
Abstract
Background
Posttraumatic stress disorder (PTSD) may stem from the formation of aberrant and enduring aversive memories. Some PTSD patients have recreationally used Cannabis, probably aiming at relieving their symptomatology. However, it is still largely unknown whether and how Cannabis or its psychotomimetic compound Δ9-tetrahydrocannabinol (THC) attenuates the aversive/traumatic memory outcomes. Here, we seek to review and discuss the effects of THC on aversive memory extinction and anxiety in healthy humans and PTSD patients.
Methods
Medline, PubMed, Cochrane Library, and Central Register for Controlled Trials databases were searched to identify peer-reviewed published studies and randomized controlled trials in humans published in English between 1974 and July 2020, including those using only THC and THC combined with cannabidiol (CBD). The effect size of the experimental intervention under investigation was calculated.
Results
At low doses, THC can enhance the extinction rate and reduce anxiety responses. Both effects involve the activation of cannabinoid type-1 receptors in discrete components of the corticolimbic circuitry, which could couterbalance the low “endocannabinoid tonus” reported in PTSD patients. The advantage of associating CBD with THC to attenuate anxiety while minimizing the potential psychotic or anxiogenic effect produced by high doses of THC has been reported. The effects of THC either alone or combined with CBD on aversive memory reconsolidation, however, are still unknown.
Conclusions
Current evidence from healthy humans and PTSD patients supports the THC value to suppress anxiety and aversive memory expression without producing significant adverse effects if used in low doses or when associated with CBD. Future studies are guaranteed to address open questions related to their dose ratios, administration routes, pharmacokinetic interactions, sex-dependent differences, and prolonged efficacy.
Similar content being viewed by others

Background
Posttraumatic stress disorder
The formation of intense and long-lasting aversive memories after threatening or stressful events affects the individual’s quality of life when it triggers the development of posttraumatic stress disorder (PTSD [1, 2];), which is categorized on the DSM-5 as a trauma- or stressor-related disorder [3]. After a traumatic event exposure, PTSD patients gradually present characteristic symptoms, such as increased anxiety, hyperarousal, and avoidance of cues associated with the trauma. The inappropriate expression of fear-related responses in non-risky situations is also frequent. Moreover, intrusive thoughts, nightmares, and resistance to extinguish the aversive/traumatic memory have been reported [4,5,6].
Fear memory extinction and reconsolidation
Fear extinction is a form of inhibitory learning that suppresses the expression of the original aversive/traumatic memory. As a consequence, individuals express less fear responses. In both laboratory animals and healthy humans, prolonged and repeated exposures to conditioned cues without presenting the aversive stimulus can induce it [7, 8]. Preclinical studies have shown that the extinction process requires activity and plasticity in several interconnected brain regions, including the infralimbic and prelimbic subregions of the medial prefrontal cortex [homologous to the human ventromedial prefrontal (vmPFC) and dorsal anterior cingulate (dACC) cortices, respectively], and some amygdala nuclei [9,10,11]. Specific PTSD psychotherapies (e.g., prolonged exposure therapy) are based on extinction learning [12]. Patients suffering from this psychiatric condition, however, often present extinction impairments [10, 13] accompanied by a hypoactive vmPFC [14, 15], hyperactive dACC and amygdala [14, 16,17,18], and a smaller and hypofunctional hippocampus [14, 18, 19]. The abnormal functioning of these brain regions could explain not only the hyperarousal and extinction deficits but also the increased responsiveness to trauma-unrelated stimuli leading to fear overgeneralization [20,21,22]. Noteworthy, over time, the original aversive/traumatic memory can spontaneously reemerge, which also limits the efficacy of the extinction approach [23].
Upon recall, a consolidated memory can become labile again and, thus, its content is destabilized and gradually reconsolidated after that, being susceptible to intervention-induced changes during this period. In both laboratory animals and healthy humans, short exposure to conditioned cues can induce memory destabilization and reconsolidation [24]. The neural substrate regulating this process and that underlying extinction is thought to be overlapping, yet distinct [24, 25]. Besides, the relative contribution of a given brain region in each case may vary. For instance, the rodent prelimbic cortex (dACC in humans) is more involved in aversive memory reconsolidation than extinction [11, 25,26,27]. The age and intensity of the memory are factors that influence the chance of destabilization upon retrieval in both laboratory and clinical settings [28,29,30,31]. Unlike extinction, however, changes in the original aversive/traumatic memory-related outcomes are permanent; therefore, impairing fear memory reconsolidation could have value for treating PTSD [32,33,34,35].
In preclinical studies, fear conditioning is a standard procedure for investigating the process of fear memory extinction and reconsolidation. It has a translational value from laboratory animals to healthy humans, and onwards to anxious/PTSD patients. However, the stimuli, the primary outcome measures, and the populations typically used differ between animal and human studies [36]. Besides, whereas in humans it is possible to assess the explicit and implicit aversive memory components, in laboratory animals, only implicit memory-related behavioral, autonomic, and hormonal measures can be assessed [37]. It should also be acknowledged that a strong aversive memory is not necessarily maladaptive. It is indispensable to evaluate not only qualitative but also quantitative aspects to infer whether the selected experimental protocol has indeed simulated one or more PTSD symptoms or features [37]. Similar considerations presumably apply while testing and modeling anxiety in rodents and humans [38, 39] and, thus, the interpretation and extrapolation of basic and clinical findings are not straightforward (Fig. 1).

The process of aversive memory formation, extinction, and reconsolidation. Immediately after an emotionally relevant experience, the acquired memory undergoes the gradual process of consolidation. Upon retrieval, a brief but sufficient conditioned stimulus exposure event induces memory reactivation or destabilization. In other words, the stable memory trace becomes labile again and, thus, its emotional content is modifiable until the reconsolidation stage is ended. Based on this, drugs aiming at interfering with reconsolidation can be administered after memory reactivation. After a prolonged or repeated period of memory retrieval, the extinction process is triggered, leading to the formation of a new memory trace that competes and inhibits the original aversive memory, reducing fear responses. Drugs that potentiate/facilitate memory extinction are usually administered before extinction learning. Other phases (e.g., consolidation) of memory extinction can also be targeted, although it has scarcely been explored. As reviewed here, low doses of THC attenuate aversive memory expression through anxiety reduction, extinction facilitation, and reconsolidation impairment (currently shown in laboratory animals only)
Evidence for the role of the endocannabinoid system in PTSD and its treatment
A low “endocannabinoid tone” has been reported in PTSD patients. Relative to healthy controls, they present reduced circulating concentrations of anandamide and 2-arachidonoylglycerol (2-AG), and lower hair concentration of palmitoylethanolamide (PEA), oleoylethanolamide (OEA) and stearoylethanolamide (SEA) [40,41,42]. They also have up-regulated cannabinoid type 1 (CB1) receptor expression in the hippocampus, dACC, and amygdala, an alteration more pronounced in women than men [40]. The genetics research focusing on variants in the genes of the CB1 receptor and the fatty acid amide hydrolase (FAAH) enzyme, which metabolizes anandamide, has shown corresponding findings. A specific variant resulting in high expression of CB1 receptors can increase the risk of developing PTSD or anxiety-related disorders [43, 44]. However, studies in healthy volunteers have shown that a genetic variant resulting in a lower FAAH activity can influence stress reactivity and fear extinction, being protective against PTSD or anxiety-related disorders [45,46,47]. Despite the advance in understanding PTSD neurobiology, the selective serotonin reuptake inhibitors (SSRIs) are mostly used to manage PTSD symptoms, with some benefit at best. Accordingly, most reviews have concluded that the benefit and effect sizes of these drugs are small [48,49,50]. Besides, PTSD comorbidities include anxiety and substance use disorders, making an effective treatment with SSRIs and psychotherapies even more challenging [51].
There is evidence relating Cannabis use in PTSD patients to relaxation, sleep improvement, attenuation of hyperarousal and anxiety [52,53,54], and reduced values in the Clinician Administered Posttraumatic Scale (CAPS [55];). Similarly, the results of open-label [56], populational [57], and double-blind placebo-controlled [56] studies have shown the benefits of using Δ9-tetrahydrocannabinol (THC), its synthetic version dronabinol or its analog nabilone to manage insomnia and nightmares in PTSD patients. However, contradictory results have also been reported, leading some authors to question the value of this Cannabis-based approach [58,59,60,61]. Differences in dose, route of administration, treatment regimen, level of THC tolerance, and current and past stress may account for the mixed findings above-mentioned [62]. Of note, the potential effects of THC/dronabinol or nabilone on aversive memory extinction and reconsolidation are under investigation. In contrast, it is still unknown whether cannabidiol (CBD), the main compound of Cannabis devoid of psychotomimetic effects, can impair the reconsolidation of aversive memories and facilitate their extinction in humans [63, 64], although its anxiolytic action has already been reported [65,66,67,68]. Similarly, associating THC with CBD could be therapeutically advantageous, but studies focusing on extinction or reconsolidation of aversive/traumatic memories are still incipient.
Based on the above, the present review aims to discuss the effects of THC, its synthetic version dronabinol, or its analog nabilone when administered alone and combined with CBD, on the extinction of aversive memories and anxiety, a common PTSD symptom, in healthy humans and PTSD patients.
Methods
Design
A qualitative systematic review of the research literature was carried out to identify relevant studies addressing the topics of our review.
Study eligibility
Types of studies
Studies presenting primary data from controlled trials in healthy adults, anxious or PTSD patients that evaluated the effects of THC or its association with CBD on fear-related memories or anxiety responses, and published in English were included. The focus was on memory extinction/reconsolidation and anxiety-related responses.
Search strategy
Medline, PubMed, Cochrane Library and Central Register for Controlled Trials databases were searched using the keywords and MeSH terms [Δ9-tetrahydrocannabinol/THC or THC and cannabidiol/THC and CBD] and [Fear extinction/memory extinction/fear memory/memory reconsolidation or anxiety] for human studies and controlled trials published between 1974 and July 2020 in the English language. The reference lists of articles included and previous systematic reviews were checked for relevant publications. Primary studies presenting data from oral or smoked THC or THC plus CBD effects on fear-related memory or anxiety were identified.
Data abstraction
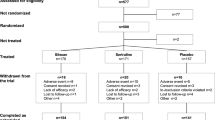
Abstracts were identified and independently analyzed by two reviewers. Three reviewers independently conducted data extraction and coding disagreements arising were discussed, and consensus coding applied. The following information was extracted for each study: age, gender, the health status of subjects, drug(s) used, doses and its management, protocols adopted, and the main results found. Figure 2 depicts the summary of the search process and the study exclusion criteria.

Flowchart of the study selection procedure
Effect size calculation
Means, standard errors, and the number of subjects per group (“n”) were collected to calculate the effect size of treatment in the selected studies (raw data are summarized in Table S1 and S2). When the study presented only the standard deviation as the dispersion measure, the standard error was calculated using the following formula: standard deviation divided by the square root of “n”. When the means and the dispersion measure were depicted in the figure only, the measure tool from the Adobe Acrobat Reader® software was used to calculate them.
The effect size of behavioral results was calculated using the formula for Cohen’s d to reflect the mean-difference (± 95% confidence interval) between two groups. A d ≥ 0.5 and ≤ 0.8 was considered a medium effect size while a large one happened when d > 0.8 [69]. When the study did not present the dispersion measure or mention the “n” per group, the effect size was not calculated. The d values were all presented as positive values, and when the study presented repeated time points of the evaluated parameter, the d expressed in the text was the highest achieved.
Results
Effects of THC/dronabinol on aversive memory extinction or reconsolidation
Table 1 summarizes the main findings from the five double-blind studies investigating the effects of an acute administration of dronabinol on aversive memory extinction in healthy men and women. Neither studies nor clinical trials investigating the THC/dronabinol effects on aversive/traumatic memory reconsolidation were identified.
Oral administration of 7.5 mg of dronabinol before fear extinction potentiated (d = 0.81) the extinction process in subjects submitted to a cued fear conditioning, as inferred by a reduced skin conductance response (SCR) during extinction recall 24 h later [70]. In a similar study [71], there were no dronabinol-induced SCR changes during extinction recall, even though it increased the activation of the vmPFC and hippocampus. In a subsequent study, dronabinol (7.5 mg/kg) attenuated the recovery of fear 24 h after extinction, and during a post-extinction resting period, the drug-treated subjects presented altered state brain dynamics (lowered amygdala-hippocampus static functional connectivity and increased amygdala-vmPFC dynamic functional connectivity) that were associated with a better extinction recall (d not calculated [72];).
Dronabinol effects on extinction have already been evaluated at a more remote time point. In the study by Hammoud et al., 2019 [73], subjects received 7.5 mg of this drug before extinction learning, and the retention test was performed both one and 7 days later. Dronabinol reduced the SCR during the extinction session (d = 0.55) and the retention test on day 1 (d = 0.55), but not on day 7. In the last case, however, the drug-treated group presented a significant increase in the functional coupling among the vmPFC, hippocampus, and dACC, which was associated with lower spontaneous recovery of fear. Moreover, healthy volunteers submitted to a neutral face presentation associated with an aversive sound and treated with 10 mg of dronabinol before extinguishing that association presented a transient reduction in SCR (d = 0.58), but unaltered fear-potentiated startle response [74].
In summary, in four of five studies, dronabinol induced significant changes in extinction-related autonomic or behavioral responses, with medium to large effect sizes (i.e., from 0.55 to 0.81). Therefore, this acute pharmacological intervention is associated with clinically relevant effects. There were no sex-dependent differences in dronabinol-induced facilitating effects on aversive memory extinction. However, such action will possibly require periodic associations of drug administration with extinction to be preserved. Noteworthy, the effects of repeated THC/dronabinol administration on aversive/traumatic memory extinction are still unknown. Despite the current knowledge about aversive memory reconsolidation and the well-documented role of the endocannabinoid system in this process in laboratory animals [75, 76], no human studies have addressed this question yet.
Effects of THC/dronabinol plus CBD on aversive memory extinction or reconsolidation
No human studies or clinical trials investigating the effects of associating THC with CBD on aversive/traumatic memory extinction or reconsolidation were identified. However, they are foreseen since the combination of THC and CBD presents advantages in comparison with THC/dronabinol alone, such as fewer and less intense adverse effects and greater safety [77, 78]. Accordingly, in the studies evaluated, whereas 10–15 mg of THC often induce psychosis in either healthy or susceptible individuals, measured by Positive and Negative Psychotic Syndrome Scale (PANSS) general scores (d = from 0.84 to 2.52 [79,80,81,82,83];), 50 mg of THC associated with CBD in a dose ratio of ~ 1:1 (nabiximols, Sativex®) no longer produced that psychotomimetic effect [78, 84]. The antipsychotic effect of CBD, which can also counteract some other undesired effects related to CB1 receptor activation by THC, probably explains this pattern of results [85, 86]. However, some studies have indicated that CBD might not counteract the THC psychotic effect (for a review, please see [87, 88]).
Effects of THC/dronabinol or its analog nabilone on anxiety-related responses
Table 2 summarizes the main findings from the 17 studies investigating the effects of THC/dronabinol or nabilone on anxiety in healthy humans and patients with PTSD or anxiety disorders.
Oral administration of 2.0 mg of nabilone to healthy subjects with no history of Cannabis consumption produced no changes in the anxiety state measured by the Hopkins Symptom Checklist Scores [89]. In contrast, 7.5 mg of THC decreased both stress reactivity, measured by the Visual Analog Scales (VAS) in the Trier Social Stress Test (d = 0.82), and amygdala activation when viewing fearful faces [90, 91]. THC (7.5 mg) also increased subjective reports of “high”, but neither impairments in task performance nor changes in anxiety, sedation, and arousal, as measured by the State-Trait Anxiety Inventory (STAI) and the Profile of Mood State (POMS), were reported [90, 91]. When administered orally at doses ≥10 mg, THC was reported to increase the anxiety state, as measured by STAI (d = from 0.93 to 2.52 [79,80,81,82,83, 92];), Visual Analogue Mood Scale (VAMS; d = 0.63 [79];), POMS (d = 1.32 [91];), subjective reports [91] or SCR during presentation of neutral and fearful faces (d = from 0.92 to 1.42 [81, 83];). This anxiogenic THC action has been associated with increased activation of CB1 receptors in the right amygdala [80, 81]. It was also accompanied by increased activation in frontal and parietal areas [83].
In healthy subjects with a history of Cannabis consumption, smoking a cigarette containing 1.8% (~ 15 mg) of THC produced no changes in the anxiety state [94, 95]. In contrast, either smoking 3.6% of THC or its oral intake of 5.0 to 15 mg increased the anxiety state measured by STAI and VAS (d not calculated [82, 95, 96];). Similarly, 10 mg of THC given orally increased the anxiety state measured by STAI in both Cannabis users and non-users (d not calculated), but the induction of psychotic symptoms was less intense in the former group [82]. Of note, the brain areas influenced by THC in users and non-users of Cannabis differed significantly [82, 83]. The explanation for the varying pattern of results is still under debate.
In anxious patients, oral administration of 1.0 or 2.0 mg of nabilone attenuated (d not calculated) the anxiety response measured using POMS, an effect accompanied by increased heart rate. At doses of 4.0 and 5.0 mg, nabilone produced orthostatic hypotension without further anxiety reduction (d not calculated [97];). Reduced anxiety was also reported in a study (d not calculated; 100) divided into two phases: the first being open-label with five patients receiving nabilone on an escalating dose regimen starting with 1.0 mg and not exceeding 10 mg, for 28 days; and the second being double-blind, placebo-controlled with 20 patients receiving nabilone 1.0 mg twice a day for 28 days. The adverse effects reported were drowsiness and dry mouth and eyes [98].
In an open-label study using PTSD patients [99], orally administering 5.0 mg of THC twice a day for 21 days improved their anxiety (d = 1.16) and global state. A recent double-blind study [100] investigated PTSD patients, trauma-exposed individuals, and healthy controls acutely treated with THC (7.5 mg). No significant changes were found in the anxiety state, but THC lowered threat-related amygdala reactivity, increased mPFC activation during the threat, and increased mPFC-amygdala functional coupling in PTSD patients [100].
In summary, in most of the studies above described, THC/dronabinol or nabilone induced statistically significant effects on anxiety-related responses, with medium to large effect sizes (i.e., from 0.63 to 2.52). However, the outcome (anxiolytic or anxiogenic effect) depends on the dose, regimen of treatment, and psychiatric status. When considered, there were no sex-related differences in drug-induced effects on anxiety.
Effects of THC/dronabinol plus CBD on anxiety-related responses
As detailed in Table 2, four double-blind studies evaluated the effects of associating THC with CBD on anxiety in healthy users and non-users of Cannabis [92, 93, 95, 96]. CBD was able to attenuate the anxiogenic effect of THC when given in a THC:CBD dose ratio of 1:1 or 1:2 (d = from 4.33 to 4.59), but not 1:0.5 or 1:0.33 [92, 93, 95]. Of note, the THC-induced increase in heart rate frequency was reduced in the presence of CBD (d = 3.41; THC:CBD dose ratio of 1:2 and d = 1.90 THC:CBD dose ratio of 1:1 [91];). Furthermore, the use of a THC:CBD dose ratio of ~ 1:1 high Sativex® (THC 16.2 mg + CBD 15 mg), but not low Sativex® (THC 5.4 mg + CBD 5 mg) induced an increase in anxiety state (d not calculated [96];). Besides, High Sativex induced higher “feeling anxious” parameters (measured by VAS) than Low Sativex (d = 0.58 [96];). Smoking a cigarette containing THC 3.6% (~ 30 mg) and CBD 1.0% (~ 8.5 mg) was associated with less anxiety (d not calculated) than the one containing only THC 3.6%. In contrast, smoking a cigarette with THC 1.8% (~ 15 mg) alone produced no changes in anxiety, but it was increased when the same THC dose was associated with CBD 1.0% [95].
In summary, CBD attenuates or even prevents the anxiogenic action produced by higher THC doses when given in a dose similar to or higher than that of THC. This outcome, however, depends on their absolute quantity and route of administration. Importantly, no study has already evaluated the effect of chronic use of CBD and THC on anxiety. When considered, there were no sex-related differences in THC-induced effects on anxiety, as well as in the potential counteracting CBD action.
Discussion
The facilitating effects of dronabinol (7.5–10 mg) in humans undergoing aversive memory extinction agree with laboratory animal data showing that the activation of CB1 receptors plays a crucial role in fear memory extinction [101,102,103,104]. For instance, THC and CBD treatment facilitates extinction acquisition and recall, respectively [101, 105]. The action of dronabinol above mentioned is also in line with human studies investigating the THC effects on procedures involving threat perception recognition, such as the emotional face-matching task, the facial emotion recognition, and the recognition of emotional pictures. At a dose range from 7.5 to 15 mg, it reduced the threat perception, the recognition of emotional pictures, and fear and anger faces [106,107,108,109]. THC also enhanced the functional connectivity of specific brain areas, such as the amygdala with both rostral anterior cingulate and medial prefrontal cortices, during the emotional face-matching task and extinction recall [106]. This pattern of results suggests that THC/dronabinol interferes with aversive memory processing and its extinction. Of note, impairments in recognition of non-emotional pictures were found after administering 15 mg of THC [110], indicating that high THC doses can affect neutral memory processing.
The studies focusing on aversive memory extinction in healthy humans and laboratory animals are compelling. However, future studies are guaranteed to examine the effects of acute and chronic THC/dronabinol administration on the extinction of traumatic memories. Currently, there are two double-blind and placebo-controlled study investigating THC effects on extinction learning in PTSD patients (ClinicalTrials.gov Identifier: NCT03008005; ClinicalTrials.gov Identifier: NCT04080427). The preliminary results indicate that PTSD patients treated with THC (7.5 mg) exhibited better recall of the extinction of a cued aversive memory, an effect accompanied by increased hippocampal activation [111]. Although this result agrees with that reported in healthy subjects, the traumatic memory was not evaluated and, thus, whether and how THC can attenuate its expression is still unknown. Moreover, as early mentioned, the extinction’s ability to suppress the original aversive/traumatic memory is thought to be temporary. Based on this, it would be interesting to investigate whether THC/dronabinol can hinder or even prevent extinction-related features that limit its clinical usefulness, such as renewal, reinstatement, and spontaneous recovery over time. Furthermore, some studies report that the long-term use of Cannabis by individuals with PTSD is associated with worse clinical outcomes [61, 112], however, investigating the effects of THC either alone or combined with CBD on PTSD symptoms at clinical settings cannot be directly compared with smoking cannabis in an uncontrolled environment, and possibly in a recreational manner. For more details, see 113.
No sex-dependent differences in THC-induced effects on extinction were identified. Women who have PTSD present more CB1 receptor expression and lower hair concentrations of PEA, OEA, and SEA, than men [40, 42]. The CB1 receptor density has been inversely correlated with anandamide and other endocannabinoid levels, implying that men had a high concentration of endogenous ligands, which might be a factor contributing not only to the increased risk of developing stress-related disorders but also to symptom severity in women [40]. In blood samples, the availability of CB1 receptors is also higher in women than men [113]. In rodents, gonadal hormones (e.g., estradiol) regulate the CB1 receptor density [114], transcription [115], and signal transduction [116], which ultimately influences the content of endocannabinoids [117] and their CB1 receptor affinity [118]. Estradiol can facilitate the fear extinction process [119, 120]. Moreover, 4 out of 5 of the studies [70,71,72,73] have reported that women that were not taking hormonal contraceptives were tested during their follicular phase only to avoid between-group effects of sex hormones. When female participants were included in the studies, they were performed about 1 week before menses onset (based on self-reports of last period and cycle length), when the estrogen level is low. This is likely a factor contributing to the lack of significant sex-dependent differences in THC effects on extinction. Of potential relevance to the present discussion are results showing that a deficient conversion of progesterone to its neuroactive metabolite allopregnanolone, which facilitates GABA action on GABAA receptors, in women with PTSD was associated with resistance to extinguish the fear memory [121, 122]. Moreover, pregnanolone, a precursor of progesterone, is a negative allosteric modulator of CB1 receptors [123]. How the interplay between steroid hormones and the endocannabinoid system contributes to fear extinction is currently under investigation. Altogether, studies support sex-dependent fluctuations in some endocannabinoid system constituents involved in extinction processing. However, animal studies have been traditionally performed almost exclusively in males. Similarly, human studies do not frequently compare THC effects on men and women. In any case, complementary analyses are guaranteed.
No human studies investigating THC effects on aversive memory reconsolidation were identified. The number of animal studies focusing on this question is still scarce, although their findings are promising. In a dose range from 0.3 to 10 mg/kg, the acute and systemic treatment with THC impaired the reconsolidation of specific and recent contextual fear memories in adult male rats, an effect dependent on prelimbic cortex CB1 receptor activation [124]. The acute treatment with 5.0 (but not 50) mg/kg of THC similarly impaired the reconsolidation of a recent cued fear memory in male rats [125]. Future studies should address whether THC can impair the reconsolidation of more remote and generalized aversive memories in both male and female animals. Noteworthy, mixed results have been reported with other drugs (e.g., propranolol) targeting aversive memory reconsolidation in rodents versus healthy humans [30, 33, 126,127,128,129,130], which suggests the existence of some boundary conditions, such as the strength, age, and specificity of the memory to be pharmacologically “fine-tuned”. Fortunately, animal findings indicate that the use of behavioral and pharmacological strategies can surmount these constraining factors [29, 131].
THC has been pointed out as a therapeutic cannabinoid for several brain diseases [132]. However, it is also the main responsible for the recreational use of Cannabis and its psychotomimetic effect, restricting Cannabis therapeutic use. Besides, some THC effects are commonly biphasic and dose-dependent [133]: low doses have potential therapeutic value in cognitive and anxiety disorders while high doses cause harmful effects, and are related to the reported side effects resulting from direct activation of CB1 receptors [51, 131]. This supraphysiological action can lead to a rapid down-regulation of these receptors [134, 135], potentially resulting in tolerance and addiction [136, 137]. CBD, on the other hand, has a safer profile because it is not a psychotic-precipitating substance, and its effects are mediated via indirect CB1 receptor activation and non-cannabinoid mechanisms as well [138, 139].
No human studies have yet investigated the effects of combining THC with CBD on aversive/traumatic memory extinction or reconsolidation. As early mentioned, however, this association could be advantageous. Accordingly, administering both THC and CBD in a sub-effective dose impaired either contextual or cued fear memory reconsolidation in adult male rats [124, 125]. In both cases, the THC:CBD dose ratio was 1:10, and combining THC with CBD allowed the use of 3-fold lower doses of them in the study by Stern et al. [124].
Regarding the effects of THC, dronabinol or nabilone on anxiety in subjects with no history of Cannabis use, orally taken doses ≥10 mg increased the anxiety level (and often produced a psychotic effect) while lower doses produced either no changes or anxiety reduction. Although the anxiogenic and psychotic effects of THC could have clinic impact (their effect sizes are as large as the ones calculated here for extinction facilitation and anxiety reduction), the dose range in which the beneficial and detrimental THC effects predominates is not necessarily the same. Therefore, its therapeutic value depends on the dose. Another aspect to keep in mind is the treatment regimen. Repeated administration of 1.0 or 5.0 mg of THC reduced anxious symptoms in anxious or PTSD patients, but acute treatment produced minor or no effects. In Cannabis users, the effects on anxiety are more variable because of the influence of the previous emotional state of individuals, how long have they used Cannabis, and when in life they started its use. Of note, inhalation is an alternative route to deliver THC. THC bioavailability after oral administration is around 10% [140], and after smoking, 25% of it reaches systemic circulation [141]. Based on that, a given smoked dose of THC will produce higher blood concentration than orally and, thus, the anxiogenic effect would be more frequent. However, in both studies in which this question was addressed indirectly, the subjects were previous Cannabis users, possibly having some degree of tolerance to the anxiogenic effect of THC. Besides, chronic users of Cannabis present altered anxiety levels relative to non-users [142]. Thus, the downregulation of CB1 receptors and differences in anxiety’s baseline could also have influenced THC effects in Cannabis users.
The biphasic effects of THC on anxiety have also been shown in laboratory animals. Low doses of THC (e.g., 0.3 mg/kg) did not alter the anxiety response in rodents tested in the elevated plus-maze, but doses between 1.0 and 10 mg/kg produced an anxiogenic-like effect [143]. In another study, THC doses ranging from 0.075 to 1.5 mg/kg produced an anxiolytic-like effect [144]. Interestingly, the anxiolytic and anxiogenic effects of THC can arise from different brain areas: infusing low doses into the ventral hippocampus or medial prefrontal cortex induced an anxiolytic-like response while higher doses had no effect or even produced an anxiogenic-like effect. In contrast, the same low THC dose infused into the basolateral amygdala increased the anxiety response, but higher doses were ineffective. Both effects relied on the activation of CB1 receptors [145], which are located on glutamatergic and GABAergic neurons. It is hypothesized that THC reduces glutamate levels in low doses and raises glutamate levels in high doses (the latter action is associated with inhibition of GABA-releasing neurons [146];). Furthermore, Bedse et al. [147] demonstrated in mice that the 2-AG depletion-induced anxiety-like behavior after stress exposure was counteracted by the administration of 0.25 mg/kg THC. Overall, these findings confirm the bidirectional effects of THC on anxiety, despite the varying dose range associated with anxiolytic and anxiogenic effects [148]. Of note, at doses ranging from 3.0 to 10 mg/kg in rats, and from 10 to 20 mg/kg in mice, THC also produced sedative-like effects [143]. The neural basis underlying THC effects on anxiety and sedation are under investigation.
Associating CBD with THC could be advantageous as CBD can minimize or even counteract some adverse effects of THC, such as the anxiogenic and psychotic effects. Oral administration of THC plus CBD in a dose ratio 1:2 attenuated the THC-associated anxiogenic effect. When using a similar THC:CBD ratio, the THC-associated anxiogenic effect was no longer observed with the use of low, but not high, doses of these drugs, a result that also depended on the previous Cannabis use history of participants. In humans, oral pretreatment with CBD 600 mg attenuated the psychotic symptoms induced by THC 10 mg [80]. Similarly, 400 mg, but not 4.0 mg, of CBD vaporized attenuated the intoxication produced by THC 8.0 mg [149]. Preclinical studies with animals also demonstrate that co-administration of THC:CBD in 1:5 and 1:10, but not 1:1, dose ratio can counteract the THC-induced anxiogenic-like effects and impairments in social interaction [150, 151]. Based on the above, the combined strategy seems to be as complex as using THC alone.
There is evidence suggesting that CBD interferes with the pharmacokinetics of THC. Indeed, CBD can inhibit the hepatic metabolism of THC [152, 153]. As a result, co-administering an equal dose of CBD doubled the brain THC amount, 30 min later, in adolescent male rats and, thus, this association produced an anxiogenic-like effect [154]. Similarly, female mice receiving a THC:CBD dose ratio of 1:2 presented an anxiogenic-like effect [155]. However, studies using proportionally more CBD than THC and performed in adult rodents and monkeys found a reduction in THC anxiogenic action [156,157,158], indicating that the hepatic biotransformation of THC may vary according to the animal’s age. In men, independent of dose ratio, the oral co-administration of CBD with THC did not alter the plasmatic THC concentration [80, 96, 159]; in women, the intake of a THC:CBD dose ratio of 1:0.5 induced a tendency of increasing both THC and its active metabolite (11-OH-THC) in the plasma, suggesting potential sex-dependent differences in THC and CBD metabolism [159]. Altogether, CBD can interfere with the pharmacokinetics of THC, but the dose, animal species, sex, and proportion of these drugs influence if CBD will potentiate or antagonize THC effects.
Conclusions
THC, dronabinol or nabilone could help with hyperarousal symptoms, insomnia, anxiety, and extinction deficits related to PTSD [51]. Indeed, despite the limited number of published studies, available data suggest that low doses of THC potentiate fear memory extinction in healthy volunteers and reduce anxiety responses in anxious and PTSD patients without inducing a psychotic effect. High doses of THC, however, do not facilitate fear memory extinction and are related to clinically relevant anxiogenic and psychotic effects in healthy volunteers. Overall, laboratory animal data corroborate human findings.
There is a lack of studies with PTSD patients using THC alone and associated with CBD focusing on aversive memory extinction and reconsolidation. Further, most studies evaluated the acute effects of THC or THC plus CBD. Therefore, it is unknown whether chronic treatment is still advantageous. Besides, some studies do not address potential sex-dependent differences in THC-induced effects, which would provide further information on whether or not it is a potential issue in humans. Animal data have shown the detrimental effects of THC following high doses. Based on that, human studies have selected an appropriate dose range of THC and, thus, neither worsening of PTSD symptoms nor strengthening of aversive memories after the use of THC has been reported. Few studies have investigated the effects of associating THC with CBD in varying dose ratios yet. Altogether, the findings encourage future controlled studies evaluating the effects of low doses of THC to attenuate aversive/traumatic memory expression in PTSD patients.
Availability of data and materials
All data generated or analyzed during this study are included in this published article [and its supplementary information files].
Abbreviations
- 2-AG:
-
2-Arachidonoylglycerol
- Caps:
-
Clinician administered posttraumatic scale
- CB1:
-
Cannabinoid type 1 receptor
- CBD:
-
Cannabidiol
- dACC:
-
Dorsal anterior cingulate cortex
- FAAH:
-
Fatty acid amide hydrolase
- mPFC:
-
Medial prefrontal cortex
- OEA:
-
Oleoylethanolamide
- PANSS:
-
Positive and negative psychotic syndrome scale
- PEA:
-
Palmitoylethanolamide
- POMS:
-
Profile of mood state
- PTSD:
-
Posttraumatic stress disorder
- SEA:
-
Stearoylethanolamide
- SCR:
-
Skin conductance response
- SSRIs:
-
Selective serotonin reuptake inhibitors
- STAI:
-
State trait anxiety inventory
- THC:
-
Δ9-tetrahydrocannabinol
- VAS:
-
Visual analog scale
- vmPFC:
-
Ventromedial prefrontal cortex
References
Brewin CR. Episodic memory, perceptual memory, and their interaction: foundations for a theory of posttraumatic stress disorder. Psychol Bull 2014 Jan;140(1):69–97. https://doi.org/https://doi.org/10.1037/a0033722.
Reul JM, Nutt DJ. Glutamate and cortisol—a critical confluence in PTSD? J Psychopharmacol 2008 Jul;22(5):469–472. https://doi.org/https://doi.org/10.1177/0269881108094617.
American Psychiatry Association (APA). Diagnostic and Statistical Manual of Mental Disorders. 5th ed. Washington, DC, USA: American Psychiatric Association; 2013.
Brewin CR. Memory and forgetting. Curr Psychiatry Rep 2018 Aug;20(10):87. https://doi.org/https://doi.org/10.1007/s11920-018-0950-7.
Ehlers A, Hackmann A, Michael T. Intrusive re-experiencing in post-traumatic stress disorder: phenomenology, theory, and therapy. Memory. 2004 Jul;12(4):403–415. https://doi.org/https://doi.org/10.1080/09658210444000025.
Yehuda R, Hoge CW, McFarlane AC, Vermetten E, Lanius RA, Nievergelt CM, et al. Post-traumatic stress disorder. Nat Rev Dis Primers. 2015;1:15057. https://doi.org/https://doi.org/10.1038/nrdp.2015.57.
Hofmann SG. Enhancing exposure-based therapy from a translational research perspective. Behav Res Ther 2007;45(9):1987–2001. https://doi.org/https://doi.org/10.1016/j.brat.2007.06.006.
Stojek MM, McSweeney LB, Rauch SAM. Neuroscience informed prolonged exposure practice: increasing efficiency and efficacy through mechanisms. Front Behav Neurosci 2018;12:281. https://doi.org/https://doi.org/10.3389/fnbeh.2018.00281.
Do-Monte FH, Manzano-Nieves G, Quiñones-Laracuente K, Ramos-Medina L, Quirk GJ. Revisiting the role of Infralimbic cortex in fear extinction with Optogenetics. J Neurosci 2015;35(8):3607–3615. https://doi.org/https://doi.org/10.1523/JNEUROSCI.3137-14.2015.
Careaga MBL, Girardi CEN, Suchecki D. Understanding posttraumatic stress disorder through fear conditioning, extinction and reconsolidation. Neurosci Biobehav Rev 2016;71:48–57. https://doi.org/https://doi.org/10.1016/j.neubiorev.2016.08.023.
Marek R, Sun Y, Sah P. Neural circuits for a top-down control of fear and extinction. Psychopharmacology 2019;236(1):313–320. https://doi.org/https://doi.org/10.1007/s00213-018-5033-2.
Morrison FG, Ressler KG. From the neurobiology of extinction to improved clinical treatments. Depress Anxiety 2014;31(4):279–290. https://doi.org/https://doi.org/10.1002/da.22214.
Horn SR, Charney DS, Feder A. Understanding resilience: new approaches for preventing and treating PTSD. Exp Neurol. 2016;284(Pt B):119–132. https://doi.org/https://doi.org/10.1016/j.expneurol.2016.07.002.
Hughes KC, Shin LM. Functional neuroimaging studies of post-traumatic stress disorder. Expert Rev Neurother 2011;11(2):275–285. https://doi.org/https://doi.org/10.1586/ern.10.198.
Markowitz S, Fanselow M. Exposure therapy for post-traumatic stress disorder: factors of limited success and possible alternative treatment. Brain Sci. 2020;10(3). pii: E167. https://doi.org/https://doi.org/10.3390/brainsci10030167.
Diamond DM, Zoladz PR. Dysfunctional or hyperfunctional? The amygdala in posttraumatic stress disorder is the bull in the evolutionary China shop. J Neurosci Res 2016;94(6):437–444. https://doi.org/https://doi.org/10.1002/jnr.23684.
Rauch SL, Shin LM, Phelps EA. Neurocircuitry models of posttraumatic stress disorder and extinction: human neuroimaging research--past, present, and future. Biol Psychiatry 2006;60(4):376–382. https://doi.org/https://doi.org/10.1016/j.biopsych.2006.06.004.
Shin LM, Rauch SL, Pitman RK. Amygdala, medial prefrontal cortex, and hippocampal function in PTSD. Ann N Y Acad Sci 2006;1071(1):67–79. https://doi.org/https://doi.org/10.1196/annals.1364.007.
Joshi SA, Duval ER, Kubat B, Liberzon I. A review of hippocampal activation in post-traumatic stress disorder. Psychophysiology 2020;57(1):e13357. https://doi.org/https://doi.org/10.1111/psyp.13357.
Olff M, Polak AR, Witteveen AB, Denys D. Executive function in posttraumatic stress disorder (PTSD) and the influence of comorbid depression. Neurobiol Learn Mem 2014;112:114–121. https://doi.org/https://doi.org/10.1016/j.nlm.2014.01.003.
Bolsinger J, Seifritz E, Kleim B, Manoliu A. Neuroimaging correlates of resilience to traumatic events-a comprehensive review. Front Psychiatry. 2018;9:693 https://doi.org/0.3389/fpsyt.2018.00693.
Fitzgerald JM, DiGangi JA, Phan KL. Functional neuroanatomy of emotion and its regulation in PTSD. Harv Rev Psychiatry. 2018;26(3):116–128. https://doi.org/https://doi.org/10.1097/HRP.0000000000000185.
Quirk GJ, Russo GK, Barron JL, Lebron K. The role of ventromedial prefrontal cortex in the recovery of extinguished fear. J Neurosci 2000;20(16):6225–6231. https://doi.org/https://doi.org/10.1523/JNEUROSCI.20-16-06225.2000.
Kida S. Function and mechanisms of memory destabilization and reconsolidation after retrieval. Proc Jpn Acad Ser B Phys Biol Sci 2020;96(3):95–106. https://doi.org/https://doi.org/10.2183/pjab.96.008.
Stern CA, Gazarini L, Vanvossen AC, Hames MS, Bertoglio LJ. Activity in prelimbic cortex subserves fear memory reconsolidation over time. Learn Mem. 2013;21(1):14–20 https://doi.org/10.1101/lm.032631.113.
da Silva TR, Raymundi AM, Bertoglio LJ, Andreatini R, Stern CA. Role of prelimbic cortex PKC and PKMζ in fear memory reconsolidation and persistence following reactivation. Sci Rep 2020;10(1):4076. https://doi.org/https://doi.org/10.1038/s41598-020-60046-x.
Vanvossen AC, Portes MA, Scoz-Silva R, Reichmann HB, Stern CA, Bertoglio, LJ. Newly acquired and reactivated contextual fear memories are more intense and prone to generalize after activation of prelimbic cortex NMDA receptors. Neurobiol Learn Mem 2017;137:154–162. https://doi.org/https://doi.org/10.1016/j.nlm.2016.12.002.
Nader K, Schafe GE, LeDoux JE. The labile nature of consolidation theory. Nat Rev Neurosci 2000;1(3):216–219. https://doi.org/https://doi.org/10.1038/35044580.
Gazarini L, Stern CA, Piornedo RR, Takahashi RN, Bertoglio LJ. PTSD-like memory generated through enhanced noradrenergic activity is mitigated by a dual step pharmacological intervention targeting its reconsolidation. Int J Neuropsychopharmacol. 2014;18(1). pii: pyu026.https://doi.org/https://doi.org/10.1093/ijnp/pyu026.
Elsey JW, Kindt M. Tackling maladaptive memories through reconsolidation: From neural to clinical science. Neurobiol Learn Mem. 2017;142(Pt A):108–117. https://doi.org/https://doi.org/10.1016/j.nlm.2017.03.007.
Monfils MH, Holmes EA. Memory boundaries: opening a window inspired by reconsolidation to treat anxiety, trauma-related, and addiction disorders. Lancet Psychiatry 2018;5(12):1032–1042. https://doi.org/https://doi.org/10.1016/S2215-0366(18)30270-0.
Thierrée S, Richa S, Brunet A, Egreteau L, Roig Q, Clarys D, El-Hage W. Trauma reactivation under propranolol among traumatized Syrian refugee children: preliminary evidence regarding efficacy. Eur J Psychotraumatol 2020;11(1):1733248. https://doi.org/https://doi.org/10.1080/20008198.2020.1733248.
Kindt M, Soeter M, Vervliet B. Beyond extinction: erasing human fear responses and preventing the return of fear. Nat Neurosci 2009;12(3):256–258. https://doi.org/https://doi.org/10.1038/nn.2271.
Barak S, Ben Hamida S. Memory erasure, enhanced extinction and disrupted reconsolidation. J Neurosci 2012;32(7):2250–2251. https://doi.org/https://doi.org/10.1523/JNEUROSCI.6123-11.2012.
Haubrich J, Bernabo M, Baker AG, Nader K. Impairments to consolidation, reconsolidation, and long-term memory maintenance lead to memory erasure. Annu Rev Neurosci. 2020;43:297–314. https://doi.org/annurev-neuro-091319-024636.
Lonsdorf TB, Menz MM, Andreatta M, Fullana MA, Golkar A, Haaker J, et al. Don't fear 'fear conditioning': methodological considerations for the design and analysis of studies on human fear acquisition, extinction, and return of fear. Neurosci Biobehav Rev 2017;77:247–285. https://doi.org/https://doi.org/10.1016/j.neubiorev.2017.02.026.
Flandreau EI, Toth M. Animal models of PTSD: a critical review. Curr Top Behav Neurosci 2018;38:47–68. https://doi.org/https://doi.org/10.1007/7854_2016_65.
Carobrez AP, Bertoglio LJ. Ethological and temporal analyses of anxiety-like behavior: the elevated plus-maze model 20 years on. Neurosci Biobehav Rev 2005;29(8):1193–1205. https://doi.org/https://doi.org/10.1016/j.neubiorev.2005.04.017.
Grillon C, Robinson OJ, Cornwell B, Ernst M. Modeling anxiety in healthy humans: a key intermediate bridge between basic and clinical sciences. Neuropsychopharmacology. 2019;44(12):1999–2010. https://doi.org/https://doi.org/10.1038/s41386-019-0445-1.
Neumeister A, Normandin MD, Pietrzak RH, Piomelli D, Zheng MQ, Gujarro-Anton A, et al. Elevated brain cannabinoid CB1 receptor availability in post-traumatic stress disorder: a positron emission tomography study. Mol Psychiatry 2013;18(9):1034–1040. https://doi.org/https://doi.org/10.1038/mp.2013.61.
Hill MN, Bierer LM, Makotkine I, Golier JA, Galea S, McEwen BS, et al. Reductions in circulating endocannabinoid levels in individuals with post-traumatic stress disorder following exposure to the world trade center attacks. Psychoneuroendocrinology 2013;38(12):2952–2961. https://doi.org/https://doi.org/10.1016/j.psyneuen.2013.08.004.
Wilker S, Pfeiffer A, Elbert T, Ovuga E, Karabatsiakis A, Krumbholz A, et al. Endocannabinoid concentrations in hair are associated with PTSD symptom severity. Psychoneuroendocrinology 2016;67:198–206. https://doi.org/https://doi.org/10.1016/j.psyneuen.2016.02.010.
Lazary J, Eszlari N, Juhasz G, Bagdy G. Genetically reduced FAAH activity may be a risk for the development of anxiety and depression in persons with repetitive childhood trauma. Eur Neuropsychopharmacol 2016;26(6):1020–1028. https://doi.org/https://doi.org/10.1016/j.euroneuro.2016.03.003.
Gee DG, Fetcho RN, Jing D, Li A, Glatt CE, Drysdale AT, et al. Individual differences in frontolimbic circuitry and anxiety emerge with adolescent changes in endocannabinoid signaling across species. Proc Natl Acad Sci U S A 2016;113(16):4500–4505. https://doi.org/https://doi.org/10.1073/pnas.1600013113.
Chiang KP, Gerber AL, Sipe JC, Cravatt BF. Reduced cellular expression and activity of the P129T mutant of human fatty acid amide hydrolase: evidence for a link between defects in the endocannabinoid system and problem drug use. Hum Mol Genet 2004;13(18):2113–2119. https://doi.org/https://doi.org/10.1093/hmg/ddh216.
Dincheva I, Drysdale AT, Hartley CA, Johnson DC, Jing D, King EC, et al. FAAH genetic variation enhances fronto-amygdala function in mouse and human. Nat Commun 2015;6(1):6395. https://doi.org/https://doi.org/10.1038/ncoms7395.
Hariri AR, Gorka A, Hyde LW, Kimak M, Halder I, Ducci F, et al. Divergent effects of genetic variation in endocannabinoid signaling on human threat- and reward-related brain function. Biol Psychiatry 2009;66(1):9–16. https://doi.org/https://doi.org/10.1016/j.biopsych.2008.10.047.
Stein DJ, Ipser JC, Seedat S, Sager C, Amos T. Pharmacotherapy for post traumatic stress disorder (PTSD). Cochrane Database Syst Rev 2006;(1):CD002795. https://doi.org/https://doi.org/10.1002/14651858.CD002795.pub2.
Friedman MJ, Marmar CR, Baker DG, Sikes CR, Farfel GM. Randomized, double-blind comparison of sertraline and placebo for posttraumatic stress disorder in a Department of Veterans Affairs setting. J Clin Psychiatry 2007;68(5):711–720. https://doi.org/https://doi.org/10.4088/JCP.v68n0508.
Berger W, Mendlowicz MV, Marques-Portella C, Kinrys G, Fontenelle LF, Marmar CR, et al. Pharmacologic alternatives to antidepressants in posttraumatic stress disorder: a systematic review. Prog Neuro-Psychopharmacol Biol Psychiatry 2009;33(2):169–180. https://doi.org/https://doi.org/10.1016/j.pnpbp.2008.12.004.
Hill MN, Campolongo P, Yehuda R, Patel S. Integrating endocannabinoid signaling and cannabinoids into the biology and treatment of posttraumatic stress disorder. Neuropsychopharmacology 2018;43(1):80–102. https://doi.org/https://doi.org/10.1038/npp.2017.162.
Betthauser K, Pilz J, Vollmer LE. Use and effects of cannabinoids in military veterans with posttraumatic stress disorder. Am J Health Syst Pharm 2015;72(15):1279–1284 https://doi.org/https://doi.org/10.2146/ajhp140523.
Bonn-Miller MO, Vujanovic AA, Feldner MT, Bernstein A, Zvolensky MJ. Posttraumatic stress symptom severity predicts marijuana use coping motives among traumatic event-exposed marijuana users. J Trauma Stress 2007;20(4):577–586. https://doi.org/https://doi.org/10.1002/jts.20243.
Bremner JD, Southwick SM, Darnell A, Charney DS. Chronic PTSD in Vietnam combat veterans: course of illness and substance abuse. Am J Psychiatry 1996;153(3):369–375. https://doi.org/https://doi.org/10.1176/ajp.153.3.369.
Greer GR, Grob CS, Halberstadt AL. PTSD symptom reports of patients evaluated for the New Mexico medical Cannabis program. J Psychoactive Drugs 2014;46(1):73–77. https://doi.org/https://doi.org/10.1080/02791072.2013.873843.
Jetly R, Heber A, Fraser G, Boisvert D. The efficacy of nabilone, a synthetic cannabinoid, in the treatment of PTSD-associated nightmares: a preliminary randomized, double-blind, placebo-controlled cross-over design study. Psychoneuroendocrinology 2015;51:585–588. https://doi.org/https://doi.org/10.1016/j.psyneuen.2014.11.002.
Cameron C, Watson D, Robinson J. Use of a synthetic cannabinoid in a correctional population for posttraumatic stress disorder-related insomnia and nightmares, chronic pain, harm reduction, and other indications: a retrospective evaluation. J Clin Psychopharmacol 2014;34(5):559–564. https://doi.org/https://doi.org/10.1097/JCP.0000000000000180.
Shishko I, Oliveira R, Moore TA, Almeida K. A review of medical marijuana for the treatment of posttraumatic stress disorder: real symptom re-leaf or just high hopes? Ment Health Clin 2018;8(2):86–94. https://doi.org/https://doi.org/10.9740/mhc.2018.03.086.
Abizaid A, Merali Z, Anisman H. Cannabis: a potential efficacious intervention for PTSD or simply snake oil? J Psychiatry Neurosci 2019;44(2):75–78. https://doi.org/https://doi.org/10.1503/jpn.190021.
Bonn-Miller MO, Boden MT, Bucossi MM, Babson KA. Self-reported cannabis use characteristics, patterns and helpfulness among medical cannabis users. Am J Drug Alcohol Abuse 2014;40(1):23–30. https://doi.org/https://doi.org/10.3109/00952990.2013.821477.
Wilkinson ST, Stefanovics E, Rosenheck RA. Marijuana use is associated with worse outcomes in symptom severity and violent behavior in patients with posttraumatic stress disorder. J Clin Psychiatry 2015;76(9):1174–1180 https://doi.org/https://doi.org/10.4088/JCP.14m09475.
Black N, Stockings E, Campbell G, Tran LT, Zagic D, Hall WD, et al. Cannabinoids for the treatment of mental disorders and symptoms of mental disorders: a systematic review and meta-analysis. Lancet Psychiatry 2019;6(12):995–1010. https://doi.org/https://doi.org/10.1016/S2215-0366(19)30401-8.
Stern CA, Gazarini L, Takahashi RN, Guimarães FS, Bertoglio LJ. On disruption of fear memory by reconsolidation blockade: evidence from cannabidiol treatment. Neuropsychopharmacology 2012;37(9):2132–2142. https://doi.org/https://doi.org/10.1038/npp.2012.63.
Bitencourt RM, Pamplona FA, Takahashi RN. Facilitation of contextual fear memory extinction and anti-anxiogenic effects of AM404 and cannabidiol in conditioned rats. Eur Neuropsychopharmacol 2008;18(12):849–859. https://doi.org/https://doi.org/10.1016/j.euroneuro.2008.07.001.
Zuardi AW, Cosme RA, Graeff FG, Guimarães FS. Effects of ipsapirone and cannabidiol on human experimental anxiety. J Psychopharmacol 1993;7(1 Suppl):82–88. https://doi.org/https://doi.org/10.1177/026988119300700112.
Crippa JA, Zuardi AW, Garrido GE, Wichert-Ana L, Guarnieri R, Ferrari L, et al. Effects of cannabidiol (CBD) on regional cerebral blood flow. Neuropsychopharmacology 2004;29(2):417–426. https://doi.org/https://doi.org/10.1038/sj.npp.1300340.
Crippa JA, Derenusson GN, Ferrari TB, Wichert-Ana L, Duran FL, Martin-Santos R, et al. Neural basis of anxiolytic effects of cannabidiol (CBD) in generalized social anxiety disorder: a preliminary report. J Psychopharmacol 2011;25(1):121–130. https://doi.org/https://doi.org/10.1177/0269881110379283.
Zuardi AW, Rodrigues NP, Silva AL, Bernardo SA, Hallak JEC, Guimarães FS, et al. Inverted U-shaped dose-response curve of the anxiolytic effect of Cannabidiol during public speaking in real life. Front Pharmacol 2017;8:259.https://doi.org/https://doi.org/10.3389/fphar.2017.00259.
Ellis PD. The essential guide to effect sizes: statistical power, meta-analysis, and the interpretation of research results. Cambridge: Cambridge University Press; 2010. https://doi.org/10.1017/CBO9780511761676.
Rabinak CA, Angstadt M, Sripada CS, Abelson JL, Liberzon I, Milad MR, et al. Cannabinoid facilitation of fear extinction memory recall in humans. Neuropharmacology 2013;64(1):396–402. https://doi.org/https://doi.org/10.1016/j.neuropharm.2012.06.063.
Rabinak CA, Angstadt M, Lyons M, Mori S, Milad MR, Liberzon I, et al. Cannabinoid modulation of prefrontal-limbic activation during fear extinction learning and recall in humans. Neurobiol Learn Mem 2014;113:125–134. https://doi.org/https://doi.org/10.1016/j.nlm.2013.09.009.
Rabinak CA, Peters C, Marusak HA, Ghosh S, Phan KL. Effects of acute Δ9-tetrahydrocannabinol on next-day extinction recall is mediated by post-extinction resting-state brain dynamics. Neuropharmacology 2018;143:289–298. https://doi.org/https://doi.org/10.1016/j.neuropharm.2018.10.002.
Hammoud MZ, Peters C, Hatfield JRB, Gorka SM, Phan KL, Milad MR, et al. Influence of Δ9-tetrahydrocannabinol on long-term neural correlates of threat extinction memory retention in humans. Neuropsychopharmacology 2019. https://doi.org/https://doi.org/10.1038/s41386-019-0416-6.
Klumpers F, Denys D, Kenemans JL, Grillon C, van der Aart J, Baas JM. Testing the effects of Δ9-THC and D-cycloserine on extinction of conditioned fear in humans. J Psychopharmacol 2012;26(4):471–478. https://doi.org/https://doi.org/10.1177/0269881111431624.
Santana F, Sierra RO, Haubrich J, Crestani AP, Duran JM, de Freitas Cassini L, et al. Involvement of the infralimbic cortex and CA1 hippocampal area in reconsolidation of a contextual fear memory through CB1 receptors: effects of CP55,940. Neurobiol Learn Mem 2016;127:42–47. https://doi.org/https://doi.org/10.1016/j.nlm.2015.11.016.
Lee JL, Amorim FE, Cassini LF, Amaral OB. Different temporal windows for CB1 receptor involvement in contextual fear memory destabilisation in the amygdala and hippocampus. PLoS One 2019;14(1):e0205781. https://doi.org/https://doi.org/10.1371/journal.pone.0205781.
Russo E, Guy GW. A tale of two cannabinoids: the therapeutic rationale for combining tetrahydrocannabinol and cannabidiol. Med Hypotheses 2006;66(2):234–246. https://doi.org/https://doi.org/10.1016/j.mehy.2005.08.026.
MacCallum CA, Russo EB. Practical considerations in medical cannabis administration and dosing. Eur J Intern Med 2018;49:12–19. https://doi.org/https://doi.org/10.1016/j.ejim.2018.01.004.
Martin-Santos R, Crippa JA, Batalla A, Bhattacharyya S, Atakan Z, Borgwardt S, et al. Acute effects of a single, oral dose of d9-tetrahydrocannabinol (THC) and cannabidiol (CBD) administration in healthy volunteers. Curr Pharm Des 2012;18(32):4966–4979. https://doi.org/https://doi.org/10.2174/138161212802884780.
Bhattacharyya S, Morrison PD, Fusar-Poli P, Martin-Santos R, Borgwardt S, Winton-Brown T, et al. Opposite effects of delta-9-tetrahydrocannabinol and cannabidiol on human brain function and psychopathology. Neuropsychopharmacology 2010;35(3):764–774. https://doi.org/https://doi.org/10.1038/npp.2009.184.
Bhattacharyya S, Egerton A, Kim E, Rosso L, Riano Barros D, Hammers A, et al. Acute induction of anxiety in humans by delta-9-tetrahydrocannabinol related to amygdalar cannabinoid-1 (CB1) receptors. Sci Rep 2017;7(1):15025. https://doi.org/https://doi.org/10.1038/s41598-017-14203-4.
Colizzi M, McGuire P, Giampietro V, Williams S, Brammer M, Bhattacharyya S. Previous cannabis exposure modulates the acute effects of delta-9-tetrahydrocannabinol on attentional salience and fear processing. Exp Clin Psychopharmacol 2018;26(6):582–598. https://doi.org/https://doi.org/10.1037/pha0000221.
Fusar-Poli P, Crippa JA, Bhattacharyya S, Borgwardt SJ, Allen P, Martin-Santos R, et al. Distinct effects of {delta}9-tetrahydrocannabinol and cannabidiol on neural activation during emotional processing. Arch Gen Psychiatry 2009;66(1):95–105. https://doi.org/https://doi.org/10.1001/archgenpsychiatry.2008.519.
Notcutt W, Langford R, Davies P, Ratcliffe S, Potts R. A placebo-controlled, parallel-group, randomized withdrawal study of subjects with symptoms of spasticity due to multiple sclerosis who are receiving long-term Sativex® (nabiximols). Mult Scler 2012;18(2):219–228. https://doi.org/https://doi.org/10.1177/1352458511419700.
Thomas A, Baillie GL, Phillips AM, Razdan RK, Ross RA, Pertwee RG. Cannabidiol displays unexpectedly high potency as an antagonist of CB1 and CB2 receptor agonists in vitro. Br J Pharmacol 2007;150(5):613–623. https://doi.org/https://doi.org/10.1038/sj.bjp.0707133.
Pertwee RG. Emerging strategies for exploiting cannabinoid receptor agonists as medicines. Br J Pharmacol 2009;156(3):397–411. https://doi.org/https://doi.org/10.1111/j.1476-5381.2008.00048.x.
Boggs DL, Nguyen JD, Morgenson D, Taffe MA, Ranganathan M. Clinical and preclinical evidence for functional interactions of Cannabidiol and Δ 9-tetrahydrocannabinol. Neuropsychopharmacology 2018;43(1):142–154. https://doi.org/https://doi.org/10.1038/npp.2017.209.
Freeman AM, Petrilli K, Lees R, Hindocha C, Mokrysz C, Curran HV, Saunders R, Freeman TP. How does Cannabidiol (CBD) influence the acute effects of delta-9-tetrahydrocannabinol (THC) in humans? a systematic review. Neurosci Biobehav Rev. 2019;107:696–712. https://doi.org/. https://doi.org/10.1016/j.neubiorev.2019.09.036.
Nakano S, Gillespie HK, Hollister LE. A model for evaluation of antianxiety drugs with the use of experimentally induced stress: comparison of nabilone and diazepam. Clin Pharmacol Ther 1978;23(1):54–62. https://doi.org/https://doi.org/10.1002/cpt197823154.
Phan KL, Angstadt M, Golden J, Onyewuenyi I, Popovska A, de Wit H. Cannabinoid modulation of amygdala reactivity to social signals of threat in humans. J Neurosci 2008;28(10):2313–2319. https://doi.org/https://doi.org/10.1523/JNEUROSCI.5603-07.2008.
Childs E, Lutz JA, de Wit H. Dose-related effects of delta-9-THC on emotional responses to acute psychosocial stress. Drug Alcohol Depend 2017 Aug;177:136–144. https://doi.org/https://doi.org/10.1016/j.drugalcdep.2017.03.030.
Zuardi AW, Shirakawa I, Finkelfarb E, Karniol IG. Action of cannabidiol on the anxiety and other effects produced by delta 9-THC in normal subjects. Psychopharmacology 1982;76(3):245–250. https://doi.org/https://doi.org/10.1007/BF00432554.
Karniol IG, Shirakawa I, Kasinski N, Pfeferman A, Carlini EA. Cannabidiol interferes with the effects of delta 9 - tetrahydrocannabinol in man. Eur J Pharmacol 1974;28(1):172–177. https://doi.org/https://doi.org/10.1016/0014-2999(74)90129-0.
Naliboff BD, Rickles WH, Cohen MJ, Naimark RS. Interactions of marijuana and induced stress: forearm blood flow, heart rate, and skin conductance. Psychophysiology. 1976;13(6):517–22.
Ilan AB, Gevins A, Coleman M, ElSohly MA, de Wit H. Neurophysiological and subjective profile of marijuana with varying concentrations of cannabinoids. Behav Pharmacol 2005;16(5–6):487–496. https://doi.org/https://doi.org/10.1097/00008877-200509000-00023.
Karschner EL, Darwin WD, McMahon RP, Liu F, Wright S, Goodwin RS, et al. Subjective and physiological effects after controlled Sativex and oral THC administration. Clin Pharmacol Ther 2011;89(3):400–407. https://doi.org/https://doi.org/10.1038/clpt.2010.318.
Glass RM, Uhlenhuth EH, Hartel FW, Schuster CR, Fischman MW. A single dose study of nabilone, a synthetic cannabinoid. Psychopharmacology 1980;71(2):137–142. https://doi.org/https://doi.org/10.1007/BF00434401.
Fabre LF, McLendon D. The efficacy and safety of nabilone (a synthetic cannabinoid) in the treatment of anxiety. J Clin Pharmacol 1981;21 S1:377S–382S. https://doi.org/https://doi.org/10.1002/j.1552-4604.1981.tb02617.x.
Roitman P, Mechoulam R, Cooper-Kazaz R, Shalev A. Preliminary, open-label, pilot study of add-on oral Δ9-tetrahydrocannabinol in chronic post-traumatic stress disorder. Clin Drug Investig 2014;34(8):587–591. https://doi.org/https://doi.org/10.1007/s40261-014-0212-3.
Rabinak CA, Blanchette A, Zabik NL, Peters C, Marusak HA, Iadipaolo A, et al. Cannabinoid modulation of corticolimbic activation to threat in trauma-exposed adults: a preliminary study. Psychopharmacology 2020. https://doi.org/https://doi.org/10.1007/s00213-020-05499-8.
Marsicano G, Wotjak CT, Azad SC, Bisogno T, Rammes G, Cascio MG, et al. The endogenous cannabinoid system controls extinction of aversive memories. Nature 2002;418(6897):530–534. https://doi.org/https://doi.org/10.1038/nature00839.
Quirk GJ, Mueller D. Neural mechanisms of extinction learning and retrieval. Neuropsychopharmacology 2008;33(1):56–72. https://doi.org/https://doi.org/10.1038/sj.npp.1301555.
Fitzgerald PJ, Giustino TF, Seemann JR, Maren S. Noradrenergic blockade stabilizes prefrontal activity and enables fear extinction under stress. Proc Natl Acad Sci U S A 2015;112(28):E3729–E3737. https://doi.org/https://doi.org/10.1073/pnas.1500682112.
Papini S, Sullivan GM, Hien DA, Shvil E, Neria Y. Toward a translational approach to targeting the endocannabinoid system in posttraumatic stress disorder: a critical review of preclinical research. Biol Psychol 2015;104:8–18. https://doi.org/https://doi.org/10.1016/j.biopsycho.2014.10.010.
Chhatwal JP, Davis M, Maguschak KA, Ressler KJ. Enhancing cannabinoid neurotransmission augments the extinction of conditioned fear. Neuropsychopharmacology 2005;30(3):516–524. https://doi.org/https://doi.org/10.1038/sj.npp.1300655.
Gorka SM, Fitzgerald DA, de Wit H, Phan KL. Cannabinoid modulation of amygdala subregion functional connectivity to social signals of threat. Int J Neuropsychopharmacol 2014;18(3):1–6. https://doi.org/https://doi.org/10.1093/ijnp/pyu104.
Ballard ME, Bedi G, de Wit H. Effects of delta-9-tetrahydrocannabinol on evaluation of emotional images. J Psychopharmacol 2012;26(10):1289–1298. https://doi.org/https://doi.org/10.1177/0269881112446530.
Ballard ME, Gallo DA, de Wit H. Pre-encoding administration of amphetamine or THC preferentially modulates emotional memory in humans. Psychopharmacology 2013;226(3):515–529. https://doi.org/https://doi.org/10.1007/s00213-012-2924-5.
Hindocha C, Freeman TP, Schafer G, Gardener C, Das RK, Morgan CJ, et al. Acute effects of delta-9-tetrahydrocannabinol, cannabidiol and their combination on facial emotion recognition: a randomised, double-blind, placebo-controlled study in cannabis users. Eur Neuropsychopharmacol 2015;25(3):325–334. https://doi.org/10.1016/j.euroneuro.2014.11.014.
Doss MK, Weafer J, Gallo DA, de Wit H. Δ9-tetrahydrocannabinol at retrieval drives false recollection of neutral and emotional memories. Biol Psychiatry 2018;84(10):743–750. https://doi.org/10.10e16/j.biopsych.2018.04.020.
Rabinak C, Peters C, Elrahal F, Milad M, Rauch S, Phan KL, et al. Cannabinoid facilitation of fear extinction in posttraumatic stress disorder. Biol Psychiatry. 2018;83:S1eS107.
O'Neil ME, Nugent SM, Morasco BJ, Freeman M, Low A, Kondo K, Zakher B, Elven C, Motu'apuaka M, Paynter R, Kansagara D. Benefits and harms of plant-based Cannabis for posttraumatic stress disorder: a systematic review. Ann Intern Med 2017;167(5):332–340.https://doi.org/https://doi.org/10.7326/M17-0477.
Onaivi ES, Chaudhuri G, Abaci AS, Parker M, Manier DH, Martin PR, et al. Expression of cannabinoid receptors and their gene transcripts in human blood cells. Prog Neuro-Psychopharmacol Biol Psychiatry 1999;23(6):1063–1077. https://doi.org/https://doi.org/10.1016/S0278-5846(99)00052-4.
Busch L, Sterin-Borda L, Borda E. Effects of castration on cannabinoid cb receptor expression and on the biological actions of cannabinoid in the parotid gland. Clin Exp Pharmacol Physiol 2006;33(3):258–263. https://doi.org/https://doi.org/10.1111/j.1440-1681.2006.04355.x.
González S, Bisogno T, Wenger T, Manzanares J, Milone A, Berrendero F, et al. Sex steroid influence on cannabinoid CB(1) receptor mRNA and endocannabinoid levels in the anterior pituitary gland. Biochem Biophys Res Commun 2000;270(1):260–266. https://doi.org/https://doi.org/10.1006/bbrc.2000.2406.
Mize AL, Alper RH. Acute and long-term effects of 17beta-estradiol on G(i/o) coupled neurotransmitter receptor function in the female rat brain as assessed by agonist-stimulated [35S] GTPgammaS binding. Brain Res 2000;859(2):326–333. https://doi.org/https://doi.org/10.1016/S0006-8993(00)01998-3.
Bradshaw HB, Rimmerman N, Krey JF, Walker JM. Sex and hormonal cycle differences in rat brain levels of pain-related cannabimimetic lipid mediators. Am J Physiol Regul Integr Comp Physiol 2006;291(2):R349–R358. https://doi.org/https://doi.org/10.1152/ajpregu.00933.2005.
Rodríguez de Fonseca F, Cebeira M, Ramos JA, Martín M, Fernández-Ruiz JJ. Cannabinoid receptors in rat brain areas: sexual differences, fluctuations during estrous cycle and changes after gonadectomy and sex steroid replacement. Life Sci 1994;54(3):159–170. https://doi.org/https://doi.org/10.1016/0024-3205(94)00585-0.
Milad MR, Zeidan MA, Contero A, Pitman RK, Klibanski A, Rauch SL, et al. The influence of gonadal hormones on conditioned fear extinction in healthy humans. Neuroscience 2010;168(3):652–658 https://doi.org/https://doi.org/10.1016/j.neuroscience.2010.04.030.
Zeidan MA, Igoe SA, Linnman C, Vitalo A, Levine JB, Klibanski A, et al. Estradiol modulates medial prefrontal cortex and amygdala activity during fear extinction in women and female rats. Biol Psychiatry. 2011;70(10):920–927. https://doi.org/https://doi.org/10.1016/j.biopsych.2011.05.016.
Rasmusson AM, Pinna G, Paliwal P, Weisman D, Gottschalk C, Charney D, et al. Decreased cerebrospinal fluid allopregnanolone levels in women with posttraumatic stress disorder. Biol Psychiatry 2006;60(7):704–713. https://doi.org/https://doi.org/10.1016/j.biopsych.2006.03.026.
Pinna G, Rasmusson AM. Ganaxolone improves behavioral deficits in a mouse model of post-traumatic stress disorder. Front Cell Neurosci 2014;8:256. https://doi.org/https://doi.org/10.3389/fncel.2014.00256.
van der Westhuizen ET, Valant C, Sexton PM, Christopoulos A. Endogenous allosteric modulators of G protein-coupled receptors. J Pharmacol Exp Ther 2015;353(2):246–260. https://doi.org/https://doi.org/10.1124/jpet.114.221606.
Stern CA, Gazarini L, Vanvossen AC, Zuardi AW, Galve-Roperh I, Guimaraes FS, et al. Δ9-tetrahydrocannabinol alone and combined with cannabidiol mitigate fear memory through reconsolidation disruption. Eur Neuropsychopharmacol 2015;25(6):958–965. https://doi.org/https://doi.org/10.1016/j.euroneuro.2015.02.001.
Murkar A, Kent P, Cayer C, James J, Durst T, Merali Z. Cannabidiol and the remainder of the plant extract modulate the effects of Δ9-tetrahydrocannabinol on fear memory reconsolidation. Front Behav Neurosci 2019;13:174. https://doi.org/https://doi.org/10.3389/fnbeh.2019.00174.
Brunet A, Orr SP, Tremblay J, Robertson K, Nader K, Pitman RK. Effect of post-retrieval propranolol on psychophysiologic responding during subsequent script-driven traumatic imagery in post-traumatic stress disorder. J Psychiatr Res 2008;42(6):503–506. https://doi.orge/https://doi.org/10.1016/j.jpsychires.2007.05.006.
Schwabe L, Nader K, Pruessner JC. Reconsolidation of human memory: brain mechanisms and clinical relevance. Biol Psychiatry 2014;76(4):274–280. https://doi.org/https://doi.org/10.1016/j.biopsych.2014.03.008.
Sevenster D, Beckers T, Kindt M. Fear conditioning of SCR but not the startle reflex requires conscious discrimination of threat and safety. Front Behav Neurosci 2014;8:32. https://doi.org/https://doi.org/10.3389/fnbeh.2014.00032.
Soeter M, Kindt M. Disrupting reconsolidation: pharmacological and behavioral manipulations. Learn Mem 2011;18(6):357–366. https://doi.org/https://doi.org/10.1101/lm.2148511.
Wood NE, Rosasco ML, Suris AM, Spring JD, Marin MF, Lasko NB, et al. Pharmacological blockade of memory reconsolidation in posttraumatic stress disorder: three negative psychophysiological studies. Psychiatry Res 2015;225(1–2):31–39. https://doi.org/https://doi.org/10.1016/j.psychres.2014.09.005.
Suzuki A, Josselyn SA, Frankland PW, Masushige S, Silva AJ, Kida S. Memory reconsolidation and extinction have distinct temporal and biochemical signatures. J Neurosci 2004;24(20):4787–4795. https://doi.org/https://doi.org/10.1523/JNEUROSCI.5491-03.2004.
Russo EB. Cannabis therapeutics and the future of neurology. Front Integr Neurosci 2018;12:51. https://doi.org/https://doi.org/10.3389/fnint.2018.00051.
Calabrese EJ, Rubio-Casillas A. Biphasic effects of THC in memory and cognition. Eur J Clin Investig 2018;48(5):e12920. https://doi.org/https://doi.org/10.1111/eci.1292.
Leweke FM, Koethe D. Cannabis and psychiatric disorders: it is not only addiction. Addict Biol 2008;13(2):264–275. https://doi.org/https://doi.org/10.1111/j.1369-1600.2008.00106.x.
Hirvonen J, Goodwin RS, Li CT, Terry GE, Zoghbi SS, Morse C, et al. Reversible and regionally selective downregulation of brain cannabinoid CB1 receptors in chronic daily cannabis smokers. Mol Psychiatry 2012;17(6):642–649. https://doi.org/https://doi.org/10.1038/mp.2011.82.
Bambico FR, Nguyen NT, Katz N, Gobbi G. Chronic exposure to cannabinoids during adolescence but not during adulthood impairs emotional behaviour and monoaminergic neurotransmission. Neurobiol Dis 2010;37(3):641–655. https://doi.org/https://doi.org/10.1016/j.nbd.2009.11.020.
Klugmann M, Klippenstein V, Leweke FM, Spanagel R, Schneider M. Cannabinoid exposure in pubertal rats increases spontaneous ethanol consumption and NMDA receptor associated protein levels. Int J Neuropsychopharmacol 2011;14(4):505–517. https://doi.org/https://doi.org/10.1017/S1461145710001562.
Mechoulam R, Parker LA, Gallily R. Cannabidiol: an overview of some pharmacological aspects. J Clin Pharmacol 2002;42 S1:11S–19S. https://doi.org/https://doi.org/10.1002/j.1552-4604.2002.tb05998.x.
Izzo AA, Borrelli F, Capasso R, Di Marzo V, Mechoulam R. Non-psychotropic plant cannabinoids: new therapeutic opportunities from an ancient herb. Trends Pharmacol Sci 2009;30(10):515–527. https://doi.org/https://doi.org/10.1016/j.tips.2009.07.006.
Grotenhermen F. Pharmacokinetics and pharmacodynamics of cannabinoids. Clin Pharmacokinet 2003;42(4):327–360. https://doi.org/https://doi.org/10.2165/00003088-200342040-00003.
Hazekamp A, Ruhaak R, Zuurman L, van Gerven J, Verpoorte R. Evaluation of a vaporizing device (volcano) for the pulmonary administration of tetrahydrocannabinol. J Pharm Sci 2006;95(6):1308–1317. https://doi.org/https://doi.org/10.1002/jps.20574.
Sethi BB, Trivedi JK, Kumar P, Gulati A, Agarwal AK, Sethi N. Antianxiety effect of Cannabis: involvement of central benzodiazepine receptors. Biol Psychiatry 1986;21(1):3–10. https://doi.org/https://doi.org/10.1016/0006-3223(86)90003-X.
Onaivi ES, Green MR, Martin BR. Pharmacological characterization of cannabinoids in the elevated plus maze. J Pharmacol Exp Ther. 1990;253(3):1002–9.
Rubino T, Sala M, Viganò D, Braida D, Castiglioni C, Limonta V, et al. Cellular mechanisms underlying the anxiolytic effect of low doses of peripheral Δ9-tetrahydrocannabinol in rats. Neuropsychopharmacology 2007;32(9):2036–2045. https://doi.org/https://doi.org/10.1038/sj.npp.1301330.
Rubino T, Guidali C, Vigano D, Realini N, Valenti M, Massi P, et al. CB1 receptor stimulation in specific brain areas differently modulate anxiety-related behaviour. Neuropharmacology 2008;54(1):151–160. https://doi.org/https://doi.org/10.1016/j.neuropharm.2007.06.024.
Steindel F, Lerner R, Häring M, Ruehle S, Marsicano G, Lutz B, et al. Neuron-type specific cannabinoid-mediated G protein signalling in mouse hippocampus. J Neurochem 2013;124(6):795–807. https://doi.org/https://doi.org/10.1111/jnc.12137.
Bedse G, Hartley ND, Neale E, Gaulden AD, Patrick TA, Kingsley PJ, et al. Functional redundancy between canonical endocannabinoid signaling Systems in the Modulation of anxiety. Biol Psychiatry 2017;82(7):488–499. https://doi.org/https://doi.org/10.1016/j.biopsych.2017.03.002.
Kubilius RA, Kaplick PM, Wotjak CT. Highway to hell or magic smoke? The dose-dependence of Δ9-THC in place conditioning paradigms. Learn Mem 2018;25(9):446–454. https://doi.org/https://doi.org/10.1101/lm.046870.117.
Solowij N, Broyd S, Greenwood LM, van Hell H, Martelozzo D, Rueb K, et al. A randomised controlled trial of vaporised Δ9-tetrahydrocannabinol and cannabidiol alone and in combination in frequent and infrequent cannabis users: acute intoxication effects. Eur Arch Psychiatry Clin Neurosci 2019;269(1):17–35. https://doi.org/https://doi.org/10.1007/s00406-019-00978-2.
Szkudlarek HJ, Desai SJ, Renard J, Pereira B, Norris C, Jobson CEL, et al. Δ-9-tetrahydrocannabinol and Cannabidiol produce dissociable effects on prefrontal cortical executive function and regulation of affective behaviors. Neuropsychopharmacology 2019;44(4):817–825. https://doi.org/https://doi.org/10.1038/s41386-018-0282-7.
Malone DT, Jongejan D, Taylor DA. Cannabidiol reverses the reduction in social interaction produced by low dose Delta(9)-tetrahydrocannabinol in rats. Pharmacol Biochem Behav 2009;93(2):91–96. https://doi.org/https://doi.org/10.1016/j.pbb.2009.04.010.
Reid MJ, Bornheim LM. Cannabinoid-induced alterations in brain disposition of drugs of abuse. Biochem Pharmacol 2001;61(11):1357–1367. https://doi.org/https://doi.org/10.1016/S0006-2952(01)00616-5.
Bornheim LM, Grillo MP. Characterization of cytochrome P450 3A inactivation by cannabidiol: possible involvement of cannabidiol-hydroxyquinone as a P450 inactivator. Chem Res Toxicol. 1998;11(10):1209–16 https://doi.org/10.1021/tx9800598.
Klein C, Karanges E, Spiro A, Wong A, Spencer J, Huynh T, et al. Cannabidiol potentiates Δ9-tetrahydrocannabinol (THC) behavioural effects and alters THC pharmacokinetics during acute and chronic treatment in adolescent rats. Psychopharmacology 2011;218(2):443–457. https://doi.org/https://doi.org/10.1007/s00213-011-2342-0.
Kasten CR, Zhang Y, Boehm SL 2nd. Acute cannabinoids produce robust anxiety-like and locomotor effects in mice, but long-term consequences are age- and sex-dependent. Front Behav Neurosci 2019;13:32. https://doi.org/https://doi.org/10.3389/fnbeh.2019.00032.
Zuardi AW, Teixeira NA, Karniol IC. Pharmacological interaction of the effects of delta 9-trans-tetrahydrocannabinol and cannabidiol on serum corticosterone levels in rats. Arch Int Pharmacodyn Ther. 1984;269(1):12–9.
Jacobs DS, Kohut SJ, Jiang S, Nikas SP, Makriyannis A, Bergman J. Acute and chronic effects of cannabidiol on Δ9-tetrahydrocannabinol (Δ9 THC)-induced disruption in stop signal task performance. Exp Clin Psychopharmacol 2016;24(5):320–330. https://doi.org/https://doi.org/10.1037/pha0000081.
Wright MJ Jr, Vandewater SA, Taffe MA. Cannabidiol attenuates deficits of visuospatial associative memory induced by Δ(9) tetrahydrocannabinol. Br J Pharmacol 2013;170(7):1365–1373. https://doi.org/https://doi.org/10.1111/bph.12199.
Nadulski T, Pragst F, Weinberg G, Roser P, Schnelle M, Fronk EM, et al. Randomized, double-blind, placebo-controlled study about the effects of cannabidiol (CBD) on the pharmacokinetics of Delta9-tetrahydrocannabinol (THC) after oral application of THC verses standardized cannabis extract. Ther Drug Monit 2005;27(6):799–810. https://doi.org/https://doi.org/10.1097/01.ftd.0000177223.19294.5c.
Acknowledgements
We thank Bruna W. Salemme for the kindly English proofreading on the manuscript.
Funding
Our study was supported by Brazilian grants from Fundação Araucária (convênio 006/2017), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; 409615/2016–1) and in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001. Fundação Araucária, CAPES and CNPq had no role in study design; in the collection, analysis and interpretation of data; in the writing of the report; and in the decision to submit the paper for publication.
Author information
Authors and Affiliations
Contributions
CAS, LJB, and AMR designed the sections and contents of the review manuscript. CAS supervised the organization to distribute the writing tasks among the authors and participated in manuscript writing. AMR, JBS, TRS, LJB, and CAS performed the literature searches and participated in the manuscript writing. All the authors critically reviewed and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1 Supplementary Table 1.
Raw data used for calculate the effect sizes of behavioral parameters from studies detailed in Table 1.
Additional file 2 Supplementary Table 2.
Raw data used for calculate the effect sizes of behavioral and autonomic parameters from studies detailed in Table 2.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Raymundi, A.M., da Silva, T.R., Sohn, J.M.B. et al. Effects of ∆9-tetrahydrocannabinol on aversive memories and anxiety: a review from human studies. BMC Psychiatry 20, 420 (2020). https://doi.org/10.1186/s12888-020-02813-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12888-020-02813-8