
Abstract
Background
In ayurvedic traditional medicine Gentiana kurroo Royle (family; Gentianaceae) is used to treat several metabolic diseases. This plant is rich in various compounds belonging to flavonoids and glycosides. Till now little work has been carried out on immunomodulatory and anti-inflammatory potential of this plant. This study confirms the presence of bioactive compounds and evaluates the anti-inflammatory and immunomodulatory effect of this plant.
Methods
To carry out this work, the methanol extract was investigated in different doses using in vivo and in vitro models. In vivo study involved haemagglutination titre and DTH methods, and in vitro study was done using splenocyte proliferation assay and LPS stimulated macrophage culture. TNF-α, IL-6 and NO were assayed using ELISA kit methods, while NF-κB was evaluated by western blotting. LC-ESI-MS/MS was used for the characterization of the methanol extract.
Results
The results showed suppression of both humoral and cell mediated immunity in vivo. This effect was also observed by inhibition of B and T cell proliferation in splenocyte proliferation assay. TNF-α, IL-6 and NO concentrations were also less in extract treated macrophage cultures. The NF-κB expression was also lowered in treated macrophages as compared to untreated macrophages. All these observations were found to be dose dependent. LC-MS characterization of this extract showed the presence of known compounds which are glycosides, alkaloids and flavonoids in nature.
Conclusion
The methanol extract of this plant was found to be rich in glycoside, alkaloid and flavonoid compounds. These compounds are probably responsible for the suppression of immune response and anti-inflammatory activity. The extract as such and identified bioactive compounds can be useful for the treatment of inflammatory disorders.
Similar content being viewed by others

Background
Inflammation is an essential protective process preserving the integrity of organisms against physical, chemical, and infective insults. Inflammatory diseases majorly develop by altering the response of T-cells and causing irregularity in immune system function starting with the migration of leukocytes, primarily neutrophils [1]. Further, during inflammation the pro-inflammatory molecules like Nitric oxide (NO) and TNF-α are produced by macrophages and the inhibition of NO synthase and TNF-α reverses inflammatory symptoms [2]. This inflammatory response is transduced by a variety of signalling pathways. Nuclear factor kappa B (NF-κB) is one of the essential signalling molecules acting as a transcription factor for the expression of iNOS, TNF-α and IL-6 genes by binding to their promoter regions [3, 4]. Many synthetic drugs like glucocorticoids are used for the treatment of inflammatory disorders by immune system modulation, but none of them is without serious adverse effects and are thus highly unsafe for use in humans. The different synthetic drugs used against inflammation are reported to produce drug related toxicities, adverse reactions and iatrogenic effects complicating the treatment process [5, 6]. Hence, anti-inflammatory treatment has observed a shift from synthetic to herbal products. These herbal products with fewer side effects are mainly the primary and secondary metabolites [7]. Such herbal drugs with the capacity to inhibit cellular and humoral immune responses can have useful applications in some immune-mediated disorders including autoimmune diseases [8].
In this perspective the current study was carried out on Gentiana kurroo Royle used as an effective drug in ayurveda for the treatment of several metabolic diseases [9]. Further the flower tops of this plant has been shown to possess the anti-inflammatory potential [10]. In our earlier studies different extracts of this plant were screened for anti-inflammatory and immuno-modulatory potential and methanol extract was found to show the maximum potential [11]. So in the current study this extract was extensively assesssed for its effect on immune response to explore its protective mechanism against inflammation.
Methods
Collection, identification and selection of herbal substance
Gentiana kurroo Royle locally known as Nilkanth (English name- Indian Gentian) was procured from sub-alpine region in Dachigam and identified in the Centre of Plant Taxonomy (COPT), Dept. of Botany, University of Kashmir. The specimen is retained in the herbarium of COPT vide voucher no.1804-KASH. Whole plant material was selected for the study.
Preparation of extracts
The plant material was cleaned, cut into small pieces and dried under a shade. It was then pulverized into coarse powder in an electric blender, weighed (~180 g dry weight) and extracted successively using petroleum ether, ethyl acetate, methanol and water respectively by soxhlet extraction each for a period of 96 h. The solvents were allowed to evaporate in a rotary evaporator at 40–45 °C and the extracts obtained were stored in a refrigerator at 4 °C. The yields of the petroleum ether, ethyl acetate, methanol and aqueous extracts were 5.6, 4.3, 5.3, and 4.2% (w/w), respectively.
Animals
Male Balb/C mice 8–10 weeks old and weighing 18–22 g, were used for the immuno-modulatory study. The animals were housed under standard laboratory conditions with a temperature of (25 ± 2)°C, relative humidity of (55 ± 10)%, 12/12 h light–dark cycles and fed with standard pellet diet (Lipton India Ltd.) and water was given ad libitum.
Experimental protocols
All experimental protocols and the number of animals used for the experimental work were duly approved by the Institutional Animals Ethics Committee (IAEC); of Indian Institute of Integrative Medicine (CSIR), Canal Road Jammu (CPCSEA registration No. 67/CPCSEA/99). The experimental work on animals was carried out as per the guidelines set by committee for the purpose of control and supervision of experiments on animals (CPCSEA).
Preliminary toxicity study
Five male Balb/C mice 8–10 weeks old were taken. Each animal was tested with the methanol extract at an oral dosage of 2000 mg/kg bw as per OECD guidelines. The mice were observed for 24 h for any signs of toxicity including change in behaviour or death.
Humoral antibody response
The mice were divided into 6 groups, each group consisting of 5 animals. Mice in group I (control) were given 1% tween 20, 0.2 mL/mice for 14 days. Mice in group II-V were given the extract at a dose range of 25, 50, 100, 200 mg/kg bw respectively for 14 days. Mice in group VI were given cyclophosphamide (50 mg/kg) on day 1 and continued for 14 days. All drugs were given orally. The animals were immunised by injecting 200 μL of 5 × 109 SRBC’s/mL intraperitonially (i.p) on day 1. Blood samples were collected in microliter tubes from individual animals in all the groups by retro-orbital vein puncture on day 7 and day 14. The blood samples were centrifuged and the serum was separated. Then haemoglutination primary and secondary titres were performed [12, 13].
Delayed type hypersensitivity
This method was carried out to determine the effect of extracts on the cell-mediated immunity. Animals were divided into six groups of 5 each. Group I served as sensitized control, as in humoral antibody titre. Mice in group II-V were administered methanol extract at a dose range of 25, 50, 100, 200 mg/kg bw respectively after SRBC’s sensitization (i.p), once daily for 14 days. Cyclophosphamide (50 mg/kg bw) was administered as standard T-cell suppressor (group VI). The mice were challenged on day 14 by injecting 0.02 mL of SRBC’s antigen subcutaneous into the right hind footpad, whereas left hind footpad served as control [14, 15]. The footpad thickness was measured with micrometre calliper (pitch 0.01 mm) after 24 and 48 h of SRBC’s challenge.
Ex-vivo TNF-α assay
The male Balb/C mice dosed for 14 days with variable concentrations (25–200 mg/kg) of methanol extract were analysed for serum TNF-α. Blood (0.5 mL) was taken on the 14th day by retro-orbital puncture from the mice used for the study of DTH reaction. Blood was kept on standing for 2 h at room temperature (24 °C). After clotting of blood, the tubes were centrifuged at 8000 rpm for 10 min, at 4 °C. The serum so collected was evaluated for the measurement of TNF-α using mouse TNF-α ELISA kit (R&D systems, USA), as per the instructions of the manufacturer.
Collection of peritoneal macrophages and nitrite assay
Male Balb/C mice were i.p. injected 4% sodium thioglycollate media 1 mL per mice. Under aseptic conditions mice were euthanized after 24 h and injected 10 mL of RPMI 1640 medium in peritoneal cavity. After 5 min, the medium was taken out and centrifuged at 1800× g for 10 min at 4 °C. The cell pellet was resuspended in RPMI 1640 medium. The macrophages (3× 106) were cultured in 96 well micro-plate in the presence of LPS (1 μg/mL), β-Methasone (0.001 μg/mL) and variable doses of methanol extract (25–200 μg/mL). The plates were incubated at 37 °C in a humid saturated atmosphere containing 5% CO2 for 24 h. Supernatants were collected after centrifugation and stored at −80 °C for determination of cytokines. The supernatants were used for nitrite assay using Griess reagent. For the NO2 assay (nitrite content), 100 μl of the culture media was incubated with 100 μL Griess reagent (0.1% naphthyl-ethylenediamide and 1% sulphanilamide in 2.5% phosphoric acid solution) at room temperature for 10 min in 96 well micro-plate [16]. Optical density was measured at 540 nm using ELISA plate reader. The nitrite concentration was determined by extrapolation from a sodium nitrite (NaNO2) standard curve and the results expressed in μM.
IL-6 and TNF-α measurements
The harvested supernatants from cultured peritoneal macrophages were used for the measurement of pro-inflammatory cytokines (IL-6 and TNF-α) using mouse IL-6 and TNF-α ELISA kits (R&D systems, USA), as per the instructions of the manufacturer.
Lymphocyte proliferation studies on mice splenocytes
Spleen collected under aseptic conditions in IRPMI, was minced using a pair of scissors and passed though a fine steel mesh to obtain a homogenous cell suspension and the erythrocytes were lysed with ammonium chloride (0.8% w/v). After centrifugation (380 × g at 4 °C for 10 min), the pelleted cells were washed three times with PBS and resuspended in complete medium [RPMI 1640 supplemented with 12 mM HEPES (pH 7.1), 0.05 mM 2-mercaptoethanol, 100 IU/mL penicillin, 100 μg/mL streptomycin and 10% FBS]. The cell number was counted with a haemocytometer by the trypan blue dye exclusion technique. Cell viability exceeded 95% [17].
To evaluate the effect of methanol extract on the proliferation of splenic lymphocytes, the spleen cell suspension (2 × 106 cells/mL) was pipetted into 96 well plates (200 μl/well) and cultured at 37 °C for 72 h in a humid saturated atmosphere containing 5% CO2 in the presence of Con-A (5 μg/mL), LPS (1 μg/mL) and variable doses of methanol extract (25–200 μg/mL). β-methasone (BMS) was used as standard drug at a concentration of 0.001 μg/mL. After 72 h, 20 μL of MTT (5 mg/mL) was added to each well and plates were incubated for 4 h. The plates were centrifuged (1400 × g, 5 min) and the untransformed MTT was removed carefully by pippeting. To each well, 100 μl of DMSO working solution (192 μl DMSO with 8 μl 1 M HCl) was added and the absorbance was evaluated in ELISA reader at 570 nm after 15 min. Reduction of MTT indicated the splenocyte proliferation.
Western blot analysis
Peritoneal macrophages collected from male Balb/C (0.5 × 106 cells) were incubated overnight and exposed to different concentrations of methanol extract (25–100 μg/mL) with DMSO as vehicle and β-methasone (0.001 μg/mL) as positive control. LPS (1 μg/mL) was also added to all wells except the vehicle, thus having two vehicles - DMSO as vehicle (unstimulated), vehicle + LPS (stimulated) so as to have a better understanding of the results. Cells were accordingly rinsed with PBS, trypsinized and collected after 24 h incubation and were lysed with lysis buffer (HEPES 1 mM/L, KCl 60 mM/L, NP-40 0.3%, EDTA 1 mM/L, DTT 1 mM/L, Sodium orthovandate 1 mM/L, PMSF 0.1 mM/L, cocktail protease inhibitor). The cell extractions were centrifuged at 12,000 rpm for 10 min at 4 °C. Supernatant was collected for protein quantification. Protein concentration was determined by the standard Bradford method [18]. Equal amount (20 μg) of protein from each sample was subjected to SDS-PAGE (12%) and proteins were transferred to membrane (Millipore), blocked with 5% (w/v) non-fat milk in PBS containing 0.1% Tween-20 and probed with anti-NF-кB (p65) and anti-β-actin for 3 h at room temperature overnight at 4 °C and subsequently washed, probed with species specific secondary antibodies coupled to horseradish peroxidase. Immuno-reactive proteins were detected by enhanced chemiluminescence plus (Amersham).
Liquid chomatography–tandem mass spectrometry (LC–ESI-MSMS)
LC–MS equipment (LC–MS QqQ-6410B Agilent Technologies) comprised a chomatographic system (1260 Infinity Agilent Technologies) coupled with an Agilent Triple Quad mass spectrometer fitted with an ESI source. MS conditions were the following: MS range 100–1200 Da, MSn spectra were obtained using both positive and negative modes, nebulizer gas 45 Psi, gas temperature 325 °C, capillary voltage 4000 V.
HPLC analysis was carried out by an Agilent 1260 infinity series. A Chomolith RP-18e column (4.6 mm ID, 50 mm length) (Merck) was used. Mobile phase consisted of (A) aqueous formic acid (0.1%) and (B) methanol. Gradient condition was; 0–8 min, linear gradient from 12 to 25% of B; 8–12 min, isocratic conditions at 25% of B; 12–16 min, linear gradient from 25 to 40% of B; 16–40 min, linear gradient from 40 to 50% of B, 40–50 min, linear gradient from 50 to 100% of B. Flow rate: 1 mL/min.
Statistical analysis
Data is expressed as Mean ± SD and statistical analysis was carried out employing the one-way ANOVA followed by Post Hoc Tukey’s HSD test and Bonferroni multiple comparison test using SPSS 16 software. p values < 0.05 are being taken as statistically significant.
Results
Preliminary toxicity study
The methanol extract was found to be non-toxic at a dose of 2000 mg/kg bw.
Haemagglutination antibody titre
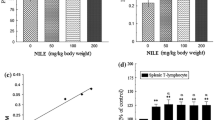
In the current study as is evident from Table 1, the methanol extract showed increased activity with the increase in dose. The effect was found to be more in primary titre than in the secondary titre. At a dose of 200 mg/kg the methanol extract had maximum effect on HA titre and also more than cyclophosphamide. The inhibition at this dose by methanol extract in primary and secondary humoral response observed was 66.21 and 55.26% respectively (Fig. 1). Cyclophosphamide treated group showed 39.39 and 52.63% inhibitory response in the primary & secondary titres. Statistical analysis showed significance at p < 0.05, of the results obtained between the control and the treated groups.

Graph showing %age inhibition of primary and secondary humoral response in Balb/C mice by methanol extract (25–200 mg/kg). Cyclophosphamide (50 mg/kg) was taken as standard drug
Delayed type hypersensitivity
DTH was carried out to study the effect of different doses of methanol extract on non-specific cellular immune response. Results obtained showed dose dependent effect in cellular immune response (Table 2). In control group edema formation was high, but was found to be less in treated groups. In positive control group the edema formation was less than control group but methanol extract treated group at a dose of 200 mg/kg had least edema formation with an inhibition of 70.33 & 75.66% in cellular response after 24 & 48 h respectively, while in cyclophosphamide treated group (50 mg/kg bw) the suppression in cellular immune response was 58.52 & 65.61% as compared to control group (Fig. 2). The results obtained were found to be significant in relation with control group at p < 0.05.

Graph showing 24 and 48 h %age inhibition of cell mediated response in Balb/C mice by methanol extract (25–200 mg/kg). Cyclophosphamide (50 mg/kg) was taken as standard drug
Ex-vivo TNF-α assay
The results indicated a decrease in serum TNF-α levels of treated groups as compared to control in a dose dependent manner. The group administered with methanol extract at a dose range of 200 mg/kg significantly (p < 0.05) decreased serum TNF-α (174.96 pg/mL) as compared to control (468.09 pg/mL). Cyclophosphamide treated group significantly (p < 0.05) decreased the TNF-α in serum (Fig. 3).

Effect of methanol extract on serum TNF-α concentration in male Balb/C mice. The results in graph include control group, methanol extract treated groups (25–200 mg/kg) and positive control group treated with cyclophosphamide (50 mg/kg). Values are mean ± SD (n = 5); *p < 0.05 (control vs. treated groups); one-way ANOVA followed by Bonferroni multiple comparison test
Effect of methanol extract on NO production in macrophages
Since NO is also known as a pro-inflammatory mediator in different acute and chronic inflammatory diseases, we addressed whether methanol extract modulates the NO production from macrophages stimulated by LPS. The extract inhibited the NO production in a dose dependent manner at a dose range of 25–200 μg/mL. The maximum inhibition was observed at 200 μg/mL dose compared to control. Increase in the dose of methanol extract indicated a significant inhibition in the production of nitrite content (Fig. 4). BMS also showed a significant inhibition (p < 0.05) in nitrite content compared to control.

Effect of methanol extract on the nitrite content in mice peritoneal macrophages. Values are means ± SD; *p < 0.05 (control vs. treated groups); one-way ANOVA followed by Bonferroni multiple comparison test. All tests were carried out in triplets and repeated twice
Effect of methanol extract on the TNF-α and IL-6 release in mice peritoneal macrophages stimulated with LPS
Results indicated that the extract inhibited the release of these two cytokines from macrophages in a dose dependent manner. At a dose of 200 μg/mL, inhibition in TNF-α (Fig. 5a) and IL-6 (Fig. 5b) was found to be most significant (p < 0.05). BMS also registered a significant inhibition (p < 0.05) in the release of TNF-α and IL-6 from LPS stimulated macrophages compared to control.

a Effect of methanol extract on TNF-α release in mice peritoneal macrophages. Values are mean ± SD., *p < 0.05 (control vs. treated groups); one-way ANOVA followed by Bonferroni multiple comparison test. All tests were carried out in triplets and repeated twice. b Effect of methanol extract on IL-6 release in mice peritoneal macrophages. Values are mean ± SD., *p < 0.05 (control vs. treated groups); one-way ANOVA followed by Bonferroni multiple comparison test. All tests were carried out in triplets and repeated twice
Effect of methanol extract on splenocyte proliferation assay
The results observed were found to show suppression of both T and B-cell proliferation of splenocytes at doses (25–200 μg/mL) in a dose dependent manner compared to control. Maximum observation was registered at a dose of 200 μg/mL (Fig. 6). The extract indicated more inhibition of T-cell than the B-cell; however the difference was not so high. β-methasone (0.001 μg/mL) showed significant inhibition (p < 0.05) of both T and B-cell proliferation compared to control.

Effect of methanol extract on ConA and LPS-stimulated splenocyte cell proliferation. Values are mean ± SD showing OD at 570 nm, *p < 0.05 (control vs. treated groups); one-way ANOVA followed by Bonferroni multiple comparison test. All tests were carried out in triplets and repeated twice
Effect of methanol extract on NF-kappa B expression in mice peritoneal macrophages stimulated with LPS
In presence of LPS (stimulated) increased concentrations of NF- кB (p65) were registered in protein extracts of peritoneal macrophages after 24 h as compared to vehicle (unstimulated). The results indicated reduced expression of NF-kappa B (p65) in presence of the methanol extract. The methanol extract was used in a dose range of 25–100 μg/mL and showed dose-dependent inhibition in the expression of NF-kappa B (p65). At a dose of 100 μg/mL maximum inhibition was observed up to 0.23 fold decrease in its expression (Fig. 7). BMS used as standard drug showed significant decrease in the expression of NF-kappa B (p65).

NF- КB (p65) expression in mice peritoneal macrophages. Extract treated cultures were found to show decreased expression of NF-kappa B (p65) as compared to stimulated and unstimulated vehicle. β-actin was used as loading control
LC–ESI-MS/MS analysis
The methanol extract was analyzed for the presence of different compounds by liquid chomatography tandem mass spectrometry (LC-ESI-MS/MS). The total ion MS chomatogram (TIC) as shown in Fig. 8 showed a number of peaks corresponding to different chemical constituents present in the extract. The major peaks were further analysed under both positive and negative ESI-MS modes. Further the molecular ion peaks along with the MS fragmentation pattern obtained were compared with the molecular ion peaks of different compounds reported in the literature for the identification of the compounds [19–22]. The compounds identified are given in the Table 3 showing their actual mass (m/z), mass obtained in both ESI+/ESI− modes, retention time and area sum%. The compounds in the ESI+ mode [M + Na]+ at m/z 399.00, 397.00, 379.00, 279.00 coincides with loganic acid (m/z = 376.13), swertiamarin (m/z = 374.12), gentiopicroside (m/z = 356.11), gentisin (m/z = 256.07) respectively. In ESI+ [M + K]+ mass spectra mode at m/z 214.90 single compound gentianine (m/z = 175.06) was identified. The compounds sweroside (m/z = 358.12) and norswertianolin (m/z = 422.07) were identified in the ESI+ [M + H]+ mass spectra mode at m/z 359.10 and 423.00, while in ESI− [M – H]− mass spectra mode at m/z 1007.10, 444.90, 551.20 and 255.10 match with the measured masses of 4′′-O-β-D-glucosyl-6′-O-(4-O-β-D- glucosyl caffeoyl) linearoside (m/z = 1008.31), swertisin (m/z = 446.12), gentioside (m/z = 552.14) and isogentisin (m/z = 256.07) (Fig. 9).

Total Ion Chomatogram (TIC) of the methanol extract

Structures of the various identified compounds in the methanol extract. 1 4″-O-β-D-glucosyl-6′-O-(4-O-β-D-glucosylcaffeoyl)-linearoside. 2 Loganic acid, 3 Swertiamarin, 4 Gentiopicroside, 5 Swertisin, 6 Sweroside, 7 Norswertianolin, 8 Gentisin, 9 Gentioside, 10 Isogentisin, 11 Gentianine
Discussion
Inflammation is a protective process that is essential for the preservation of the integrity of the organism in the event of physical, chemical and infectious damage. Often, it is found that inflammatory response to severe lesions erroneously damages normal tissue [23]. It is well established that activated immunocytes are involved in inflammation process, particularly macrophages, which play a crucial role in specific and nonspecific immune responses during inflammation [24]. So the methanol fraction was used to demonstrate the effect on the immune system using HA titre and DTH method for determining its effect on humoral and cell mediated immune responses respectively. The methanol fraction was observed to have an immunosuppressive effect by inhibiting the antibody formation and the cellular immune response in a dose dependent manner. One of the major pro-inflammatory cytokine is TNF-α which is produced by different immune cells and its rate of production and hence its concentration in blood demonstrates the extent of inflammatory response [25]. Blood samples were taken from the treated and untreated groups of animals used in the HA titre and DTH models. The results indicated marked difference in the concentration of serum TNF-α between the treated and untreated groups. Further the serum TNF-α concentration in serum of treated animals showed decrease with increase in dose of the methanol extract and hence the dose dependent effect. During the progression of inflammation, the most known important mediators produced by macrophages include NO, prostanoids, TNF-\( \alpha \), and Interleukins [26, 27]. Among these inflammatory cytokines, TNF-\( \alpha \), IL-1\( \beta \), and IL-6 are considered the most crucial mediators for the inflammatory process, mediating immunity and activating macrophages [28, 29]. Lipopolysaccharide (LPS)-stimulated macrophages using Raw264.7 cells is a well-established in vitro inflammation model [29, 30]. This model was used for investigating suppressive effect of methanol extract on the release of inflammatory mediators-NO, TNF-α and IL-6 by LPS activated macrophages. The extract inhibited the production of these mediators in a dose dependent manner from LPS stimulated macrophages. These results infer that the contribution of methanol extract in anti-inflammatory activity might be by inhibition of the production of these pro-inflammatory mediators from immune cells. The effect of methanol extract on the proliferation of B-cells and T-cells in response to LPS and ConA respectively, was investigated using MTT assay. The methanol extract was found to significantly inhibit the proliferation of both cells in a dose dependent manner. The extract indicated more inhibition of T-cell than the B-cell; however the difference was not so high. These results are consistent with above results where the methanol extract inhibited the humoral and cell mediated immune response. It is well known that B-cells produce antibodies in response to the antigens while T-cells help in framing the cellular response against the antigens. Because NF-\( \kappa \)B acts as transcription factor and thus causes induction of iNOS, TNF-\( \alpha \), and IL-6 genes by binding to their promoter regions [3, 4, 31–33]. So it was essential to investigate its expression while studying the production of these cytokines. The expression of NF-кB (p65) in mice peritoneal macrophages stimulated with LPS was reduced in presence of different concentrations of the methanol extract. So the reduced production of the pro-inflammatory mediators observed above may be attributed to inhibition in the expression of NF-кB (p65) and hence the effect against the inflammation.
The plant secondary metabolites - Iridoids and flavonoids are reported to have anti-inflammatory potential [34, 35]. Various compounds belonging to Iridoids and flavonoids were found to be present in methanol extract. Among them loganic acid, swertiamarin and gentianine are reported to possess the anti-inflammatory potential [36–38]. The anti-inflammatory potential of methanol extract is probably due to these compounds.
Conclusion
Our research results demonstrate that G. kurroo possess anti- inflammatory activity. This activity was mediated by suppression of immune system- where both the immune cells as well as the cytokines were found to be inhibited. The plant extract as such and the identified bioactive compounds like the loganic acid, swertiamarin and gentianine may be used as natural anti-inflammatory compounds.
Abbreviations
- %:
-
Percent
- BMS:
-
Betamethasone
- bw:
-
Body weight
- C:
-
Degree centigrade
- DMSO:
-
Dimethyl sulfoxide
- ELISA:
-
Enzyme linked immunosorbent assay
- g:
-
Gram
- h:
-
Hour
- HCl:
-
Hydrochloric acid
- IL-6:
-
Interleukin 6
- iNOS:
-
Induced nitric oxide synthase
- kg:
-
Kilogram
- LPS:
-
Lipopolysaccharide
- mg:
-
Milligram
- min:
-
Minutes
- mL:
-
Millilitre
- mM:
-
Millimolar
- NF-\( \kappa \)B:
-
Nuclear factor kappa B
- NO:
-
Nitric oxide
- PBS:
-
Phosphate buffered saline
- pg:
-
Pictogram
- SD:
-
Standard deviation
- SRBC:
-
Sheep red blood cells
- TNF-α:
-
Tumor necrosis factor alpha
- μg:
-
Microgram
- μl:
-
Microlitre
References
Bettini M, Vignali DA. Regulatory T cells and inhibitory cytokines in autoimmunity. Curr Opin Immunol. 2009;21(6):612–8.
Amin AR, Vyas P, Attur M, Leszcynsca-piziak J. The mode of action of aspirin-like drugs: effect on inducible nitric oxide synthase. Proc Natl Acad Sci U S A. 1995;92(17):7926–30.
Chen F, Kuhn DC, Sun SC, Gaydos LJ, Demers LM. Dependence and reversal of nitric oxide production on NF-\( \kappa \)B in silica and lipopolysaccharide-induced macrophages. Biochem Biophys Res Commun. 1995;214(3):839–46.
Yoshimura A. Signal transduction of inflammatory cytokines and tumor development. Cancer Sci. 2006;97(6):439–47.
Lanas A. Nonsteroidal anti-inflammatory drugs and cyclooxygenase inhibition in the gastrointestinal tract: a trip from peptic ulcer to colon cancer. Am J Med Sci. 2009;338:96–106.
Kandulski A, Venerito M, Malfertheiner P. Non steroidal anti-inflammatory drugs (NSAIDs) - balancing gastrointestinal complications and the cardiovascular risk. Dtsch Med Wochenschr. 2009;134:1635–40.
Souza-Fagundes EM, Gazzinelli G, Parreira GG, Martins-Filho OA, Amarante-Mendes GP, Correa-Oliveira R, Zani CL. In vitro activity of labdane diterpene from Alomia myriadenia (Asteraceae): immunosuppression via induction of apoptosis in monocytes. Int Immunopharmacol. 2003;3(3):383–92.
Mirshafiey A, Khorramizadeh MR, Saadat F, Rehm BHA. Chemopreventive effect of M2000, a new anti-inflammatory agent. Med Sci Monit. 2004;10(10):I105–9.
Shahi VK. A significant of drug of Ayurveda: Vangamaya-katuki. Sachitra Ayurda. 1993;46(1):21–7.
Latif A, Khan TF, Afaq SH. Anti-inflammatory activity of flower tops of Gentiana kurroo Royale extract. Pharmacologyonloine. 2006;3:575–80.
Mubashir K, Ghazanfar K, Ganai BA, Akbar S, Malik AH, Masood A. Scientific validation of Gentiana kurroo Royle for anti-inflammatory and immunomodulatory potential. ISRN Inflammation. 2014;2014:Article ID 701765.
Bhagwat DP, Kharya MD, Sarang B, Kaul A, Kour K, Chauhan PS. Immunosuppressive properties of Pluchea lanceolata leaves. Indian J Pharmacol. 2010;42(1):21–6.
Thakur M, Connellan P, Deseo MA, Morris C, Dixit VK. Immunomodulatory polysaccharide from Chlorophytum borivilianum roots. Evid Based Complement Alternat Med. 2011;2011:Article ID 598521.
Jayathirtha MG, Misha SH. Preliminary immunomodulatory activities of methanol extract of Lelipta alba and Centella asiatica. Phytomedicine. 2004;11:361–5.
Mangathayarua K, Umadevi M, Reddy U. Evaluation of the immunomodulatory and DNA protective activities of the shoots of Cynodon dactylon. J Ethnopharmacol. 2009;123:181–4.
Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal Biochem. 1982;126:131–8.
Wang YP, Li XY. Effect of astragaloside IV on T, B lymphocyte proliferation and peritoneal macrophage function in mice. Acta Pharmacol Sin. 2002;3:263–6.
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–54.
Anonymous. The wealth of India: a dictionary of Indian raw material and industrial products. New Delhi: Publication and Information Directorate, CSIR. 1956;4:123–5.
Szucs Z, Danos B, Nyiredy S. Comparative analysis of the underground parts of Gentiana species by HPLC with diode-array and mass spectrometric detection. Chomatographia. 2002;56:19–23.
Singh A. Phytochemicals of Gentianaceae: a review of pharmacological properties. International Journal of Pharmaceutical Sciences and Nanotechnology. 2008;1(1):33–6.
Urbain A, Marston A, Marsden-Edwards E, Hostettmann K. Ultra-performance liquid chomatography/time-of-flight mass spectrometry as a chemotaxonomic tool for the analysis of Gentianaceae species. Phytochem Anal. 2009;20:134–8.
Lunardelli A, Leite CE, Pires MGS, de Oliveira JR. Extract of the bristles of Dirphia sp. increases nitric oxide in a rat pleurisy model. Inflamm Res. 2006;55:129–35.
Romeo GR, Lee J, Shoelson SE. Metabolic syndrome, insulin resistance and roles of inflammation: mechanisms and therapeutic targets. Arteriosclerosis, Thombosis, and Vascular Biology. 2012;32:1771–6.
Beutler B, Cerami A. The biology of cachectin/TNF—a primary mediator of the host response. Annu Rev Immunol. 1989;7:625–55.
Watson WH, Zhao Y, Chawla RK. Sadenosylmethionine attenuates the lipopolysaccharide induced expression of the gene for tumour necrosis factor \( \alpha \). Biochem J. 1999;342:21–5.
Kubes P, McCafferty DM. Nitric oxide and intestinal inflammation. Am J Med. 2000;109(2):150–8.
Dai R, Phillips RA, Karpuzoglu E, Khan D, Ahmed SA. Estrogen regulates transcription factors STAT-1 and NF-\( \kappa \)B to promote inducible nitric oxide synthase and inflammatory responses. J Immunol. 2009;183(11):6998–7005.
Yang JH, Kim SC, Shin BY, Jin SH, Jo MJ, Jegal KH, Kim YW, Lee JR, Ku SK, Cho IJ, Ki SH. O-methylated flavonols isorhamnetin prevents acute inflammation though blocking of NF-\( \kappa \)B activation. Food Chem Toxicol. 2013;59:362–72.
Kim YW, Zhao RJ, Park SJ, Lee JR, Cho IJ, Yang CH, Kim SG, Kim SC. Anti-inflammatory effects of liquiritigenin as a consequence of the inhibition of NF-\( \kappa \)B dependent iNOS and pro-inflammatory cytokines production. Br J Pharmacol. 2008;154(1):165–73.
Roshak AK, Jackson JR, McGough K, Chabot-Fletcher M, Mochan E, Marshall LA. Manipulation of distinct NF\( \kappa \)B proteins alters interleukin-1\( \beta \)-induced human rheumatoid synovial fibroblast prostaglandin E2 formation. J Biol Chem. 1996;271(49):31496–501.
Liu H, Sidiropoulos P, Song G, Pagliari LJ, Birrer MJ, Stein B, Anrather J, Pope RM. TNF-\( \alpha \) gene expression in macrophages: regulation by NF-\( \kappa \)B is independent of c-Jun or C/EBP\( \beta \). J Immunol. 2000;164(8):4277–85.
Vila-del Sol V, Fresno M. Involvement of TNF and NF-\( \kappa \)B in the transcriptional control of cyclooxygenase-2 expression by IFN-\( \gamma \) in macrophages. J Immunol. 2005;174(5):2825–33.
Aquila S, Giner RM, Recio MC, Speqazzi ED, Rios JL. Anti-inflammatory activity of flavonoids from Cayaponia tayuya roots. J Ethnopharmacol. 2009;121(2):333–7.
Ocampo DC, Gomez SB, Barbosa JRB, Amar RA, Lopez VR. Anti-inflammatory activity of iridoids and verbacoside isolated from Castilleja tenuifloraI. Molecules. 2013;18:12109–18.
Kwak WJ, Kim JH, Ryu KH, Cho YB, Geon SD, Moon CK. Effects of gentianine on the production of pro-inflammatory cytokines in male Sprague-Dawley rats treated with lipolysaccharide (LPS). Biol Pharm Bull. 2005;28(4):750–4.
Wei S, Chi H, Kodama H, Chen G. Anti-inflammatory effect of thee iridoids in human neutrophils. Nat Prod Res. 2012;27(10):911–5.
Saravanan S, Islam VI, Babu NP, Pandikumar P, Thirugnanasambantham K, Chellappandian M, Raj CS, Paulraj MG, Ignacimuthu S. Swertiamarin attenuates inflammation mediators via modulating NF-kB/I-kB and JAK2/STAT3 transcription factors in adjuvant induced arthitis. Eur J Pharm Sci. 2014;56:70–86.
Acknowledgements
The authors are highly thankful to the Indian Institute of Integrative Medicine (IIIM), Jammu, for providing the facilities for accomplishing this research work.
Fundings
The authors declare that they have received no funding for the research reported.
Availability of data and materials
All the available data and material used in this study is presented in the main paper.
Authors’ contributions
KM, BAG, SA and AM designed the study and were involved in drafting and correcting the manuscript. KM and KG carried out the in vivo analysis and in vitro analysis was together carried out with BR. MT carried out the LCMS of the extract and also interpreted their results. All the authors read the manuscript, critically revised it for important intellectual content and approved the final version of the manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable in this section.
Ethics approval and consent to participate
All experimental protocols and the number of animals used for the experimental work were duly approved by the Institutional Animals Ethics Committee (IAEC); of Indian Institute of Integrative Medicine (CSIR), Canal Road Jammu (CPCSEA registration No. 67/CPCSEA/99). The experimental work on animals was carried out as per the guidelines set by committee for the purpose of control and supervision of experiments on animals (CPCSEA).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Mubashir, K., Ganai, B.A., Ghazanfar, K. et al. Anti-inflammatory and immuno-modulatory studies on LC-MS characterised methanol extract of Gentiana kurroo Royle. BMC Complement Altern Med 17, 78 (2017). https://doi.org/10.1186/s12906-017-1593-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-017-1593-7