
Abstract
Background
Preingestive behavioral modulations of herbivorous insects on the host plant are abundant over insect taxa. Those behaviors are suspected to have functions such as deactivation of host plant defenses, nutrient accumulation, or modulating plant-mediated herbivore interactions. To understand the functional consequence of behavioral modulation of insect herbivore, we studied the girdling behavior of Phytoecia rufiventris Gautier (Lamiinae; Cerambycidae) on its host plant Erigeron annuus L. (Asteraceae) that is performed before endophytic oviposition in the stem.
Results
The girdling behavior significantly increased the larval performance in both field monitoring and lab experiment. The upper part of the girdled stem exhibited lack of jasmonic acid induction upon larval attack, lowered protease inhibitor activity, and accumulated sugars and amino acids in compared to non-girdled stem. The girdling behavior had no effect on the larval performance of a non-girdling longhorn beetle Agapanthia amurensis, which also feeds on the stem of E. annuus during larval phase. However, the girdling behavior decreased the preference of A. amurensis females for oviposition, which enabled P. rufiventris larvae to avoid competition with A. amurensis larvae.
Conclusions
In conclusion, the girdling behavior modulates plant physiology and morphology to provide a modulated food source for larva and hide it from the competitor. Our study implies that the insect behavior modulations can have multiple functions, providing insights into adaptation of insect behavior in context of plant-herbivore interaction.
Similar content being viewed by others

Background
Coevolutionary theories predict that plant defense and insect offense exert selective pressure that favors diversification of each other [1]. Central to understanding the arms race between plants and insect herbivores have been the phytochemistry and the molecular adaptation of insects to it [2, 3]. However, the chemical conflict occurs at the latest stage of herbivory, only after the ingestion occurs. Before ingestion, an interesting class of insect behavior is observed in many clades: behavioral modulation. Vein cutters cut the veins of the leaf, trenchers cut a line through the leaf, and girdlers chew around the petiole or stem [4]. These behavioral modulations are found in various insect taxa of Lepidoptera [5], Coleoptera [6, 7], Hemiptera [8], and Orthoptera [9]. The convergent evolution of similar behavioral modulations from distant taxa indicates it is an adaptation.
One well-characterized function of the behavioral modulation is the neutralization of plant defense, especially canal-borne exudates that are blocked by vein cutting and trenching [6, 10]. However, though behavioral modulation is abundantly conducted on plants without secretory canals, their functional consequences in plant-herbivore interaction is rarely tested. Particularly, only a few studies have proposed the potential role of the girdling behavior as exposure of vasculature to apply saliva [11] or nutrient accumulation at the girdled part [7, 8] and the downstream effects on insect performance are not known, raising questions on the functional consequence of girdling behavior.
Herbivorous insects largely depend on the quality of the host plants [12]; the dependency suggests that examination of the effect of the girdling behavior on the metabolic and defensive traits of plants is required to assess the functional consequences of the behavior. Insect attack has significant effects on the nutrient status of plants and the effect of modified plant chemistry on insect performance is diverse [13]. Moreover, insect herbivore faces plant defenses comprising unpalatable substances (e.g., secondary metabolites [14], proteinase inhibitor [15], and hardened cell walls [16]), and natural enemies of herbivores [17]. A substantial part of plant defense is activated by attack primarily through jasmonic acid (JA) signaling [18], which is referred induced defense.
Different herbivores feeding on the same plant interact with each other via plant traits including systemic responses [19] and altered plant appearance [10]. As behavioral modulation changes plant morphology [20] and palatability [21], it may alter the responses of other herbivores sharing the same host plant. Indeed, canal cutting promotes the feeding of other herbivores that originally do not feed on plants with secretory canals [21]. However, it is not known whether the effect of behavioral modulation on other herbivores are beneficial to the modulating insects.
To systematically assess the functional consequence of behavioral modulation of insect herbivores, we studied a girdling longhorn beetle Phytoecia rufiventris Gautier (Lamiinae; Cerambycidae) and a non-girdling longhorn beetle Agapanthia amurensis Kraatz (Lamiinae; Cerambycidae) which both oviposit on an introduced species Erigeron annuus L. (Asteraceae) in South Korea. The P. rufiventris female girdles around the stem of Asteraceae host plants including E. annuus before oviposition (Supplementary Video S1) while A. amurensis female lays an egg inside the stem without girdling. To determine the functional consequences of the girdling behavior using experimental girdling, we tested the following three hypotheses: (1) The girdling behavior of female P. rufiventris facilitates larval growth inside the girdled stem, (2) The girdling behavior impairs the plant resistance and enhances nutrients in the girdled stem, and (3) The appearance of the girdled stem is decreased to a competing herbivore.
Results
Two longhorn beetles sharing Erigeron annuus as the host plant show distinct natural histories
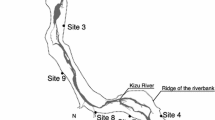
We observed the natural history of two Lamiinae longhorn beetles, Phytoecia rufiventris and Agapanthia amurensis, by monitoring Erigeron annuus (Asteraceae) as a shared host plant of two beetles in our field sites in Korea (Fig. 1, Supplementary Table S1). Phytoecia rufiventris laid eggs in E. annuus predominately while they also laid eggs in Artemisia princeps. Erigeron annuus bolted in April and P. rufiventris adults emerged from mid-April to July, girdled and laid eggs inside the stem of E. annuus plants. Every E. annuus stem with a feeding pattern at the lower part also had a feeding pattern at the upper part but not vice versa i.e., the larvae first fed on the upper part of the girdled stem and then move down beyond the girdles. In September, the larva cut the basal part of the stem and pupated inside the root-shoot junction. The adult overwintered inside the stem until subsequent April.

Natural history of Erigeron annuus and two longhorn beetles, Phytoecia rufiventris and Agapanthia amurensis. Adult emergence and oviposition timing in field observation sites are indicated as the adult. The larval feeding pattern is indicated as the larval position inside the stem
In contrast to P. rufiventris, A. amurensis overwintered as a larva and pupated during April. The adult A. amurensis thus emerged from May to July and laid eggs at the lower part of E. annuus stem without girdling. The hatched larva first moved and fed towards the shoot apex, then moved down to the lower part of the stem and cut in the middle of the dead stem. The larva stayed inside the stem until subsequent April.
Girdling behavior of Phytoecia rufiventris cuts all vascular bundles to induce cell death at the upper part of the E. annuus stem
To examine the functional consequence of girdling behavior of P. rufiventris, we first characterized the girdling behavior (Fig. 2a and b, Supplementary Video S1, approximately 6 min). Phytoecia rufiventris female girdles two arcs (lower and upper girdles) with about 1 cm apart. The female makes an oviposition cavity by chewing the epidermis and then lays an egg inside the stem. The upper part of the girdled E. annuus plant lost its turgor within an hour and showed dropping morphology (Fig. 2c, Supplementary Video S2, approximately 9 min). The girdles divided the stem into the upper (14.76 ± 4.07 cm) and lower part (50.72 ± 16.13 cm) respectively (N = 73 and 61, respectively). Though neither of the lower and upper girdles was a full circle, they together cut every vascular bundle (Fig. 2b). Two semicircle artificial girdles mimicked the natural girdling without fracture of the upper part (Supplementary Fig. S1). The upper part of the girdled stem underwent cell death within 1-week post girdling (wpg) while the lower part of the girdled stem remained viable (Fig. 2d). The cell death in the upper part of the girdled stem became more severe at 3 wpg, as shown by stringer Trypan-blue staining at the upper part of the girdled stem.

Observations on girdling behavior of Phytoecia rufiventris. (a) Behavioral sequence of girdling and oviposition of P. rufiventris. (b) Girdle angles of lower and upper girdling (N = 100). Box indicates 1st and 3rd quantiles. (c) Morphology of naturally girdled E. annuus. Two girdles are indicated with white triangles. (d) Trypan-blue staining of the experimentally girdled and non-girdled stem of E. annuus at 1 and 3 wpg. Blue staining indicates cell death
Larval performance of P. rufiventris is enhanced by girdling behavior
We found the girdled E. annuus occasionally reconnected the damaged vascular bundles and restored the turgor to the upper part in field observation. Phloroglucinol staining of the longitudinal section of experimentally girdled and recovered stem showed reconnection of the xylem in recovered E. annuus plants (Fig. 3a). In such recovered stems, larval survival of P. rufiventris was significantly lower than in successfully girdled E. annuus stem (Fig. 3a).

Effect of girdling behavior on the larval performance of P. rufiventris. (a) Larval survival rates in successfully girdled (N = 47) and recovered (N = 42) E. annuus plants measured in naturally girdled plants (* P < 0.05, Chi-squared test). The left images represent the morphology of girdled and recovered E. annuus plants while the right images represent Phloroglucinol staining of the longitudinal section of girdled and recovered part. (b) Larval mass measured from the experimentally girdled and non-girdled E. annuus plants. Boxes indicate the 1st and 3rd quantiles (significant differences are indicated as different letters; P < 0.05, One-way ANOVA followed by Tukey’s HSD)
We then mimicked the girdling behavior and inoculated P. rufiventris eggs in the E. annuus stem (Supplementary Fig. S1a). At three wpg, the larval mass was nearly five times higher in girdled plants than in non-girdled plants (Fig. 3b). As in the field monitoring, the experimentally girdled stems occasionally recovered (Supplementary Fig. S1b). The larvae in such recovered stems showed a similar mass to the larvae in non-girdled plants (Fig. 3b).
The upper part of the girdled stem is modulated into a better food source
As in our field observation, the larva preferentially fed on the upper part during the first week after egg inoculation whereas larvae in non-girdled stems showed no feeding preference (Fig. 4a). Moreover, larval survival and mass were not facilitated when the upper part of the stem was removed by decapitation (Fig. 4b and c). Thus, we suspected that the upper part of the girdled E. annuus stem is modulated into a better food source for the larva.

The upper part of the E. annuus plant is modulated by girdling behavior. (a) Feeding patterns of P. rufiventris larva between the upper and lower parts of the stem one week after egg inoculation. Empty dots indicate the upper limit of the feeding pattern, whereas filled dots indicate the lower limit of the feeding pattern. Boxes indicate the 1st and 3rd quantiles. Statistic comparisons were performed between the upper and lower limits in the same treatment groups. (n.s., no significant difference; *** P < 0.001, Student’s T-test). (b-c) Larval performance in decapitated plants, with (b) larval mass and (n.s., no significant difference, Chi-squared test) (c) larval survival rates (n.s., no significant difference, Student’s T-test). (d-e) Jasmonic acid levels were measured with and without experimental girdling and egg inoculation at (d) the upper part and € the lower part of the stem. (Significant differences are indicated as different letters; P < 0.05, One-way ANOVA followed by Tukey’s HSD). (f) Normalized % proteinase inhibition in experimentally girdled and non-girdled stems (significant differences are indicated as different alphabet letters; P < 0.05, Kruskal-Wallis’ test followed by Dunn’s test). (g) Metabolic profiles of the upper and lower stems of E. annuus with and without experimental girdling. Metabolites that accumulated only at the upper part of the girdled stem are indicated with the red box and their compound names (P < 0.05, One-way ANOVA followed by Tukey’s HSD). The abundance values were log-transformed for plotting. GUS, girdled upper stem; GLS, girdled lower stem; CUS, control upper stem; CLS, control lower stem
To understand the mechanistic background of the enhanced larval performance in girdled plants, we compared phytohormonal responses of the upper and lower part of experimentally girdled and non-girdled plants. First, we found that while the upper part of non-girdled plants showed increased JA levels upon larval attack, the upper part of girdled plants was not able to increase JA levels during the first week of larval feeding (Fig. 4d). This impaired defense in the girdled E. annuus was only present in the upper part of the stem while the lower parts of the stem showed increased JA levels regardless of experimental girdling, 3 weeks after the egg inoculation (Fig. 4e). In contrast, decapitated stems showed increased JA levels upon larval attack (Supplementary Fig. S2), highlighting importance of the upper apart of the girdled stem in facilitating larval growth. While salicylic acid (SA) levels at the upper part of the stem were not affected by both experimental girdling and egg inoculation (Supplementary Fig. S3a), abscisic acid (ABA) levels were significantly increased by experimental girdling in the same region (Supplementary Fig. S3b). Egg inoculation had no significant effect on ABA levels in both girdled and non-girdled E. annuus plants.
Additionally, we measured the proteinase inhibitor activity of girdled and non-girdled stems to assess the palatability of the girdled tissues (Fig. 4f). The proteinase inhibitor activity was not induced by the larval attack but it was reduced by experimental girdling. The protease inhibitor activity was marginally restored in girdled E. annuus stem by larval attack but the activity level was not significantly different from not attacked and girdled E. annuus stem.
We then tested the nutritional value of the girdled stem by sampling the upper and lower parts of stems with and without experimental girdling 2 days after experimental girdling, when the larval feeding starts. Primary metabolites were analyzed using gas chromatography–mass spectrometry (GC-MS) (Supplementary Table S2). Principal component analysis (PCA) of the 40 identified metabolites showed that the metabolomes of the upper parts were largely altered by experimental girdling, while the lower parts were only affected to a modest degree (Supplementary Fig. S4). We identified nine metabolites that were significantly accumulated in the upper part of the experimentally girdled stems while not accumulated in the upper part of the non-girdled stems (Fig. 4g): palmitic acid, glycerol, valine, proline, sucrose, turanose, galactose, fructose, and arabinose. The accumulation of soluble sugar in the upper part of girdled stems was confirmed by an anthrone assay (Supplementary Fig. S5).
Subsequently, we tested whether the accumulated nutrient sugars and amino acids in the upper part of the stems are sufficient to facilitate larval growth. We reared P. rufiventris larvae on semi-artificial diets made of non-girdled stem powder supplemented with sugars and amino acids, corresponding to the previous measurements. The three most abundant sugars (fructose, sucrose, and galactose) and two amino acids (proline and valine) in the upper part of the girdled stems were selected. Unexpectedly, the larval relative growth rate was not significantly altered by sugar and/or amino acid supplementation, even with the double amount of observed difference between the girdled and non-girdled upper part of the stem (Fig. 4h, Supplementary Fig. S6).
Non-girdling Agapanthia amurensis does not benefit from girdling behavior
With our experimental evidence on the adaptive value of girdling behavior, we tested whether girdling behavior provides general benefits to a non-girdling longhorn beetle, A. amurensis. Unlike in P. rufiventris, experimental girdling did not significantly facilitate larval growth of A. amurensis, regardless of the presence of P. rufiventris larva in the same stem (Fig. 5a).

Effect of girdling behavior on Agapanthia amurensis. (a) Larval mass of A. amurensis measured from the egg inoculation experiment. Boxes indicate the 1st and 3rd quantiles (n.s., no significant differences). (b) Number of putative detoxification genes identified from de novo assembled transcriptome of P. rufiventris and A. amurensis (CYP450; Cytochrome P450, UGT; UDP-glucosyltransferase, GST; Glutathione-S-transferase, ABC transporter; ATP-binding cassette transporter, COE; Carboxylesterase). (c) Experimental scheme of the co-infestation assay. The scientific names of insects indicate egg inoculation. (d) Larval survival of P. rufiventris and A. amurensis larvae in co-infested E. annuus stem. Treatments for each group are indicated in Fig. 5c (e-f) Oviposition preference of A. amurensis on girdled and non-girdled E. annuus in (e) field monitoring (***; P < 0.001, Chi-squared test) and (f) choice assay (**; P < 0.01, Fisher’s exact test). The field-girdled E. annuus contained P. rufiventris egg inside the stem while experimentally girdled E. annuus stem did not
The asymmetry in the effect of girdling on two species led us to speculate about the differential chemical adaptation of two species to plant metabolites. To test this idea, we investigated the detoxification capacity of two species at the transcriptomic level. We used de novo assembled gut transcriptome of two species to extract putative detoxification genes of five families: Cytochrome P450s (CYP450s), UDP-glycosyltransferases (UGTs), Glutathione-S-Transferases (GSTs), Carboxylesterases (COEs), and ABC transporters. Interestingly, the number of putative detoxification genes was higher in the assembled transcriptome of A. amurensis than that of P. rufiventris for all 5 families of genes investigated (Fig. 5b).
Phytoecia rufiventris avoids competition with Agapanthia amurensis via the decreased appearance of girdled stem
We observed co-infestation of P. rufiventris and A. amurensis on same E. annuus plants in the field monitoring. Notably, A. amurensis larvae mostly persisted in stems of such plants while P. rufiventris larvae were dead (Fig. 5c and d). To test whether competition is sufficient to decrease the larval survival of P. rufiventris, we conducted a co-infestation assay (Fig. 5c). Regardless of experimental girdling, in most stems where two longhorn beetles were inoculated together, A. amurensis larvae showed priority (Fig. 5d).
To test whether the girdled E. annuus plants are hidden from the oviposition choice of A. amurensis, we measured the oviposition rate of A. amurensis on naturally girdled and non-girdled E. annuus plants in field sites. The proportion of plants chosen for A. amurensis oviposition was higher in non-girdled E. annuus plants than in girdled plants (Fig. 5e). To confirm the decreased appearance of girdled E. annuus plants to A. amurensis, we conducted an oviposition choice assay of A. amurensis on girdled and non-girdled E. annuus plants. The female A. amurensis significantly preferred non-girdled E. annuus plants for oviposition over girdled E. annuus plants (Fig. 5f).
Discussion
Natural history observations on behavioral modulations suggest various functions of behavioral modulations. As most behavioral modulations damage the vascular bundles [9], the subsequent effects have been predicted to be the inhibition of defense signaling [20] and accumulation of nutrients [22, 23]. Moreover, the modified morphology of modulated plants is suspected to be utilized for evasion of natural enemies [20]. However, other than disarming defense mediated by canal-borne exudates, experimental testing on functional consequences of behavioral modulations at the molecular level has been seldom compared to copious hypotheses.
In consistent to the cell death observed at the upper part of the girdled stem, JA induction and proteinase inhibitor activity were decreased at the region. Inhibition of JA signaling is strong evidence of inhibited induced defense against chewing herbivores [24]. Although the inhibition of defenses was restricted to the upper part of the stem, the girdling-dependent feeding preference on the upper part enabled the larvae to selectively consume the modulated food source during the early stages. The susceptible region provided could boost the growth of vulnerable early larval stages and face the induced defense of the lower part of the stem with a better capacity to tolerate plant defense. The inhibition of plant defense at the physiological level is achieved by various insect strategies including symbionts [25] and effectors [26]. We propose that the plant tissue which is freshly killed by the girdling behavior also exhibit inhibited defense.
In addition to deactivated defenses, accumulation of nutrients also occurred in the upper part of the girdled stem, as in girdling of alfalfa hopper and twig girdler [7, 8]. However, the nutrient, especially soluble sugar, can be negatively correlated with insect performance [13]. In our study, the nutrient supplementation in an artificial diet had no significant effect solely on larval growth. Nevertheless, the joint effect of accumulated nutrients and impaired defense in girdled E. annuus stem remains to be studied.
Interestingly, experimental girdling had no significant effect on the growth of A. amurensis larvae. In studies performed using generalists to test the effect of trenching, the behavioral modulation was a prerequisite for feeding but not always sufficient [27]. As A. amurensis is capable of feeding on E. annuus without girdling behavior, possible explanation are that A. amurensis may not induce a strong defense response in E. annuus or cope well with the defenses. To test the latter possibility, we mined the transcriptomes of P. rufiventris and A. amurensis and compared the number of putative detoxification genes, as the gene duplication events are thought to be favored by pressure exerted by plant substances [28, 29]. We interrogated the enzymes that are generally but not exclusively associated to plant metabolite detoxification as the specialized detoxification mechanisms of P. rufiventris and A. amurensis are not known. Insects detoxify toxic metabolites by cleavage (e.g., Carboxylesterase; COE [30]), oxidation (e.g., Cytochrome P450; CYP450 [31]), and the addition of moiety (e.g., Glutathione-S-Transferase; GST [32], UDP-Glycolic Transferase; UGT [33]). Moreover, plant metabolites are rapidly transported by transporters such as ATP-binding cassette (ABC) transporters [34]. The higher putative detoxification gene numbers of A. amurensis in compared to P. rufiventris might be associated with their ability to digest plant metabolites.
Nevertheless, our current analyses do not include identification of bioactive resistive compound produced by E. annuus and deactivated by the girdling behavior, leaving an interesting topic for further study. Erigeron annuus synthesizes a variety of secondary metabolites including terpenoids, flavonoids, organic acid glycosides, and polyacetylenes [35,36,37,38]. The bioactivities of plant secondary metabolites are highly context-dependent; indeed, untargeted metabolomics coupled with bioassays are further required to the chemical interaction between E. annuus and P. rufiventris, along with the effect of behavioral modulation on plant secondary metabolism.
The drooping stem is a widespread phenotype in herbaceous plants that decreases appearance to herbivores [20]. The girdling-induced drooping of E. annuus decreased the appearance to A. amurensis which is a harmful competitor of P. rufiventris. Although co-infestation of multiple herbivores on a plant can be beneficial [39] or neutral [40] for herbivores, restricted unidirectional movement of larvae inside the stem causes unavoidable competition between P. rufiventris and A. amurensis. This indirect effect of girdling behavior, evading a competitor, is a novel function of behavioral modulation, which we propose is an exaptation of counteradaptation to plant defense. Rodent herbivores also modulate the physical properties of plants to decrease their appearance, implying that disguising through plant modulation has a general adaptive value across animals [41].
Conclusion
In conclusion, we demonstrated that the girdling behavior of P. rufiventris decreases host plant defense and increases nutrients. The girdling behavior also decreases the risk of harmful competition with A. amurensis via the lowered appearance of the girdled plants. Insect herbivores have developed various array of offensive strategies to deal with plant defense [42]. We conclude that the behavioral modulation on plants can have manipulating effects on host plant physiology and morphology, which clearly are adaptive for insect herbivores.
Methods
Plant growth and insect rearing
Adult Phytoecia rufiventris beetles were collected from our field sites (Supplementary Table S1) between April and July 2019–2022. The adult beetles were maintained at 25–27 °C in transparent acrylic cages (40 × 40 × 40 cm3) with Erigeron annuus plants under long-day (16 L:8D) conditions. Eggs were collected every three days and inoculated inside the E. annuus stem following the previously described method [43] with experimental girdling. The larvae were maintained in E. annuus stems in the lab until the basal part of the stem was cut by the larvae, then the larvae were collected from the stems and moved to the short-day condition (12 L:12D) at 25 °C to induce pupation [44]. To break adult sexual diapause, the adults were placed at 0 °C for 16 weeks.
Adult A. amurensis adults were collected from the field sites (Supplementary Table S1) from May to July 2019–2022 and reared under the same conditions as those of P. rufiventris. The collected eggs were inoculated inside the E. annuus stem without experimental girdling and reared inside the stem until the stem is cut by larvae then collected and placed at 0 °C for 16 weeks to induce pupation.
We collected E. annuus seeds from a single individual at the Daejeon_1 site (Supplementary Table S1) in July 2019 and dried them with silica gel for 3 weeks. Seeds were germinated a 9 cm x 9 cm x 9 cm pot and were grown under long-day conditions (16 L:8D), 26 °C.
Field monitoring
Field monitoring was conducted from 2019 to 2022 at field sites in Daejeon, Cheongju, and Jeungpyeong, Korea (Supplementary Table S1). The angles of the girdled scars were measured by visually classifying the angle of the girdle arc by 45°. The positions of the girdles were measured from the ground to the lower girdle and from the shoot apical meristem to the upper girdle. The survival of P. rufiventris larvae in successfully girdled and recovered E. annuus stems was investigated. The oviposition of A. amurensis on girdled and non-girdled E. annuus plants was measured by checking the oviposition cavity. The larval survival of P. rufiventris in girdled E. annuus, with and without A. amurensis competition was measured by randomly dissecting naturally girdled E. annuus plants.
Egg inoculation and experimental girdling
Experimental girdling was performed by cutting the vascular bundles of E. annuus stems into two lines 1 cm apart, each forming a semicircle. Decapitation was performed by cutting the stem at the corresponding height. Phytoecia rufiventris eggs were inoculated between two girdles by digging a scar with a needle and gently inserting it with forceps, as described previously [43]. The egg inoculation was performed right below the cut stem in decapitated E. annuus plants. Experimental girdling and egg inoculation were performed 8 weeks after the germination of E. annuus. We sampled the upper part of the girdled stem one wpg (weeks post girdling) while the lower part was sampled three wpg, when the actual larval attack was occurring at each part. To enable sufficient growth of larvae for measurement, larval growth was measured in three wpg.
Trypan blue staining
The E. annuus stems were longitudinally cut, stained in 0.4% Trypan blue (aq) for 5 min, and destained with 100% ethanol overnight.
Primary metabolites and total soluble sugar measurement
The upper and lower parts (each 15 cm) of the girdled stem were collected 2 days after experimental girdling. Samples were freeze-dried and ground by mortar and pestle.
The metabolites in each sample were analyzed as described previously [45]. Briefly, 10 mg of each freeze-dried sample was used for extraction with 1 ml of methanol and the solvent was evaporated by applying a nitrogen flow rate of 0.625 L/min for 5 min. To induce oximation, 30 µL of 20 mg/mL methoxyamine hydrochloride in pyridine, 50 µL of BSTFA (N, O-Bis (trimethylsilyl)trifluoroacetamide) with 1% TMCS (chlorotrimethylsilane), 30 µL of 300 µg/mL 2-chloronaphtalene in pyridine was added. 10 µL of 300 µg/mL 2-chloronaphtalene in pyridine for internal standards, was added. The samples were incubated at 65 °C for 60 min. One microliter of the 1/100 diluted sample was injected into the Rtx-5MS column at an injection temperature of 250 °C. A GCMS-QP2020 system (Shimadzu, Kyoto, Japan) was used for analysis.
The total amount of soluble sugar was measured following previously described methods [46,47,48] with modifications. 5 mg of freeze-dried sample was used. Disrupting pigments were discarded using 100% acetone and soluble sugar was extracted using 0.5 mL of 80% ethanol. 0.5 mL of each sugar extract was reacted with 0.5 mL of ice-cold anthrone (Daejung, Korea) in 72% sulfuric acid (2 mg/mL) at 100 °C for 11 min, and the absorbance at 630 nm was measured.
Artificial Diet
The semi-artificial diet was prepared by 3.33% agar solution with 5% freeze-dried E. annuus stem powder. The total amount of supplemented sugars and amino acids were determined by the observed difference between girdled upper part and non-girdled upper part of the stem using GC-MS. In addition, diets comprising double the amounts of supplements were prepared. The composition of supplemented nutrients in the diet was determined according to the data acquired from GC-MS (Fig. 4g): 37.97 mg/g galactose, 2.93 mg/g sucrose, 13.45 mg/g fructose, 0.11 mg/g valine, and 1.10 mg/g proline. Each larva was posited in an Eppendorf tube with a block of artificial diet. Every 3 days, larval mass was measured and a fresh diet was provided.
Phytohormone quantification
The upper and lower parts (each 15 cm) of the girdled stem were collected one wpg and three wpg, respectively, when the larval feeding at each part was occurring. For the decapitated stems, only lower parts were sampled. To exclude bias originating from different water contents, the upper part samples were freeze-dried. Phytohormones were quantified according to the previously described method [49], with modifications [16]. One milliliter of ethyl acetate spiked with internal standards (20 ng each of d4-salicylic acid, d5-jasmonic acid, and d6-abscisic acid) was added to 10 mg of each sample. The solvent was evaporated using a centrifugal vacuum concentrator VC2124 (LaboGene, South Korea) at 30 °C and resuspended in 70% methanol and centrifuged at 13,000 rpm for 10 min. 200 µL of the supernatant was collected.
The phytohormones were measured using LC-MS. The extract was injected into a C18 column (UPLC BEH, 1.7 μm particle size, 100 mm length × 2.1 mm inner diameter, Waters, Ireland) and separated using a high-performance liquid chromatography system (Shimadzu, LCMS-8050). Solvent A consisted of deionized water containing 0.1% (v/v) acetonitrile and 0.05% formic acid. Solvent B consisted of 100% MeOH. The following gradient conditions were used for chromatography: 0.01–0.5 min 5% B; 0.6–6.7 min linear gradient 50–95% B; 6.7–8.7 min 95% B; and re-equilibration at 5% B for 1 min. The flow rate was set at 400 µL/min. The mass spectrometry conditions were as follows: capillary temperature, 300 °C; capillary voltage, 3.0 kV.
Protease inhibitor assay
The upper lower part (15 cm) of the girdled stem was collected one wpg for protease inhibitor activity measurement. The procedure used here was modified from previous studies [50, 51]. Protein extraction was performed by adding 180 µL of extraction buffer (50 mM phosphate pH 7.2, 150 mM NaCl, and 2.0 mM EDTA) to 45 mg of each sample.
The protease activity of papain with the extracted protein was measured for each sample. The papain solution was prepared as 50 µL/mL papain in 25 mM sodium phosphate buffer (pH 7.0). 0.05 mL of papain solution, 0.1 mL of protein extract, and 0.1 mL of incubation buffer (0.25 M sodium phosphate buffer, pH 6.0, 2.5 mM EDTA, and 25 mM 2-mercaptoethanol) were mixed. After 5 min of incubation at 37 °C, the protease reaction was initiated by adding 0.1 mL of 1 mM Nα-benzoyl-DL-arginine β-naphthylamide hydrochloride (BANA) solution as the substrate. The reaction was terminated by adding 0.5 mL of 2% HCl in ethanol after 10 min of incubation at 37 °C. For color development, 0.5 mL of 0.06% p-dimethylaminocinnamaldehyde/ethanol was added. The resulting absorbance was measured at 540 nm as an indicator of protease activity and normalized by the fresh mass of the samples and the amount of total protein, which was measured using the BCA assay (Thermo Scientific kit 23,227, Waltham, MA, USA).
Larval gut RNA extraction and RNA-sequencing
Larval gut RNA was extracted from P. rufiventris and A. amurensis. Three, one, and three biological replicates were sequenced for P. rufiventris reared in girdled stems, ungirdled stems, and A. amurensis larvae, respectively. The larvae were sterilized with 70% ethanol, rinsed with distilled water, and ice-cooled. The midguts were collected, rinsed with Ringer’s solution, and frozen in liquid nitrogen for grinding by pestle. The RNA was extracted using RNeasy Micro Kit (QIAGEN, Germany) and purified by RNeasy MinElute Cleanup Kit (QIAGEN). The library preparation was conducted using TruSeq Stranded mRNA LT Sample Prep Kit. Illumina paired-end 151 bp platform was used for sequencing.
De novo transcriptome assembly
The quality of the raw reads was assessed using FastQC [52] (v. 0.11.8), and the reads were trimmed using Trimmomatic [53] (v 0.38). To remove contamination of plant-driven RNA, Kraken2 [54] (v. 2.1.2) was used to classify reads belonging to the plantdb. Processed reads were assembled using Trinity [55] (v. 2.10.0). Open reading frames were predicted using TransDecoder (v. 5.5.0; https://github.com/TransDecoder/TransDecoder) and the extracted sequences were annotated using Trinotate [56] (v. 3.2.2).
Gene search
The Pfam annotations in Trinotate was used to initially search putative CYP450s, UGTs, COEs, GSTs, and ABC transporters in de novo assembled transcriptome of P. rufiventris and A. amurensis. The blastx search to SWISS-PROT was used to confirm predicted genes.
Agapanthia amurensis choice assay
The oviposition preference of Agapanthia amurensis for girdled and non-girdled E. annuus plants was investigated. One week after experimental girdling, two girdled and two non-girdled E. annuus plants were placed symmetrically in a tent (47 × 47 × 91 cm3) in an open field site at Cheongju_1 (Supplementary Table S1). Each plant was placed 30 cm from the release point of a female A. amurensis. Each female was used for the choice assay within one month post emergence. E. annuus plants were exchanged once an oviposition choice was made and each choice was recorded.
Co-infestation assay
The effect of the A. amurensis larva on P. rufiventris larva was investigated by inoculating an egg of A. amurensis into the bottom part of the E. annuus stem 3 d after P. rufiventris egg inoculation. Four weeks after egg inoculation, E. annuus stems were dissected to measure the survival rate of two larvae.
Statistical analysis
All statistical analyses were conducted using the R software [57] (v. 4.0.5). The normality of raw data was checked using the Shapiro-Wilk test and similar variances between groups were tested using Levene’s test. For parametric tests, Student’s t-test and one-way ANOVA followed by Tukey’s honestly significant difference (HSD) test were used. For non-parametric tests, the Mann-Whitney U test and Kruskal-Wallis’s test followed by Dunn’s test were used.
(h) Relative growth rates (RGR) of P. rufiventris larva at 12 days reared with artificial diets containing supplementary sugars and amino acids (n.s., no significant differences, One-way ANOVA).
Data availability
The raw sequence files are deposited at NCBI (Bioproject: PRJNA1008891). All data supporting the results are available at Dryad (https://datadryad.org/stash/share/IyxFJF58UWR6tAdwLnlvsSqD4e4tC0UCS4IpXrv_wRY).
Code availability
Codes used for bioinformatic analyses are deposited in Github (https://github.com/Phytoecia/Girdling_transcriptomics).
Abbreviations
- JA:
-
jasmonic acid
- GO:
-
gene ontology
- GC-MS:
-
gas chromatography-mass spectrometry
- CYP450:
-
cytochrome P450
- UGT:
-
UDP-glycosyltransferase
- GST:
-
glutathione-S-transferase
- COE:
-
carboxylesterase
References
Futuyma DJ, Agrawal AA. Macroevolution and the biological diversity of Plants and Herbivores. Proceedings of the National Academy of Sciences. 2009;106:18054–61.
Ehrlich PR, Raven PH. Butterflies and plants: a study in coevolution. Evol (N Y). 1964;18:586–608.
Heckel DG. Insect Detoxification and Sequestration Strategies. 2014.
Dussourd DE. Behavioral sabotage of plant defenses by insect folivores. Annu Rev Entomol. 2016;62:15–34.
Clarke AR, Zalucki MP. Foraging and vein-cutting behaviour of Euploea core corinna (W. S. Macleay) (Lepidoptera: Nymphalidae) caterpillars feeding on latex-bearing leaves. Aust J Entomol. 2000;39:283–90.
Dussourd DE, Eisner T. Vein-cutting behavior: Insect counterploy to the latex defense of plants. Science (1979). 1987;237:898–901.
Forcella F. Why twig-girdling beetles girdle twigs. Naturwissenschaften. 1982;69:398–400.
Andersen PC, Brodbeck Bv, Herzog DC. Girdling-induced nutrient accumulation in above ground tissue of peanuts and subsequent feeding by Spissistilus festinus, the three-cornered alfalfa hopper. Entomol Exp Appl. 2002;103:139–49.
Dussourd DE, Denno RF. Deactivation of Plant Defense: Correspondence between Insect Behavior and Secretory Canal Architecture. J Ecol. 1991;72:1383–96.
Hu X, Su S, Liu Q, Jiao Y, Peng Y, Li Y, et al. Caterpillar-induced rice volatiles provide enemy-free space for the offspring of the brown planthopper. Elife. 2020;9:1–19.
Ganong CN, Dussourd DE, Swanson JD. Girdling by notodontid caterpillars: distribution and occurrence. Arthropod Plant Interact. 2012;6:621–33.
Awmack CS, Leather SR. Host plant quality and fecundity in herbivorous insects. Annu Rev Entomol. 2001.
Machado RAR, Arce CCM, Ferrieri AP, Baldwin IT, Erb M. Jasmonate-dependent depletion of soluble sugars compromises plant resistance to Manduca sexta. New Phytol. 2015;207:91–105.
Steppuhn A, Gase K, Krock B, Halitschke R, Baldwin IT. Nicotine’s defensive function in nature. PLoS Biol. 2004;2.
Hartl M, Giri AP, Kaur H, Baldwin IT. Serine protease inhibitors specifically defend Solanum nigrum against generalist herbivores but do not influence plant growth and development. Plant Cell. 2012;22:4158–75.
Joo Y, Kim H, Kang M, Lee G, Choung S, Kaur H, et al. Pith-specific lignification in Nicotiana attenuata as a defense against a stem‐boring herbivore. New Phytol. 2021. https://doi.org/10.1111/nph.17583.
Dicke M. Local and systemic production of volatile herbivore-induced terpenoids: their role in plant-Carnivore mutualism. J Plant Physiol. 1994;143:465–72.
Schuman MC, Baldwin IT. The layers of plant responses to insect herbivores. Annu Rev Entomol. 2015;61:373–94.
Heil M, Ton J. Long-distance signalling in plant defence. Trends Plant Sci. 2008;13:264–72.
Yamazaki K. The role of ducking stems in plant anti-herbivore defense and its exploitation by herbivorous insects. J Plant Interact. 2012;7:283–9.
Dussourd DE, Denno RF. Host range of Generalist caterpillars: trenching permits feeding on plants with secretory canals. Ecology. 1994;75:69–78.
Chen Y, Duan J, Yang S, Yang E, Jiang Y. Effect of girdling on levels of catechins in fresh leaf in relation to quality of Huang Zhi Xiang Oolong Tea. Plant Foods Hum Nutr. 2009;64:293–6.
Delaney KJ, Higley LG. An insect countermeasure impacts plant physiology: Midrib vein cutting, defoliation and leaf photosynthesis. Plant Cell Environ. 2006;29:1245–58.
Erb M, Reymond P. Molecular interactions between plants and insect herbivores. Annu Rev Plant Biol. 2019;70:annurev–arplant.
Hansen AK, Moran NA. The impact of microbial symbionts on host plant utilization by herbivorous insects. Mol Ecol. 2014;23:1473–96.
Elzinga DA, De Vos M, Jander G. Suppression of plant defenses by a Myzus persicae (Green peach aphid) salivary effector protein. Mol Plant-Microbe Interraction. 2014;27:747–56.
Tune R, Dussourd DE. Specialized generalists: constraints on host range in some plusiine caterpillars. Oecologia. 2000;123:543–9.
Edger PP, Heidel-Fischer HM, Bekaert M, Rota J, Glöckner G, Platts AE, et al. The butterfly plant arms-race escalated by gene and genome duplications. Proc Natl Acad Sci U S A. 2015;112:8362–6.
Calla B, Noble K, Johnson RM, Walden KKO, Schuler MA, Robertson HM, et al. Cytochrome P450 diversification and hostplant utilization patterns in specialist and generalist moths: birth, death and adaptation. Mol Ecol. 2017;26:6021–35.
Jackson CJ, Liu JW, Carr PD, Younus F, Coppin C, Meirelles T, et al. Structure and function of an insect α-carboxylesterase (αesterase7) associated with insecticide resistance. Proc Natl Acad Sci U S A. 2013;110:10177–82.
Zhu F, Moural TW, Nelson DR, Palli SR. A specialist herbivore pest adaptation to xenobiotics through up-regulation of multiple cytochrome P450s. Sci Rep. 2016;6:1–10.
Sun XQ, Zhang MX, Yu JY, Jin Y, Ling B, Du JP et al. Glutathione S-Transferase of Brown Planthoppers (Nilaparvata lugens) is essential for their adaptation to gramine-containing host plants. PLoS ONE. 2013;8.
Wouters FC, Reichelt M, Glauser G, Bauer E, Erb M, Gershenzon J, et al. Reglucosylation of the Benzoxazinoid DIMBOA with Inversion of Stereochemical Config.uration is a detoxification strategy in Lepidopteran herbivores. Angewandte Chemie - Int Ed. 2014;53:11320–4.
Strauss AS, Peters S, Boland W, Burse A. ABC transporter functions as a pacemaker for sequestration of plant glucosides in leaf beetles. Elife. 2013;2013.
Iijima T, Yaoita Y, Kikuchi M. Five new sesquiterpenoids and a new diterpenoid from Erigeron annuus (L.) Pers., Erigeron philadelphicus L. and Erigeron Sumatrensis Retz. Chem Pharm Bull (Tokyo). 2003;51:545–9.
Nazaruk J, Kalemba D. Chemical composition of the essential oils from the roots of Erigeron acris L. and Erigeron annuus (L.) Pers. Molecules. 2009;14:2458–65.
Li X, Pan J, Gao K. γ-pyranone derivatives and other constituents from Erigeron annuus. Pharmazie. 2006;61:474–7.
Jang DS, Yoo NH, Kim NH, Lee YM, Kim CS, Kim J, et al. 3,5-Di-O-caffeoyl-epi-quinic acid from the leaves and stems of Erigeron annuus inhibits protein glycation, aldose reductase, and cataractogenesis. Biol Pharm Bull. 2010;33:329–33.
Soler R, Badenes-Pérez FR, Broekgaarden C, Zheng SJ, David A, Boland W, et al. Plant-mediated facilitation between a leaf-feeding and a phloem-feeding insect in a brassicaceous plant: from insect performance to gene transcription. Funct Ecol. 2012;26:156–66.
Lee G, Joo Y, Kim SG, Baldwin IT. What happens in the pith stays in the pith: tissue-localized defense responses facilitate chemical niche differentiation between two spatially separated herbivores. Plant J. 2017;92:414–25.
Zhong Z, Li G, Sanders D, Wang D, Holt RD, Zhang Z. A rodent herbivore reduces its predation risk through ecosystem engineering. Curr Biol. 2022;:1–6.
Karban R, Agrawal AA. Herbivore Offense. Annu Rev Ecol Syst. 2002;33:641–64.
Lee G, Joo Y, Diezel C, Lee EJ, Baldwin IT, Kim SG. Trichobaris weevils distinguish amongst toxic host plants by sensing volatiles that do not affect larval performance. Mol Ecol. 2016;25:3509–19.
Shintani Y. Quantitative short-day photoperiodic response in larval development and its adaptive significance in an adult-overwintering cerambycid beetle, Phytoecia Rufiventris. J Insect Physiol. 2011;57:1053–9.
Suh HW, Hyun SH, Kim SH, Lee SY, Choi HK. Metabolic profiling and enhanced production of phytosterols by elicitation with methyl jasmonate and silver nitrate in whole plant cultures of Lemna paucicostata. Process Biochem. 2013;48:1581–6.
Marshall JD. Drought and shade interact to cause fine-root mortality in Douglas-fir seedlings. Plant Soil. 1986;91:51–60.
Oren R, Schulze ED, Werk KS, Meyer J, Schneider BU, Heilmeier H. Performance of two Picea abies (L.) Karst. Stands at different stages of decline - I. Carbon relations and stand growth. Oecologia. 1988;75:25–37.
Hansen J, Møller I. Percolation of starch and soluble carbohydrates from plant tissue for quantitative determination with anthrone. Anal Biochem. 1975;68:87–94.
Schäfer M, Brütting C, Baldwin IT, Kallenbach M. High-throughput quantification of more than 100 primary- and secondary-metabolites, and phytohormones by a single solid-phase extraction based sample preparation with analysis by UHPLC-HESI-MS/MS. Plant Methods. 2016;12.
Abe M, Abe K, Iwabbuch K, Domoto C, Arai S. Corn cystatin I expressed in Escherichia coli: investigation of its inhibitory profile and occurrence in Corn Kernels. J Biochem. 1994;116:488–92.
Zhao Y, Botella MA, Subramanian L, Niu X, Nielsen SS, Bressan RA, et al. Two wound-inducible soybean cysteine proteinase inhibitors have Greater Insect Digestive proteinase inhibitory activities than a constitutive homolog. Plant Physiol. 2002;111:1299–306.
Andrews S, FastQC:. A Quality Control Tool for High Throughput Sequence Data [Online].
Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30:2114–20.
Lu J, Rincon N, Wood DE, Breitwieser FP, Pockrandt C, Langmead B, et al. Metagenome analysis using the Kraken software suite. Nat Protoc. 2022;17:2815–39.
Grabherr MG, Haas BJ, Yassour M, Levin JZ, Thompson DA, Amit I, et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat Biotechnol. 2011;29:644–52.
Haas BJ, Papanicolaou A, Yassour M, Grabherr M, Blood PD, Bowden J, et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat Protoc. 2013;8:1494–512.
R Core Team. R: A language and environment for statistical computing. 2022.
Acknowledgements
We are grateful to Dr. Gisuk Lee for valuable discussions and field monitoring. We appreciate advices from Dr. Ian T. Baldwin regarding significance of the study.
Funding
National Research Foundation of Korea (NRF) funds this work through the Basic Research Lab Program [NRF-2022R1A4A3024451] and Basic Science Research Program [NRF-2021R1C1C1006907], which are funded by the Ministry of Science and Technology. This work was supported by the New Faculty Startup Fund from Seoul National University. This work was supported by the research grant of the Chungbuk National University in 2020.
Author information
Authors and Affiliations
Contributions
MSC, JL, JMK, SGK, and YJ designed the study. MSC, JL, JMK, and YJ performed experiments. MSC analyzed the data. MSC, SGK, and YJ wrote the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
As all insects were collected from public field sites, no permission for collection was required throughout the study.
Consent of publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Choi, MS., Lee, J., Kim, JM. et al. Girdling behavior of the longhorn beetle modulates the host plant to enhance larval performance. BMC Ecol Evo 24, 49 (2024). https://doi.org/10.1186/s12862-024-02228-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12862-024-02228-z