
Abstract
Background
Indian jointvetch (Aeschynomene indica) is a common and pernicious weed found in the upland direct-seeding rice fields in the lower reaches of the Yangtze River in China. However, there are few reports on the degree of harm, genetic characteristics, and management methods of this weed. The purpose of this study is to clarify the harm of Indian jointvetch to upland direct-seeding rice, analyze the genetic characteristics of this weed based on chloroplast genomics and identify its related species, and screen herbicides that are effective in managing this weed in upland direct-seeding rice fields.
Results
In a field investigation in upland direct-seeding rice paddies in Shanghai and Jiangsu, we determined that the plant height and maximum lateral distance of Indian jointvetch reached approximately 134.2 cm and 57.9 cm, respectively. With Indian jointvetch present at a density of 4/m2 and 8/m2, the yield of rice decreased by approximately 50% and 70%, respectively. We further obtained the first assembly of the complete chloroplast (cp.) genome sequence of Indian jointvetch (163,613 bp). There were 161 simple sequence repeats, 166 long repeats, and 83 protein-encoding genes. The phylogenetic tree and inverted repeat region expansion and contraction analysis based on cp. genomes demonstrated that species with closer affinity to A. indica included Glycine soja, Glycine max, and Sesbania cannabina. Moreover, a total of 3281, 3840, and 3838 single nucleotide polymorphisms were detected in the coding sequence regions of the cp. genomes of S. cannabina voucher IBSC, G. soja, and G. max compared with the A. indica sequence, respectively. A greenhouse pot experiment indicated that two pre-emergence herbicides, saflufenacil and oxyfluorfen, and two post-emergence herbicides, florpyrauxifen-benzyl and penoxsulam, can more effectively manage Indian jointvetch than other common herbicides in paddy fields. The combination of these two types of herbicides is recommended for managing Indian jointvetch throughout the entire growth period of upland direct-seeding rice.
Conclusions
This study provides molecular resources for future research focusing on the identification of the infrageneric taxa, phylogenetic resolution, and biodiversity of Leguminosae plants, along with recommendations for reliable management methods to control Indian jointvetch.
Similar content being viewed by others


Background
The genus Aeschynomene in the family Leguminosae is widely distributed in tropical and subtropical regions of the world, mostly in America, Africa, and Asia, with approximately 200 species identified to date [1,2,3]. Aeschynomene is an annual herbaceous plant that grows as shrubs, with shield-shaped stipules, butterfly-shaped flowers, and segmented leaves connected by diaphragms [4]. Only one species of this genus has been found in China to date, namely A. indica (Indian jointvetch) [2]. It usually germinates in April and its seeds mature in October. The germination of Indian jointvetch seeds mainly depends on the temperature during the day [5]. Research has shown that the seeds of Indian jointvetch have dormancy characteristics, which can survive for about six months in the soil seed bank and mechanical scarification is an effective method to break dormancy [6].
Although this plant does not germinate under flooded conditions, once it grows, Indian jointvetch indirectly affects crop productivity by competing for resources, hindering harvesting operations, and reducing grain and seed quality [7,8,9]. Moreover, it has been confirmed that mixing seeds of A. indica into rice as pig feed can induce the development of pig diseases [1]. A. indica was reported to be a weed in irrigated rice fields previously [10, 11] and was considered the third most troublesome weed after weedy rice and barnyard grass in certain rice-producing areas [12]. However, A. indica was recently found to impair the growth of upland direct-seeding rice in the lower reaches of the Yangtze River in China (Additional File 3, Figure S1). Because the planting area of upland direct-seeding rice is increasing owing to the global shortage of labor and water resources, more and more attention has been paid to the serious weed problem [13,14,15,16]. Distinguishing plants of the genus Aeschynomene is very difficult, and managing these weeds in rice fields is also very challenging presently [3]. Therefore, further research is required to elucidate the characteristics of this species, including the genetic characteristics, degree of harm induced, and responses to available herbicides for effective management and control.
Chloroplasts are the main organelles involved in photosynthesis in photosynthetic plants or algae [17]. Chloroplasts have their own distinct genetic material with characteristics of non-recombination, haploid, and single-parent inheritance, which leads to a highly conserved genome; therefore, analysis of the chloroplast (cp.) genome can provide very rich evolutionary information [18,19,20]. Additionally, the cp. genome is small and easy to obtain, offering unique research value in phylogeny, species identification, and population genetics [21]. The cp. genome has a typical tetrad structure, with large single copy (LSC) and small single copy (SSC) regions separated by inverted repeats (IRs) [22,23,24]. Previous studies have shown that in the context of network evolution (i.e., hybridization) and polyploidy, analysis of the cp. genome is particularly useful for characterizing the phylogenetic and historical aspects of most plant lineages [25,26,27]. However, information on the composition, structure, interspecific differences, and evolutionary relationships of A. indica based on its cp. genome is limited. Therefore, exploring the cp. genomic information of A. indica can provide a theoretical basis for the management or utilization of this species and predict the possibility of evolution of related species into paddy weeds.
Current weed control methods in upland direct-seeding rice fields mainly include cultural weed management practices and physical methods such as deep plowing, germination, mechanical weeding, and hand-pulling weeding, along with chemical methods using herbicides [28,29,30]. Chemical methods for weed control in upland direct-seeding rice fields have specific advantages of obtaining higher yields with relatively lower labor costs [30,31,32]. Aeschynomene weeds are often controlled using a combination of imazapyr and imazapic herbicides [3]. However, this genus of weed has been found to exhibit imazapic resistance [33]. In addition, this herbicide is typically considered to be unsafe for rice growth. To our knowledge, there have been no systematic studies on the toxicity of common herbicides used for A. indica in rice fields. Therefore, screening for suitable herbicides to effectively control A. indica is of great significance to achieve a high and optimal yield of upland direct-seeding rice.
To address these questions, we performed an in-depth analysis of the genetic characteristics and herbicide response of A. indica growing in the upland direct-seeding rice fields of Shanghai, China. We studied the morphological characteristics of this weed in rice fields and its effect on rice yield. The cp. genome of A. indica was then sequenced and compared to those of other species to analyze their genetic characteristics and evolutionary relationships. Finally, the toxicities of common herbicides currently used in rice fields in China against A. indica were evaluated in a greenhouse pot experiment to determine a reasonable chemical control method. Our study thus provides a theoretical and practical foundation for gaining a better understanding of the damage of the troublesome dicotyledonous weed A. indica imposes in upland direct-seeding rice fields and facilitating strategies for its effective management.
Methods
Plant materials
Indian jointvetch seeds were collected from upland direct-seeding rice fields in Jinshan District, Shanghai, China (N 30.81°, E 121.18°) in October 2021. The seeds were dried to constant weight after collection and stored at 4 °C until planted.
Determination of field traits
Plant height and the maximum lateral distance of Indian jointvetch growing in upland direct-seeding rice fields in Shanghai and Jiangsu Province were measured in October 2021. More than 20 plants were randomly selected; however, no more than three plants were sampled from each paddy field. Seeds were collected, dried to a constant weight, and weighed. A total of 294 upland direct-seeding rice fields were investigated, fields with Indian jointvetch were recorded, and their frequency was calculated.
Subsequently, 0, 1, 2, 3, and 4 Indian jointvetch seedlings grown in a greenhouse previously were transplanted into a 1 m2 upland direct-seeding rice planting area and all other weeds were manually removed. When the upland direct-seeding rice had matured, the effective spike number, grains per spike, and 1000-seed weight of upland direct-seeding rice seeds were measured. Each treatment contains three biological replicates and the entire experiment was repeated twice. Data of percentages of fresh weight compared to the control (none transplanted Indian jointvetch) were subjected to ANOVA. For comparison of the differences in the rice yield indicators among the five groups, the Duncan’s Multiple Range Test (P < 0.05) was used. ANOVA was conducted using SPSS version 20 (SPSS, Chicago, IL, USA).
Construction of the A. indica cp. genome
DNA sequencing and genome assembly
Total genomic DNA from ten Indian jointvetch at the seedling stage was extracted using a modified cetyltrimethylammonium bromide method and applied to construct a 500-bp paired-end library using the NEBNext Ultra DNA Library Prep Kit (New England Biolabs, Ipswich, MA, USA) for next-generation sequencing on the Illumina NovaSeq 6000 platform (Berry Genomics Co., Ltd., Beijing, China). Approximately 2.35 Gb of raw data from Indian jointvetch were generated with 150-bp paired-end read lengths. De novo assembly was performed using NOVOPlasty v4.2 software (https://github.com/ndierckx/NOVOPlasty, accessed March 24, 2023). Detailed sequencing and assembly methods are described in our previous study [34].
Genome component analysis and gene annotation
Genes encoding proteins, tRNAs, and rRNAs in the cp. genome of Indian jointvetch were predicted using GeSeq software (https://chlorobox. mpimp-golm. mpg. de/geseq. com ml/, accessed March 24, 2023). The protein sequences of cp. genes were compared with known proteins in databases using BLASTP (https://ncbiinsights.ncbi.nlm.nih.gov/tag/blastp/, accessed March 24, 2023) (e-value < 1 × 10− 5). Because there may be more than one alignment result for each sequence, only one optimal alignment result was reserved for database alignment for the given gene to ensure its biological significance. The amino acid sequences of Indian jointvetch were aligned with sequences included in the Non-Redundant Protein Sequence (http://www.ncbi.nlm.nih.gov/, accessed March 24, 2023), Swiss-Prot (http://www.ebi.ac.uk/uniprot, accessed March 24, 2023), Clusters of Orthologous Groups, Kyoto Encyclopedia of Genes and Genomes (KEGG; http://www.genome.jp/kegg/, accessed March 24, 2023), and Gene Ontology (GO; http://geneontology.org/, accessed March 24, 2023) databases to obtain functional annotation information for the coding genes.
Analysis of genetic relationships and identification characteristics
Phylogenetic analysis
Nineteen cp. genomes of common plants, including the model plant of dicotyledons Arabidopsis thaliana, common plants growing in rice fields (Oryza sativa Indica, O. sativa Tropical Japonica, Echinochloa oryzoides, Eclipta prostrata, Ammannia arenaria, Ammannia multiflora, Cyperus iria, Cyperus difformis), and common Leguminosae plants (Vicia sepium, Sesbania cannabina, Medicago sativa, Medicago truncatula, Medicago polymorpha, Glycine max, Glycine soja, Astragalus sinicus), were downloaded from the National Center for Biotechnology Information (NCBI) database (see Additional File 1, Table S1 for accession numbers) and subject to phylogenetic analysis to determine their evolutionary relationships with Indian jointvetch. The sequences were aligned using ClustalW (v2.0.12) (http://www.clustal.org/clustal2/, accessed April 4, 2023) with default settings. The DNA substitution model was utilized based on the Akaike information criterion [35]. The phylogenetic tree was constructed by the maximum-likelihood (ML) method using PhyML v3.0 (htp://ww.atgc-montpeller. fr/phyml/, accessed April 4, 2023) and bootstrap values were calculated for 1000 replicates [36, 37]. The tree building model was finally evaluated using jModelTest 2.1.10 (https://github.com/ddarriba/jmodeltest2, accessed April 4, 2023), with the best model “GTR + I + G.”
Contraction and expansion analysis of IR regions
We performed IR contraction and expansion analysis for the cp. genomes of Indian jointvetch and species with closer genetic relationships selected based on the phylogenetic analysis. The four quadripartite structures (LSC, SSC, and two IR repeat regions) of each cp. genome were compared, and changes in the copy number of related genes caused by contraction and expansion of the IR or pseudogenes, resulting in boundary regions, were analyzed. Genes that crossed or were adjacent to these boundaries were identified. In addition, the length and distance from the boundaries of these genes were analyzed.
Single nucleotide polymorphism (SNP) analysis
Using MUMmer software (http://mummer.sourceforge.net/, accessed April 4, 2023), the cp. genome sequences of Indian jointvetch (as the reference) and closely related species, based on phylogenetic and contraction/expansion IR analyses, were completely aligned, and preliminary filtering was performed to detect potential SNP sites. Sequences of 100 bp on both sides of the candidate SNP site of the reference sequence were extracted and aligned with the assembly results using BLAT v35 software (http://hgdownload.soe.ucsc.edu/admin/exe/linux.x86_64/blat/, accessed April 4, 2023) to verify the SNP site. If the alignment length was less than 101 bp, it was considered an unreliable SNP and was removed; if the alignment was repeated multiple times, the SNP was considered a repetitive region and was removed, resulting in only reliable SNPs.
Herbicide sensitivity assays
We determined the susceptibility of Indian jointvetch to common herbicides applied in rice fields using a pot experiment in the greenhouse. The soil was a middle loam obtained from a farmland in the suburbs of Shanghai, China, where herbicides had not been used. Five seeds were sown in each plastic cup (7 × 7 × 7 cm), water was added until saturation, a fine layer of soil was added, and the seeds were treated with herbicides (i.e., pre-emergence treatment) using a 3WP-2000 walking-type spray system (Nanjing Agricultural Mechanization Research Institute of the Ministry of Agriculture, China). Each treatment was performed with 30 mL of liquid (450 L ha− 1 water) using a fan-shaped nozzle. Control plants were sprayed with the same amount of water as the herbicide for the treated plants. The seedlings were then placed in a greenhouse for cultivation. When the seeds of the control group germinated and the treatment group showed a significant gradient, the Indian jointvetch plants were cut and weighed. Five seeds were sown and cultivated at the three-leaf stage for post-emergence herbicide application using the same method. After 21 days, the above-ground Indian jointvetch in each cup was cut and weighed. The dose and details of the herbicides are listed in Table 1. The experiment included three to four biological replicates and the entire experiment was repeated twice. The effective rate of each herbicide causing 50% reduction in plant growth (GR50) was determined using the four-parameter logistic function with the “drc” add-on package [38] in the R 3.1.3 Language and Environment for Statistical Computing [39]. This model is defined as follows:
where parameter e represents GR50 as the dose producing a response halfway between the upper limit d and the lower limit c, and parameter b denotes the relative slope around e.
Results
Impact of A. indica on rice growth
We first investigated the morphological features of Indian jointvetch during seed maturity in upland paddy fields in the lower reaches of the Yangtze River in China. The average height of Indian jointvetch was 134.2 ± 3.7 cm, which was higher than that of the rice (approximately 90 cm). The mean maximum lateral distance was 57.9 ± 2.1 cm. The seeds presented a kidney shape (Additional File 3, Figure S2) with a 1000-seed weight of 10.39 ± 0.055 g. The frequency of occurrence of this weed in upland paddy fields in our survey was 13.85 ± 2.22%. From the perspective of rice damage, as the number of Indian jointvetch plants increased, the effective spike number and grains per spike of upland direct-seeding rice exhibited a significant downward trend. Rice seed weights also showed an overall downward trend with an increasing number of Indian jointvetch plants. Specifically, when the number of Indian jointvetch was 0, 2, 4, 6, and 8, the yields of upland direct-seeding rice were 8019.3, 5534.4, 3783.2, 3611.4, and 2092.5 kg/ha, respectively (Fig. 1).

Effect of the number of Indian jointvetch plants on rice yield in the field. The standard errors of the means are described by vertical bars. ANOVA significance groupings are shown as a, b, c, and d (P < 0.05)
Characteristics of the A. indica cp genome
Cp genome map
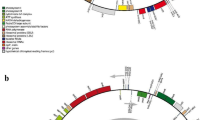
After trimming low-quality fragments from the raw data, 33,624,994 clean reads with a GC content of 35.93% were mapped to the complete cp. genome of Indian jointvetch. De novo assembly resulted in a circular genome of 163,613 bp in length (Fig. 2). The raw reads were deposited in the NCBI GenBank database (accession number: PRJNA963187). The complete cp. genome displayed the typical quadripartite structure of most angiosperms, including an LSC region, SSC region, and pair of IRs (IRa and IRb). The lengths of the LSC, SSC, and IR regions were 88,562, 19,801, and 27,625 bp, respectively, and the intergenic region length was 86,150 bp. The Indian jointvetch cp. genome contains 83 protein-coding genes (Table 2).

Assembly, size, and features of the chloroplast genome of Indian jointvetch. The genes outside the circle are transcribed in the counterclockwise direction and the genes inside the circle are transcribed in the clockwise direction. Different colors of genes represent different functions. The dark gray area and light gray area of the inner circle represent the GC and AT content of the genome, respectively
Cp genome components
The cp. genome of Indian jointvetch contained 39 tRNA genes and eight rRNA genes (Table 3). There were 70 protein-coding and 26 tRNA genes located within the LSC; 10 protein-coding genes, 10 tRNA-coding genes, and four rRNA-coding genes located within IRb or IRa; and 13 protein-coding genes and one tRNA gene located within the SSC region (Fig. 2). All 83 genes encoding proteins in the cp. genome of Indian jointvetch were functionally annotated, which mainly belonged to the photosynthesis and self-replication categories. The gene names, groups, and categories are listed in Table 4. The genes were mainly associated with GO biological processes (Additional File 3, Figure S3), and were associated with KEGG energy production and conversion, translocation, ribosomal structure, and biogenesis pathways (Additional File 3, Figure S4). In total, 161 simple sequence repeats (SSRs) were identified in the Indian jointvetch cp. genome. There were eight SSRs on IRa or IRb, 114 on the LSC region, and 31 on the SSC region. In total, 166 long repeats (LRs) were identified in the cp. genome of Indian jointvetch (Table 5).
Genetic similarity analysis
Phylogenetic analysis
Bayesian inference of 19 complete cp. genomes showing the same topology demonstrated that Indian jointvetch clustered into a single clade (Fig. 3). Indian jointvetch, A. sinicus, M. polymorpha, G. soja, and G. max have the most recent common ancestor (MRCA) (BS = 100 for the ML tree), and this group has an MRCA with S. cannabina voucher IBSC voucher (BS = 100 for the ML tree). The closest relatives to these plants were E. prostrata, O. sativa Indica, O. sativa Tropical Japonica, and E. oryzicola (BS = 100 for the ML tree). In addition, the group formed by S. cannabina-1, M. sativa, M. polymorpha, A. thaliana, C. iria, C. difformis, A. arenaria, and A. multiflora was closely related to the Indian jointvetch group. The most distantly related plant to Indian jointvetch was V. sepium.

Phylogenetic tree constructed using the maximum-likelihood method based on alignments of complete chloroplast genome sequences. The numbers at the nodes indicate bootstrap values from 1000 replicates. If the bootstrap value is 100, this number is not shown on the nodes. The species newly sequenced in this study is shown in bold
IR expansion and contraction
To further observe the potential genetic relationships among the most closely related species to Indian jointvetch, namely S. cannabina voucher IBSC, G. soja, and G. max, based on the contraction and expansion of IR regions, the gene variations at the IR/SSC and IR/LSC boundary regions of the four species were compared (Fig. 4); A. sinicus and M. polymorpha do not have complete IR regions and were therefore excluded from this analysis. The genes rps19/ rpl2, trnN/ ndhF, ycf1/ trnN, and rpl2/ trnH were identified at the junctions of the LSC/IRb, IRb/SSC, SSC/IRa, and IRa/LSC regions, respectively, in Indian jointvetch. Among them, ndhF crosses IRb/SSC and ycf1 crosses SSC/IRa. Among all tested species, the closest to Indian jointvetch was S. cannabina voucher IBSC with respect to the expansion and contraction of IR regions; these two species exhibited identical boundary genes with the only difference being the length of the genes or the distance between the genes and the boundary.

Comparison of large sequence copy (LSC), inverted repeat (IRb, IRa), and small sequence copy (SSC) border regions of the chloroplast genomes of Indian jointvetch and related species
SNP analysis
SNP analysis was performed to compare the differentiation among Indian jointvetch, S. cannabina voucher IBSC, G. soja, and G. max. A total of 4659 SNPs were detected in S. cannabina voucher IBSC, including 1378 SNPs in the intergenic spacer regions and 3281 SNPs in the coding sequence regions (Table 6; Additional File 2, Table S2). The nonsynonymous to synonymous substitution ratio was 0.626. Two SNPs were detected at the start codon and eight SNPs were detected at the stop codon. The numbers of SNPs in the cp. genomes of G. soja and G. max were much higher than that for S. cannabina voucher IBSC compared to the Indian jointvetch cp. genome.
Chemical herbicide effects
Toxicity of pre-emergence herbicides
In pre-emergence application, as the doses of oxyfluorofen and saflufenacil increased, Indian jointvetch was significantly inhibited on the 14th day after herbicide treatment and showed a gradient decrease pattern thereafter. However, upon treatment with the other two herbicides, bensulfuron-methyl and butralin, Indian jointvetch did not show significant growth inhibition until 21 days after treatment (Fig. 5). The toxicity calculation showed that the GR50 value of oxyfluorofen on Indian jointvetch was 34.9 g a.i. ha− 1 and the GR90 value was 157.5 g a.i. ha− 1; the GR50 value of saflufenacil on Indian jointvetch was 1.3 g a.i. ha− 1 and the GR90 value was 10.3 g a.i. ha− 1 (Fig. 5).

Dose–response analyses for the response of Indian jointvetch to pre-emergence herbicides. The X-axis represents the dose (g a.i. ha− 1) and the Y-axis represents the percentage of fresh weight to that of the untreated control. “Bm,” “Sf,” “Of,” and “Bl” represent the herbicides bensulfuron-methyl, saflufenacil, oxyfluorofen, and butralin, respectively
Toxicity of post-emergence herbicides
With post-emergence application, with the increase in the doses of florpyrauxifen-benzyl, pyrazosulfuron-ethyl, and penoxsulam, Indian jointvetch was significantly inhibited on the 21st day after herbicide treatment and showed a decreasing gradient thereafter; the other three herbicides, quinclorac, 2-methyl-4-chlorophenoxyacetic acid, and pyraquinate, did not have a significant inhibitory effect on the growth of Indian jointvetch. From the toxicity calculation, the GR50 value of florpyrauxifen-benzyl on Indian jointvetch was 0.3 g a.i. ha− 1 and the GR90 value was 1.3 g a.i. ha− 1. The GR50 value of penoxsulam on Indian jointvetch was 4.0 g a.i. ha− 1 and the GR90 value was 16.8 g a.i. ha− 1. The GR50 value of pyrazosulfuron-ethyl on Indian jointvetch was 11.9 g a.i. ha− 1 and the GR90 value was g a.i. ha− 1 737.5, but this far exceeds its safe usage limit (45.0 g a.i. ha− 1) (Fig. 6).

Dose–response analyses for response of Indian jointvetch to post-emergence herbicides. The X-axis represents the dose (g a.i. ha− 1) and the Y-axis represents the percentage of fresh weight to that of the untreated control. “Fb,” “Ps,” “MCPA,” “Qc,” “Pq,” and “Pz” represent the herbicides florpyrauxifen-benzyl, penoxsulam, 2-methyl-4-chlorophenoxyacetic acid, quinclorac, pyraquinate, and pyrazosulfuron-ethyl, respectively
Discussion
According to the results of our on-site investigation, Indian jointvetch causes serious harm to upland direct-seeding rice fields in the lower reaches of the Yangtze River in China. Upland direct-seeding rice is an important food crop grown in areas with limited water resources presently. However, rampant weeds are an obstacle to the cultivation of upland direct-seeding rice [14,15,16]. The rough management practice of the upland direct-seeding rice field system has further aggravated weed ravages. Although studies have shown that Indian jointvetch can be considered a green fertilizer that is beneficial for rice growth [40], we obtained contrasting results. In fact, the tall Indian jointvetch plant type has reduced the survival resources of rice, thereby seriously restricting its yield. However, research on Indian jointvetch in rice fields is limited.
Given the serious harm caused by Indian jointvetch in rice fields, genetic research is important to gain a deeper understanding of this weed and predict its evolution in rice fields in the future. Because the cp. genome has unique research value in phylogeny, species identification, and population genetics [21], we sequenced the cp. genome of Indian jointvetch for the first time in this study, demonstrating a typical tetrad structure, including genes encoding proteins, tRNAs, and rRNAs, similar to the cp. genomes reported in other plants [22, 34, 41]. The complete cp. genome of Indian jointvetch was 163,613 bp long, which is larger than that of other common plants found in rice fields with reported cp. genomes, including those of the genera Oryza [42], Echinochloa [41], and Ammannia [34].
Repeating sequences are unique labels of a species, including SSRs and LRs. SSRs, composed of 1–6 nucleotide repeat units, are widely distributed molecular genetic markers in the cp. genome of plants [43, 44]. We identified 161 SSRs in the cp. genome of Indian jointvetch, which can be used for population genetics analyses and plant genotyping [45,46,47,48]. We also identified 166 LRs in the Indian jointvetch cp. genome. LRs are a special type of DNA repeat sequence that typically occupy a large proportion of the genome [49]. These repetitive structures exhibit diversity in the cp. genome and promote molecular recombination [50], which has important molecular and biological significance in the study of plant evolution [51]. Thus, the SSRs and LRs detected in this study offer important biological information resources for the identification of Indian jointvetch and to facilitate genetic diversity and population structure research of Aeschynomene.
In order to explore the closely related plants of Indian jointvetch, we conducted phylogenetic tree analysis, IR expansion and contraction comparisons, and SNP studies based on the cp. genome. Genome assembly and characterization provides great value in defining species, revealing phylogenetic relationships, and determining taxonomic status [52,53,54,55]. Phylogenetic analysis based on the cp. genomes showed that species with a close genetic relationship to Indian jointvetch included A. sinicus, M. polymorpha, G. soja, G. max, and Sesbania cannabina voucher IBSC. Notably, the genetic relationship between the two species of S. cannabina registered in the NCBI database is not close, suggesting that they may in fact represent different species or diverged due to geographical isolation and local adaptation. IR expansion and contraction in the cp. genome are common phenomena in plants [18], mainly occurring at the IR/SC junction [56], which is a key driving force in plant evolution [57,58,59,60]. The results of IR expansion and contraction analyses further indicated that the genetic relationship between Indian jointvetch and S. cannabina voucher IBSC was closer. Because their boundary genes are completely identical, except for differences in length of the genes or the distance between the genes and the boundary (Fig. 4). SNPs are important indicators of evolutionary differences between closely related species. These direct molecular markers can clearly display the exact nature and location of allele variations [61]. Using the cp. genome of Indian jointvetch as reference, we identified 3281, 3840, and 3838 SNPs in the cp. genomes of S. cannabina voucher IBSC, G. soja, and G. max, respectively. The fewer SNPs detected represents the fewer variations in single nucleotides. This result further indicated that the Indian jointvetch and S. cannabina voucher IBSC have a relatively close genetic relationship, which is consistent with the phylogeny and IR expansion and contraction results. These SNPs can be used as important differential nucleotide databases to distinguish the species.
From the perspective of crop protection, it is necessary to study reasonable measures to eliminate Indian jointvetch from paddy fields. The advantages of chemical methods for weed control in upland direct-seeding rice fields have been reported for decades in terms of higher yields and lower labor costs [30,31,32]. However, the chemical control of Indian jointvetch is currently very challenging [3]. Our results also indicated that the availability of herbicides for Indian jointvetch control is limited. Herbicides commonly used to control broadleaf weeds in rice fields in China, such as bensulfuron-methyl and 2-methyl-4-chlorophenoxyacetic acid [62,63,64], had poor effects on inhibiting the germination of Indian jointvetch. Nevertheless, we found that among the 10 herbicides tested, four met the requirements, including two pre-emergence herbicides (saflufenacil and oxyfluorfen) and two post-emergence herbicides (florpyrauxifen-benzyl and penoxsulam). Given that mixing Indian jointvetch seeds with rice seriously reduces the quality of the rice [1], we strongly recommend using herbicides containing these four components to control Indian jointvetch, which is rampant in rice fields; however, further research is needed to establish effective control strategies.
It should be pointed out that the understanding of the genetic characteristics of Indian jointvetch and the prediction of the evolution of related plants into rice field weeds in this study are only based on the chloroplast genome. Further research should be based on broader morphological and whole-genome. Additionally, this study only provides suggestions for chemical control of Indian jointvetch. Further research is needed on control strategies that are more conducive to biodiversity and even the value of resource utilization of this weed.
Conclusions
In this study, we obtained the first complete cp. genome of Indian jointvetch (A. indica), a common weed found in upland direct-seeding rice fields in the lower reaches of the Yangtze River in China. Overall, our study indicates that species with closer affinity to Indian jointvetch include (but are not limited to) Glycine soja, Glycine max, and Sesbania cannabina based on the cp. genome. Therefore, it is necessary to be vigilant about these plants becoming farmland weeds (especially S. cannabina), entering rice fields, and causing harm to rice. Note that because A. sinicus and M. polymorpha do not have complete IR regions, further analyses of these two species were not conducted in this study, which may warrant further exploration of their potential harm in upland direct-seeding rice fields. Toxicity evaluation of common herbicides currently used in rice fields indicate the need for the combination of herbicides to achieve effective control of Indian jointvetch and improve the yield and quality of upland direct-seeding rice.
Data availability
The datasets generated and/or analyzed during the current study are available in the [National Center for Biotechnology Information] repository, [Accession number: PRJNA963187].
Abbreviations
- cp:
-
Chloroplast
- GO:
-
Gene Ontology
- GR50:
-
The herbicide dose needed to cause 50% growth reduction of plant
- IBSC:
-
South China Botanical Garden, Chinese Academy of Sciences
- IR:
-
Inverted Repeat
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- LR:
-
Long repeat
- LSC:
-
Large single copy
- ML:
-
Maximum likelihood
- MRCA:
-
Most recent common ancestor
- NCBI:
-
National Center for Biotechnology Information
- SSC:
-
Small single copy
- SNP:
-
Single nucleotide polymorphism
- SSR:
-
Simple sequence repeat
References
Riet-Correa F, Timm C, Barros S, Summers B. Symmetric focal degeneration in the cerebellar and vestibular nuclei in swine caused by ingestion of Aeschynomene indica seeds. Vet Pathol. 2003;40(3):311–6.
Flora Reipublicae Popularis Sinicae. Available online: http://www.iplant.cn/.
Martins MB, Agostinetto D, Fogliatto S, Vidotto F, Andres A. Aeschynomene spp. Identification and Weed Management in Rice fields in Southern Brazil. Agronomy. 2021;11(3):453.
Hartmann LS, Rodrigues RS, Flores AS. O gênero Aeschynomene (Leguminosae-Papilionoideae) no estado de Roraima. Brasil Rodriguésia. 2019;70:1–14.
Ouyang W, Chen L, Yang Y. The effect of temperature on the germination rate of Sesbania cannabina and Aeschynomene indica and the allelopathic effect between them. South China Agric. 2021;15(20):179–80. (in Chinese).
Martins MB, Munhos TF, Schaedler CE, Agostinetto D, Andres A. Seed dynamics of Aeschynomene denticulata and Aeschynomene indica. Weed Biol Manag. 2021;21(3):172–80.
Fleck NG, Lazaroto CA, Schaedler CE, Ferreira FB. Suscetibilidade De três espécies de angiquinho (Aeschynomene spp.) a herbicidas de utilização em pós-emergência em arroz irrigado. Curr Agricultural Sci Technol 2008;14(3).
Dornelles SHB, Sanchotene DM, Macedo L, Rodrigues SN. Controle pós-tardio de angiquinho Aeschynomene denticulata na cultura do arroz.: Vivências. 2014;10(19):42–9.
Galon L, Guimarães S, Radünz AL, Lima AMD, Burg GM, Zandoná RR, Bastiani MO, Belarmino JG, Perin GF. Competitividade relativa de cultivares de arroz irrigado com Aeschynomene denticulata. Bragantia. 2015;74:67–74.
Kissmann KG, Groth D. Plantas infestantes e nocivas. Volume 1. Basf Brasileira São Paulo; 1991.
Oliveira M. Aeschynomene indica L. no Brasil: Primeira citação. Inheringia. 2001;41:3–8.
Fruet BDL, Junior AM, Ulguim ADR. Survey of rice weed management and public and private consultant characteristics in Southern Brazil. Weed Technol. 2020;34(3):351–6.
Mahajan G, Chauhan BS, Kumar V. Integrated weed management in rice. Recent advances in weed management. New York: Springer; 2014. pp. 125–53.
Fofana, Rauber. Weed suppression ability of upland rice under low-input conditions in West Africa. Weed Res. 2000;40(3):271–80.
Saito K. Weed pressure level and the correlation between weed competitiveness and rice yield without weed competition: an analysis of empirical data. Field Crop Res. 2010;117(1):1–8.
Saito K, Futakuchi K. Improving estimation of weed suppressive ability of upland rice varieties using substitute weeds. Field Crop Res. 2014;162:1–5.
Howe CJ, Barbrook AC, Koumandou VL, Nisbet RER, Symington HA, Wightman TF. Evolution of the chloroplast genome. Philosophical Trans Royal Soc Lond Ser B: Biol Sci. 2003;358(1429):99–107.
Plunkett GM, Downie SR. Expansion and contraction of the chloroplast inverted repeat in Apiaceae subfamily Apioideae. Syst Bot. 2000;25(4):648–67.
Wicke S, Schneeweiss GM, Depamphilis CW, Müller KF, Quandt D. The evolution of the plastid chromosome in land plants: gene content, gene order, gene function. Plant Mol Biol. 2011;76(3):273–97.
Cosner ME, Raubeson LA, Jansen RK. Chloroplast DNA rearrangements in Campanulaceae: phylogenetic utility of highly rearranged genomes. BMC Evol Biol. 2004;4:27.
Hong Z, Wu Z, Zhao K, Yang Z, Zhang N, Guo J, Tembrock LR, Xu D. Comparative analyses of five complete chloroplast genomes from the genus Pterocarpus (Fabacaeae). Int J Mol Sci. 2020;21(11):3758.
Wu FH, Chan MT, Liao DC, Hsu CT, Lee YW, Daniell H, Duvall MR, Lin CS. Complete chloroplast genome of Oncidium Gower Ramsey and evaluation of molecular markers for identification and breeding in Oncidiinae. BMC Plant Biol. 2010;10(1):68.
Li P, Lu RS, Xu WQ, Ohi-Toma T, Cai MQ, Qiu YX, Cameron KM, Fu CX. Comparative genomics and phylogenomics of east Asian tulips (Amana, Liliaceae). Front Plant Sci. 2017;8:451.
Bi Y, Zhang MF, Xue J, Dong R, Du YP, Zhang XH. Chloroplast genomic resources for phylogeny and DNA barcoding: a case study on Fritillaria. Sci Rep-UK. 2018;8(1):1184.
Kugita M, Kaneko A, Yamamoto Y, Takeya Y, Matsumoto T, Yoshinaga K. The complete nucleotide sequence of the hornwort (Anthoceros Formosae) chloroplast genome: insight into the earliest land plants. Nucleic Acids Res. 2003;31(2):716–21.
Henry RJ. Plant diversity and evolution: genotypic and phenotypic variation in higher plants. Biologist. 2005;(1):121.
Yamane K, Yasui Y, Ohnishi O. Intraspecific cpDNA variations of diploid and tetraploid perennial buckwheat, Fagopyrum Cymosum (Polygonaceae). Am J Bot. 2003;90(3):339–46.
Singh R, Ghosh D. Effect of cultural practices on weed management in rainfed upland rice. Int J Pest Manage. 1992;38(2):119–21.
Renu S, George T, Raharn C. Stale seedbed: A technique for non-chemical weed control for direct seeded upland rice. In: Proceedings of the 19th Kerala Science Congress, Kerala, India: 2007;29–31.
Adigun J, Kolo E, Adeyemi O, Daramola O, Badmus A, Osipitan O. Growth and yield response of upland rice to nitrogen levels and weed control methods. Int J Agron Agricultural Res. 2017;11(6):92–101.
Ogungbile A, Lagoke S. On-farm evaluation of the economics of chemical weed control in oxen‐mechanized maize production in Nigerian savanna. Int J Pest Manage. 1986;32(4):269–73.
Kolo E, Adigun JA, Adeyemi OR, Daramola OS, Bodunde JG. Effect of nitrogen levels and weed management methods on weed abundance and yield of upland rice (Oryza sativa L). J Agricultural Sci (Belgrade). 2020;65(2):137–50.
Concenço G, Andres A, Schreiber F, Moisinho I, Martins M. Control of jointvetch (Aeschynomene spp.), establishment and productivity of rice as a function of [imazapic + imazapyr] doses. J Agr Sci. 2018;10(4):287.
Gao Y, Li S, Yuan G, Fang J, Shen G, Tian Z. Comparison of Biological and genetic characteristics between two most common broad-leaved weeds in Paddy Fields: Ammannia Arenaria and A. multiflora (Lythraceae). Biology. 2023;12(7):936.
Sakamoto Y, Ishiguro M, Kitagawa G. Akaike information criterion statistics. Dordrecht, The Netherlands: D Reidel. 1986;81(10.5555):26853.
Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30(9):1312–13.
Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39(4):783–91.
Jiao B, Wang K, Chang Y, Dong F, Pan X, Wu X, Xu J, Liu X, Zheng Y. Photodegradation of the Novel Herbicide Pyraquinate in Aqueous Solution: kinetics, photoproducts, mechanisms, and Toxicity Assessment. J Agric Food Chem. 2023;71(10):4249–57.
Shinozaki K, Ohme M, Tanaka M, Wakasugi T, Hayashida N, Matsubayashi T, Zaita N, Chunwongse J, Obokata J, Yamaguchi-Shinozaki K. The complete nucleotide sequence of the tobacco chloroplast genome: its gene organization and expression. EMBO J. 1986;5(9):2043–9.
Nilanthi D, Inoka K, Madhushani P, Dissanayaka D, Kumarasinghe S. Effect of weeds Diya Siyambala (Aeschynomene indica), Kudamatta (Fimbristylis miliacea) and Thunassa (Cyperus iria) on growth and yield of Rice (Oryza sativa L.) variety Bg 379-2, ld 365, Bg 357. Int J Sci Res Publications. 2015;5(6):473–5.
Gao Y, Shen G, Yuan G, Tian Z. Comparative analysis of whole chloroplast genomes of three common species of Echinochloa (Gramineae) in Paddy Fields. Int J Mol Sci. 2022;23(22):13864.
National Center for Biotechnology Information. 2023; Available online: https://www.ncbi.nlm.nih.gov/nuccore/?term=Oryza+sativa+chloroplast+complete+genome (accessed on 4 April 2023).
Wu M, Li Q, Hu Z, Li X, Chen S. The complete Amomum Kravanh chloroplast genome sequence and phylogenetic analysis of the commelinids. Molecules. 2017;22(11):1875.
Zhou J, Cui Y, Chen X, Li Y, Xu Z, Duan B, Li Y, Song J, Yao H. Complete chloroplast genomes of Papaver rhoeas and Papaver orientale: molecular structures, comparative analysis, and phylogenetic analysis. Molecules. 2018;23(2):437.
Doorduin L, Gravendeel B, Lammers Y, Ariyurek Y, Chin-A-Woeng T, Vrieling K. The complete chloroplast genome of 17 individuals of pest species Jacobaea vulgaris: SNPs, microsatellites and barcoding markers for population and phylogenetic studies. DNA Res. 2011;18(2):93–105.
He S, Wang Y, Volis S, Li D, Yi T. Genetic diversity and Population structure: implications for conservation of wild soybean (Glycine soja Sieb. Et zucc) based on nuclear and chloroplast microsatellite variation. Int J Mol Sci. 2012;13:12608–28.
Yang AH, Zhang JJ, Yao XH, Huang HW. Chloroplast microsatellite markers in Liriodendron tulipifera (Magnoliaceae) and cross-species amplification in L. Chinense. Am J Bot. 2011;98(5):e123–6.
Xue J, Wang S, Zhou SL. Polymorphic chloroplast microsatellite loci in Nelumbo (Nelumbonaceae). Am J Bot. 2012;99(6):e240–4.
Han Y, Gao Y, Zhai X, Zhou H, Ding Q, Ma L. Assembly and comparative analysis of chloroplast genome of wheat K-CMS line and maintainer line. BMC Genom. 2020.
Guo H, Liu J, Luo L, Wei X, Zhang J, Qi Y, Zhang B, Liu H, Xiao P. Complete chloroplast genome sequences of Schisandra chinensis: genome structure, comparative analysis, and phylogenetic relationship of basal angiosperms. Sci China Life Sci. 2017;60:1286–90.
Cavalier-Smith T. Chloroplast Evolution: secondary Symbiogenesis and multiple losses. Curr Biol. 2002;12(2):R62–4.
Yang JB, Yang SX, Li HT, Yang J, Li DZ. Comparative chloroplast genomes of Camellia species. PLoS ONE. 2013;8(8):e73053.
Li HT, Yi TS, Gao LM, Ma PF, Zhang T, Yang JB, Gitzendanner MA, Fritsch PW, Cai J, Luo Y. Origin of angiosperms and the puzzle of the jurassic gap. Nat Plants. 2019;5(5):461–70.
Li X, Yang Y, Henry RJ, Rossetto M, Wang Y, Chen S. Plant DNA barcoding: from gene to genome. Biol Rev. 2015;90(1):157–66.
Zhang Z, Zhang Y, Song M, Guan Y, Ma X. Species identification of Dracaena using the complete chloroplast genome as a super-barcode. Front Pharmacol. 2019;10:1441.
Wang W, Messing J. High-throughput sequencing of three Lemnoideae (duckweeds) chloroplast genomes from total DNA. PLoS ONE. 2011;6(9):e24670.
Raubeson LA, Peery R, Chumley TW, Dziubek C, Fourcade HM, Boore JL, Jansen RK. Comparative chloroplast genomics: analyses including new sequences from the angiosperms Nuphar advena and Ranunculus macranthus. BMC Genomics. 2007;8(1):174.
Yang M, Zhang X, Liu G, Yin Y, Chen K, Yun Q, Zhao D, Al-Mssallem IS, Yu J. The complete chloroplast genome sequence of date palm (Phoenix dactylifera L). PLoS ONE. 2010;5(9):e12762.
Daniell H, Lin CS, Yu M, Chang WJ. Chloroplast genomes: diversity, evolution, and applications in genetic engineering. Genome Biol. 2016;17(1):1–29.
Logacheva MD, Krinitsina AA, Belenikin MS, Khafizov K, Konorov EA, Kuptsov SV, Speranskaya AS. Comparative analysis of inverted repeats of polypod fern (Polypodiales) plastomes reveals two hypervariable regions. BMC Plant Biol. 2017;17(2):61–73.
Germano J, Klein AS. Species-specific nuclear and chloroplast single nucleotide polymorphisms to distinguish Picea glauca, P. mariana and P. rubens. Theor Appl Genet. 1999;99(1):37–49.
Shi XR, Zhao HY, Gong B, Cao WS. Study on the Control Effect of Different Herbicide Treatments in Direct Water Rice Fields. Shanghai Agricultural Sci Technol 2013;(4):132–3. (in Chinese).
Zhang JL, Qiao JL, Meng ZH, Du JL, Li CG, Liu YW. Experiment on the control of weeds in Rice Transplant fields with 10% bensulfuron WP. Heilongjiang Agricultural Sci 2009;(2):76. (in Chinese).
Jin DJ, Chi XC, Gu XJ, Yang XY, Zhang YH, Fu Y. Efficacy and selectivity of 65% of MCPA-Dimethylamine Salt (AS) to Rice in transplanted Rice Fields. J Weed Sci. 2017;035(004):25–9. (in Chinese).
Acknowledgements
We thank Shanghai BIOZERON Biotechnology Co., Ltd. for performing the high-throughput sequencing.
Funding
This research was funded by Shanghai Agriculture Applied Technology Development Program, China (Grant number T20210104).
Author information
Authors and Affiliations
Contributions
Study design, Y.G.; concept, Y.G., Z.T., and G.S.; data collection, Y.G., T.C., and J.L.; analysis, Y.G., T.C., and J.L.; manuscript writing, Y.G; review, Z.T., and G.S.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The seeds of Aeschynomene indica collected in this study were approved by the landowner.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gao, Y., Chen, T., Long, J. et al. Complete chloroplast genome and comparison of herbicides toxicity on Aeschynomene indica (Leguminosae) in upland direct-seeding paddy field. BMC Genomics 25, 277 (2024). https://doi.org/10.1186/s12864-024-10102-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-024-10102-x