
Abstract
Background
Low-density lipoprotein (LDL)-cholesterol is positively associated with cardiovascular disease (CVD) and inversely associated with type 2 diabetes, which could detract from lipid modification. Here, we examined whether lipid traits potentially relevant to CVD aetiology, i.e. apolipoprotein B (apoB), triglycerides (TG) and lipoprotein(a) [Lp(a)] exhibited the same associations. We investigated sex-specifically, including the role of sex hormones, because sex disparities exist in lipid profile and type 2 diabetes. We also replicated where possible.
Methods
We used Mendelian randomization (MR) to examine sex-specific associations of apoB, TG and Lp(a) with type 2 diabetes, HbA1c, fasting insulin, fasting glucose, testosterone and estradiol in the largest relevant sex-specific genome-wide association studies (GWAS) in people of European ancestry and replicated where possible. We also assessed sex-specific associations of liability to type 2 diabetes with apoB, TG and Lp(a).
Results
Genetically predicted apoB and Lp(a) had little association with type 2 diabetes or glycemic traits in women or men. Genetically predicted higher TG was associated with higher type 2 diabetes risk [odds ratio (OR) 1.44 per standard deviation (SD), 95% confidence interval (CI) 1.26 to 1.65], HbA1c and fasting insulin specifically in women. Higher TG was associated with lower testosterone in women and higher testosterone in men, but with lower estradiol in men and women. Genetic liability to type 2 diabetes was associated with higher TG in women, and possibly with lower apoB in men.
Conclusions
Lipid traits potentially relevant to CVD aetiology do not exhibit contrasting associations with CVD and type 2 diabetes. However, higher TG is associated with higher type 2 diabetes risk and glycemic traits, which in turn further increases TG specifically in women, possibly driven by sex hormones.
Similar content being viewed by others


Introduction
Low-density lipoprotein (LDL)-cholesterol is associated with higher risk of cardiovascular disease (CVD) and lower risk of type 2 diabetes [1, 2]. Similar associations have been observed for a major lipid modifier, statins [3], and for familial hypercholesteremia [4], indicating either lipid traits are overlooked causes of type 2 diabetes or additional underlying factors with opposing effects on CVD and type 2 diabetes exist, as previously suggested [5]. Understanding whether other lipids potentially relevant to the aetiology of CVD have opposing effects on CVD and type 2 diabetes has public health implications for clinical interventions and drug development for CVD prevention.
Apolipoprotein B (apoB) is emerging as the predominant trait in the aetiology of CVD, probably accounting for the effect of LDL-cholesterol on CVD and lifespan [6, 7]. The causal roles of triglycerides (TG) [8] and lipoprotein(a) [Lp(a)] [9] in CVD are gaining acceptance, whilst the roles of high-density lipoprotein (HDL)-cholesterol and correspondingly apolipoprotein A (apoA) are in doubt given drugs targeting HDL-cholesterol do not reduce CVD risk in trials [10, 11]. However, the association of apoB with type 2 diabetes risk remains unclear [7]. Previous studies investigating the associations of TG and Lp(a) with type 2 diabetes risk have yielded contradictory results, suggesting inverse [1, 12], null [13,14,15] or positive [2, 7, 16] associations of TG, and inverse [17] or null [18] associations of Lp(a).
Women usually have a less atherogenic lipid profiles than men [19] and lipids have differing associations with CVD by sex [20], while diabetes incidence is similar in men and women although differences by sex exist in pathophysiology and complications of type 2 diabetes, possibly partly driven by sex hormones [21, 22]. Estrogen protects against type 2 diabetes in women [23], while testosterone protects against type 2 diabetes in men [24]. Few studies have evaluated sex-specific associations of lipid traits with type 2 diabetes risk [16]. Randomized controlled trials (RCTs) are not usually powered to detect sex differences. Given the correlation between lipid fractions, it is also difficult to disentangle the role of each lipid fraction in the development of type 2 diabetes in an RCT.
To examine sex-specific associations of lipid traits potentially relevant to the aetiology of CVD, i.e. apoB, TG and Lp(a), with type 2 diabetes risk, glycemic traits and sex hormones, we conducted a Mendelian randomization (MR) study, i.e., an instrumental variable analysis with genetic instruments [25]. MR studies take advantage of genetic randomization at conception to obtain less confounded estimates [25]. We used multivariable MR to assess the robustness of the findings [26], and replicated, where possible, i.e. in East Asians.
Methods
Study design
We performed a two-sample MR study to examine sex-specific associations of apoB, TG and Lp(a) with type 2 diabetes, glycemic traits and sex hormones, taking advantage of the largest relevant publicly available sex-specific genetic summary statistics. We extracted sex-specific independent (r2 < 0.001) genome-wide significant (p value < 5 × 10–8) genetic instruments for each lipid trait from the UK Biobank (http://www.nealelab.is/uk-biobank/), and where possible, ancestry-specific genetic instruments from the Global Lipids Genetics Consortium (GLGC) excluding the UK Biobank participants [27] for replication. We applied them to the largest sex-specific genome-wide association study (GWAS) of type 2 diabetes, with different participants from the UK Biobank, in people of European ancestry [28], and then replicated in East Asians [29]. To give greatly granularity, we also applied these genetic instruments to the largest relevant sex-specific GWAS of HbA1c, fasting insulin, fasting glucose, testosterone and estradiol in people of European ancestry [30,31,32]. We used multivariable MR to assess the role of each lipid trait taking into account the others and additionally body mass index (BMI) [26], because BMI affects both lipid traits and type 2 diabetes [33], and might confound their associations given genetic instruments for lipid traits may also predict BMI. We also assessed sex-specific associations of genetic liability to type 2 diabetes with lipid traits, because bidirectional relationships between lipids and glycemic traits have previously been suggested [34].
Genetic predictors for lipid traits
We extracted sex-specific independent (r2 < 0.001) genome-wide significant (p value < 5 × 10–8) genetic instruments for apoB (184,377 women/158,213 men), TG (184,885 women/159,107 men) and Lp(a) (147,684 women/126,212 men) from sex-specific GWAS of the UK Biobank (http://www.nealelab.is/uk-biobank). The quality controlled GWAS included people of white British ancestry, intended age 40–69 years, adjusted for age, age2, and the first 20 principal components. We also extracted ancestry-specific independent (r2 < 0.001) genome-wide significant (p value < 5 × 10–8) genetic instruments for TG (864,240/83,965 people of European/East Asian ancestry) from GLGC excluding the UK Biobank participants [27] for replication. Summary statistics were adjusted for age, age2, sex, principal components of ancestry and study-specific covariates [27].We excluded genetic variants located on the GCKR or FADS1 genes as previously [13, 14, 16], because they are strongly associated with other traits relevant to type 2 diabetes.
To assess the validity of sex-specific genetic instruments from the UK Biobank in East Asians, we used coronary artery disease (CAD) (cases = 7708 women/21,611 men, controls = 95,398 women/87,736 men) from Biobank Japan [35] as a positive control outcome. Summary statistics were adjusted for age and top five principal components [35].
We conducted multivariable MR using sex-specific genetic instruments from the UK Biobank to identify the direct effect of each lipid, i.e. apoB, TG and Lp(a), accounting for potential confounding or mediation by the other lipids considered [26]. As the effects of lipid traits on type 2 diabetes might also be confounded by BMI [33], we additionally included BMI in the multivariable MR analyses. We obtained sex-specific genetic associations with BMI (193,570 women/166,413 men) from a GWAS of the UK Biobank (http://www.nealelab.is/uk-biobank), adjusted for age, age2, and the first 20 principal components. In multivariable MR, we combined all the genetic instruments, dropped duplicated SNPs and removed correlated (r2 ≥ 0.001) SNPs based on the minimum p value for genetic association with each trait. We extracted associations of the remaining SNPs with the exposures and the outcome, then fitted multivariable models.
Genetic associations with type 2 diabetes
We obtained sex-specific genetic associations with type 2 diabetes from the DIAbetes Meta-ANalysis of Trans-Ethnic association studies (DIAMANTE) consortium in people of European ancestry excluding UK Biobank participants (cases = 23,197 women/29,583 men, controls = 201,329 women/193,076 men) [28]. Summary genetic associations were adjusted for study-specific covariates and principal components [28]. We extracted sex-specific independent (r2 < 0.001) genome-wide significant (p value < 5 × 10–8) genetic instruments for liability to type 2 diabetes from the GWAS [28].
We also obtained sex-specific associations with type 2 diabetes (cases = 27,370 women/28,027 men, controls = 135,055 women/89,312 men) from the Asian Genetic Epidemiology Network (AGEN) meta-analysis in East Asians, adjusted for age, sex, study-specific covariates and principal components of ancestry [29].
Genetic associations with HbA1c, fasting insulin and fasting glucose
We obtained sex-specific genetic associations with HbA1c from a GWAS of the UK Biobank (185,022 women/159,160 men), adjusted for age, age2 and 20 principal components (http://www.nealelab.is/uk-biobank/). We obtained sex-specific genetic associations with fasting insulin (50,404 women/47,806 men) and fasting glucose (73,089 women/67,506 men) from the Meta-Analyses of Glucose and Insulin-related traits Consortium (MAGIC) in people of European ancestry without diabetes, which were adjusted for age, study site and principal components [30].
Genetic associations with testosterone and estradiol
We obtained genetic associations with total testosterone in women (N = 230,454) and bioavailable testosterone, hereafter testosterone in men (N = 178,782), from a GWAS conducted in the UK Biobank, because they have little correlation with sex hormone-binding globulin (SHBG) [31]. Estimates were adjusted for genotyping chip/release of genetic data, age at baseline, fasting time, center and 10 genetically derived principal components [31].
We obtained sex-specific genetic associations with estradiol (above detection limit = 37,461 women/13,367 men, below detection limit = 126,524 women/134,323 men) from a GWAS of the UK Biobank [32]. Summary quality controlled genetic associations were adjusted for age, BMI, the first 10 genetic principal components, genotyping array, and additionally hormone replacement therapy, oral contraceptive use, number of live births, menopausal status, and hysterectomy in women [32].
Statistical analysis
We used the F-statistic to assess instrument strength, obtained from the mean of the square of each SNP-exposure association divided by the square of its standard error [36]. An F-statistic larger than 10 suggests weak instrument bias is unlikely. In multivariable MR, we used the conditional F-statistic FTS to examine the instrument strength for each exposure conditional on the other exposures, and the Q-statistic to assess heterogeneity [37].
We aligned the SNPs based on alleles and/or allele frequency and excluded palindromic SNPs with intermediate effect allele frequency (i.e. 0.42–0.58) when the strand direction was uncertain. We used proxy SNPs (r2 ≥ 0.8), where possible, when SNPs were not in the outcome GWAS. We obtained MR estimates by meta-analyzing Wald estimates (i.e. genetic association with the outcome divided by genetic association with the exposure) using inverse variance weighting (IVW) with first-order weights, and fixed effects for three SNPs or less and random effects for four SNPs or more [38]. IVW assumes all the genetic variants are valid or have balanced pleiotropy [38]. IVW using first-order weights gives unbiased estimates with or without the presence of heterogeneity, when the mean F-statistic is high [39]. To assess the validity of the IVW estimates, we conducted sensitivity analyses using methods with different assumptions, i.e. the weighted median [40], MR Egger [41] and the contamination mixture method [42]. The weighted median is valid when more than half of the information derives from valid SNPs [40]. MR Egger assumes no consequence of the instruments confounds exposure on outcome [41]. We used the MR Egger intercept to assess whether the IVW estimate might be affected by violation of the exclusion-restriction assumption [41]. The contamination mixture method is robust to outliers and horizontal pleiotropy, with well-controlled type 1 error rates [42, 43]. We used multivariable IVW to assess the direct effect of each lipid trait controlling for the others. We used multivariable MR Egger and additional adjustment for BMI as sensitivity analysis.
We used Steiger filtering to identify SNPs explaining more of the variance in the outcome than in the exposure (p value < 0.05) [44], given potentially bidirectional relationships between lipids and glycemic traits [34]. To assess whether MR estimates were affected by reverse causality, we conducted analyses including and excluding these SNPs.
Power was estimated based on the approximation that the sample size required for an MR study is the sample size for the conventional observational study divided by the variance in the exposure explained by the SNPs [45]. This variance was estimated as the sum of 2*beta2*MAF*(1-MAF), where beta is the standardized genetic association with the exposure and MAF is the minor allele frequency.
Sex differences were assessed using a two-sided z-test. A Bonferroni corrected significance level was set at α = 0.05/3 = 0.017, because 3 lipid fractions were included. All statistical analyses were conducted using R version 4.1.1 and the packages “TwoSampleMR” for harmonizing data, “MendelianRandomization” for univariable and multivariable MR, “MVMR” for conditional F-statistics and Q-statistics, and “ieugwasr” for removing correlated SNPs. All analyses were based on publicly available summary statistics, which does not require ethical approval.
Results
Genetic instruments
We extracted 111 (women) and 80 (men) independent (r2 < 0.001) genome-wide significant (p value < 5 × 10–8) SNPs for apoB, 144 (women) and 96 (men) for TG, and 15 (women) and 10 (men) for Lp(a) from the UK Biobank. All the SNPs had an F-statistic larger than 10. After excluding 2 genetic variants (rs780094 and rs1260326) located on GCKR, the mean F-statistics were 170.9 (women) and 188.9 (men) for apoB, 117.7 (women) and 139.9 (men) for TG, and 673.3 (women) and 1101.5 (men) for Lp(a). When instrumented by these SNPs, genetically predicted apoB, TG and Lp(a) were positively associated with the positive control outcome, i.e. CAD in East Asians, despite wide confidence intervals for Lp(a) (Additional file 1: Table S1).
We also extracted 238 (European) and 29 (East Asian) independent (r2 < 0.001) genome-wide significant (p value < 5 × 10–8) SNPs for TG from GLGC excluding the UK Biobank participants for replication. We excluded 2 SNPs (rs150419156 and rs1260326) on GCKR and 1 SNP (rs7394579) on FADS1. The F-statistics were all greater than 10, with mean 236.3 and 192.6 for people of European and East Asian ancestry, respectively.
In multivariable MR including apoB, TG and Lp(a), the conditional F-statistics were 68.1 (women) and 96.8 (men) for apoB, 69.6 (women) and 80.2 (men) for TG, and 46.8 (women) and 72.5 (men) for Lp(a). We extracted 148 (women) and 134 (men) independent (r2 < 0.001) genome-wide significant (p value < 5 × 10–8) SNPs for BMI, and additionally included BMI in the multivariable MR, when the conditional F-statistics were 43.4 (women) and 53.3 (men) for apoB, 49.3 (women) and 50.2 (men) for TG, 34.4 (women) and 45.7 (men) for Lp(a), and 21.7 (women) and 24.2 (men) for BMI.
We also extracted 33 (women) and 48 (men) independent (r2 < 0.001) genome-wide significant (p value < 5 × 10–8) genetic instruments for liability to type 2 diabetes from DIAMANTE in people of European ancestry excluding UK Biobank participants. The F-statistics were all greater than 10, with mean 63.5 and 68.3 for women and men, respectively.
Results of power calculations are shown in Additional file 1: Table S2.
Sex-specific associations of lipid traits with type 2 diabetes and glycemic traits
In the univariable MR analyses, genetically predicted apoB was not associated with type 2 diabetes risk or with any glycemic trait in women or men (Figs. 1, 2). Genetically predicted higher TG was associated with higher type 2 diabetes risk, HbA1c and fasting insulin in women, but not men (Figs. 1, 2). Findings were similar when using ancestry-specific TG SNPs from GLGC excluding the UK Biobank participants, although the MR Egger intercept indicated possible pleiotropy (Additional file 1: Table S3). Genetically predicted Lp(a) was not associated with type 2 diabetes risk in women or men (Fig. 1), despite a positive association with HbA1c (Fig. 2).

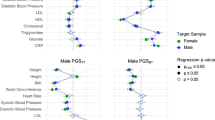
Ancestry- and sex-specific associations of genetically predicted lipid fractions (instrumented by the SNPs from the UK Biobank) with type 2 diabetes. a. MVMR multivariable MR, UVMR univariable MR, b. MVMR estimates for each lipid fraction were adjusted for the other two traits; c. Estimates were derived using inverse variance weighted approach, and were expressed in standard deviation for lipid fractions, and in odds ratio for type 2 diabetes

Sex-specific associations of genetically predicted lipid fractions (instrumented by the SNPs from the UK Biobank) with glycemic traits in people of European ancestry. a. MVMR multivariable MR, UVMR univariable MR, b. MVMR estimates for each lipid fraction were adjusted for the other two traits; c. Estimates were derived using inverse variance weighted approach, and were expressed in standard deviation for lipid fractions and HbA1c, in pmol/L (natural log transformed) for fasting insulin, and in mmol/L for fasting glucose
In the multivariable MR analyses, the associations of apoB with type 2 diabetes and glycemic traits were similar when taking into account TG and Lp(a) (Figs. 1, 2). The positive association of TG with type 2 diabetes became pronounced in men of European ancestry after controlling for apoB and Lp(a) (Fig. 1); however, the p value did not reach Bonferroni corrected significance. The positive association of Lp(a) with HbA1c was attenuated towards the null after controlling for apoB and TG (Fig. 2). These associations were robust to different analytic methods, additional adjustment for BMI and exclusion of SNPs explaining more of the variance in the outcome than in the exposure (Additional file 1: Table S4, S5, S6, S7). The associations of lipid traits with type 2 diabetes were replicated in East Asians (Fig. 1).
Sex-specific associations of lipid traits with testosterone and estradiol
In the univariable MR analyses, genetically predicted apoB had little association with sex hormones in women or men (Fig. 3). Genetically predicted higher TG was associated with lower testosterone in women and higher testosterone in men, but with lower estradiol in men and women (Fig. 3). Findings were similar when instrumented by TG SNPs from GLGC excluding the UK Biobank (Additional file 1: Table S3). Genetically predicted higher Lp(a) was possibly associated with higher estradiol in women, but not in men (Fig. 3). Multivariable MR analyses taking into account the effects of other lipid traits gave consistent results (Fig. 3).

Sex-specific associations of genetically predicted lipid fractions (instrumented by the SNPs from the UK Biobank) with sex hormones in people of European ancestry. a. MVMR multivariable MR, UVMR univariable MR, b. MVMR estimates for each lipid fraction were adjusted for the other traits; c. Estimates were derived using inverse variance weighted approach, and were expressed in standard deviation for lipid fractions and testosterone, and in log odds for estradiol (above and below the limit of detection)
The MR Egger intercept indicated possible pleiotropy, and the Q-statistics suggested heterogeneity in multivariable MR (Additional file 1: Table S8). Nevertheless, findings were similar when using different analytic methods, additionally adjusting for BMI and excluding SNPs explaining more of the variance in the outcome than in the exposure (Additional file 1: Table S8, S9).
Sex-specific associations of liability to type 2 diabetes with lipid traits
Genetic liability to type 2 diabetes was associated with higher TG in women, and possibly with lower apoB in men (Table 1, p values for sex difference 0.002 and 0.067, respectively). Findings were similar when using different analytic methods, and the MR Egger intercept did not indicate possible pleiotropy (Table 1). The inverse association of liability to type 2 diabetes with apoB became pronounced in men after excluding SNPs explaining more of the variance in the outcome than in the exposure (Table 1, p values for sex difference 0.001).
Discussion
Consistent with previous studies [2, 7, 16, 18], we found a positive association of TG with type 2 diabetes but little association of Lp(a) with type 2 diabetes. We added by showing apoB was not associated with type 2 diabetes or glycemic traits, but TG was positively associated with type 2 diabetes and glycemic traits specifically in women. The finding that lipid traits potentially relevant to the aetiology of CVD do not exhibit contrasting associations with risk of CVD and type 2 diabetes is novel and has implications for interventions and drug development for CVD prevention.
We found apoB, the predominant trait in the aetiology of CVD [6, 46], was not associated with type 2 diabetes or any glycemic trait, in contrast to previous MR studies showing an inverse association of LDL-cholesterol with type 2 diabetes [1, 2]. Despite the high correlation between apoB and LDL-cholesterol, the mass of cholesterol per apoB particle is not uniform [47]. For a given value of apoB, the level of LDL-cholesterol increases when apoB particles are cholesterol-enriched [47]. In this situation, the uptake of cholesterol via the LDL receptor reduces, which might improve beta cell function and protect against type 2 diabetes [48]. Although diabetogenic effects of lipid modifiers have been observed, these effects are drug-specific rather than a general property of lipid lowering. Meta-analysis of RCTs suggest that only statins, of lipid modifiers currently in use, increase the risk of incident diabetes [49], possibly due to underlying factors with opposite effects on CVD and type 2 diabetes, such as sex hormones [5].
Our finding that higher TG was associated with higher type 2 diabetes risk and glycemic traits specifically in women is inconsistent with some previous MR studies showing inverse [1, 12] or null [13,14,15] associations of TG with type 2 diabetes. However, these studies used fewer SNPs [13,14,15], had smaller sample sizes [12,13,14,15], did not use analytic methods robust to pleiotropy [1, 12], and did not assess sex-specific associations [1, 12,13,14,15]. Our finding is consistent with an RCT showing bezafibrate lowering TG decreases type 2 diabetes incidence [50]. Differences by sex are consistent with observational studies showing a stronger relation of TG with diabetes in women than men [51, 52], but inconsistent with the Tehran Lipid and Glucose Study suggesting women experience more adverse changes in BMI and TG than men before the onset of diabetes [53]. However, this observation is possibly due to higher insulin sensitivity in women [21], and thus they may experience a greater burden of metabolic risk factors than men before diabetes becomes evident. Notably, the causal relation of BMI with type 2 diabetes is also stronger in women than men [54].
Genetically predicted TG had an inverse association with estradiol, and had sex-specific associations with testosterone, which may underlie sex-specific associations of TG with type 2 diabetes and glycemic traits. Estrogen protects against type 2 diabetes in women in trials [23], mainly through activating estrogen receptor α in various tissues (e.g. brain, liver, skeletal muscle, adipose tissue and pancreatic beta cells) and thereby improving adiposity, insulin sensitivity and glucose tolerance [21]. TG lowering estradiol in women could increase the risk of type 2 diabetes specifically in women. Consistently, higher TG is also associated with lower breast cancer risk [55] and lower bone mineral density [56]. Testosterone protects against type 2 diabetes in men [24], possibly due to improvement in body composition and insulin sensitivity [57]. Thus, TG increasing testosterone in men may ameliorate any effects of TG on type 2 diabetes. TG may also work via altering insulin-like growth factor-1 (IGF-1) and increase type 2 diabetes risk [58, 59]. However, the causal association of IGF-1 with type 2 diabetes is complex and differs by underlying molecular pathway [60]. Furthermore, sex disparities in the treatment of dyslipidemia could contribute to the positive association of TG with type 2 diabetes in women [61], but would not explain why the difference is specific to TG.
Consistent with a previous MR study suggesting bidirectional relationships between lipids and glycemic traits [34], we add by showing these relationships may differ by sex. Specifically, higher TG was associated with higher type 2 diabetes risk and glycemic traits, which in turn would increase TG in women. It has clinical implications that women might benefit more from early control of TG than men. Observationally, the relative risk of CVD associated with TG is higher in women than men [62], though not consistently so [63]. Further studies are warranted to assess whether the associations of TG with CVD and lifespan differ by sex, and to investigate the role of sex hormones in mediating these associations.
This is the first MR study comprehensively assessing sex-specific associations of lipid traits relevant to the aetiology of CVD with type 2 diabetes and glycemic traits, in both people of European and East Asian ancestry. Nevertheless, this study has several limitations. First, MR should fulfill three rigorous assumptions of relevance, independence and exclusion-restriction [25]. To fulfill the relevance assumption, we checked that F-statistics and conditional F-statistics were > 10, which suggests little weak instrument bias. To satisfy the independence assumption, we excluded the genetic variants located on the GCKR or FADS1 genes as previously [13, 14, 16], because these genes are strongly associated with other traits relevant to type 2 diabetes. To address the exclusion restriction assumption, we used analytic methods with different assumptions and multivariable MR to control for potential pleiotropy via other lipid traits and BMI, which gave consistent conclusions. Second, we extracted genetic instruments for lipid traits from the same study (i.e. the UK Biobank) as we obtained the genetic effects on sex hormones. However, two-sample MR methods in a one-sample setting perform well within large biobanks, except for the MR Egger estimate, when the variability of instrument strength across variants (I2GX) is < 97% [64], which was not the case here. The bias due to overlapping sample is proportional to 1/F-statistic [65] and unlikely changes the results substantially given the high F-statistics (average F-statistics larger than 100 for all lipid traits in women and men). In addition, the associations of TG with sex hormones were replicated using the genetic instruments extracted from GLGC excluding the UK Biobank participants. Third, we extracted genetic instruments for lipid traits from a GWAS performed in people of European ancestry and used them to derive MR estimates in East Asians. However, using these SNPs gave expected results for CAD in East Asians. In addition, findings were similar when using TG SNPs obtained from East Asians. Fourth, MR, particularly for dichotomous outcomes, could be open to selection bias [66]. The associations of lipid traits with type 2 diabetes could be biased when the underlying sample is selected on surviving to recruitment on genetic make-up and competing risks of type 2 diabetes [66]. However, the participants were relatively young likely obviating selective survival to recruitment on genetic endowment. Furthermore, we obtained similar findings when using continuous outcomes, i.e. glycemic traits which are less likely affected by selection bias [66]. We obtained genetic associations of fasting insulin and fasting glucose from MAGIC only including individuals without diabetes [30], which may underestimate the associations. Fifth, the associations in people of European ancestry may not be transportable to other populations. However, causal effects should act consistently across settings, unless the mediating mechanisms differ [67]. We replicated the associations in East Asians; further investigation in other populations would be worthwhile. Sixth, we assessed whether MR estimates were affected by reverse causality using Steiger filtering, which calculated the variance explained in the exposure and the outcome by the genetic instruments [44]. However, Steiger filtering might have reduced statistical power or infer the wrong direction when measurement error differs between the exposure and the outcome [44]. Seventh, estradiol was taken as a binary phenotype (above and below the limit of detection) [32], and thus it is difficult to interpretate the magnitude of the estimates and compare the associations of lipid traits with estradiol between women and men. Finally, MR assesses lifetime effects of lipids, which could differ from the short-term effects of lipid-lowering drugs.
Conclusions
Lipid traits potentially relevant to the aetiology of CVD do not exhibit contrasting associations with risk of CVD and type 2 diabetes. However, higher TG was positively associated with type 2 diabetes risk and glycemic traits, which in turn would increase TG specifically in women, possibly driven by sex hormones. These insights have implications for public health interventions and drug development for CVD prevention including identifying potential side-effects and reinforcing the importance of using sex-specific approaches in the investigation, prevention, treatment and management of CVD and type 2 diabetes.
Availability of data and materials
Summary statistics are available in the website http://www.nealelab.is/uk-biobank/ for UK Biobank (Neale lab), http://jenger.riken.jp/en/result for Biobank Japan, http://csg.sph.umich.edu/willer/public/glgc-lipids2021/ for GLGC, http://diagram-consortium.org/downloads.html for DIAMANTE, https://magicinvestigators.org/downloads/ for MAGIC, https://blog.nus.edu.sg/agen/summary-statistics/t2d-2020/ for AGEN, and https://www.ebi.ac.uk/gwas/ for GWAS of testosterone and estradiol. The data analyzed in the current study are provided in Additional file 2: Table S10, S11, S12, S13, S14, S15, S16, S17.
Abbreviations
- AGEN:
-
Asian genetic epidemiology network
- ApoA:
-
Apolipoprotein A
- ApoB:
-
Apolipoprotein B
- BMI:
-
Body mass index
- CI:
-
Confidence interval
- CVD:
-
Cardiovascular diseases
- DIAMANTE:
-
DIAbetes Meta-ANalysis of Trans-Ethnic association studies
- GLGC:
-
Global lipids genetics consortium
- GWAS:
-
Genome-wide association studies
- HDL:
-
High-density lipoprotein
- IGF-1:
-
Insulin-like growth factor-1
- IVW:
-
Inverse variance weighting
- LDL:
-
Low-density lipoprotein
- Lp(a):
-
Lipoprotein(a)
- MAGIC:
-
Meta-analyses of glucose and insulin-related traits consortium
- MR:
-
Mendelian randomization
- OR:
-
Odds ratio
- RCT:
-
Randomized controlled trial
- SD:
-
Standard deviation
- SHBG:
-
Sex hormone-binding globulin
- TG:
-
Triglycerides
References
White J, Swerdlow DI, Preiss D, Fairhurst-Hunter Z, Keating BJ, Asselbergs FW, Sattar N, Humphries SE, Hingorani AD, Holmes MV. Association of lipid fractions with risks for coronary artery disease and diabetes. JAMA Cardiol. 2016;1(6):692–9.
Fall T, Xie W, Poon W, Yaghootkar H, Mägi R, Knowles JW, Lyssenko V, Weedon M, Frayling TM, Ingelsson E. Using genetic variants to assess the relationship between circulating lipids and type 2 diabetes. Diabetes. 2015;64(7):2676–84.
Swerdlow DI, Preiss D, Kuchenbaecker KB, Holmes MV, Engmann JE, Shah T, Sofat R, Stender S, Johnson PC, Scott RA, et al. HMG-coenzyme a reductase inhibition, type 2 diabetes, and bodyweight: evidence from genetic analysis and randomised trials. Lancet. 2015;385(9965):351–61.
Besseling J, Kastelein JJ, Defesche JC, Hutten BA, Hovingh GK. Association between familial hypercholesterolemia and prevalence of type 2 diabetes mellitus. JAMA. 2015;313(10):1029–36.
Schooling CM, Au Yeung SL, Leung GM. Why do statins reduce cardiovascular disease more than other lipid modulating therapies? Eur J Clin Invest. 2014;44(11):1135–40.
Richardson TG, Sanderson E, Palmer TM, Ala-Korpela M, Ference BA, Davey Smith G, Holmes MV. Evaluating the relationship between circulating lipoprotein lipids and apolipoproteins with risk of coronary heart disease: a multivariable Mendelian randomisation analysis. PLoS Med. 2020;17(3):e1003062.
Richardson TG, Wang Q, Sanderson E, Mahajan A, McCarthy MI, Frayling TM, Ala-Korpela M, Sniderman A, Smith GD, Holmes MV. Effects of apolipoprotein B on lifespan and risks of major diseases including type 2 diabetes: a Mendelian randomisation analysis using outcomes in first-degree relatives. Lancet Healthy longev. 2021;2(6):e317–26.
Holmes MV, Asselbergs FW, Palmer TM, Drenos F, Lanktree MB, Nelson CP, Dale CE, Padmanabhan S, Finan C, Swerdlow DI, et al. Mendelian randomization of blood lipids for coronary heart disease. Eur Heart J. 2015;36(9):539–50.
Saleheen D, Haycock PC, Zhao W, Rasheed A, Taleb A, Imran A, Abbas S, Majeed F, Akhtar S, Qamar N, et al. Apolipoprotein(a) isoform size, lipoprotein(a) concentration, and coronary artery disease: a Mendelian randomisation analysis. Lancet Diabetes Endocrinol. 2017;5(7):524–33.
Schwartz GG, Olsson AG, Abt M, Ballantyne CM, Barter PJ, Brumm J, Chaitman BR, Holme IM, Kallend D, Leiter LA, et al. Effects of dalcetrapib in patients with a recent acute coronary syndrome. N Engl J Med. 2012;367(22):2089–99.
Lincoff AM, Nicholls SJ, Riesmeyer JS, Barter PJ, Brewer HB, Fox KAA, Gibson CM, Granger C, Menon V, Montalescot G, et al. Evacetrapib and cardiovascular outcomes in high-risk vascular disease. N Engl J Med. 2017;376(20):1933–42.
Klimentidis YC, Chougule A, Arora A, Frazier-Wood AC, Hsu CH. Triglyceride-increasing alleles associated with protection against type-2 diabetes. PLoS Genet. 2015;11(5):e1005204.
De Silva NM, Freathy RM, Palmer TM, Donnelly LA, Luan J, Gaunt T, Langenberg C, Weedon MN, Shields B, Knight BA, et al. Mendelian randomization studies do not support a role for raised circulating triglyceride levels influencing type 2 diabetes, glucose levels, or insulin resistance. Diabetes. 2011;60(3):1008–18.
Islam M, Jafar TH, Wood AR, De Silva NM, Caulfield M, Chaturvedi N, Frayling TM. Multiple genetic variants explain measurable variance in type 2 diabetes-related traits in Pakistanis. Diabetologia. 2012;55(8):2193–204.
Marott SC, Nordestgaard BG, Tybjærg-Hansen A, Benn M. Components of the metabolic syndrome and risk of type 2 diabetes. J Clin Endocrinol Metab. 2016;101(8):3212–21.
Qi Q, Liang L, Doria A, Hu FB, Qi L. Genetic predisposition to dyslipidemia and type 2 diabetes risk in two prospective cohorts. Diabetes. 2012;61(3):745–52.
Gudbjartsson DF, Thorgeirsson G, Sulem P, Helgadottir A, Gylfason A, Saemundsdottir J, Bjornsson E, Norddahl GL, Jonasdottir A, Jonasdottir A, et al. Lipoprotein(a) concentration and risks of cardiovascular disease and diabetes. J Am Coll Cardiol. 2019;74(24):2982–94.
Kamstrup PR, Nordestgaard BG. Lipoprotein(a) concentrations, isoform size, and risk of type 2 diabetes: a Mendelian randomisation study. Lancet Diabetes Endocrinol. 2013;1(3):220–7.
Freedman DS, Otvos JD, Jeyarajah EJ, Shalaurova I, Cupples LA, Parise H, D’Agostino RB, Wilson PW, Schaefer EJ. Sex and age differences in lipoprotein subclasses measured by nuclear magnetic resonance spectroscopy: the framingham study. Clin Chem. 2004;50(7):1189–200.
Cupido AJ, Asselbergs FW, Schmidt AF, Hovingh GK. Low-density lipoprotein cholesterol attributable cardiovascular disease risk is sex specific. J Am Heart Assoc. 2022;11(12):e024248.
Tramunt B, Smati S, Grandgeorge N, Lenfant F, Arnal JF, Montagner A, Gourdy P. Sex differences in metabolic regulation and diabetes susceptibility. Diabetologia. 2020;63(3):453–61.
Kautzky-Willer A, Harreiter J, Pacini G. Sex and gender differences in risk, pathophysiology and complications of type 2 diabetes mellitus. Endocr Rev. 2016;37(3):278–316.
Margolis KL, Bonds DE, Rodabough RJ, Tinker L, Phillips LS, Allen C, Bassford T, Burke G, Torrens J, Howard BV. Effect of oestrogen plus progestin on the incidence of diabetes in postmenopausal women: results from the women’s health initiative hormone trial. Diabetologia. 2004;47(7):1175–87.
Wittert G, Bracken K, Robledo KP, Grossmann M, Yeap BB, Handelsman DJ, Stuckey B, Conway A, Inder W, McLachlan R, et al. Testosterone treatment to prevent or revert type 2 diabetes in men enrolled in a lifestyle programme (T4DM): a randomised, double-blind, placebo-controlled, 2 year, phase 3b trial. Lancet Diabetes Endocrinol. 2021;9(1):32–45.
Lawlor DA, Harbord RM, Sterne JA, Timpson N, Davey Smith G. Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med. 2008;27(8):1133–63.
Burgess S, Thompson SG. Multivariable Mendelian randomization: the use of pleiotropic genetic variants to estimate causal effects. Am J Epidemiol. 2015;181(4):251–60.
Graham SE, Clarke SL, Wu KH, Kanoni S, Zajac GJM, Ramdas S, Surakka I, Ntalla I, Vedantam S, Winkler TW, et al. The power of genetic diversity in genome-wide association studies of lipids. Nature. 2021;600(7890):675–9.
Mahajan A, Taliun D, Thurner M, Robertson NR, Torres JM, Rayner NW, Payne AJ, Steinthorsdottir V, Scott RA, Grarup N, et al. Fine-mapping type 2 diabetes loci to single-variant resolution using high-density imputation and islet-specific epigenome maps. Nat Genet. 2018;50(11):1505–13.
Spracklen CN, Horikoshi M, Kim YJ, Lin K, Bragg F, Moon S, Suzuki K, Tam CHT, Tabara Y, Kwak SH, et al. Identification of type 2 diabetes loci in 433,540 East Asian individuals. Nature. 2020;582(7811):240–5.
Lagou V, Mägi R, Hottenga JJ, Grallert H, Perry JRB, Bouatia-Naji N, Marullo L, Rybin D, Jansen R, Min JL, et al. Sex-dimorphic genetic effects and novel loci for fasting glucose and insulin variability. Nat Commun. 2021;12(1):24.
Ruth KS, Day FR, Tyrrell J, Thompson DJ, Wood AR, Mahajan A, Beaumont RN, Wittemans L, Martin S, Busch AS, et al. Using human genetics to understand the disease impacts of testosterone in men and women. Nat Med. 2020;26(2):252–8.
Schmitz D, Ek WE, Berggren E, Höglund J, Karlsson T, Johansson Å. Genome-wide association study of estradiol levels and the causal effect of estradiol on bone mineral density. J Clin Endocrinol Metab. 2021;106(11):e4471–86.
Dale CE, Fatemifar G, Palmer TM, White J, Prieto-Merino D, Zabaneh D, Engmann JEL, Shah T, Wong A, Warren HR, et al. Causal associations of adiposity and body fat distribution with coronary heart disease, stroke subtypes, and type 2 diabetes mellitus: a Mendelian randomization analysis. Circulation. 2017;135(24):2373–88.
Zhu Z, Wang K, Hao X, Chen L, Liu Z, Wang C. Causal graph among serum lipids and glycemic traits: a Mendelian randomization study. Diabetes. 2022;71(8):1818–26.
Ishigaki K, Akiyama M, Kanai M, Takahashi A, Kawakami E, Sugishita H, Sakaue S, Matoba N, Low SK, Okada Y, et al. Large-scale genome-wide association study in a Japanese population identifies novel susceptibility loci across different diseases. Nat Genet. 2020;52(7):669–79.
Bowden J, Del Greco MF, Minelli C, Davey Smith G, Sheehan NA, Thompson JR. Assessing the suitability of summary data for two-sample Mendelian randomization analyses using MR-Egger regression: the role of the I2 statistic. Int J Epidemiol. 2016;45(6):1961–74.
Sanderson E, Spiller W, Bowden J. Testing and correcting for weak and pleiotropic instruments in two-sample multivariable Mendelian randomization. Stat Med. 2021;40(25):5434–52.
Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37(7):658–65.
Bowden J, Del Greco MF, Minelli C, Zhao Q, Lawlor DA, Sheehan NA, Thompson J, Davey Smith G. Improving the accuracy of two-sample summary-data Mendelian randomization: moving beyond the NOME assumption. Int J Epidemiol. 2019;48(3):728–42.
Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in Mendelian Randomization with some invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40(4):304–14.
Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015;44(2):512–25.
Burgess S, Foley CN, Allara E, Staley JR, Howson JMM. A robust and efficient method for Mendelian randomization with hundreds of genetic variants. Nat Commun. 2020;11(1):376.
Slob EAW, Burgess S. A comparison of robust Mendelian randomization methods using summary data. Genet Epidemiol. 2020;44(4):313–29.
Hemani G, Tilling K, Davey Smith G. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genet. 2017;13(11):e1007081.
Freeman G, Cowling BJ, Schooling CM. Power and sample size calculations for Mendelian randomization studies using one genetic instrument. Int J Epidemiol. 2013;42(4):1157–63.
Yuan S, Tang B, Zheng J, Larsson SC. Circulating lipoprotein lipids, apolipoproteins and ischemic stroke. Ann Neurol. 2020;88(6):1229–36.
Sniderman AD, Thanassoulis G, Glavinovic T, Navar AM, Pencina M, Catapano A, Ference BA. Apolipoprotein B particles and cardiovascular disease: a narrative review. JAMA Cardiol. 2019;4(12):1287–95.
Kruit JK, Kremer PH, Dai L, Tang R, Ruddle P, de Haan W, Brunham LR, Verchere CB, Hayden MR. Cholesterol efflux via ATP-binding cassette transporter A1 (ABCA1) and cholesterol uptake via the LDL receptor influences cholesterol-induced impairment of beta cell function in mice. Diabetologia. 2010;53(6):1110–9.
Khan SU, Rahman H, Okunrintemi V, Riaz H, Khan MS, Sattur S, Kaluski E, Lincoff AM, Martin SS, Blaha MJ. Association of lowering low-density lipoprotein cholesterol with contemporary lipid-lowering therapies and risk of diabetes mellitus: a systematic review and meta-analysis. J Am Heart Assoc. 2019;8(7):e011581.
Tenenbaum A, Motro M, Fisman EZ, Adler Y, Shemesh J, Tanne D, Leor J, Boyko V, Schwammenthal E, Behar S. Effect of bezafibrate on incidence of type 2 diabetes mellitus in obese patients. Eur Heart J. 2005;26(19):2032–8.
Lin D, Qi Y, Huang C, Wu M, Wang C, Li F, Yang C, Yan L, Ren M, Sun K. Associations of lipid parameters with insulin resistance and diabetes: a population-based study. Clin Nutr. 2018;37(4):1423–9.
Wang YL, Koh WP, Talaei M, Yuan JM, Pan A. Association between the ratio of triglyceride to high-density lipoprotein cholesterol and incident type 2 diabetes in Singapore Chinese men and women. J Diabetes. 2017;9(7):689–98.
Ramezankhani A, Azizi F, Hadaegh F. Sex differences in rates of change and burden of metabolic risk factors among adults who did and did not go on to develop diabetes: two decades of follow-up from the tehran lipid and glucose study. Diabetes Care. 2020;43(12):3061–9.
Censin JC, Peters SAE, Bovijn J, Ferreira T, Pulit SL, Mägi R, Mahajan A, Holmes MV, Lindgren CM. Causal relationships between obesity and the leading causes of death in women and men. PLoS Genet. 2019;15(10):e1008405.
Johnson KE, Siewert KM, Klarin D, Damrauer SM, Chang KM, Tsao PS, Assimes TL, Maxwell KN, Voight BF. The relationship between circulating lipids and breast cancer risk: a Mendelian randomization study. PLoS Med. 2020;17(9):e1003302.
Yang XL, Cui ZZ, Zhang H, Wei XT, Feng GJ, Liu L, Liu YZ, Pei YF, Zhang L. Causal link between lipid profile and bone mineral density: a Mendelian randomization study. Bone. 2019;127:37–43.
Kelly DM, Jones TH. Testosterone and obesity. Obes Rev. 2015;16(7):581–606.
Tan VY, Bull CJ, Biernacka KM, Teumer A, Richardson TG, Sanderson E, Corbin LJ, Dudding T, Qi Q, Kaplan RC, et al. Investigation of the interplay between circulating lipids and IGF-I and relevance to breast cancer risk: an observational and Mendelian randomization study. Cancer Epidemiol Biomarkers Prev. 2021;30(12):2207–16.
Larsson SC, Michaëlsson K, Burgess S. IGF-1 and cardiometabolic diseases: a Mendelian randomisation study. Diabetologia. 2020;63(9):1775–82.
Wang W, Tesfay EB, van Klinken JB, van Willems DK, Bartke A, van Heemst D, Noordam R. Clustered Mendelian randomization analyses identify distinct and opposing pathways in the association between genetically influenced insulin-like growth factor-1 and type 2 diabetes mellitus. Int J Epidemiol. 2022. https://doi.org/10.1093/ije/dyac119.
Aggarwal NR, Patel HN, Mehta LS, Sanghani RM, Lundberg GP, Lewis SJ, Mendelson MA, Wood MJ, Volgman AS, Mieres JH. Sex differences in ischemic heart disease: advances, obstacles, and next steps. Circ Cardiovasc Qual Outcomes. 2018;11(2):e004437.
Hokanson JE, Austin MA. Plasma triglyceride level is a risk factor for cardiovascular disease independent of high-density lipoprotein cholesterol level: a meta-analysis of population-based prospective studies. J Cardiovasc Risk. 1996;3(2):213–9.
Sarwar N, Danesh J, Eiriksdottir G, Sigurdsson G, Wareham N, Bingham S, Boekholdt SM, Khaw KT, Gudnason V. Triglycerides and the risk of coronary heart disease: 10,158 incident cases among 262,525 participants in 29 Western prospective studies. Circulation. 2007;115(4):450–8.
Minelli C, Del Greco MF, van der Plaat DA, Bowden J, Sheehan NA, Thompson J. The use of two-sample methods for Mendelian randomization analyses on single large datasets. Int J Epidemiol. 2021. https://doi.org/10.1093/ije/dyab084.
Burgess S, Davies NM, Thompson SG. Bias due to participant overlap in two-sample Mendelian randomization. Genet Epidemiol. 2016;40(7):597–608.
Schooling CM, Lopez PM, Yang Z, Zhao JV, Au Yeung SL, Huang JV. Use of multivariable Mendelian randomization to address biases due to competing risk before recruitment. Front Genet. 2020;11:610852.
Lopez PM, Subramanian SV, Schooling CM. Effect measure modification conceptualized using selection diagrams as mediation by mechanisms of varying population-level relevance. J Clin Epidemiol. 2019;113:123–8.
Acknowledgements
The authors acknowledge the UK Biobank (Neale lab), Biobank Japan, GLGC, DIAMANTE, MAGIC, AGEN, Ruth KS, et al. and Schmitz D, et al. for their publicly available summary data.
Funding
This study received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
GYY analyzed the data, wrote the manuscript and contributed to the study design. CMS contributed to the study design, reviewed and edited the manuscript. GYY is the guarantor of this work and takes responsibility for the integrity of the data analysis. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
: Table S1. Mendelian Mendelian randomization estimates for sex-specific associations of genetically predicted lipid traits (instrumented by the SNPs from the UK Biobank) with coronary artery disease in East Asians. Table S2. Odds ratio or mean difference in z-score that was detectable at 80% power (α = 0.05) for each analysis. Table S3. Mendelian randomization estimates for ancestry- and sex-specific associations of genetically predicted triglycerides (instrumented by the SNPs from GLGC excluding the UK Biobank participants) with type 2 diabetes, glycemic traits and sex hormones. Table S4. Mendelian randomization estimates for ancestry- and sex-specific associations of genetically predicted lipid traits (instrumented by the SNPs from the UK Biobank) with type 2 diabetes (including SNPs explaining more of the variance in the outcome than in the exposure). Table S5. Mendelian randomization estimates for sex-specific associations of genetically predicted lipid traits (instrumented by the SNPs from the UK Biobank) with glycemic traits in people of European ancestry (including SNPs explaining more of the variance in the outcome than in the exposure). Table S6. Mendelian randomization estimates for ancestry- and sex-specific associations of genetically predicted lipid traits (instrumented by the SNPs from the UK Biobank) with type 2 diabetes (excluding SNPs explaining more of the variance in the outcome than in the exposure). Table S7. Mendelian randomization estimates for sex-specific associations of genetically predicted lipid traits (instrumented by the SNPs from the UK Biobank) with glycemic traits in people of European ancestry (excluding SNPs explaining more of the variance in the outcome than in the exposure). Table S8. Mendelian randomization estimates for sex-specific associations of genetically predicted lipid traits (instrumented by the SNPs from the UK Biobank) with sex hormones in people of European ancestry (including SNPs explaining more of the variance in the outcome than in the exposure). Table S9. Mendelian randomization estimates for sex-specific associations of genetically predicted lipid traits (instrumented by the SNPs from the UK Biobank) with sex hormones in people of European ancestry (excluding SNPs explaining more of the variance in the outcome than in the exposure).
Additional file 2
: Table S10. Genetic variants for apolipoprotein B and their associations with type 2 diabetes, glycemic traits and sex hormones in women. Table S11. Genetic variants for apolipoprotein B and their associations with type 2 diabetes, glycemic traits and sex hormones in men. Table S12. Genetic variants for triglycerides and their associations with type 2 diabetes, glycemic traits and sex hormones in women. Table S13. Genetic variants for triglycerides and their associations with type 2 diabetes, glycemic traits and sex hormones in men. Table S14. Genetic variants for lipoprotein(a) and their associations with type 2 diabetes, glycemic traits and sex hormones in women. Table S15. Genetic variants for lipoprotein(a) and their associations with type 2 diabetes, glycemic traits and sex hormones in men. Table S16. Genetic variants for multivariable Mendelian randomization analyses and their associations with type 2 diabetes, glycemic traits and sex hormones in women. Table S17. Genetic variants for multivariable Mendelian randomization analyses and their associations with type 2 diabetes, glycemic traits and sex hormones in men.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yang, G., Schooling, C.M. Investigating sex-specific associations of lipid traits with type 2 diabetes, glycemic traits and sex hormones using Mendelian randomization. Cardiovasc Diabetol 22, 3 (2023). https://doi.org/10.1186/s12933-022-01714-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12933-022-01714-2