF6 were explored. After knocking down KLF6 in PATU-8988 and PANC1 cell lines, as expected, the proliferative activity was remarkedly suppressed (Fig. 9E–J). Moreover, similar results were observed in organoid proliferation (Fig. 9K, L and Figure S4A-D). Subsequently, the transwell assays determined that KLF6 enhanced the migratory and invasive abilities in pancreatic cancer cells (Fig. 9M–P). Above all, these results suggested that KLF6 may contribute to the progression of PDAC by promoting proliferation and metastasis activities.

KLF6 promoted malignant progression of pancreatic cancer in vitro. A The expression of KLF6 in tumor and adjacent tissues collected from 20 PDAC patients in Ruijin Hospital. B IHC staining images of KLF6 in clinical samples. C The IHC score revealed the infiltration of KLF6 in tumor and normal tissues. D Relative expression of KLF6 in pancreatic cancer cell lines and normal epithelial cell line. E The protein levels of KLF6 after being knocked down in PATU-8988 and PANC1. F–G CCK-8 assays revealed the proliferative activity of KLF6 in PDAC. H–J Plate colony assays in PATU-8988 and PANC1 after transfection of shNC, sh1-KLF6, and sh2-KLF6. K–L The proliferative capacity of KLF6 in different patient-derived organoids on day 0, day 5, and day 10. M–P Transwell assays demonstrated the migration and invasion abilities of KLF6 in PDAC. (*p < 0.05; **p < 0.01; ***p < 0.001)
Pan-cancer analysis of KLF6
To investigate the potential value of KLF6 in other tumors, pan-cancer analysis was performed. As shown in Fig. 10A, KLF6 was significantly and differentially expressed across multiple tumor types. Moreover, KLF6 was most highly expressed in kidney renal clear cell carcinoma tissues (Fig. 10B).

Analysis of KLF6 in pan-cancer level. A The expression of KLF6 in tumor and normal tissues of multiple cancers. B Relative expression of KLF6 in pan-cancer. C–P The overall survival (OS), disease-free survival (DFS), disease-specific survival (DSS), and progression-free survival (PFS) analysis of KLF6 in pan-cancer. (*p < 0.05; **p < 0.01; ***p < 0.001)
In addition, KLF6 was closely related to overall survival, disease-free survival, disease-specific survival, and progression-free survival in pan-cancer (Fig. 10C–F). Further survival curves analysis confirmed it (Fig. 10G–P). Subsequently, we assessed the impact of KLF6 in TIME of multiple cancers and found that the KLF6 was closely relevant to the immune microenvironment in a range of tumors (Fig. S5A-Q). Notably, KLF6 is also associated with TMB and MSI in various types of cancer (Fig. S5R, S). Furthermore, as Fig. S5T, U demonstrated, KLF6 may act as a key factor in regulating the immune activity of pan-cancer.
Discussion
Pancreatic cancer is a global health issue, characterized by rising incidence and mortality rates alongside limited clinical treatment options and low survival rates [26]. The need for novel biomarkers to facilitate early diagnosis and tailored therapies, and to improve prognosis remains critical and urgent.
KLFs, a family of 17 transcription factors with highly conserved zinc-finger structural domains, play crucial roles in tumor development, influencing proliferation, apoptosis, metastasis, and survival [6]. Despite understanding individual KLF genes’ impact on tumor progression, the potential interconnections within the KLF family in pancreatic cancer remain unexplored [27]. Consequently, this study aimed to explore the clinical value of whole KLF family genes in pancreatic cancer.
First, we investigated transcriptional differences and potential interactions among KLFs. Subsequently, according to differentially expressed KLFs, 178 patients with PDAC were classified into two distinct molecular phenotypes, which revealed significant differences both in the prognosis and clinicopathological characteristics. Enrichment analysis demonstrated that patients in cluster 1 exhibited increased activation of cancer-related pathways, including hypoxia, glycolysis, and the p53 pathway. TIME is infiltrated by a variety of immune cells, among which CD8 + or cytotoxic T lymphocytes exert tumor-killing function, while regulatory T cells attenuate effector T cell activity and promote immunosuppression in TIME [28, 29]. Different immune cell ratios could promote tumor growth and metastasis by shaping TIME and thus mediating the immune escape of tumor cells [30]. Cluster 1 showed a lower proportion of naive B cells infiltrated with CD8+ T cells. Furthermore, notable variations in stromal score, immune score, and estimate score were observed, indicating cluster 1 exhibited an immunosuppressive status and validating the prognostic differences between subgroups.
To enhance the clinical relevance of our study, we screened three genes (KLF3, KLF5, and KLF6) to construct the KLF_score prognostic model using Cox and LASSO regression analyses. KLF3 was considered a novel tumor-promoting transcription factor in colorectal cancer [31]. Specifically, highly expressed in ovarian cancer, KLF5 facilitated the process of ovarian cancer proliferation and metastasis and promoted PARP inhibitor resistance by remodeling the transcription of the key gene for homologous recombination, RAD51 [32]. KLF6 was a tumor suppressor and inhibited the progression of a variety of tumors [33,34,35]. Based on the median KLF_score, patients were divided into high- and low-risk groups. The high-risk group displayed a worse prognosis than the low-risk group, along with heightened activity in cancer-related pathways such as glucose metabolism, p53, and the E2F pathway. Validation via K-M-plots and ROC curves of the validation cohort confirmed the robustness and reliability of the KLF_score signature. Additionally, immune checkpoint expression, immune pathway enrichment, TIDE scores, and TMB scores differed significantly between the high- and low-risk groups, suggesting higher immunological activation in the low-risk group, which aligns with their better overall survival. At the same time, it also meant that patients in the high-risk group were more suitable for immunotherapy to release their own immunosuppression and restore their immune activity, thus improving the anti-tumor effect.
Combination therapies usually work better than monotherapies, and this is especially true for cancer treatments based on checkpoint blockade with immunosuppression [36]. The drug sensitivity analysis could be better guidance in clinical treatment. Additionally, to screen for novel and potential targeted drugs, we utilized molecular docking and found three drugs that targeted KLF3, KLF5, and KLF6. These findings could provide valuable clinical relevance, guiding the choice of chemotherapy and targeted immunotherapy. Clinical characteristics impact prognosis. Therefore, we performed a multifactorial Cox analysis integrating clinical characteristics with the risk signature. Age, risk score, and N stage were independent prognostic factors in PDAC. The resultant graphic and calibration curves demonstrated the significant and efficient predictive ability of KLF score.
To further investigate the impact of key genes on tumor progression, we performed single-cell analysis and found KLF6 was highly expressed in immune cells and strongly elevated in tumor cells. Then, we assessed the biological role of KLF6 in PDAC. KLF6 expression was highly elevated in pancreatic cancer samples compared to adjacent normal tissue samples. The proliferation and metastatic capacity were significantly suppressed by the downregulation of KLF6 in pancreatic cancer cells. Moreover, the clinical value of KLF6 was assessed at the pan-cancer level.
Although we developed a KLF_score to serve as a reliable predictor of pancreatic cancer prognosis and demonstrated the contribution of KLF6 to the development of the disease, our study has some limitations. First, the dataset employment had a small sample size. In some cases, the clinical characteristics of those patients were incomplete. Future validation using larger datasets from multiple sites is needed. Second, the mechanisms underlying the impact of KLF6 on pancreatic cancer progression are still unknown. Moreover, single-cell sequencing data revealed that KLF6 was strongly expressed in tumor immune cells, such as macrophages, indicating that KLF6 may participate in remodeling TIME, enhancing the progression of pancreatic cancer. These challenges warrant future exploration.
Conclusion
In general, our study illustrated the clinical relevance and significance, tumor microenvironment characteristics, and immune infiltration features of the KLF family in pancreatic cancer. In addition, a reliable and accurate KLF gene-based prognostic model was presented. Further cellular analysis on KLF6 confirmed the role it plays in the malignancy of pancreatic cancer. These findings might be useful for early diagnosis and targeted treatment on pancreatic cancer.
Availability of data and materials
The datasets presented in this study can be found in the UCSC Xena and GEO databases.
Abbreviations
- PDAC:
-
Pancreatic ductal adenocarcinoma
- KLFs:
-
Krüppel-like factors
- NMF:
-
Non-negative matrix factorization
- TIME:
-
Tumor immune microenvironment
- LASSO:
-
Least absolute shrinkage and selection operator
- scRNA-seq:
-
Single-cell RNA sequencing
- CNV:
-
Copy number variation
- DEGs:
-
Differentially expressed genes
- GSVA:
-
Gene set variation analysis
- GSEA:
-
Gene set enrichment analysis
- ROC:
-
Receiver operating characteristic
- PCA:
-
Principal component analysis
- TMB:
-
Tumor mutation burden
- MSI:
-
Microsatellite instability
- AUC:
-
Area under the ROC curve
- K-M:
-
Kaplan–Meier
- PDO:
-
Patient-derived organoid
- ETCs:
-
Epithelial tumor cells
References
Vincent A, Herman J, Schulick R, Hruban RH, Goggins M. Pancreatic cancer. Lancet. 2011;378(9791):607–20.
Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73(1):17–48.
McConnell BB, Yang VW. Mammalian Krüppel-like factors in health and diseases. Physiol Rev. 2010;90(4):1337–81.
Ilsley MD, Gillinder KR, Magor GW, Huang S, Bailey TL, Crossley M, Perkins AC. Krüppel-like factors compete for promoters and enhancers to fine-tune transcription. Nucleic Acids Res. 2017;45(11):6572–88.
Dang DT, Pevsner J, Yang VW. The biology of the mammalian Krüppel-like family of transcription factors. Int J Biochem Cell Biol. 2000;32(11–12):1103–21.
Tetreault M-P, Yang Y, Katz JP. Krüppel-like factors in cancer. Nat Rev Cancer. 2013;13(10):701–13.
Sun H, Peng Z, Tang H, Xie D, Jia Z, Zhong L, Zhao S, Ma Z, Gao Y, Zeng L, et al. Loss of KLF4 and consequential downregulation of Smad7 exacerbate oncogenic TGF-β signaling in and promote progression of hepatocellular carcinoma. Oncogene. 2017;36(21):2957–68.
Yu R, Han L, Ni X, Wang M, Xue P, Zhang L, Yuan M. Kruppel-like factor 4 inhibits non-small cell lung cancer cell growth and aggressiveness by stimulating transforming growth factor-β1-meidated ERK/JNK/NF-κB signaling pathways. Tumour Biol. 2017;39(6):1010428317705574.
Liu P, Wang Z, Ou X, Wu P, Zhang Y, Wu S, Xiao X, Li Y, Ye F, Tang H. The FUS/circEZH2/KLF5/ feedback loop contributes to CXCR4-induced liver metastasis of breast cancer by enhancing epithelial-mesenchymal transition. Mol Cancer. 2022;21(1):198.
Xu T-P, Ma P, Wang W-Y, Shuai Y, Wang Y-F, Yu T, Xia R, Shu Y-Q. KLF5 and MYC modulated LINC00346 contributes to gastric cancer progression through acting as a competing endogeous RNA and indicates poor outcome. Cell Death Differ. 2019;26(11):2179–93.
He P, Yang JW, Yang VW, Bialkowska AB. Krüppel-like factor 5, increased in pancreatic ductal adenocarcinoma, promotes proliferation, acinar-to-ductal metaplasia, pancreatic intraepithelial neoplasia, and tumor growth in mice. Gastroenterology. 2018;154(5):1494–508.e13.
Ho WJ, Jaffee EM, Zheng L. The tumour microenvironment in pancreatic cancer—clinical challenges and opportunities. Nat Rev Clin Oncol. 2020;17(9):527–40.
Winkelmann R, Sandrock L, Porstner M, Roth E, Mathews M, Hobeika E, Reth M, Kahn ML, Schuh W, Jäck H-M. B cell homeostasis and plasma cell homing controlled by Krüppel-like factor 2. Proc Natl Acad Sci U S A. 2011;108(2):710–5.
Wu Q, Liu Z, Gao Z, Luo Y, Li F, Yang C, Wang T, Meng X, Chen H, Li J, et al. KLF5 inhibition potentiates anti-PD1 efficacy by enhancing CD8+ T-cell-dependent antitumor immunity. Theranostics. 2023;13(4):1381–400.
Kirby MK, Ramaker RC, Gertz J, Davis NS, Johnston BE, Oliver PG, et al. RNA sequencing of pancreatic adenocarcinoma tumors yields novel expression patterns associated with long-term survival and reveals a role for ANGPTL4. Mol Oncol. 2016;10:1169–82.
Lin W, Noel P, Borazanci EH, Lee J, Amini A, Han IW, Heo JS, Jameson GS, Fraser C, Steinbach M, Woo Y, Fong Y, Cridebring D, Von Hoff DD, Park JO, Han H. Single-cell transcriptome analysis of tumor and stromal compartments of pancreatic ductal adenocarcinoma primary tumors and metastatic lesions. Genome Med 2020;12(1):80. https://doi.org/10.1186/s13073-020-00776-9.
Liberzon A, Birger C, Thorvaldsdóttir H, Ghandi M, Mesirov JP, Tamayo P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015;1:417–25.
Yang W, Soares J, Greninger P, Edelman EJ, Lightfoot H, Forbes S, Bindal N, Beare D, Smith JA, Thompson IR, et al. Genomics of Drug Sensitivity in Cancer (GDSC): a resource for therapeutic biomarker discovery in cancer cells. Nucleic Acids Res. 2013;41:D955–61.
Davis AP, Wiegers TC, Johnson RJ, Sciaky D, Wiegers J, Mattingly CJ. Comparative toxicogenomics database (CTD): update 2023. Nucleic Acids Res. 2023;51(D1):D1257–62.
Kim S, Chen J, Cheng T, Gindulyte A, He J, He S, Li Q, Shoemaker BA, Thiessen PA, Yu B, et al. PubChem 2023 update. Nucleic Acids Res. 2023;51(D1):D1373–80.
Hu C, Li T, Xu Y, Zhang X, Li F, Bai J, Chen J, Jiang W, Yang K, Ou Q, et al. Cell Marker 2.0: an updated database of manually curated cell markers in human/mouse and web tools based on scRNA-seq data. Nucleic Acids Res. 2023;51:D870–6.
Raghavan S, Winter PS, Navia AW, Williams HL, DenAdel A, Lowder KE, Galvez-Reyes J, Kalekar RL, Mulugeta N, Kapner KS, et al. Microenvironment drives cell state, plasticity, and drug response in pancreatic cancer. Cell. 2021;184(25):6119–37.e26.
Connor AA, Gallinger S. Pancreatic cancer evolution and heterogeneity: integrating omics and clinical data. Nat Rev Cancer. 2022;22(3):131–42.
Fan J-Q, Wang M-F, Chen H-L, Shang D, Das JK, Song J. Current advances and outlooks in immunotherapy for pancreatic ductal adenocarcinoma. Mol Cancer. 2020;19(1):32.
Padrón LJ, Maurer DM, O’Hara MH, O’Reilly EM, Wolff RA, Wainberg ZA, Ko AH, Fisher G, Rahma O, Lyman JP, et al. Sotigalimab and/or nivolumab with chemotherapy in first-line metastatic pancreatic cancer: clinical and immunologic analyses from the randomized phase 2 PRINCE trial. Nat Med. 2022;28(6):1167–77.
Topham JT, Karasinska JM, Lee MKC, Csizmok V, Williamson LM, Jang GH, Denroche RE, Tsang ES, Kalloger SE, Wong H-L, et al. Subtype-discordant pancreatic ductal adenocarcinoma tumors show intermediate clinical and molecular characteristics. Clin Cancer Res. 2021;27(1):150–7.
Zeng L, Zhu Y, Moreno CS, Wan Y. New insights into KLFs and SOXs in cancer pathogenesis, stemness, and therapy. Semin Cancer Biol. 2023;90:29–44.
Lerner EC, Woroniecka KI, D’Anniballe VM, Wilkinson DS, Mohan AA, Lorrey SJ, Waibl-Polania J, Wachsmuth LP, Miggelbrink AM, Jackson JD, et al. CD8+ T cells maintain killing of MHC-I-negative tumor cells through the NKG2D-NKG2DL axis. Nat Cancer. 2023;4(9):1258–72.
Sakaguchi S, Miyara M, Costantino CM, Hafler DA. FOXP3+ regulatory T cells in the human immune system. Nat Rev Immunol. 2010;10(7):490–500.
Liu W, Yu X, Yuan Y, Feng Y, Wu C, Huang C, Xie P, Li S, Li X, Wang Z, et al. CD73, a promising therapeutic target of diclofenac, promotes metastasis of pancreatic cancer through a nucleotidase independent mechanism. Adv Sci (Weinh). 2023;10(6):e2206335.
Li J, Mo R, Zheng L. Inhibition of the cell migration, invasion and chemoresistance of colorectal cancer cells through targeting KLF3 by miR-365a-3p. J Cancer. 2021;12(20):6155–64.
Wu Y, Chen S, Shao Y, Su Y, Li Q, Wu J, Zhu J, Wen H, Huang Y, Zheng Z, et al. KLF5 promotes tumor progression and parp inhibitor resistance in ovarian cancer. Adv Sci (Weinh). 2023;10(31):e2304638.
Narla G, Heath KE, Reeves HL, Li D, Giono LE, Kimmelman AC, Glucksman MJ, Narla J, Eng FJ, Chan AM, et al. KLF6, a candidate tumor suppressor gene mutated in prostate cancer. Science. 2001;294(5551):2563–6.
Reeves HL, Narla G, Ogunbiyi O, Haq AI, Katz A, Benzeno S, Hod E, Harpaz N, Goldberg S, Tal-Kremer S, et al. Kruppel-like factor 6 (KLF6) is a tumor-suppressor gene frequently inactivated in colorectal cancer. Gastroenterology. 2004;126(4):1090–103.
Zhang Q, Shi R, Bai Y, Meng L, Hu J, Zhu H, Liu T, De X, Wang S, Wang J, et al. Meiotic nuclear divisions 1 (MND1) fuels cell cycle progression by activating a KLF6/E2F1 positive feedback loop in lung adenocarcinoma. Cancer Commun (Lond). 2021;41(6):492–510.
Heinhuis KM, Ros W, Kok M, Steeghs N, Beijnen JH, Schellens JHM. Enhancing antitumor response by combining immune checkpoint inhibitors with chemotherapy in solid tumors. Ann Oncol. 2019;30(2):219–35.
Acknowledgements
Thanks for the clinical samples provided by Standardized Clinical Biobank, National Facility for Translational Medicine (Shanghai), Ruijin Hospital, Shanghai Jiao Tong University School of Medicine.
Funding
Not available.
Author information
Authors and Affiliations
Contributions
JYL and PYL designed the study and performed bioinformatics analysis; JYL and BYS designed experiments and drafted the manuscript; KYS, YL, JL, and YSH were responsible for sample collection and conducted experiments; MMS, TW, JZ, and LXJ were responsible for data analysis and figures generation; BYS and JZ discussed and revised the manuscript. All authors approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Sample collections were approved by the Ethics Committee of Ruijin Hospital, Shanghai Jiaotong University School of Medicine (2021 Clinical Ethnics Review No.161).
Consent for publication
Not applicable.
Competing interests
There is no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
12935_2024_3369_MOESM1_ESM.tif
Supplementary material 1: Figure S1. NMF clustering applied on differently expressed KLFs. (A) Nonnegative matrix factorization rank survey. (B) Consensus matrix heatmaps for k=2-10.
12935_2024_3369_MOESM2_ESM.tif
Supplementary material 2: Figure S2. Enrichment analysis of high- and low-risk group. (A) GSVA analysis indicated the different biological pathways of high- and low-risk groups. The yellow and blue respectively represented activated and suppressive pathways. (B-G) GSEA displayed enriched gene sets in patients with high- and low-risk score groups.
12935_2024_3369_MOESM3_ESM.tif
Supplementary material 3: Figure S3. Drug sensitivity analysis. (A-L) Difference IC50 of axitinib, camptothecin, gefitinib, JNK.Inhibitor.VIII, nilotinib, pazopanib, salubrinal, sunitinib, bleomycin, lapatinib, paclitaxel, and vinorelbine in high- and low-risk groups. (* p < 0.05; ** p < 0.01; *** p < 0.001).
12935_2024_3369_MOESM4_ESM.tif
Supplementary material 4: Figure S4. KLF6 inhibited the proliferation of patient-derived organoids from PDAC. (A-B) Representative pictures of PDO 1# (A) and PDO 2# (B) on day 0, day 5, and day 10 after transfected shNC, sh1 KLF6 and sh2 KLF6. (C-D) The area size was compared on day 0, day 5, and day 10 respectively in shNC, sh1 KLF6, and sh2 KLF6 groups. (* p < 0.05; ** p < 0.01; *** p < 0.001).
12935_2024_3369_MOESM5_ESM.tif
Supplementary material 5: Figure S5. Relevance of KLF6 and tumor immune microenvironment in pan-cancer. (A-Q) The correlation of stromal score and KLF6 expression, estimate score and KLF6 expression, immune score, and KLF6 expression in pan-cancer. (R) Tumor mutation burden of KLF6 in pan-cancer. (S) Microsatellite instability of KLF6 in pan-cancer. (U) The co-expression analysis of immunostimulators and KLF6 in pan-cancer. (V) The co-expression analysis of immunoinhibitors and KLF6 in pan-cancer. (* p < 0.05; ** p < 0.01; *** p < 0.001).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Lin, J., Liu, P., Sun, K. et al. Comprehensive analysis of KLF family reveals KLF6 as a promising prognostic and immune biomarker in pancreatic ductal adenocarcinoma. Cancer Cell Int 24, 177 (2024). https://doi.org/10.1186/s12935-024-03369-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-024-03369-3