
Abstract
Analysis of insects can provide evidence in death cases, for example, by answering the question about the time of death. Apart from flies, beetles are the second most useful insect group in forensic entomology. To elucidate the time of death based on insect evidence, developmental models of a given species are necessary. In this study, we developed such models for Necrodes littoralis, a necrophagous beetle, which is common in the Palearctic region and has great potential in forensic entomology. We monitored the development at 10 constant temperatures (14–30 °C). Larvae were reared in aggregations. Thermal summation models, isomorphen and isomegalen diagrams and growth curves were derived using the data. Depending on the temperature, development lasted between about 23 and 89 days. Mortality was high at the extremes of the temperature range. The thermal summation constant for the total development was 434.7 ± 28.86 accumulated degree-days above a developmental threshold of 9.04 ± 0.55 °C. This is the first comprehensive dataset on the development of N. littoralis. Implications for its use in forensic casework are discussed.
Similar content being viewed by others
Introduction
Forensic entomology uses insects and other arthropods as evidence in legal investigations. Arthropods are used mainly in homicide, suicide and mysterious death cases, when insect evidence allows to establish some circumstances of death1,2. For instance, due to the ecological specialization of specific insect taxa, it is possible to determine the environment where death occurred and to answer the question whether the body was relocated after death3,4. Most often, however, insects are used to elucidate the time of death, usually through the estimation of minimum postmortem interval (PMImin)5. Two main methods can be used for this purpose. The first one uses the regularity of insect succession on carcasses6,7. Using models of insect succession on carrion for given environmental conditions and a set of insect taxa found on the death scene, we can try to estimate how much time has passed since death8. The second method uses the regularity of insect development, in particular a close relationship between temperature and the rate of development1,9. Developmental studies of necrophagous insects allow for the creation of developmental models that can be used to estimate PMImin.
Forensic entomologists created various kinds of developmental models. Some of them, such as the isomegalen diagram or growth curve, represent the relationship between development time and size of the larvae at a given temperature. The isomorphen diagram shows the duration of developmental stages at given temperatures. The thermal summation model (TSM) assumes that within the certain temperature range, there is a linear relationship between the rate of development and temperature and that development stops below a certain temperature10. Accordingly, there is some constant amount of heat that needs to be accumulated by insects of a given species to reach certain developmental landmark (e.g. hatching, pupation or eclosion). This value is called a thermal summation constant (K) and is expressed in accumulated degree hours (ADH) or accumulated degree days (ADD). The temperature below which the insect species ceases to develop is termed a lower developmental threshold (D0) and is expressed in temperature units10. One of the most frequently used methods to derive TSM, is the one proposed by Ikemoto and Takai11. It is a modification of the classic thermal summation method. Authors proposed an equation:
where D is the duration of development, D0 is the lower developmental threshold, K is the thermal summation constant and T is the environmental temperature. The authors also suggested using the Reduced Major Axis (RMA) regression to derive TSM. Slope of the RMA model is the lower developmental threshold (D0) and the y intercept is the thermal summation constant (K). Although TSMs simplify complexity of insect development and ignore its substantial intraspecific variation (e.g. variation in development time between the sexes12,13 or between insects of different sizes14,15), their use enables satisfactorily accurate estimation of insect age and eventually PMImin8,16,17.
Flies are the most frequently used group of insects in forensic entomology. They usually appear on cadavers as first colonizers and age of their immature stages allow to estimate the PMImin that is close to the true PMI18. Typically, the first colonizers are blow flies (Calliphoridae). They appear on the corpse within the first hours or even minutes after death10,19. During feeding, their larvae form large aggregations20,21,22. Beetles appear later on carcasses, and are often present there until the remains stage. Hence, they can be an important tool in estimating the PMI in the advanced stages of cadaver decomposition23. Numerous studies of insect succession on carrion and descriptions of specific forensic cases demonstrated that beetles may be useful for the estimation of PMI6,17,24,25,26,27,28,29,30,31,32,33,34. Among the forensically important families of beetles, the most frequently mentioned ones are carrion beetles (Silphidae), rove beetles (Staphylinidae), checkered beetles (Cleridae), skin beetles (Dermestidae), clown beetles (Histeridae) and sap beetles (Nitidulidae). The greatest limitation for the use of beetles in forensic entomology is the lack of developmental models.
Developmental models for only 15 species of forensically important beetles have been published so far. First models were created for Thanatophilus micans (Silphidae), population from South Africa35. The other carrion beetles with published models comprise: Oxelytrum discicolle population from northern part of South America36, Thanatophilus mutilatus population from South Africa37, Necrodes littoralis15 population from Central Europe, Necrophila (Calosilpha) brunnicollis population from East Asia38 and populations from Central Europe of Thanatophilus sinuatus39 and Thanatophilus rugosus40. Models were created also for four species of Dermestidae: populations from south-western Europe of Dermestes frischi, D. undulatus and D. maculatus and Chinese population of D. tessellatocollis41,42. Developmental models were also published for central European population of Sciodrepoides watsoni (Leiodidae: Cholevinae)43, Chinese44 and central European45 populations of Creophilus maxillosus (Staphylinidae) and Chinese populations of Necrobia rufipes (Cleridae)46 and Omosita colon (Nitidulidae)47. Although there are beetle taxa that were extensively studied (e.g. Thanatophilus or Creophilus), still many forensically useful beetle species lack developmental models.
Necrodes littoralis (Silphidae) is widely distributed in the Palearctic region. It prefers open and forest natural habitats, but has also been recorded in urban open and quasi-indoor habitats24,31,34,48,49. Larvae feed on carrion mainly in spring and summer, adult beetles have also been recorded in the fall24,48. The species is associated with cadavers at active and advanced decay7,31,48,50. N. littoralis prefers large carrion, on which its larvae frequently form aggregations that allow them to drive active decay similarly to blow flies50,51,52. N. littoralis has been reported from many forensic cases, however it has infrequently been used to estimate PMI17,31,34,49,53,54,55. Comprehensive data on its occurrence on human cadavers in France was provided by Charabidze et al.31. According to their analysis, N. littoralis (larvae or adult beetles) were present in 154 cases (1 in 8 cases examined), with 91.6% of outdoor cases (mainly forests and bushes). Most of them occurred during spring or summer. In more than 85% of the cases N. littoralis was observed from the early to advanced decomposition stages. Moreover, the pre-appearance interval (PAI) of adult and larval N. littoralis was found to be strongly related to the preceding ambient temperature56 and for this reason, it may easily be estimated using temperature methods for PAI57. Furthermore, the size of adult N. littoralis was found to be negatively correlated with its physiological age at maturity. Thus, such features as length or weight of adult N. littoralis can be useful to calibrate developmental constants for this species and to improve the accuracy of age estimation15. Some partial developmental datasets for N. littoralis were already published15,58. However, there is no comprehensive dataset providing all development models and based on the full temperature range for this species. This deficiency partially explains low frequency of N. littoralis use for the estimation of PMI. Current study aims to create the first comprehensive set of robust development models for N. littoralis. Some of the data used in this work was also used for previously published analyzes15.
Results
Life cycle
Adult Necrodes littoralis is mainly active after dark. Mating occurs usually during the night. The female lays eggs in the soil, in batches, usually 50–70 eggs in each. The first instar larvae hatch creamy white and migrate quickly in search for food. It is the period when they are very fragile and particularly vulnerable to injuries. They darken, and their cuticle hardens with time. Likewise, the second and third instar larvae are white and non-sclerotized shortly after ecdysis. When feeding is complete, third instar larvae burrow into the ground. Then they form pupal chambers by thrashing the abdomen and thus compacting the soil around them. They go through the prepupal, pupal and teneral adult stages inside the chambers. The pupa is creamy white at the beginning and with time it gradually sclerotizes and darkens. The metamorphosis ends with the appearance of tenerals—creamy white and non-sclerotized adult beetles. Over time, their cuticle hardens and darkens, giving the insects their final black color. The beetles dig out of the pupal chamber after they became fully sclerotized and colored.
Influence of in vivo measurements
In general, non-measured beetles developed longer (Fig. 1), with significant differences recorded at 15 °C (Mann–Whitney U test: Z = 6.02, p < 0.001), 16 °C (Mann–Whitney U test: Z = 5.92, p < 0.001), 19 °C (Mann–Whitney U test: Z = 3.56, p < 0.001) and 20 °C (Mann–Whitney U test: Z = 3.20, p = 0.001). The development time of non-measured beetles was shorter only at 14 °C (Mann–Whitney U test: Z = − 3.16, p = 0.001). The largest difference was observed in 16 °C, where non-measured beetles developed 5.39% longer than measured beetles. However, in higher temperatures, starting from 17 °C, the differences between measured and non-measured beetles were very small (Fig. 1).

Differences in duration of total immature development between measured and non-measured specimens of N. littoralis at ten constant temperatures. *Statistically significant difference in Mann–Whitney U test at α = 0.001.
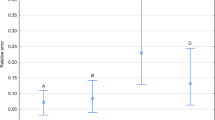
Developmental models calculated using measured and non-measured specimens were only slightly different (Table 1). There were, however no significant differences in the relative error of age estimation between the tested models (Wilcoxon signed-rank test: Z = 0.96, p = 0.34, N = 110). Both models yielded estimates of age with the average relative error below 0.08 (Fig. 2).

Relative error of age estimation using models calculated with measured or non-measured beetles.
Mortality and development
The highest mortality was observed at extreme temperatures: 86.5% at 14 °C and 64.75% at 30 °C. At the other temperatures mortality was below 25%, except for 17 °C where it was 53% (Fig. 3). There were differences between the temperatures in the distribution of deaths across the life stages (Fig. 3.) At the lowest and the highest temperatures deaths occured during all developmental stages, while at the optimal temperatures beetles died mostly at postfeeding larval or pupal stages. At 17 °C deaths were also distributed across all developmental stages. The hypothesis as to why and consequences will be presented in the discussion section.

Mortality of N. littoralis at immature developmental stages in different rearing temperatures.
The development from egg to adult stage took between 22.84 days at 30 °C and 89.11 days at 14 °C (Table 2, Fig. 4.). Results from 17 °C deviated from the general pattern of decrease in development time with an increase in temperature. The first and second instar larvae at 17 °C developed much longer than larvae at 16 °C and 15 °C, and thus the total development time at 17 °C lengthened on average to 60.98 days. Due to this inconsistency and the exceptionally high mortality of larvae at 17 °C, we decided not to use the data from this temperature when developing the models (except for the model for hatching).

Isomorphen diagram for N. littoralis based on median times to reach developmental events at each of the nine temperatures tested (Supplementary Table 1). Horizontal bars represent interquartile ranges. Areas between lines represent developmental stages. Different symbols represent developmental events.
Immediately after hatching, the larvae had an average length of 6.91 ± 0.79 mm (Isomegalen diagram: Supplementary Fig. 1, Growth curves: Supplementary Figs. 2–10). At the growth peak, larvae were on average 25.14 ± 1.82 mm in length. Moulting usually occurred at a similar length of the larvae: 10.71 ± 0.79 mm for the first ecdysis and 16.10 ± 1.30 mm for the second ecdysis. Growth curves were distinctly sigmoidal (Supplementary Figs. 2–10).
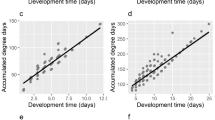
All included temperature points were within 95% confidence interval for all reduced major axis regression models (Fig. 5). The coefficient of determination for each thermal summation model exceeded 0.96, indicating high fit of the models. Lower developmental thresholds (D0) ranged from 9.04 °C for eclosion to 10.86 °C for second ecdysis. Thermal summation constants (K) were between 39.97 ADD for hatching and 434.705 ADD for eclosion (Table 3).

Reduced major axis (RMA) regression models for five developmental events of N. littoralis. DT is the time in days to reach developmental event multiplied by the constant rearing temperature. Temperature values are noted next to the points. Dashed lines represent 95% confidence intervals.
Discussion
Current results on the development time are consistent with the development times of N. littoralis obtained by Dekeirsschieter58. She studied the development at two temperatures only. The development (without the egg stage) took on average 42.79 days at 18 °C and 23.60 days at 23 °C. By subtracting the egg stage from the mean development time at equivalent temperature, in the current study, we get consistent values. TSM for the eclosion from this paper is different compared to the general model for the total immature development given in our previous paper (Table 1 in Gruszka & Matuszewski15). The previous model was created without data from 17 °C and 22 °C, as regression analysis showed that they were outside the 95% confidence interval. In the current study, we excluded the data from 17 °C before the analysis and therefore they were not used while calculating initial TSMs (we give the reasons for this in the results section and discuss proximate and distant causes later in this section). Omitting data from 17 °C changed the results of the regression analysis, and the data point from 22 °C was now within the 95% confidence interval, so it was not omitted while deriving the current final TSMs. Therefore, current article gives more accurate thermal summation values for the eclosion of N. littoralis and we encourage using the current TSM for the eclosion instead of the less accurate previous one.
The value of thermal summation constant (K) for the eclosion of N. littoralis is larger compared to those obtained for other silphid beetles (Table 4). Larger accumulation of thermal units needed to reach maturity by an insect species indicates its longer development time. The development of N. littoralis is the longest among studied silphid beetles. Furthermore, this species usually colonizes cadavers later than for instance Thanatophilus species59,60. Therefore, Necrodes littoralis expands the timeframe in which PMI may be estimated using carrion beetles. In other beetle families, the thermal summation values are larger than in carrion beetles (Table 4).
The pattern of mortality across temperatures is consistent with the patterns revealed for other forensically important beetles. In T. micans, T sinuatus, D. tessellatocollis, C. maxillosus and N. rufipes the highest mortality was observed at extreme temperatures35,39,42,45,46. The same pattern is reported in this study. At intermediate temperatures, mortality of N. littoralis was below 25%, which is also in line with mortality patterns reported for the other beetle species42,45. In the study of Dekeirsschieter at 23 °C, mortality of N. littoralis was surprisingly high. It amounted to 30%58, whereas only 3.75% of the beetles died in 22 °C in the current study. Probably the higher mortality reported by Dekeirsschieter resulted from individual rearing of larvae, since mortality rate of individually reared larvae of N littoralis was found to be higher compared to the larvae that were reared in aggregations61. However, other effects might have been important, as well.
Mortality was exceptionally high at 17 °C in this study. More than half of the beetles died. Moreover, the development at this temperature deviated significantly from the general pattern, with the first and second instar larvae developing longer than at lower temperatures (16 °C and 15 °C). For this reason, we decided not to use data from 17 °C to calculate TSMs. Development extension and high mortality were probably caused here by rearing problems and were not directly related to the temperature itself. At 17 °C large numbers of nematodes were recorded in all containers. They were sticking to the larvae, which probably interrupted larval development and finally led to death of some larvae.
In order to create an isomegalen diagram and growth curves, it is necessary to measure larvae throughout their development. For this purpose, larvae can be killed and then measured. However, killing and storage methods may deform larvae62. Therefore, we decided to measure larvae in vivo. This method also has some drawbacks. First, the temperature changes when larvae are removed from the temperature chambers for measurement. Second, by taking a measurement, stress is induced that in turn can cause an increase in juvenile hormone levels, which interfere with the development63. Previous study64 revealed differences in development time between measured and non-measured Creophilus maxillosus beetles. However, after TSMs were built, it turned out that the estimation error did not differ significantly between the two models. The authors concluded that differences in development time were an effect of repeated stress rather than temperature changes during the measurements. Moreover, lack of significant differences between measured and non-measured TSMs was due to a small size of the differences and moderate size of the validation sample. In the current study, differences in the estimation error between the models were also insignificant. Based on this finding it can be concluded that in vivo measurements of forensically useful beetles have negligible impact on the resultant development models.
Third instar larvae in this study had longer maximum body length than those in previous studies of N. littoralis58,65. In the studies of Dekeirsschieter58, larvae were reared individually on small pieces of meat that could result in their smaller size. In the second study, larvae were collected from pig carcasses on the 17th, 20th and 24th days of decomposition65. The largest third instar larvae used in that study were therefore not the largest (longest) larvae present during decomposition, since after 24th day of decay larvae continued to feed and grow. In the current study, a substantial increase in larval length (above 22 mm) took place at the very end of the feeding phase (Supplementary Figs. 3–10). Fully mature Silphinae larvae range in size from 12 to 40 mm66, current results on larval length are therefore consistent with this range.
When rearing larvae in aggregations, it is difficult to ensure that manual samples are taken at random. We tried to sample larvae irrespective of their position inside the aggregation and their size. However owing to a natural tendency to pick larger larvae, there may be a size bias in our analysis.
A transition of the third instar larva to the post-feeding phase is a critical moment in N. littoralis development. This is when the larva buries itself into the soil to form a pupal chamber. The transition was impossible in the containers in which larvae were reared in aggregations. Therefore, it was necessary to transfer them to separate, smaller containers after they ceased feeding. This transfer might slightly interfere with development. Identification of the exact moment of transition to the post-feeding phase is, however, difficult due to rather long duration and lack of unambiguous markers of transition. The same problem was reported in the study of D. tessellatocollis development, in case of which durations of the last larval stage and the prepupal stage were summed up due to the difficulties in identifying the transition point42. In the study on T. sinuatus, T. rugosus and N. brunnicollis, the post-feeding stage was distinguished38,39,40. Similarly, in the studies of N. littoralis by Dekeirsschieter, the post-feeding stage was identified, supposedly due to the rearing of larvae individually in Petri dishes that facilitated monitoring of the larvae58.
Necrodes littoralis is a species with great forensic potential. However, the lack of development models has largely limited its use in forensic cases. By providing the first comprehensive developmental dataset for this species, current study makes a significant contribution to the advancement of forensic entomology, particularly in central and northern Europe.
Materials and methods
Laboratory colony
Adult beetles came from our laboratory colony established in 2017 using insects collected in the alder forest of Biedrusko military range (Western Poland, Central Europe, 52° 31ʹ N, 16° 54ʹ E). They were kept at room temperature in medium-sized terrariums, 20–30 adult beetles per box (2–3 boxes maintained simultaneously). Boxes contained soil and cotton wool soaked with water. Beetles were fed with pork ad libitum.
Experimental rearing
Laboratory rearing was conducted at ten constant temperatures: 14, 15, 16, 17, 18, 19, 20, 22, 26 and 30 °C. We followed the same protocol for each of the tested temperatures. Rearing was carried out inside the temperature chambers (type ST 1/1 BASIC or ST 1/ 1 + , POL-EKO, Poland). We controlled the chambers using temperature recorders. Deviations from the settings were consistent with the manufacturer's assurances (they did not exceed ± 0.5 °C). In order to induce oviposition adult beetles from the main colony were matched in pairs and placed in 0.5-L containers filled with soil. They had constant access to water and pork ad libitum. Containers were inspected every four hours for the presence of fresh eggs. If eggs were recorded, adult beetles were taken out from the container and eggs were left for hatching. Further inspections were carried at intervals representing no more than 10% of the egg stage duration. Inspection times were calculated based on the results of pilot studies. After hatching, first instar larvae were counted, and 50 larvae were placed in a small terrarium (18 cm × 11 cm × 14 cm) for further rearing. There were eight replicates per temperature (8 containers with 50 larvae each). Rearing boxes contained soil, cotton wool soaked with water, and pork meat ad libitum. Meat was covered with aluminum foil to avoid drying out. After feeding was completed and larvae started to bury themselves, they were transferred to new 0.5-L containers (8–10 larvae per container), filled with soil to allow them to form pupal chambers and complete their development.
Inspections and measurements
Immature beetles from every terrarium were checked for transition to the next developmental stage: second instar larva, third instar larva, post-feeding larva, pupa and adult beetle. Inspections were carried out at time intervals that were no longer than 10% of the stage duration. In the case of the second instar, third instar and post-feeding larvae data were collected per container. Transitions were identified when more than half of the larvae passed to the next stage. Data for pupae and adult beetles were collected per insect. Mortality was measured for each stage of development. For this purpose, live individuals were counted after transition to the next stage. In addition, 24 sampled larvae from four out of eight containers (six larvae per container) were measured in vivo during each inspection. Length of their body was measured from the anterior margin of clypeus to the posterior end of the last abdominal segment. Measurements were made using a geometrical micrometer67. Larvae in the other four containers were inspected for transitions and mortality only.
Data analysis
Influence of in vivo measurements
To test the influence of multiple in vivo measurements on the development of N. littoralis, we compared the total development time of measured and non-measured beetles. 100 randomly selected individuals were used per temperature (54 beetles at 14 °C due to high mortality). Non-parametric Mann–Whitney U test was used in this comparison. To account for the multiple tests we used Bonferonni correction (since there were 10 tests, we used 0.005 level of significance). Then, using data for measured and non-measured beetles, we created separate thermal summation models with the Ikemoto and Takai method11. In order to validate these models we used 110 non-measured beetles (15 beetles per temperature; insects from 17 °C were omitted; due to the high mortality, we had no specimens from 14 °C and only 5 beetles from 30 °C). The true physiological age of the insects (known based on laboratory rearing) was compared with the age estimated using the models. The differences between true and estimated age were used to quantify the estimation errors for measured and non-measured TSM. These errors were compared using the Wilcoxon signed-rank test. Analyzes were performed with Statistica 13 (TIBCO Software Inc.).
Mortality and development
We calculated the percentage mortality for each developmental stage and each of the temperatures tested. Due to the difficulty in accurate separation of the active feeding and post-feeding third instar larvae, we summed up the mortality for these stages. They were similarly combined for the purpose of all the subsequent analyzes. Using data for all individuals, we calculated the mean duration for each developmental stage at each temperature. Median times to reach developmental landmarks (i.e. hatching, first ecdysis, second ecdysis, pupation and eclosion) were calculated and used to build the isomorphen diagram. Using larval measurements, we built isomegalen diagram and growth curves. Thermal summation models were derived using the Ikemoto and Takai method11. For this purpose, we used measured and non-measured beetles. For hatching, first ecdysis and second ecdysis, the sample comprised eight observations per temperature, since we had eight containers for each temperature and could use only “container data” in these analyzes. For pupation and eclosion, the sample comprised 100 observations per temperature, since we could use “individual data” in these analyzes (for 14 °C the sample comprised 63 beetles upon pupation and 54 beetles upon eclosion). Growth curves and thermal summation models were created in R 3.5.2. The other analyzes were performed using Statistica 13 (TIBCO Software Inc.).
Ethical approval
The study comprised laboratory experiments using insect species Necrodes littoralis (Coleoptera: Silphidae). The species is not under protection. No permission or approval from Ethic Commission were needed.
Data availability
The datasets generated and/or analyzed during the study are available from the corresponding author on a reasonable request.
References
Smith, K. G. V. Manual of Forensic Entomology (Trustees of the British Museum, 1986).
Catts, E. P. & Goff, M. L. Forensic entomology in criminal investigations. Annu. Rev. Entomol. 37, 253–272. https://doi.org/10.1146/annurev.en.37.010192.001345 (1992).
Charabidze, D., Gosselin, M. & Hedouin, V. Use of necrophagous insects as evidence of cadaver relocation: Myth or reality?. PeerJ 5, e3506. https://doi.org/10.7717/peerj.3506 (2017).
Matuszewski, S., Szafałowicz, M. & Jarmusz, M. Insects colonising carcasses in open and forest habitats of Central Europe: Search for indicators of corpse relocation. Forens. Sci. Int. 231, 234–239. https://doi.org/10.1016/j.forsciint.2013.05.018 (2013).
Tarone, A. M. & Sanford, M. R. Is PMI the hypothesis or the null hypothesis?. J. Med. Entomol. 54, 1109–1115. https://doi.org/10.1093/jme/tjx119 (2017).
Grassberger, M. & Frank, C. Initial study of arthropod succession on pig carrion in a central European urban habitat. J. Med. Entomol. 41, 511–523. https://doi.org/10.1603/0022-2585-41.3.511 (2004).
Matuszewski, S., Bajerlein, D., Konwerski, S. & Szpila, K. An initial study of insect succession and carrion decomposition in various forest habitats of Central Europe. Forens. Sci. Int. 180, 61–69. https://doi.org/10.1016/j.forsciint.2008.06.015 (2008).
Amendt, J. et al. Best practice in forensic entomology—standards and guidelines. Int. J. Legal Med. 121, 90–104. https://doi.org/10.1007/s00414-006-0086-x (2007).
Higley, L. G. & Haskell, N. In Forensic Entomology. The Utility of Arthropods in Legal Investigations (eds Byrd, J. H. & Castner, J. L.) 389–407 (CRC Press, 2010).
Amendt, J., Richards, C. S., Campobasso, C. P., Zehner, R. & Hall, M. J. R. Forensic entomology: Applications and limitations. Forens. Sci. Med. Pathol. 7, 379–392. https://doi.org/10.1007/s12024-010-9209-2 (2011).
Ikemoto, T. & Takai, K. A new linearized formula for the law of total effective temperature and the evaluation of line-fitting methods with both variables subject to error. Environ. Entomol. 29, 671–682. https://doi.org/10.1603/0046-225x-29.4.671 (2000).
Picard, C. J. et al. Increasing precision in development-based postmortem interval estimates: What’s sex got to do with it?. J. Med. Entomol. 50, 425–431. https://doi.org/10.1603/me12051 (2013).
Frątczak-Łagiewska, K. & Matuszewski, S. Sex-specific developmental models for Creophilus maxillosus (L.) (Coleoptera: Staphylinidae): Searching for larger accuracy of insect age estimates. Int. J. Legal Med. 132, 887–895. https://doi.org/10.1007/s00414-017-1713-4 (2018).
Matuszewski, S. & Frątczak-Łagiewska, K. Size at emergence improves accuracy of age estimates in forensically-usefull beetle Creophilus maxillosus L. (Staphylinidae). Sci. Rep. https://doi.org/10.1038/s41598-018-20796-1 (2018).
Gruszka, J. & Matuszewski, S. Estimation of physiological age at emergence based on traits of the forensically useful adult carrion beetle Necrodes littoralis L. (Silphidae). Forens. Sci. Int. 314, 110407. https://doi.org/10.1016/j.forsciint.2020.110407 (2020).
Matuszewski, S. Post-mortem interval estimation based on insect evidence: Current challenges. Insects 12, 314. https://doi.org/10.3390/insects12040314 (2021).
Bajerlein, D., Taberski, D. & Matuszewski, S. Estimation of postmortem interval (PMI) based on empty puparia of Phormia regina (Meigen) (Diptera: Calliphoridae) and third larval stage of Necrodes littoralis (L.) (Coleoptera: Silphidae)—advantages of using different PMI indicators. J. Forens. Legal Med. 55, 95–98. https://doi.org/10.1016/j.jflm.2018.02.008 (2018).
Greenberg, B. Flies as forensic indicators. J. Med. Entomol. 28, 565–577. https://doi.org/10.1093/jmedent/28.5.565 (1991).
Catts, E. P. Problems in estimating the postmortem interval in death investigations. J. Agric. Entomol. 9, 245–255 (1992).
Payne, J. A. A summer carrion study of the baby pig Sus scrofa Linnaeus. Ecology 46, 592–602 (1965).
Ives, A. R. Aggregation and coexistence in a carrion fly community. Ecol. Monogr. 61, 75–94 (1991).
Slone, D. H. & Gruner, S. V. Thermoregulation in larval aggregations of carrion-feeding blow flies (Diptera: Calliphoridae). J. Med. Entomol. 44, 516–523. https://doi.org/10.1603/0022-2585(2007)44[516:tilaoc]2.0.co;2 (2007).
Kulshrestha, P. & Satpathyb, D. K. Use of beetles in forensic entomology. Forens. Sci. Int. 120, 15–17. https://doi.org/10.1016/s0379-0738(01)00410-8 (2001).
Matuszewski, S., Bajerlein, D., Konwerski, S. & Szpila, K. Insect succession and carrion decomposition in selected forests of Central Europe. Part 2: Composition and residency patterns of carrion fauna. Forens. Sci. Int. 195, 42–51. https://doi.org/10.1016/j.forsciint.2009.11.007 (2010).
Kadej, M., Szleszkowski, Ł, Thannhäuser, A. & Jurek, T. A mummified human corpse and associated insects of forensic importance in indoor conditions. Int. J. Legal Med. 134, 1963–1971 (2020).
Mashaly, A., Al-Khalifa, M., Al-Qahtni, A. & Alshehri, A. Analysis of insects colonised on human corpses during autopsy in Riyadh Saudi Arabia. Entomol. Res. 50, 351–360. https://doi.org/10.1111/1748-5967.12441 (2020).
Meira, L. M. R., Barbosa, T. M., Jales, J. T., Santos, A. N. & Gama, R. A. Insects associated to crime scenes in the northeast of Brazil: Consolidation of collaboration between entomologists and criminal investigation institutes. J. Med. Entomol. 57, 1012–1020. https://doi.org/10.1093/jme/tjaa040 (2020).
Wang, M. et al. Forensic entomology application in China: Four case reports. J. Forensic Leg. Med. 63, 40–47. https://doi.org/10.1016/j.jflm.2019.03.001 (2019).
Moemenbellah-Fard, M. D., Keshavarzi, D., Fereidooni, M. & Soltani, A. First survey of forensically important insects from human corpses in Shiraz, Iran. J. Forens. Legal Med. 54, 62–68. https://doi.org/10.1016/j.jflm.2017.12.016 (2018).
Bonacci, T., Vercillo, V. & Benecke, M. Dermestes frischii and D. undulatus (Coleoptera: Dermestidae) on a Human Corpse in Southern Italy: First Report*. Roman. J. Legal Med. 25, 180–184. https://doi.org/10.4323/rjlm.2017.180 (2017).
Charabidze, D., Vincent, B., Pasquerault, T. & Hedouin, V. The biology and ecology of Necrodes littoralis, a species of forensic interest in Europe. Int. J. Legal Med. 130, 273–280. https://doi.org/10.1007/s00414-015-1253-8 (2016).
Charabidze, D., Colard, T., Vincent, B., Pasquerault, T. & Hedouin, V. Involvement of larder beetles (Coleoptera: Dermestidae) on human cadavers: a review of 81 forensic cases. Int. J. Legal Med. 128, 1021–1030. https://doi.org/10.1007/s00414-013-0945-1 (2014).
Arnaldos, M. I., Garca, M. D., Romera, E., Presa, J. J. & Luna, A. Estimation of postmortem interval in real cases based on experimentally obtained entomological evidence. Forens. Sci. Int. 149, 57–65. https://doi.org/10.1016/j.forsciint.2004.04.087 (2005).
Matuszewski, S. & Mądra-Bielewicz, A. Post-mortem interval estimation based on insect evidence in a quasi-indoor habitat. Sci. Justice 59, 109–115. https://doi.org/10.1016/j.scijus.2018.06.004 (2019).
Midgley, J. M. & Villet, M. H. Development of Thanatophilus micans (Fabricius 1794) (Coleoptera: Silphidae) at constant temperatures. Int. J. Legal Med. 123, 103–108. https://doi.org/10.1007/s00414-008-0280-0 (2009).
Velásquez, Y. & Viloria, A. L. Effects of temperature on the development of the Neotropical carrion beetle Oxelytrum discicolle (Brullé, 1840) (Coleoptera: Silphidae). Forens. Sci. Int. 185, 107–109. https://doi.org/10.1016/j.forsciint.2008.12.020 (2009).
Ridgeway, J. A., Midgley, J. M., Collett, I. J. & Villet, M. H. Advantages of using developmental models of the carrion beetles Thanatophilus micans (Fabricius), and T. mutilatus (Castelneau) (Coleoptera: Silphidae) for estimating minimum post mortem intervals, verified with case data. Int. J. Legal Med. 128, 207–220. https://doi.org/10.1007/s00414-013-0865-0 (2014).
Jakubec, P., Qubaiová, J., Novák, M. & Růžička, J. Developmental Biology of Forensically Important Beetle, Necrophila (Calosilpha) brunnicollis (Coleoptera: Silphidae). J. Med. Entomol. 58, 64–70. https://doi.org/10.1093/jme/tjaa170 (2020).
Montoya-Molina, S. et al. Developmental Models of the Forensically Important Carrion Beetle, Thanatophilus sinuatus (Coleoptera: Silphidae). J. Med. Entomol. https://doi.org/10.1093/jme/tjaa255 (2020).
Montoya-Molina, S. et al. Developmental models of the carrion beetle Thanatophilus rugosus (Linnaeus, 1758) (Coleoptera: Silphidae). Sci. Rep. https://doi.org/10.1038/s41598-021-98833-9 (2021).
Martín-Vega, D., Díaz-Aranda, L. M., Baz, A. & Cifrián, B. Effect of temperature on the survival and development of three forensically relevant Dermestes species (Coleoptera: Dermestidae). J. Med. Entomol. 54, 1140–1150 (2017).
Wang, Y. et al. Development of Dermestes tessellatocollis Motschulsky under different constant temperatures and its implication in forensic entomology. Forens. Sci. Int. https://doi.org/10.1016/j.forsciint.2021.110723 (2021).
Jakubec, P. Thermal summation model and instar determination of all developmental stages of necrophagous beetle, Sciodrepoides watsoni (Spence) (Coleoptera: Leiodidae: Cholevinae). PeerJ https://doi.org/10.7717/peerj.1944 (2016).
Wang, Y. et al. Development of the forensically important beetle Creophilus maxillosus (Coleoptera: Staphylinidae) at constant temperatures. J.Med. Entomol. https://doi.org/10.1093/jme/tjw193 (2016).
Frątczak-Łagiewska, K., Grzywacz, A. & Matuszewski, S. Development and validation of forensically useful growth models for Central European population of Creophilus maxillosus L. (Coleoptera: Staphylinidae). Int. J. Legal Med. https://doi.org/10.1007/s00414-020-02275-3 (2020).
Hu, G. et al. Development of Necrobia rufipes (De Geer, 1775) (Coleoptera: Cleridae) under constant temperatures and its implication in forensic entomology. Forens. Sci. Int. 311, 110275. https://doi.org/10.1016/j.forsciint.2020.110275 (2020).
Wang, Y. et al. Temperature-dependent development of Omosita colon at constant temperature and its implication for PMI min estimation. J. Forens. Leg. Med. 72, 101946. https://doi.org/10.1016/j.jflm.2020.101946 (2020).
Anton, E., Niederegger, S. & Beutel, R. G. Beetles and flies collected on pig carrion in an experimental setting in Thuringia and their forensic implications. Med. Vet. Entomol. 25, 353–364. https://doi.org/10.1111/j.1365-2915.2011.00975.x (2011).
Bonacci, T. et al. First report of the presence of Necrodes littoralis (L) (Coleoptera: Silphidae) on a human corpse in Italy. J. Forens. Sci. 66, 2511–2514. https://doi.org/10.1111/1556-4029.14821 (2021).
Matuszewski, S., Bajerlein, D., Konwerski, S. & Szpila, K. Insect succession and carrion decomposition in selected forests of Central Europe. Part 1: Pattern and rate of decomposition. Forens. Sci. Int. 194, 85–93. https://doi.org/10.1016/j.forsciint.2009.10.016 (2010).
Matuszewski, S., Konwerski, S., Frątczak, K. & Szafałowicz, M. Effect of body mass and clothing on decomposition of pig carcasses. Int. J. Legal Med. 128, 1039–1048. https://doi.org/10.1007/s00414-014-0965-5 (2014).
Gruszka, J. et al. Patterns and mechanisms for larval aggregation in carrion beetle Necrodes littoralis (Coleoptera: Silphidae). Anim. Behav. 162, 1–10. https://doi.org/10.1016/j.anbehav.2020.01.011 (2020).
Saloña-Bordas, M. I. & Perotti, M. A. First contribution of mites (Acari) to the forensic analysis of hanged corpses: A case study from Spain. Forens. Sci. Int. https://doi.org/10.1016/j.forsciint.2014.08.005 (2014).
Dekeirsschieter, J., Frederickx, C., Verheggen, F., Boxho, P. & Haubruge, E. Forensic entomology investigations from doctor marcel leclercq (1924–2008): A review of cases from 1969 to 2005. J. Med. Entomol. 50, 935–954. https://doi.org/10.1603/me12097 (2013).
Lutz, L., Zehner, R., Verhoff, M. A., Bratzke, H. & Amendt, J. It is all about the insects: A retrospective on 20 years of forensic entomology highlights the importance of insects in legal investigations. Int. J. Legal Med. 135, 2637–2651. https://doi.org/10.1007/s00414-021-02628-6 (2021).
Matuszewski, S. & Szafałowicz, M. Temperature-dependent appearance of forensically useful beetles on carcasses. Forens. Sci. Int. 229, 92–99. https://doi.org/10.1016/j.forsciint.2013.03.034 (2013).
Matuszewski, S. & Mądra-Bielewicz, A. Validation of temperature methods for the estimation of pre-appearance interval in carrion insects. Forens. Sci. Med. Pathol. 12, 50–57. https://doi.org/10.1007/s12024-015-9735-z (2016).
Dekeirsschieter, J. Etude des interactions entre l’entomofaune et un cadavre: approches biologique, comportementale et chémo-écologique du coléoptère nécrophage, Thanatophilus sinuatus Fabricius (Col., Silphidae) PhD thesis, Gembloux Agro-Bio Tech - University of Liège, (2012).
Matuszewski, S., Bajerlein, D., Konwerski, S. & Szpila, K. Insect succession and carrion decomposition in selected forests of Central Europe. Part 3: Succession of carrion fauna. Forens. Sci. Int. 207, 150–163. https://doi.org/10.1016/j.forsciint.2010.09.022 (2011).
Matuszewski, S. et al. Effect of body mass and clothing on carrion entomofauna. Int. J. Legal Med. 130, 221–232. https://doi.org/10.1007/s00414-015-1145-y (2016).
Gruszka, J. & Matuszewski, S. Insect rearing protocols in forensic entomology: benefits from collective rearing of larvae in a carrion beetle Necrodes littoralis L. (Silphidae). PLoS ONE 16, e0260680. https://doi.org/10.1371/journal.pone.0260680 (2021).
Midgley, J. M. & Villet, M. H. Effect of the killing method on post-mortem change in length of larvae of Thanatophilus micans (Fabricius 1794) (Coleoptera: Silphidae) stored in 70% ethanol. Int. J. Legal Med. 123, 103–108. https://doi.org/10.1007/s00414-008-0260-4 (2009).
Tauchman, S. J., Lorch, J. M., Orth, A. P. & Goodman, W. G. Effects of stress on the hemolymph juvenile hormone binding protein titers of Manduca sexta. Insect Biochem. Mol. Biol. 37, 847–854. https://doi.org/10.1016/j.ibmb.2007.05.015 (2007).
Frątczak-Łagiewska, K. & Matuszewski, S. The quality of developmental reference data in forensic entomology: Detrimental effects of multiple, in vivo measurements in Creophilus maxillosus L. (Coleoptera: Staphylinidae). Forens. Sci. Int. 298, 316–322. https://doi.org/10.1016/j.forsciint.2019.02.059 (2019).
Frątczak, K. & Matuszewski, S. Instar determination in forensically useful beetles Necrodes littoralis (Silphidae) and Creophilus maxillosus (Staphylinidae). Forens. Sci. Int. 241, 20–26. https://doi.org/10.1016/j.forsciint.2014.04.026 (2014).
Newton, A. F. J. In Immature insects Vol. 2 (ed. Stehr, F. W.) 339–341 (Kendall/Hunt, 1991).
Villet, M. H. An inexpensive geometrical micrometer for measuring small, live insects quickly without harming them. Entomol. Exp. Appl. 122, 279–280. https://doi.org/10.1111/j.1570-7458.2006.00520.x (2007).
Acknowledgements
The study was funded by the National Science Center of Poland (grant no. 2016/21/B/NZ8/00788).
Author information
Authors and Affiliations
Contributions
J.G. performed laboratory rearing and data collection, analyzed the results and prepared figures. S.M. obtained funds and supervised the research. Both authors designed the study and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gruszka, J., Matuszewski, S. Temperature models of development for Necrodes littoralis L. (Coleoptera: Silphidae), a carrion beetle of forensic importance in the Palearctic region. Sci Rep 12, 9689 (2022). https://doi.org/10.1038/s41598-022-13901-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-13901-y
- Springer Nature Limited
This article is cited by
-
Development of Phormia regina at seven constant temperatures for minimum postmortem interval estimation
International Journal of Legal Medicine (2024)
-
Initial laboratory validation of temperature development models for Necrodes littoralis L. (Staphylinidae: Silphinae)
International Journal of Legal Medicine (2023)
-
A beetle for prosecution: exogenous DNA detection from larval and adult gut of a Neotropical carrion beetle
Forensic Science, Medicine and Pathology (2023)