
Abstract
A wide variety of substances and agents are released into the atmosphere due to anthropogenic activities. Higher levels of such entities have given rise to four major environmental problems: air, light and noise pollution and global warming, all of which severely affect birds and other animals. These four issues have been overlooked, although climate change is receiving increasing attention. The four challenges often occur simultaneously and are likely to exert composite impacts. Most studies have focused on the effects of a specific problem at a particular time and have never taken into account their cumulative consequences. This review tries to address this shortcoming. It aims to evaluate the composite impacts of the problems on flying birds beyond an understanding of their individual impacts. The review initially sheds light on the individual impacts based on existing scientific literature. Composite impacts were then estimated by assigning suitable scores to the literature to convert it into empirical data. Scores were then analysed. Through this assessment, it was found that the health of birds is highly vulnerable to the composite effects of the problems. Additionally, statistical analysis revealed that the effects of all problems on different aspects of avian biology are likely to be magnified simultaneously in the future. Thus, the impact of these problems on birds should not be neglected, and further studies should be conducted to understand their mechanisms.
Similar content being viewed by others

1 Introduction
A wide range of substances/agents are released into the environment due to anthropogenic activities. Higher levels of such entities result in effects that reduce the quality of the biosphere (Morrison 1986). In general, the ambient environment is negatively affected by the harmful effects of four major problems arising due to the emission of substances/agents from human activities; air pollution, light pollution, noise pollution and global warming/climate change. With regard to the latter, birds and amphibians are the most threatened animal groups (Foden et al. 2013; Bird Life International 2013).
These four problems exert impacts that vividly extend beyond mankind into the animal world. However, except for global warming, their impact on wildlife has received minimal attention in terms of research. Issues related to global warming and birds have been studied (Sekercioğlu et al. 2012). But the scientific literature does not provide any insight on how the problem acts in the presence of other environmental factors. There is a need to assess the composite effects of these issues as they commonly occur together at the same time. The gravity of the situation can be well understood by the fact that the Aichi Biodiversity Targets (20 targets under five strategic goals) have emphasised the reduction in different types of pollution to levels that are not detrimental to ecosystem function and biodiversity (Target 8) as well as the minimisation of climate change to maintain ecosystem function and integrity (Target 10) (Convention on Biological Diversity 2013; O’Connor et al. 2015). These targets form an integral part of the ‘Strategic Plan for Biodiversity 2011–2020’ adopted by the Convention on Biological Diversity (CBD) in 2010 in view of drastic declines in global biodiversity. The targets aim to facilitate global biodiversity conservation and guide regional and national-scale target setting (O’Connor et al. 2015), and the international community has agreed to attempt to achieve these by 2020 (Tittensor et al. 2014).
Flying birds are commonly exposed to all four problems and are at a high risk of their negative effects. They provide much scope for the study of their composite effects, which has not been performed previously. In fact, this knowledge deficit exists because no study has concentrated on more than one problem simultaneously, even though all issues often occur together. Such an evaluation is also essential because birds are much overlooked in this regard (except for the impact of climate change, as mentioned above). According to the UN Millennium Ecosystem Assessment, birds provide four types of ecosystem services: provisioning, regulating, cultural and supporting services. Additionally, they play important roles in predation, pollination, scavenging, seed dispersal, seed predation and ecosystem engineering (Whelan et al. 2008). Hence, birds must be conserved and for successful conservation, the negative effects of all four problems must be understood and mitigated.
Taking the above into consideration, the present review attempts to understand the individual and composite consequences of these four problems on flying birds as a whole. It also tries to identify possible synergistic associations among the issues. It attempts to fulfil this aim using published scientific literature collected from peer-reviewed sources, supplemented with information from databases of organisations/institutes associated with wildlife and environmental research. The review is the first to use such an approach and has two general objectives:
-
(a)
To review the individual effects of the problems on birds through analysis of the scientific literature.
-
(b)
To assess the composite and synergistic effects of the problems through analyses (including statistical analysis) of scores assigned based on secondary data.
2 Air pollution
Atmospheric pollutants that arise from human activities exert widespread effects on all living organisms. In this regard, human health has always been the primary concern of research (Gupta and Bakre 2013). However, it also has a number of implications for birds, as outlined below.
2.1 Direct impacts
Birds have narrow lung capillaries and higher breathing rates and spend considerable amount of time in open air. Hence, they are highly vulnerable to atmospheric particulate matter. Ground-level ozone and nitrogen oxides also lead to negative effects on avian lungs (e.g. inflammation and lung failure). In addition, these pollutants can also rupture of blood vessels upon long-term exposure (Qin 2015). In Beijing and Manila, emissions have given rise to black lungs and enlarged testes (Lovett 2012). In passerine birds, pollutants that arise from coal-fired power plants bring about changes in the tracheal epithelium including increased mucus cover, shortening of the cilia, and an increase in the number of secretory granules and vesicles (Llacuna et al. 1993). Emission of NO x , SO2 and particulates from such plants has led to variation in the percentage of ciliated and non-ciliated cells, as well as the organisation, orientation and morphology of cilia in the tracheal epithelium of goldfinch (Carduelis carduelis), rock bunting (Emberiza cia), great tit (Parus major) and blackbird (Turdus merula) (Gorriz et al. 1994).
The erythrocyte count decreases, whereas erythrocyte size increases in rock bunting and blackbird under the influence of SO2, NO x and particles released from coal-fired power plants. Pre-albumins are increased, and β-globulins are decreased in rock bunting. In blackbird, transaminases (GOT and GPT) are increased and weight is decreased (Llacuna et al. 1996). Haematological effects are also evident in sparrows that live in highly polluted urban areas as these tend to have very low levels of haemoglobin and antioxidant capacity. The intensity of this impact is directly related to the level of pollution (European Commission 2013). Another group of harmful pollutants is polycyclic aromatic hydrocarbons (PAHs) emitted from traffic. Their long-term exposure can lead to the inhibition of growth, reduction in the production and hatching of eggs and higher abandonment rates of clutches or broods. PAHs could also trigger DNA mutations that can persist through several generations (as evident in Double-crested Cormorant Phalacrocorax autitus in Canada). These mutations can also be carcinogenic (Qin 2015).
The above harmful pollutants are accumulated in avian bodies following increased emission. For instance, the lungs and livers of birds in Beijing have been found to contain three to four times more PAHs and common by-products of fossil fuel burning, compared with birds in higher air quality areas (Lovett 2012). Heavy metal accumulation also occurs due to pollution by coal-fired plants, and this varies among different species. This is evident from the fact that blackbirds accumulate significantly more heavy metal residues than great tit or rock bunting (Llacuna et al. 1995).
In addition to the above, a number of miscellaneous problems also occur, such as reduction in breeding success, as is evident in great tit in southern Poland (Nyholm et al. 1995). Further, in addition to affecting the physiology of great tit, air pollution can also affect their song. This is evident from the fact that males of great tit have been found to sing less near emission sources (Gorissen et al. 2005; Isaksson et al. 2005). Similarly, copper smelter emissions have been found to reduce the thickness of egg shells, egg volume and content, clutch size and hatching success of great tit and pied flycatcher (Ficedula hypoleuca) (Eeva and Lehikoinen 1995). Next, ozone (and some other atmospheric pollutants) affects feeding activity in ruby-throated hummingbird (Archilochus colubris). As these birds breathe higher volumes of air with respect to their bodyweight, they are also at risk of experiencing greater adverse effects of ozone (Lewis 2006).
2.2 Indirect impacts
Air pollution can also affect birds by triggering habitat changes, increasing parasites and reducing food sources (Morrison 1986; Graveland 1990; Eeva et al. 1994; Hörnfeldt and Nyholm 1996).
Emissions can also suppress plant carotenoid levels, and this reduces their availability for phytophagous insects. Lower carotenoid accumulation in phytophagous insects in turn lowers their levels in insectivorous passerine birds. Consequently, the supply of carotenoids (which provide antioxidant protection and immune-competence) from parents to nestlings is inadequate in such birds. Carotenoids are also required for the development of eggs. Therefore, the development of both nestlings and eggs could be affected (Sillanpää 2010). Emissions have also been found to decrease the yellow plumage colour in great tit nestlings by affecting the abundance of prey insects (green caterpillars and sawfly larvae fed to nestlings) that provide carotenoids. Pollutants can also obstruct carotenoid metabolism and transportation to feathers and/or affect the concentration of carotenoid derivatives in organs. Thus, nestlings at greater distances from pollution sources have better colour intensity and are also heavier. This fact indicates lower survival probability of pale nestlings. Plumage colour can affect selection of mates by female birds, male–male competition, social dominance in winter flocks and the ability to avoid predation via crypticism (Eeva et al. 1998).
Air pollution (heavy metals) from copper smelters negatively affects many invertebrate species. This in turn affects the diet of insectivorous birds such as great tit (caterpillar specialist) and pied flycatcher, an opportunistic forager. Feeding frequency or food quantity is not affected. However, the type of food consumed is altered in both species. Breeding success of great tit is affected by the change in food quality, whereas the same in pied flycatcher is not affected. This indicates a greater vulnerability of the former to the lack of carotenoid-rich caterpillars. However, pied flycatcher is more sensitive to a decreased amount of calcium-rich food items due to heavy metal pollution (Eeva et al. 2005).
Sulphur dioxide and nitrogen oxides, which arise from fossil fuel combustion, can cause acidification of water bodies. This in turn affects the quantity and quality of avian food resources (Hare et al. 2002). Several molluscs and crustaceans have high concentrations of calcium and are sensitive to pH. They disappear rapidly upon acidification of wetlands (Hare et al. 2002). On the other hand, higher levels of aluminium that arise due to acidification cause thinning of eggshells in some bird species like the great tit and pied flycatcher (Dudley and Stolton 1996). However, acidification can exert varied consequences. For example, ospreys (Pandion haliaetus) find fewer fish to consume in an acidic lake because there are far fewer fish present. However, the same could be helpful for divers because clearer water in acidic lakes makes hunting easier (Hare et al. 2002). In addition, eutrophication can arise due to higher levels of atmospheric nitrogen oxides. Consequently, negative affects occur in fish and invertebrates which are used by birds as food resources. Over time, nitrogen oxide accumulation can facilitate the propagation of nitrogen-loving invasive plants that flourish at the expense of native plants (such as lichens) which provide forage and nesting material to birds. As a whole, the overall impacts of emissions can also extend up to bird habitats and trigger remarkable alterations in landscape (Qin 2015).
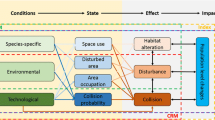
Furthermore, accumulation of atmospheric dioxins in soil leads to their greater absorption by earthworms. Although earthworms remain unaffected, they can accumulate up to fivefold higher levels of these chemicals than their concentration in the surrounding soil. Birds then acquire this chemical by consuming earthworms, which can lead to carcinogenic, reproductive and immuno-toxic effects (Hare et al. 2002). The overall impact of emissions on birds is depicted in Fig. 1.

Impacts of air pollution on birds and their mechanisms of action
3 Noise
3.1 Noise as a problem
Anthropogenic noise is on the rise due to the introduction of more powerful sources, greater geographic spread and mobility of noise sources, and the exposure of a greater proportion of the day to noise (Berglund and Lindvall 1995). It is a unique evolutional selection pressure that exerts consequences at both individual and population levels (Slabbekoorn and Ripmeester 2008). Noise propagates through natural ecosystems throughout the world and imposes pressure on wildlife processes (Barber et al. 2010; Tennessen et al. 2014). Physiological effects of noise on animals include changes in endocrine, digestive, blood, immune and reproductive function (US Department of Transportation and Federal Administration 2004).
The effects of noise on natural populations and communities are not well understood (Francis et al. 2012). Although its consequences have generated much interest, little is known about the impact of noise on wildlife (Tennessen et al. 2014). For instance, sublethal consequences of noise such as physiological stress and impaired reproduction have not been properly reported (Kight and Swaddle 2011). Noise is predicted to increase with increasing human population growth (Babisch et al. 2005), and even urban bird species adapted to a variety of environmental conditions could be sensitive to noise (Francis et al. 2011).
3.2 Impact of noise on birds
3.2.1 Avian calls
The purpose of bird song is to indicate territorial occupancy and/or reproductive readiness (Gil et al. 2015). In fact, acoustic communication provides sexually selected cues for mating and defending a territory (Mockford and Marshall 2009). Increasing levels of anthropogenic noise interferes with such communication (Diaz et al. 2011), which is a fundamental determinant of avian sexual selection and social integration (Catchpole and Slater 1995). It is in fact the background noise that primarily decides which individuals receive vocal signals and the viability of such signals (Patricelli and Blickley 2006). In this way, noise exerts selection pressure on acoustic signals (Ryan and Brenowitz 1985). When low-frequency male calls are interrupted by noise, song-based assessments in females are masked. As a result, lower energy is invested in egg production. This is why great tits nesting in noisy areas has reduced clutch sizes (Francis et al. 2011). In addition, such masking can also hamper the ability of female birds to identify high-quality males. Consequently, pairing could take place with inferior males whose higher frequency signals are less interrupted by noise (Francis et al. 2011).
Facultative behavioural responses to anthropogenic noise include alterations in amplitude, frequency, timing and duration of signals to minimise acoustic competition (Fuller et al. 2007). In Europe, bird populations have been found to greatly advance their song timing, when singing overlaps with air traffic. The extent of advancement varies among species, according to the disturbance level and species resilience. This modification indicates adaptive change and does not take place due to circadian constraints. Such modification of normal singing times enables survival of avian populations in noisy environments. But it entails a higher energetic cost and altered competitive ability and resilience (Gil et al. 2015).
Another important example is urban traffic noise, which directly interferes with avian acoustics due to its low frequency (Mockford and Marshall 2009). Serins (Serinus serinus) have been found to increase singing at both spatial and temporal scales under such noise exposure (Diaz et al. 2011). Adjustment is also evident in male blackbirds that attempt to adjust their dawn song timing to periods of lower noise in response to traffic. Consequently, they are bound to become active several hours before humans (Helmholtz Centre for Environmental Research 2013). European Robins (Erithacus rubecula) reduce acoustic interference by singing at night in areas of high diurnal noise (Fuller et al. 2007). Robins in Britain have been found to avoid their dawn chorus at peak rush hour and instead shift to singing at night (Francis et al. 2009). Nocturnal singing by diurnal birds may minimise interference from ambient urban noise. However, this could increase the metabolic rate at the cost of sleep (Fuller et al. 2007).
Birds also change the frequency and amplitude of their songs in response to noise. Increasing the calling amplitude by the signaller under greater noise exposure is termed the Lombard effect (Lombard 1911). Free-ranging male nightingales (Luscinia megarhynchos) sing louder territorial songs at noisier locations (Brumm 2004). German nightingales have taken to singing more loudly (95 decibels), which can be harmful to human hearing (Francis et al. 2009). In urban song sparrows (Melospiza melodia), a positive relationship has been found between the minimum frequency of male song and the amplitude of anthropogenic noise (Wood and Yezerinac 2006). great tits, house finches (Carpodacus mexicanus) and oscine birds also adjust their call frequency to prevent masking by noise. However, high-frequency sounds might not efficiently propagate and lead to decreased efficiency of the acoustic signal. Some birds might increase the amplitude of their calls. The extent of such increase depends upon body size as well as energy cost. In this case, energy costs may reduce the benefit of vocal adjustment (Patricelli and Blickley 2006). Singing under noisy situations may also take place at the expense of vigilance and hence may pose a greater risk of survival (Diaz et al. 2011).
Human development has been proceeding at a great pace. Consequently, there is likely to be greater acoustic interference with animal communication, which in turn can lead to important behavioural consequences (Fuller et al. 2007). Larger bird species emit lower-frequency signals. Therefore, larger birds are more vulnerable to noisy areas than smaller species which transmit higher frequency signals. Body size, vocal amplitude and frequency are important factors that determine tolerance (Francis et al. 2011). Smaller species that rely upon high-frequency transmissions could persist in noisy environments. In addition, these could also benefit from reduced predation risk in such areas. But this benefit entails costs to male–female communication, pairing success and reproductive success in the absence of predation (Francis et al. 2011).
3.2.2 Impacts on health and physiology
Developing nestlings are sensitive to noise, but they could not move away from it (they are confinement to their nests). They could suffer from immediate effects like suppression of growth and immune function (Crino et al. 2013). Long-term impacts could lead to physiological, morphological and behavioural consequences. Such consequences could include lifelong and trans-generational effects on reproductive success and survival. In fact, exposure to even short periods of stress, such as noise, during development could lead to large-scale effects (Crino et al. 2013). This has been observed in nestlings of white-crowned sparrows (Zonotrichia leucophrys) that suffer from phenotypic effects under higher levels of traffic noise (Crino et al. 2013). Birds could use various strategies to overcome the effects of noise. This could lead to evolutionary changes in signal characteristics or short-term adaptations (Herrera-Montes and Aide 2010).
3.2.3 Ecological impacts
Bird abundance, occurrence and species richness are reduced near roads in response to noise. The largest reductions occur where traffic levels are high (Summers et al. 2011). Similar effects are also evident near airports. A small percentage of species with sufficiently plastic behaviour to escape noise can thrive near airports. Even so, this diminishes the bird community and consequently the ecosystem is affected (Gil et al. 2015). Diversity of bird communities is reduced by noise, especially in secondary lowland forest sites (Francis et al. 2009; Herrera-Montes and Aide 2010). However, sometimes noise exerts a beneficial effect on smaller birds if it cannot be tolerated by egg-eating predators such as the Western Scrub Jay (Aphelocoma californica) which causes nest failure. However, this can also have significant ecological consequences, since Scrub Jays are important for pollination (Francis et al. 2009). On the other hand, noise indirectly increases pollination by hummingbirds (Francis et al. 2012).
4 Light pollution
4.1 Light as a problem
An increase in artificial lighting has impacted heavily on natural light regimes and has led to several ecological consequences (ranging from changes in animal behaviour to community composition). Potential impacts include extension of foraging time, sexual competition of diurnal and crepuscular animals into night, improvement in prey detection and predator avoidance capabilities, changes in the capacity to navigate and ability of pollinators to detect nectar sources. Studies on these aspects are limited because the problem has only been recently recognised as an environmental issue (Davies et al. 2013). The impacts of light can be subtle, and such effects have not received the required attention (Poot et al. 2008; Kempenaers et al. 2010). However, various characteristics of light have the potential to influence ecological and evolutionary processes (Navara and Nelson 2007; van Langevelde et al. 2011). The evolution of plants and animals has occurred on the basis of day–night cycle, and artificial light disrupts this cycle (Cinzano et al. 2001). Light has in fact been an ecological issue for a long time, and the problem is likely to increase with developments in lighting technologies (Gaston et al. 2013). There are a number of implications which need to be studied (Gaston et al. 2013), some of which are discussed in the following section.
4.2 Impact of light on birds
4.2.1 Natural behaviour
The avian circadian rhythm (which determines the time of mating, breeding, foraging and migration) is dependent upon light (Mahr et al. 2012). Therefore, alterations in activity patterns take place under the influence of light (Mahr et al. 2012). In other words, when natural day and night rhythms are affected by artificial light, natural behavioural patterns may be altered (Mahr et al. 2012). Hence, artificial light can prepone or postpone bird migration because migration primarily depends upon cues available from properly timed seasonal schedules. Consequently, birds can miss optimal conditions for nesting, foraging and other behaviours (International Dark Sky Association). The importance of light on behaviour is demonstrated by the fact that Greater Flamingos (Phoenicopterus roseus) and Peahens (Pavo cristatus) exhibit greater vigilance behaviour at night under lower light levels (Yorzinski et al. 2015). In some birds, foraging is extended upon exposure to artificial light. Mockingbirds (Minimus sp.) feed their nestlings in the late evening under the influence of greater artificial light. European blackbirds continue foraging longer into the evening and begin their mornings earlier when exposed to artificial night lighting. Thus, light can have a substantial impact on the fitness of birds by changing their behaviour (Yorzinski et al. 2015).
Light exerts an important influence on avian breeding behaviour. It leads to a number of consequences which are yet to be studied (Kempenaers et al. 2010). Birds have been found to prepone breeding under the influence of longer day length (Florida Fish and Wildlife Conservation Commission). This phenomenon has been reported in urban blackbirds in which light can prepone breeding by almost a month. Moulting could also be preponed in these birds by up to 3 months (in comparison with birds living in rural areas) under the influence of light (Helm et al. 2013; Partecke et al. 2005). Another bird in which reproductive behaviour is affected by artificial light is the Blue Tit (Cyanistes caeruleus) (Mahr et al. 2012). In this case, males near light sources at forest edges have been found to be more successful in attracting additional mates. In other words, these often have offspring with females other than their primary social partners (Kempenaers et al. 2010). Thus, nocturnal light could influence avian strategies for choosing partners, as males and females may have separate preferences for light (Mahr et al. 2012). The overall effects of night lighting on breeding birds are expected to be more magnified with increases in spring temperatures due to global warming (Kempenaers et al. 2010).
The effects of night light could be intensified by traffic noise. The combined impact of may make birds active up to 5 h earlier in morning compared with birds exposed to natural light regimes (Helmholtz Centre for Environmental Research 2013). Such composite consequences are also well reflected in the behavioural patterns and natural cycles of city blackbirds (Helmholtz Centre for Environmental Research 2013). This is a relevant problem of modern times because road networks and traffic volumes are projected to increase worldwide. Noise and artificial light are important effects of roads that can also act synergistically with other effects to cause greater ecological impacts (Kociolek et al. 2011). However, there are several anthropogenic factors that determine the impact of artificial night lighting on natural rhythms. The impacts on fitness of such changes in seasonal timing of behaviour are also not completely understood (Da Silva et al. 2015).
4.2.2 Disorientation
Artificial light acts as a strong false orientation cue that can trap birds (Verheijen 1985). Nocturnal migrants are the most vulnerable to this issue (Gauthreaux and Belser 2006; Watson et al. 2016) since light obstructs orientation (Gauthreaux and Belser 2006; Mahr et al. 2012). Visual pigments of migratory birds are bleached by artificial light. Consequently, they can lose sight of the horizon, which makes them circle within a cone of light. This can result in exhaustion or collision with the light source (Florida Fish and Wildlife Conservation Commission). Artificial light can also cause the deviation of birds away from the correct migration routes towards harmful city landscapes at night. This is demonstrated by the fact that large numbers of birds die due to collisions with illuminated structures every year (International Dark Sky Association). For instance, about 98 million to one billion birds die following collisions with man-made structures across North America alone (Chepesiuk 2009). Many nocturnally migrating birds also die or lose a large amount of energy upon encountering artificial light sources such as offshore platforms (Poot et al. 2008). The reasons why birds are attracted towards artificially lit structures are not well understood (Poot et al. 2008). In addition to triggering changes in migratory routes and causing birds to pass over illuminated sites, light can also lead to flight at lower altitudes, increased calling and resulting in birds spending more time over lit areas (Watson et al. 2016).
Disorientation due to light can have fatal consequences for threatened seabirds such as Petrels (Balearic Shearwater Puffinus mauretanicus, Scopoli’s Shearwater Calonectris diomedea, and European Storm-petrel Hydrobates pelagicus). Fledglings are attracted to artificial lights during their maiden night flights from the nest to the sea. As a result, they can become grounded and exposed to multiple threats. This could lead to high mortality (Rodríguez et al. 2015). Disorientation also leads to inefficient feeding by seabirds in feeding grounds because they prey on bioluminescent sea animals under low light levels (Florida Fish and Wildlife Conservation Commission). However, nocturnal lights can enable some shorebirds to utilise visual foraging at night instead of tactile foraging (Rojas et al. 1999). On the whole, bird responses to all light conditions are strongest on nights devoid of moon and starlight (Poot et al. 2008).
4.2.3 Mating calls
When the intensity of artificial light exceeds a threshold, earlier onset of dawn song occurs in some birds (Helmholtz Centre for Environmental Research 2013). In fact, under the influence of artificial light, many songbirds prepone singing around dawn, postpone singing around dusk and even engage in nocturnal singing (Da Silva et al. 2015). Earlier singing results in sleep loss in male birds and increases the risk of predation (Kempenaers et al. 2010). Night lights can elongate the day length of diurnal songbirds and make them more prone to predators as they sing beyond their location (Florida Fish and Wildlife Conservation Commission). The presence of street lights leads to earlier morning singing in the males of some bird species. In the corresponding females, it induces earlier egg laying (Kempenaers et al. 2010). The females of some birds are considered to engage in additional copulations with high-quality sires to increase the quality of their offspring. These could use early singing, due to light as a cue to decide the quality of male. Light could thus disrupt the link between the cue: early singing—male quality, so that females would end up having lower-quality males (Kempenaers et al. 2010).
4.2.4 Additional effects of light
Road lighting affects the density of nests for some species (Longcore and Rich 2004). However, certain species tend to shift between periods of light absence and presence (McClure et al. 2013). In addition, disruption of melatonin production may occur in birds exposed to night light. Such an impairment of melatonin production could cause severe physiological consequences (Gaston et al. 2013).
Moreover, indirect effects are also apparent. For instance, many artificial lights act as fatal distraction for insects and result in their decline. This is well evident in Germany where attraction towards artificial lights causes the death of a huge number of insects every summer. This in turn negatively impacts on birds that use insects as a food resource (International Dark Sky Association; Eisenbeis 2006).
5 Global warming
5.1 Global warming as a problem
The current trend in global warming is highly significant because most of it is human induced, and it has been taking place at an unprecedented rate in the past 1300 years (Whelan et al. 2008). Average world temperature has increased by 0.83 °C during the previous century, and it is expected to surge by another 0.28–4.78 °C over the next century. The average rate of global annual temperature increase is 0.07 °C per decade since 1880 and 0.17 °C per decade since 1970. The average global land and ocean temperature for 2016 was 0.94 °C higher than the twentieth-century average of 13.9 °C. It surpassed the earlier record of 2015 by 0.04 °C. This was the fifth time that a record high annual temperature was set in the twenty-first century (along with 2005, 2010, 2014, and 2015) (Global Climate Report-Annual 2016).
This increased heating is the result of higher atmospheric levels of carbon dioxide, methane, nitrous oxide and chlorofluorocarbons (US EPA 2016). Global warming is in turn triggering remarkable changes in climate (Sahu et al. 2015). Approximately 60% of global warming has been linked with carbon dioxide (Grace 2004). The atmospheric concentration of this greenhouse gas has increased drastically since the start of the industrial revolution (World Meteorological Organization Website; British Geological Society Website). Carbon dioxide is released in huge quantities due to deforestation, fossil fuel combustion and cement production (World Meteorological Organisation Website; British Geological Society Website). About half of the anthropogenic CO2 emissions between 1750 and 2011 have occurred in the last 40 years. It is predicted that more frequent heat waves and extreme precipitation events will occur as a result, in addition to greater warming and acidification of the ocean (IPCC 2014).
Methane is produced in landfill sites as well as the agricultural and animal husbandry sectors (World Meteorological Organisation Website; British Geological Society Website). Nitrous oxide arises predominantly due to nitrification and denitrification in agricultural soils. From every 1000 kg of reactive nitrogen (mainly fertiliser) in such soils, approximately 10–50 kg is released as nitrous oxide gas. This is a serious issue because a huge increase in cultivated land is expected in the near future (Tilman et al. 2001; IPCC 2007; Singh et al. 2010). Chlorofluorocarbons (regulated under the Montreal Protocol) are synthetic compounds utilised mainly for refrigeration and air conditioning. Growth in industrialisation is directly related to the increase in emission of these greenhouse gases (tenfold increase since the beginning of the twentieth century) (World Meteorological Organization Website).
Climate change is likely to increase the frequency of certain climatic extremes that can exert drastic impacts on biodiversity (Jentsch et al. 2007; Seneviratne et al. 2014; US EPA 2015). This is especially likely to occur if meteorological thresholds are surpassed and lead to population collapses (Oliver et al. 2015). In fact, prominent environmental parameters of climate change such as temperature, solar radiation, humidity, cloud cover and precipitation have implications for biodiversity (Bickford et al. 2010). Consequently, biodiversity is already being impacted due to climate change, and many species are likely to suffer declines or even undergo extinction (Foden et al. 2013). Impacts depend upon inherent sensitivity, adaptability, and the extent of exposure to climatic change (Bird Life International 2013). In addition, climate change can also lead to more frequent wildfires (Silvestrini et al. 2011), which in turn have their own ecological consequences. Among the most prominent biotic effects of global warming are poleward and altitudinal range shifts (evident in some butterflies), changes in precipitation and consequently decreased water availability for animals, complex responses for migratory species (evident in birds by the fact that Red Knot Calidris canutus are becoming smaller, with smaller bills, resulting in decreased survival rates in Africa) as well as changes in phenology and community composition and abundance (WWF 2016). Climate change can also synergise with land use changes and biological invasions to cause the extinction of many bird species (Benning et al. 2002).
There is an immediate necessity to take initiatives to suitably protect wildlife from global warming, and combating climate change and its impacts has been included by UNEP as one of its sustainable development goals (Goal 13) (UNEP 2015).
5.2 Impact of global warming on birds
5.2.1 Extinction
Climate change accelerates the extinction rates of birds (Foden et al. 2013). According to the Intergovernmental Panel on Climate Change (IPCC), 20–30% of species around the globe are likely to be at a high risk of extinction within this century if global mean temperatures exceed 2–3 °C above pre-industrial levels (Barnosky et al. 2011). Indeed, about 90% of some bird populations have already declined due to climate change (WWF 2015). It has been estimated that in South Africa, mid-range climate change will result in a 33–40% extinction rate if birds are unable to shift to new suitable habitats (Thomas et al. 2004). Avian extinction rates in Europe are predicted to be 38, and 72% has been predicted for north-east Australia, if global warming surpasses 2 °C above pre-industrial levels (currently 0.8 °C above) (WWF 2015). In fact, 600–900 terrestrial birds are likely to be extinct by 2100 if the surface temperature rises by 3.5 °C. Additionally, 100–500 bird species may go extinct for every 1° increase in temperature above this value (Sekercioğlu et al. 2012). Birds that survive within narrow environmental ranges are likely to decline the most. Rising temperatures may also help invasive species to outcompete native species (WWF 2015). Migratory, mountain, island, wetland, Arctic, Antarctic and sea birds are highly vulnerable to the problem (WWF 2015). However, up to 83% of all birds highly vulnerable to climate change are not present in the current IUCN Red List of Threatened Species. These species have shown sharp declines in population or a shrinking in geographic range (Foden et al. 2013). Limitations imposed on species range by climatic, ecological and physiological effects of elevation are important determinants of extinction risk (Sekercioglu et al. 2008).
5.2.2 Habitat and distribution
Distribution losses and gains have been forecasted to occur for many bird species due to climate change. An important factor in this regard is the natural history of a given species and its particular habitat requirements. Black-throated sparrows (Amphispiza bilineata) and grey vireos (Vireo vicinior) are likely to gain breeding habitat. But huge losses have been predicted for pygmy nuthatches (Sitta pygmaea), sage thrashers (Oreoscoptes montanus) and Williamson sapsuckers (Sphyrapicus thyroideus). Such losses could result in future population declines. On the whole, such changes may profoundly affect the distribution and viability of many species (US Geological Survey 2014). According to The National Audubon Society Website (2014), climate change could imperil 314 species of birds due to habitat shrinkage. Some of the birds likely to be affected are listed in Table 1.
Climate change might shift the potential distribution of bird species in Europe by about 550 km north-east by the end of the present century. The average distribution of regular European breeding birds has been projected to decrease by a fifth and end up overlapping the present range by only 40%, due to climate change. About 75% of all such species are likely to experience shrinkage in range. If such changes occur, very few areas of habitat with a suitable climate would be left for certain species in Europe (RSPB 2008). In addition, rises in sea-level threaten coastal ecosystems and their biota. Climate change in the Arctic tundra is likely to impact on waterbird breeding habitat (Bird Life International 2013).
Northward latitudinal and elevation shifts are evident in several North American bird species distributions due to climatic changes (King and Finch 2013). It must be added that the constraints of area and resources at higher altitudes could trigger interspecific competition and threaten high elevation residents (Jankowski et al. 2010). Under some circumstances, migrant species might dominate their upland congeners and push them further upwards or lead to their extinction (Jankowski et al. 2010). The shifting of birds to climatically suitable ranges could also be complicated by landscapes which are fragmented or damaged by anthropogenic activities. Island and mountain birds are most vulnerable as these are confined to small habitat patches (WWF 2006). In addition, if climatic changes occur too rapidly for vegetation to respond or take place beyond potential vegetation ranges, then bird populations might be forced to move into unsuitable habitats. As a result, they may suffer from reduced survival and reproduction (Crick 2004). Climate change may also alter community composition by inducing variation in the rate at which species shift their range. Communities could change further due to shifting of constituent species along different trajectories (Gillings et al. 2015). Future indirect climate change effects on birds include alteration of habitats due to rises in sea level, as well as changes in fire regimes, vegetation and land use (WWF 2006). Climate change can also induce drought conditions that elevate the intensity and occurrence of wildfires, which could impact birds by destroying nests and modifying habitats. This is already evident in parts of the western US (King and Finch 2013).
An important effect of climate change propagates through the prevalence of droughts. Open water, vegetation biomass and native over-story trees are important determinants of bird diversity along river reaches. All three factors are reduced when water flow in rivers decreases due to drought. As a result, key sensitive species are affected. This is evident in the Colorado River Delta in Mexico (Hinojosa-Huert et al. 2013). In addition, blackbirds, robins, thrushes and starlings also face difficulty in reaching worms and insects in soil under dry conditions. Moreover, as water bodies dry up, growth of vegetation is hampered. Consequently, the risk of predation increases for wading birds (The Wildlife Trusts Website). Droughts can also lead to changes in salinity and thereby eliminate many insects such as brine flies (Ephydridae sp.) used as food by birds. Insect populations can also decrease due to lower hatching rates under water scarcity (Frost 2014). Dry periods have been linked with bird community changes as well as amphibian and reptilian losses (Pounds et al. 2006). Protected areas help to counteract the effects of droughts by protecting ecosystem services such as water flow and water quality, and conserving habitats (World Bank 2003). They also maintain ecosystem integrity and buffer local climate against drought (Stolton et al. 2008). Hence, the effects of drought are likely to be more pronounced in unprotected areas.
5.2.3 Reproduction
Breeding failure and population reduction are already common in birds due to climate change across the world (Sekercioğlu et al. 2012). In fact, climate change has resulted in complete and unprecedented avian reproduction failures (WWF 2015). This is because breeding performance is dependent upon climate trends (Crick 2004). Changes in climate affect avian metabolism and behaviour, including the ability to mate (Crick 2004). Climate change can also alter display sites for singing birds and also lead to changes in optimal song choice, variance in mating success and predator–prey interactions (Moller 2010).
Breeding success can also be hampered due to the chilling and starvation of young birds under extreme conditions (frozen spills and droughts) occurring due to climate change (Crick 2004). Drought due to climate change also reduces the breeding success of wading birds, as demonstrated in England during the spring of 2011. This could have serious implications because reduced breeding success for more than two consecutive years could threaten vulnerable wading birds such as Redshank (Tringa totanus). Such birds also cannot feed in hard, dry ground (The Wildlife Trusts Website). In addition, temporal partitioning of the breeding period due to changes induced by climate can lead to destructive interspecific competition (Ahola et al. 2007).
Arrival of migratory birds at breeding and over-wintering grounds at the appropriate time is the primary determinant of reproductive success, survival and fitness (Cotton 2003). This is because synchrony between offspring needs and food availability is crucial (Visser et al. 2004). Climate change exerts a negative impact on this aspect by extending growing seasons, changing flowering phenology and altering distribution patterns (Cotton 2003). Increasing spring temperatures have altered vegetation phenology. Consequently, the timing of the food supply for young offspring is not synchronised with reproductive behaviour in some birds. This has led to reproductive failure in some birds such as the great tit in the UK (Visser et al. 1998). Similar effects caused a breeding crash in seabirds in the North Sea around the UK in 2004. This occurred due to the shortage of food resources arising from warming ocean waters and shifts in species that affected the ocean food web (WWF-India 2006). Many birds have advanced their timing of reproduction in response to climate change. Species that do not advance their breeding adequately become mismatched with the availability of food sources and suffer from reproduction failures. The greater the extent of this mismatch, the greater is likely to be the decline in populations (Moller et al. 2008).
However, climate change affects different species differently. This is evident from two groups of sea birds in Siberia, namely planktivorous auklets (Aethia cristatella and Cyclorhynchus psittacula) and piscivorous puffins (Fratercula corniculata and Lunda cirrhata). The former feed on macro-zooplanktons that are favoured in cooler waters. The latter feed on fish that are favoured by warmer waters (warmer water supports mesoplankton that are consumed by fish). Thus, changes in sea surface temperatures affect the reproductive success in these two groups differently (Crick 2004).
Birds can also exhibit individual responses. Pied flycatcher not only advances egg laying to prepone hatching, but also adjusts clutch size and initiation of incubation in response to climate change (Visser et al. 2004). Warmer springs have led to increased egg and clutch sizes in this species in Germany and Finland (Crick 2004). On the other hand, some species attempt to rear a second brood instead of advancing egg laying (Visser et al. 2004).
5.2.4 Incidence of disease
The fact that climate change can lead to increased incidences of some avian pathogens and parasites is exemplified by the spread of West Nile disease (King and Finch 2013). Climate change is also likely to push avian malaria from the tropics and temperate areas towards the north. In fact, it has been estimated that by 2080, the range of this disease will increase and threaten bird populations in currently unaffected areas (Loiseau et al. 2012). Another example is in Hudson Bay, Canada, where mosquito numbers now peak earlier in the spring due to climate change. However, breeding seabirds have not changed their behaviour accordingly. Thus, a combination of heat and mosquitoes has led to greater egg losses and increased adult mortality (Gaston et al. 2002). The effects of climate change are likely to be more severe in Hawaii, where the transmission of avian pox and malaria is affected by fluctuations in temperature and rainfall. Warming of 2 °C can severely increase the risk of incidence of these diseases (Atkinson and LaPointe 2009). Global warming could also move mosquitoes that carry avian malaria to higher elevation forests. Hawaiian honeycreepers (Drepanidinae sp.) would then be exposed to this deadly disease and as a result, face extinction (Benning et al. 2002). Avian pox and influenza demonstrate the impacts that can arise due to range extensions of pathogens and vectors (Atkinson and LaPointe 2009). In addition, climate change can also alter the composition of local species. Consequently, the redistribution of avian influenza across different age groups, species and flyways can take place (Sehgal 2010).
5.2.5 Physical effects of rising temperatures
Hot weather events in the past have resulted in heavy mortality of birds (Finlayson 1932; Keast 1960; Miller 1963). In this regard, it must be mentioned that desert birds are highly vulnerable to increasing temperatures. This is because several such species inhabit areas which are extremely hot in summer and where water resources are scarce and highly unpredictable (African Climate and Development Initiative 2015). Heat waves have led to widespread avian deaths in hot desert environments, and this phenomenon has been projected to increase in intensity, frequency and duration by climate change models. By the 2080s, desert birds are likely to suffer from reduced survival at greater rates during mid-summer (McKechnie and Wolf 2010). With rising air temperatures, increases in avian thermoregulatory water requirements will occur. Mass-specific evaporative water loss rates increase with decreasing body mass, and so vulnerability to acute dehydration will be greater in smaller birds (McKechnie and Wolf 2010). In general, birds have a small body size and are predominantly diurnal. They also make less use of thermally buffered micro-sites. All these characteristics make them highly vulnerable to extreme heat waves. Even in shaded micro-sites, and under inactive conditions, small species can suffer from rates of evaporative water loss that could exceed 5% of body mass per hour. Under such situations, birds could quickly reach and exceed their dehydration tolerance threshold (Wolf and Walsberg 1996). Thus, the smaller the size of a bird, the greater is its vulnerability to heat waves.
5.2.6 Migration
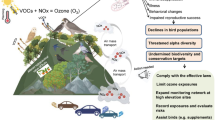
Migration in certain birds has been advancing, and this phenomenon has been correlated with early nesting and hatching due to climate change. Species which do not migrate early undergo population decline. Advances in the time of migration are not common in long-distance migratory birds. Hence, these have been rapidly declining (Gill et al. 2014). European and North American long-distance migratory species have declined. These birds are unable to predict changes and hence have been deprived of food sources during migration. This also explains why some species are unable to advance their arrival date in spring breeding grounds sufficiently (WWF 2006). The onset of migration in some long-distant migrants is determined by endogenous rhythms. Therefore, their migration may not be adequately advanced by climate change, which prepones the phenology of their breeding grounds. Consequently, such species are unable to arrive at their breeding sites at the appropriate time. This is evident in pied flycatcher (Both and Visser 2001). The insectivorous pied flycatcher (family: Muscicapidae) winters in tropical West Africa and breeds in spring in Europe. It is an obligatory long-distant trans-Saharan migrant (Wormworth and Mallon 2006; Harnos et al. 2016). The bird migrates between August and September in Europe (Harnos et al. 2016) and uses South West France, North West Spain and North Portugal as stopover sites for refuelling (Jones 2002). During the recent times, the pied flycatcher is unable to reproduce in presence of adequate food resources. This is because climate change has been accelerating pupation and consequently triggering emergence of caterpillars before the bird breeds (Wormworth and Mallon 2006). Hence, the population of the bird is declining under the influence of climate change (Both et al. 2006). Even though some populations have adopted early migration, they are unable to reach early as their journey is delayed by harsh climatic conditions. This indicates the potential of this bird to adjust itself with climate change, but it also highlights the vulnerability of migration to environmental conditions (Both 2010); Fig. 2.

Source of bird image: BBC Nature (2014)
Process in which climate change causes the population decline of the insectivorous migratory bird pied flycatcher.
Take-off is the migratory phase of birds most sensitive to climate change. However, changes in climate can also drastically affect migration state, routes and direction, as well as stopovers for food and rest (Zalakevicius 2000). Many wetlands are used by migrating birds as staging grounds as well as for breeding. As these dry up due to climate change, many birds will be forced to continue migrating without stopping. They will also be forced to crowd into limited areas, and this can increase the risk of spreading disease (Frost 2014).
The departure of migratory birds from winter areas is dependent on photoperiod. But the availability of food sources depends on plant phenology, a factor which advances due to changes in climate. Therefore, some migratory birds alter their time of breeding to match food supply (Both and Visser 2001). However, due to warmer climate, some Arctic-breeding species do not need to travel long distances to seek milder places that allow them to find food. This has conservation implications because under such circumstances, birds make less use of sites designated for their protection (Lehikoinen et al. 2013). Meanwhile, in some higher altitude sites, initiation of the growing season is determined by the melting of snowpack that has remained unaffected by warmer spring temperatures. The consequent disjunction between altered bird phenology (due to warm spring temperatures) at low altitude and high altitude environments creates problems for altitudinal migrants (Inouye et al. 2000).
6 Assessment of the impact of composite effects through scoring
6.1 Scoring type
In order to assess the composite effects of the four problems for the first time, two scoring types are proposed; viz. (a) impact score (number of general mechanisms through which a particular problem exerts its influence) and (b). hypothesized synergy score (number of mechanisms which are common to any two problems influencing a particular biological aspect. The greater the score, the greater is the synergistic influence of the problems). The scoring method was devised after taking into account the similarities and dissimilarities in the mechanisms of action of the four environmental problems; hence, it is primarily based on these aspects.
6.2 Significance of the scores
Impact score enables simple representation of the cumulative consequence of the four problems. Researchers can utilise this scoring system to obtain an empirical aspect to otherwise qualitative data and thus facilitate statistical analyses. It also provides a unique way to link the problems and can be utilised to develop hypotheses. Synergy scores can be used to deduce synergistic relationships among problems and predict the problems that are likely to escalate when two or more problems occur simultaneously. The basis of both the scores is published scientific literature.
6.3 Assignment of scores
Based on these scores, the five most prominent aspects related to avian biology affected by the environmental problems have been assessed, viz. (a) health, (b) mating, (c) migration, (d) reproduction and (e) general behaviour. The method of scoring for each aspect is described below.
6.3.1 Health impacts
Avian health is affected by emissions, noise and global warming. Emissions exert effects through direct toxicity and indirectly through ecological changes. Noise acts through physiological and behavioural consequences. Global warming leads to diseases and direct heat stress. Since there are two mechanisms of action for each problem, all were awarded impact scores of two (health impact: air pollution = 2, noise pollution = 2, global warming = 2; Table 2). The problems act differently on health, and so I hypothesised that there is no synergistic health effect among the three issues (hypothesised synergy score: air pollution on noise pollution and viceversa = 0, air pollution on global warming and viceversa = 0, noise pollution on global warming and viceversa = 0).
6.3.2 Impacts on mating
Noise and light hamper avian mating calls. Noise modifies timing and frequency, whereas light modifies only the timing of mating calls. Thus, noise acts through two mechanisms and light acts through a single mechanism. Therefore, these were awarded impact scores of 2 and 1, respectively (impact on mating call: noise pollution = 2, light pollution = 1; Table 2).
Light can synergise with only a single action of light (i.e. modification of time, which accounts for half of the negative effect of noise on mating calls). Thus, the hypothesised synergy score of light on noise is 0.5. Noise can increase the entire negative effect of light on mating. Hence, the hypothesised synergy score of noise on light is 1 (hypothesised synergy score of light pollution on noise pollution = 0.5, and noise pollution on light pollution = 1, with respect to mating; Table 3).
6.3.3 Impacts on migration
Light and climate change affect migration through two (disorientation of migrating birds and modification of circadian rhythms) and three (alteration in migration onset, changing habitats by affecting plant phenology, and changing migration characteristics such as state, route and direction) mechanisms, respectively (migration impact: light pollution = 2, global warming = 3; Table 2). No mode of action is common between light and climate change; hence, there is no scope for synergy (hypothesized synergy score: global warming on light pollution and viceversa = 0).
6.3.4 Impacts on reproduction
Reproduction (i.e. breeding success) is affected by light in a single way (it triggers phenological changes through changes in day length), by climate change in two ways (it triggers phenological changes that affect and hampers offspring survival), and by emissions in a single way (it obstructs egg development indirectly by causing carotenoid deficiency in food sources). Hence, the scoring for reproduction impact is light pollution = 1, global warming = 2, air pollution = 1 (Table 2).
Global warming has the maximum potential mechanisms of action (two), out of which one (changing phenology; 50%) is also affected by light. However, global warming can synergise with the entire negative effect of light on reproduction (hypothesised synergy score: global warming on light pollution = 1, light pollution on global warming = 0.5; Table 3). Air pollution has no synergistic effect on either of the problems, and viceversa.
6.3.5 Impacts on behaviour patterns
General behavioural changes are only triggered by light in birds due to changes in circadian rhythms (behaviour impact: light pollution = 1; Table 2).
6.4 Score analysis
The impact scores of the four problems regarding all the aspects are summarised in Table 2. Impact scores for all the five biological variables considered under each of the four environmental problems were summed up to obtain four composite impact scores (CIS). The CISs represented the cumulative effect of each of the four problems (Table 2). The Chi-square test was performed among the CISs to find out whether the intensity of the most dominant effect was significant (Table 2). The total number of mechanisms (T) acting on every biological aspect was calculated (Table 2). For this, the impact scores obtained by every biological variable for all the environmental problems were summed up. The same was converted into percentage for every biological variable. Synergy scores were tabulated after estimation and prediction of synergistic effects (Table 3).
6.5 Results
Among the four environmental problems, light was found to affect the highest number of biological aspects (four; migration, reproduction, mating, and general behaviour), followed by global warming (three: health, migration and reproduction), noise (two: health and mating) and emissions (one: health). However, in terms of overall effects, global warming was identified as the most multi-dimensional, because it exerted its impacts on birds through the highest number (seven) of mechanisms (Table 2). However, there was no significant difference in the number of mechanisms of the problems (χ 2 = 1.84, df = 3, p > 0.05; Table 2).
The physical health of birds is the most affected aspect, as the highest proportion of impacts (31.6%) exerts an influence on this aspect (Table 2). In other words, if all four problems occur simultaneously, poor bird health will be the main symptom. Light does not affect this aspect but it does alter behaviour patterns. It is, however, likely to amplify the effects of noise and global warming on mating and reproduction of birds. The reverse is also true; noise and global warming would intensify its impact on these aspects based on the hypothesised synergy scores (Table 3). Therefore, effects on mating and reproduction could be larger than expected, and the factor by which it will increase will depend upon the intensity of the problems. Under all circumstances, light would accelerate the effects of noise and global warming on mating and reproduction, respectively, by 50%. The problems will in turn enhance the effect of light on the aspects by 100%. The overall composite effect of the four problems on birds is depicted in Fig. 3.

Composite mechanisms of action of the four problems of the terrestrial biosphere (air pollution, light pollution, noise pollution and global warming) on flying birds
7 Discussion
7.1 Individual effects
Emissions, noise, light and global warming exert widespread impacts on birds, but further study is needed in this area. In the context of air pollution, species-specific differences in responses should be considered during planning (Eeva and Lehikoinen 1995). Mammalian faecal matter is a good bio-indicator for atmospheric metal exposure and provides a reasonable and reliable method for long-term pollution assessments (Gupta and Bakre 2013).
Environmental noise affects the quality of territory in songbirds and in this way alters their behavioural ecology (Brumm 2004). Noise can also trigger evolutionary changes in the signal structure of vocalisations. The spectral energy distributions of vocalisations can therefore be shaped to frequency bands that are resistant to noise (Brumm 2004). Under the influence of noise, selection may favour individuals whose vocalisations are unmasked, and species that have greater vocal adjustment ability. Conversely, receivers with hearing better attuned for detecting signals in noisy urban environments might be positively selected (Patricelli and Blickley 2006). In addition, noise can also have reproductive consequences. For instance, clutch size and fledgling mass in great tits are reduced in locations with high traffic noise (Halfwerk et al. 2011). Noise also prominently affects health and mating in birds (as discussed above). Life history and communication traits are important determinants of adaptability of organisms to noise (Kaiser et al. 2010). The effects of anthropogenic noise such as traffic on acoustically communicating animals like birds and amphibians are poorly understood, and there is a lack of sufficient data (Perry et al. 2008; Kaiser et al. 2010). However, traffic noise is likely to have greater negative effects on birds compared to other groups. Mitigation measures in this regard include noise reduction strategies, changes to roadway lighting and vegetation and traffic flow (Kociolek et al. 2011). Suggestions include closure of key roads during the breeding season, as well as the reduction in vehicle speed and/or volume on particular roads (Parris and Schneider 2008).
Light mainly exerts its impacts by altering the biological clock. It triggers changes in diurnal patterns of behaviour and causes diurnal animals to extend their active period, as well as affects endogenous circadian rhythmicity (Dominoni et al. 2013). In addition, it causes disorientation in migrating birds. This fact that has received much attention (Gaston et al. 2013), but additional research is required to develop deeper insight. In order to reduce the negative effects of light on birds, Rodríguez and Rodríguez (2009) suggested reducing artificial lighting sources and changing light signatures, revision of current artificial light sources, switching off lights when unnecessary, rectifying the position of light sources (so that projection into the night sky is eliminated) and increasing public awareness. Suggested mitigation measures also include development of bird-friendly artificial lighting by manipulating the wavelength characteristics of offshore platforms (Poot et al. 2008). The introduction of broad spectrum street lamps can alter the balance of species interactions in artificially lit environments (Davies et al. 2013) and hence should be avoided. Awareness should be generated because mitigation measures cannot be applied without public support. In fact, public awareness and regulations have addressed the challenge of light effectively in some areas (Salmon 2006). In general, lighting of appropriate duration should be maintained and lights turned off after a predetermined hour, and this practice should be properly regulated.
Proper regulations should also be enforced to control emissions, light and noise. Various methods of regulation have been discussed in the literature with regard to emissions. Regulation could also be temporal or seasonal case of light and noise. For example, a species might mate in a particular season, and this could be hampered by noise/light. In this case, noise/light should be reduced during the breeding season in areas where the particular species is present. Light and noise are not toxic. Therefore, if their repulsive nature on animals is better understood, they could be used for managing human-wildlife conflict.
Birds can act as efficient bioindicators of climate change (Crick 2004) because their mobility can lead to rapid and evident changes in distribution and chosen migration paths. Shifts in the timing of important life cycle events (especially reproduction) and in their distribution ranges are the two major ways that birds exhibit strong responses to climate change (WWF 2006). Changes in reproductive phenology in birds are mostly triggered by temperature fluctuations; the same is also true in the case of diseases and pathogens. Flight enables birds to easily move to new surroundings in search of suitable habitats. But the drawback in this regard is that climate change also impacts on migrating birds. This aspect urgently requires attention since climate change is projected to worsen further and cause more drastic consequences. Inappropriate or no phenological responses to climate change and a lack of phonological synchrony with the environment are important factors that determine the extent of the consequence of climate change (Crick 2004).
7.2 Composite effects
My hypothetical analyses identified the heath of birds as the factor most vulnerable to the composite effects of the four problems. However, outcome was not statistically significant. This suggested that the health of birds is at a high risk, but the consequences on the other aspects are not negligible. In other words, all problems could intensify simultaneously and impacts on health would be higher. A significant value of the Chi-square analysis would have led to a different conclusion. Such a value would imply that health effects would solely dominate. The effects on remaining variables would be negligible in comparison to health impacts. Thus, all aspects should be addressed, with special emphasis on health effects. A suitable theory could be developed in this regard, but only after performing experimental studies. This would aid to quantify the effects and appropriate mathematical formulae could then be developed for modelling and quantification of impacts. Most studies consider a particular environmental problem and concentrate on specific animal groups. There is a need to study composite effects be across different ecosystems and consider their synergistic effects.
This review analysed avian biological variables. However, a number of ecological variables (e.g. changes in diversity and prey-predator relations) are also affected by the environmental problems. For example, emissions and noise negatively affect the abundance and variability of birds in forests (Saha and Panday 2011). These could be assessed properly only in the light of prevailing ecological problems such as forest fragmentation, which can also trigger changes in wildlife (Dutta and Dutta 2016). These variables combined with biological variables would give a complete estimation of impacts. More animals should be brought under comparative research into the synergistic effects, and additional ecological problems such as landscape changes considered. Such approaches will likely identify other determinants of the problems. In addition, data generation on species and habitat vulnerabilities, investigation of synergies among threat factors and identification of climate change vulnerabilities are also necessary.
Trees could prove effective against all four problems because trees reflect and dissipitate noise, provide shade and perform ecological functions that regulate the ambient atmosphere. Trees can therefore be used to counteract the effects of emissions, noise and light if adequate and appropriate planning takes place. However, selection of appropriate species is important because trees themselves are affected by light (Chaney 2002). Trees can also be used as mitigation measure against noise because sound is scattered by their structures. Moreover, mechanical vibrations are caused in plants by sound waves. In this way, sound energy is converted into heat (Tang et al. 1986; Nasiri et al. 2015). Species with a low Height to DBH ratio and wide crown are most suitable for decreasing noise. Evergreen trees are preferred for this purpose because they bear leaves throughout the year (Nasiri et al. 2015). Forest stands located at the edges of roads play an important role in controlling noise. Hence, the properties of such trees are important. In this regard, shape and growth form are the main determinants (Spellerberg 1998; Demir et al. 2009; Maleki and Hosseini 2011). Further, forests also act as sinks that moderate global climate; indeed, forest ecosystems worldwide store more carbon than the entire atmosphere (Sheikh et al. 2011).
7.3 Implications
All four problems are accelerating at a high rate, and emissions, noise and global warming have the greatest affect on avian health. Thus, as the issues intensify in the future, bird deaths are likely to escalate and newer health problems would emerge. It is likely that such consequences are already occurring but remain in under the radar due to a lack of studies. In other words, the rate of bird extinction will likely increase and global avian diversity would decrease. Extinction due to health problems will also be accompanied by extinction due to negative impacts on migration. This is because migration is an equally important component of the lifecycle of many bird species. Based on the existing literature, it is difficult to predict whether extinction will be caused more by the former or the latter.
Reproduction is the main factor determining variation and inheritance and is the primary requirement of organic evolution (East 1918). Reproduction is affected by emissions, light and global warming, and worsening of these problems is likely to accelerate avian extinction. Light and noise have severe negative effects on the mating behaviour of many bird species. Alteration in mating behaviour can lead to speciation (Ritchie et al. 1999) and result in evolutionary consequences. Alterations in vocalisations occur due to noise pollution, and this can also have evolutionary implications (Emerson and Boyd 1999). This effect can also extend to aquatic ecosystems, as several fish produce sounds associated with courtship that might be hampered by noise. Courtship and spawning sounds are important factors that relate reproduction in fish and the survival of larvae in oceans (Lobel 2002). Amphibians are another important group that would suffer reproduction failure on account of these problems because, like birds and fish, they also depend on an acoustic mode of reproduction. Indeed, some female tree frogs are attuned to the songs of mates that share an identical number of chromosomes as themselves (Tucker and Gerhardt 2011). In other words, mating behaviour has a genetic basis in such organisms, and this might also be true for birds.
The consequences of the four problems occur simultaneously because the problems themselves often take place at the same time and in the same place. In addition, there are a number of ecological effects such as changes in behaviour and distribution that might alter their mechanisms of action and lead to actual effects that differ markedly from those expected. Both biotic (among effects) and abiotic (among physico-chemical changes triggered in the terrestrial environment) linkage is evident, but a lack of research means that the cumulative effects are difficult to estimate. The present work is a first attempt at addressing this issue, and the findings will hopefully help to formulate future investigations. The theoretical analyses included herein could form the basis of theories and models that could be developed for prediction and impact analyses.
8 Conclusion
The review summarised the negative effects of anthropogenic agents on the biosphere, with the emphasis on how a particular group of fauna (birds) is affected. The consequent impacts are clearly widespread and diverse. In the modern world, environmental problems often occur simultaneously. Therefore, rather than addressing each of the four main problems separately, they should be considered together due to synergy among their effects. This is because the impact of one problem could be more intense in the presence of another. This work only took into account a single group of fauna (birds), and future analyses should include other animal groups so that the overall status of the entire biosphere can be appreciated. Herpetofauna in particular provide much scope in this regard. Ultimately, our findings suggest that development should be planned carefully to protect wildlife from major anthropogenic problems, especially where industrial development and roads are concerned.
References
African climate and development initiative (2015) Hot Birds Programme: predicting the impacts of climate change on desert birds. http://www.acdi.uct.ac.za/research/hot-birds-programme-predicting-impacts-climate-change-desert-birds. Accessed 09 May 2017
Ahola MP, Laaksonen T, Eeva T, Lehikoinen E (2007) Climate change can alter competitive relationships between resident and migratory birds. J Anim Ecol 76:1045–1052
Atkinson CT, LaPointe DA (2009) Introduced avian diseases, climate change, and the future of Hawaiian Honeycreepers. J Avian Med Surg 23(1):53–63
Babisch W, Beule B, Schust M, Kersten N, Ising H (2005) Traffic noise and risk of myocardial infarction. Epidemiology 16:33–40
Barber JR, Crooks KR, Fristrup KM (2010) The costs of chronic noise exposure for terrestrial organisms. Trends Ecol Evol 25:180–189
Barnosky AD, Matzke N, Tomiya S, Wogan GOU, Swartz B et al (2011) Has the earth’s sixth mass extinction already arrived? Nature 471:51–57
BBC Nature (2014) http://www.bbc.co.uk/nature/life/European_Pied_Flycatcher. Accessed 03 Aug 2017
Benning TL, LaPointe D, Atkinson CT, Vitousek PM (2002) Interactions of climate change with biological invasions and land use in the Hawaiian Islands: modeling the fate of endemic birds using a geographic information system. Proc Natl Acad Sci USA 99(22):14246–14249
Berglund B, Lindvall T (eds) (1995) Community noise. World Health Organization, Stockholm
Bickford D, Howard SD, Ng DJJ, Sheridan JA (2010) Impacts of climate change on the amphibians and reptiles of Southeast Asia. Biodivers Conserv 19:1043–1062
Bird Life International (2013) The biological traits of some bird species render them particularly vulnerable to climate change. Presented as part of the Bird Life State of the world’s birds website. http://www.birdlife.org/datazone/sowb/c asestudy/291. Accessed 09 May 2017
Both C (2010) Flexibility of timing of avian migration to climate change masked by environmental constraints en route. Curr Biol. doi:10.1016/j.cub.2009.11.074
Both C, Visser ME (2001) Adjustment to climate change is constrained by arrival date in a long-distance migrant bird. Nature 411:296–298
Both C, Bouwhuis S, Lessells CM, Visser MW (2006) Climate change and population declines in a long-distance migratory bird. Nature 441:81
British Geological Society Website. http://www.bgs.ac.uk/discoveringGeology/climateChange/CCS/man-madeEf fect.html. Accessed 09 May 2017
Brumm H (2004) The impact of environmental noise on song amplitude in a territorial bird. J Anim Ecol 73:434–440
Catchpole CK, Slater PJB (1995) Bird song: themes and variations. Cambridge University Press, Cambridge
Chaney WR (2002) Does night lighting harm trees?. Purdue University Cooperative Extension Service, West Lafayette
Chepesiuk R (2009) Missing the dark: health effects of light pollution. Environ Health Perspect 117(1):A20–A27
Cinzano P, Falchi F, Elvidge CD (2001) The first world atlas of the artificial night sky brightness. Mon Not R Astron Soc 328:689–707
Convention on Biological Diversity (2013) Quick guides to the Aichi Biodiversity Targets (Version 2). https://www.cbd.int/doc/strategic-plan/targets/compilation-quick-guide-en.pdf. Accessed 09 May 2017
Cotton PA (2003) Avian migration phenology and global climate change. Proc Natl Acad Sci 100(21):12219–12222
Crick HQP (2004) The impact of climate change on birds. Ibis 146:48–56
Crino OL, Johnson EE, Blickley JL, Patricelli GL, Breuner CW (2013) Effects of experimentally elevated traffic noise on nestling white-crowned sparrow stress physiology, immune function and life history. J Exp Biol 216:2055–2062
Da Silva A, Valcu M, Kempenaers B (2015) Light pollution alters the phenology of dawn and dusk singing in common European songbirds. Phil Trans R Soc B 370:20140126
Davies TW, Bennie J, Inger R, de Ibarra NH, Gaston KJ (2013) Artificial light pollution: are shifting spectral signatures changing the balance of species interactions? Glob Change Biol 19:1417–1423
Demir M, Küçükosmanolu A, Hasdemir M, Acar HH (2009) Assessment of forest roads and firebreaks in Turkey. Afr J Biotech 8:4553–4561
Diaz M, Parra A, Gallardo C (2011) Serins respond to anthropogenic noise by increasing vocal activity. Behav Ecol. doi:10.1093/beheco/arq210
Dominoni DM, Helm B, Lehmann M, Dowse HB, Partecke J (2013) Clocks for the city: circadian differences between forest and city songbirds. Proc R Soc B 280:20130016
Dudley A, Stolton F (1996) Air pollution and biodiversity. Air Pollution and Climate Secretariat
Dutta A, Dutta H (2016) The biotic implications of forest fragmentation. Int Res J Environ Sci 5(1):58–62
East EM (1918) The role of reproduction in evolution. Am Nat 52(618/619):273–289
Eeva T, Lehikoinen E (1995) Egg shell quality, clutch size and hatching success of the great tit (Parus major) and the pied flycatcher (Ficedula hypoleuca) in an air pollution gradient. Oecologia 102(3):312–323
Eeva T, Lehikoinen E, Nurmi J (1994) Effects of ectoparasites on breeding success of great tits (Parus major) and pied flycatchers (Ficedula hypoleuca) in an air pollution gradient. Can J Zoo 72:624–635
Eeva T, Lehikoinen E, Rönkä M (1998) Air pollution fades the plumage of the great tit. Funct Ecol 12:607–612
Eeva T, Ryömä M, Riihimäki J (2005) Pollution-related changes in diets of two insectivorous passerines. Oecologia 145(4):629–639
Eisenbeis G (2006) Artificial night lighting and insects: attraction of insects to streetlamps in a rural setting in Germany. In: Rich C, Longcore T (eds) Ecological consequences of artificial night lighting. Island Press, Washington, pp 281–304
Emerson SB, Boyd SK (1999) Mating vocalizations of female frogs: control and evolutionary mechanisms. Brain Behav Evol 53(4):187–197
European Commission (2013) Sparrows could be used to monitor air pollution. Science for environment policy (European Commission DG Environment News Alert Service, Edited by SCU), The University of the West of England, Bristol
Finlayson HH (1932) Heat in the interior of South Australia: holocaust of bird-life. S Aust Ornithol 11:158–160
Florida Fish and Wildlife Conservation Commission. Impacts of artificial lighting on birds. http://myfwc.com/conservation/you-conserve/lighting/pollution/birds/. Accessed 09 May 2017
Foden WB, Butchart SHM, Stuart SN, Vié J-C, Akcakaya HR et al (2013) Identifying the world’s most climate change vulnerable species: a systematic trait-based assessment of all birds, amphibians and corals. PLoS ONE 8(6):e65427
Francis C, Ortega C, Cruz A (2009) Noise pollution changes avian communities and species interactions. Curr Biol. doi:10.1016/j.cub.2009.06.052
Francis CD, Ortegaand CP, Cruz A (2011) Noise pollution filters bird communities based on vocal frequency. PLoS ONE 6(11):e27052
Francis CD, Kleist NJ, Ortega CP, Cruz A (2012) Noise pollution alters ecological services: enhanced pollination and disrupted seed dispersal. Proc R Soc B 279:2727–2735
Frost G (2014) How the drought is affecting birds. http://ca.audubon.org/news/how-drought-affecting-birds. Accessed 09 May 2017
Fuller RA, Warren PH, Gaston KJ (2007) Daytime noise predicts nocturnal singing in urban robins. Biol Lett 3:368–370
Gaston AJ, Hipfner JM, Campbell D (2002) Heat and mosquitoes cause breeding failures and adult mortality in an arctic-nesting seabird. Ibis 144(2):185
Gaston KJ, Bennie J, Davies TW, Hopkins J (2013) The ecological impacts of night time light pollution: a mechanistic appraisal. Biol Rev 88:912–927
Gauthreaux SA, Belser CG (2006) Effects of artificial night lighting on migrating birds. In: Rich C, Longcore T (eds) Ecological consequences of artificial night lighting. Island Press, Washington, pp 67–93
Gil D, Honarmand M, Pascual J, Pérez-Mena E, Garcia MC (2015) Birds living near airports advance their dawn chorus and reduce overlap with aircraft noise. Behav Ecol 26(2):435–443
Gill JA, Alves JA, Sutherland WJ, Appleton GF, Potts PM, Gunnarsson TG (2014) Why is timing of bird migration advancing when individuals are not? Proc R Soc B 281:20132161
Gillings S, Balmer DE, Fuller RJ (2015) Directionality of recent bird distribution shifts and climate change in Great Britain. Global Change Biol 21(6):2155–2168
Global Climate Report - Annual 2016. https://www.ncdc.noaa.gov/sotc/global/201613. Accessed 09 May 2017
Gorissen L, Snoeijs T, Van Duyse E, Eens M (2005) Heavy metal pollution affects dawn singing behaviour in a small passerine bird. Oecologia 145:504–509
Gorriz A, Llacuna S, Durfort M, Nadal J (1994) A study of the ciliar tracheal epithelium on passerine birds and small mammals subjected to air pollution: Ultrastructural study. Arch Environ Contam Toxicol 27(1):137–142
Grace J (2004) Understanding and managing the global carbon cycle. J Ecol 92:189–202
Graveland J (1990) Effects of acid precipitation on reproduction in birds. Experientia (Basel) 46:962–970
Gupta V, Bakre P (2013) Mammalian faeces as bioindicator of urban air pollution in captive mammals of Jaipur Zoo. World Environ 3(2):60–65
Halfwerk W, Holleman LJM, Lessells CM, Slabbekoorn H (2011) Negative impact of traffic noise on avian reproductive success. J Appl Ecol 48:210–219
Hare S, Cresswell L, Twigg R, Buchdahl J (2002) Air pollution. Fact sheet series for key-stage 4 and a-level. Atmosphere, Climate and Environment Information Programme, Manchester Metropolitan University, Manchester, UK
Harnos A, Fehérvári P, Piross IS, Karcza Z, Ágh N, Kovács S, Csörgo T (2016) Exploratory analyses of migration timing and morphometrics of the pied flycatcher (Ficedula hypoleuca). Ornis Hung 24(2):109–126
Helm B, Ben-Shlomo R, Sheriff MJ, Hut RA, Foster R, Barnes BM, Dominoni DM (2013) Annual rhythms that underlie phenology: biological time-keeping meets environmental change. Proc R Soc B 280:20130016
Helmholtz Centre for Environmental Research (2013) Artificial lighting and noise alter biorhythms of birds. https://www.ufz.de/index.php?en=32043. Accessed 09 May 2017
Herrera-Montes MI, Aide M (2010) The effect of anthropogenic noise on the calling behavior of amphibians and birds in urban areas of Puerto Rico. J Acoustical Soc Am 127(3):1840
Hinojosa-Huert AO, Nagler PL, Carrillo-Guererro YK, Glenn EP (2013) Effects of drought on birds and riparian vegetation in the Colorado River Delta, Mexico. Ecol Eng 51:275–281
Hörnfeldt B, Nyholm NEI (1996) Breeding performance of Tengmalm’s owl in a heavy metal pollution gradient. J Appl Ecol 33:377–386
Inouye DW, Billy Barr B, Armitage KB, Inouye BD (2000) Climate change is affecting altitudinal migrants and hibernating species. Proc Natl Acad Sci 97(4):1630–1633
International Dark Sky Association. Light pollution effects on wildlife and ecosystems. http://darksky.org/light-pollution/wildlife/. Accessed 09 May 2017
IPCC (2007) Assessment of observed changes and responses in natural and managed systems. In: Rosenzweig C, Casassa G, Karoly DJ, Imeson A, Liu C et al (eds) Climate change 2007: impacts, adaptation and vulnerability, contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press Cambridge, Cambridge
IPCC (2014) Climate Change 2014: Synthesis Report. In: Core Writing Team, Pachauri RK, Meyer LA (eds) Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, Switzerland
Isaksson C, Örnborg J, Stephensen E, Andersson S (2005) Plasma glutathione and carotenoid coloration as potential biomarkers of environmental stress in great tits. Eco Health 2:138–146
Jankowski JE, Robinson SK, Levey DJ (2010) Squeezed at the top: interspecific aggression may constrain elevational ranges in tropical birds. Ecology 91:1877–1884
Jentsch A, Kreyling J, Beierkuhnlein C (2007) A new generation of climate change experiments: events, not trends. Front Ecol Environ 5:365–374
Jones P (2002) Pied flycatcher. In: Wernham C, Toms M, Marchant JH, Clark J, Siriwardena G, Baillie SR (eds) The migration atlas: movements of the birds of Britain and Ireland. British Trust for Ornithology, Thetford, pp 584–586
Kaiser K, Scofield DG, Alloush M, Jones RM, Marczak S, Martineau K, Oliva MA, Narins PM (2010) When sounds collide: the effect of anthropogenic noise on a breeding assemblage of frogs in Belize, Central America. Behaviour 148:215–232
Keast A (1960) Bird adaptations to aridity on the Australian continent. Proc Int Ornithol Congr 12:373–375
Kempenaers B, Borgström P, Loës P, Schlicht E, Valcu M (2010) Artificial night lighting affects dawn song, extra-pair siring success, and lay date in songbirds. Curr Biol. doi:10.1016/j.cub.2010.08.028
Kight CR, Swaddle JP (2011) How and why environmental noise impacts animals: an integrative, mechanistic review. Ecol Lett 14:1052–1061
King D, Finch DM (2013) The effects of climate change on terrestrial birds of North America. U.S. Department of Agriculture, Forest Service, Climate Change Resource Center, Washington
Kociolek AV, Clevenger AP, St. Clair CC, Proppe DS (2011) Effects of road networks on bird populations. Conserv Biol 25:241–249
Lehikoinen A, Jaatinen K, Vähätalo AV, Clausen P, Olivia Crowe O, Deceuninck B et al (2013) Rapid climate driven shifts in wintering distributions of three common waterbird species. Global Change Biol 19(7):2071–2081
Lewis TE (2006) Air pollution and hummingbirds. ORD Newslett 11:1
Llacuna S, Gorriz A, Durfort M, Nadal J (1993) Effects of air pollution on passerine birds and small mammals. Arch Environ Contam Toxicol 1:59–66
Llacuna S, Gorriz A, Sanpera C, Nadal J (1995) Metal accumulation in three species of passerine birds (Emberiza cia, Parus major, and Turdus merula) subjected to air pollution from a coal-fired power plant. Arch Environ Contam Toxicol 28(3):298–303
Llacuna S, Gorriz A, Riera M, Nadal J (1996) Effects of air pollution on hematological parameters in passerine birds. Arch Environ Contam Toxicol 31(1):148–152
Lobel PS (2002) Diversity of fish spawning sounds and the application of passive acoustic monitoring. Bioacoustics 12:286–289
Loiseau C, Harrigan RJ, Cornel AJ, Guers SL, Dodge M, Marzec T et al (2012) First evidence and predictions of Plasmodium transmission in Alaskan bird populations. PLoS ONE 7(9):e44729
Lombard E (1911) Le signe de l’élévation de la voix. Annales Des Maladies de l’Oreille et Du Larynx 37:101–119
Longcore T, Rich C (2004) Ecological light pollution. Front Ecol Environ 2(4):191–198
Lovett RA (2012) How birds are used to monitor pollution. Nature. doi:10.1038/nature.2012.11848
Mahr K, Griggio M, Granatiero M, Hoi H (2012) Female attractiveness affects paternal investment: experimental evidence for male differential allocation in Blue Tits. Front Zoo 19:14
Maleki K, Hosseini SM (2011) Investigation of the effect of leaves, branches and canopies of trees on noise pollution reduction. Ann Environ Sci 5:2–7
McClure CJW, Ware HE, Carlisle J, Kaltenecker G, Barber JR (2013) An experimental investigation into the effects of traffic noise on distributions of birds: avoiding the phantom road. Proc R Soc B Biol Sci 280(1773):20132290. doi:10.1098/rspb.2013.2290
McKechnie AE, Wolf BO (2010) Climate change increases the likelihood of catastrophic avian mortality events during extreme heat waves. Biol Lett 6:253–256
Miller AH (1963) Desert adaptations in birds. In: Proceedings of XIII international ornithological congress, Ithaca, New York, pp 666–674
Mockford EJ, Marshall RC (2009) Effects of urban noise on song and response behaviour in great tits. Proc R Soc B 276:2979–2985
Moller AP (2010) When climate change affects where birds sing. Behav Ecol 12:2012–2017
Moller AP, Rubolini D, Lehikoinen E (2008) Populations of migratory bird species that did not show a phenological response to climate change are declining. Proc Natl Acad Sci 105(42):16195–16200
Morrison ML (1986) Bird populations as indicators of environmental change. Curr Ornithol 3:429–451
Nasiri M, Fallah A, Nasiri B (2015) The effects of tree species on reduction of the rate of noise pollution at the edge of hyrcanian forest roads. Environ Eng Manag J 14(5):1021–1026
Navara KJ, Nelson RJ (2007) The dark side of light at night: physiological, epidemiological and ecological consequences. J Pineal Res 34:215–224
Nyholm NEI, Sawicka-Kapusta K, Swiergosz R, Laczewska B (1995) Effects of environmental pollution on breeding populations of birds in southern Poland. Water Air Soil Pollut 85(2):829–834
O’Connor B, Secades C, Penner J, Sonnenschein R, Skidmore A, Burgess ND, Hutton JM (2015) Earth observation as a tool for tracking progress towards the Aichi Biodiversity Targets. Remote Sens Ecol Conserv 1(1):19–28
Oliver TH, Marshall HH, Morecroft MD, Brereton T, Prudhomme C, Huntingford C (2015) Interacting effects of climate change and habitat fragmentation on drought sensitive butterflies. Nat Clim Change 5:941–945
Parris KM, Schneider A (2008) Impacts of traffic noise and traffic volume on birds of roadside habitats. Ecol Soc 14(1):29
Partecke J, Van’t Hof T, Gwinner E (2005) Underlying physiological control of reproduction in urban and forest-dwelling European Blackbirds Turdus Merula. J Avian Biol 36:295–305
Patricelli GL, Blickley JL (2006) Avian communication in urban noise: causes and consequences of vocal adjustment. Auk 123(3):639–649
Perry G, Buchanan BW, Fisher RN, Salmon M, Wise SE (2008) Effects of artificial night lighting on amphibians and reptiles in urban environments. In: Mitchell JC, Jung Brown RE, Bartholomew B (eds) Urban herpetology. Society for the Study of Amphibians and Reptiles, Salt Lake City, pp 239–256
Poot H, Ens BJ, de Vries H, Donners MAH, Wernand MR, Marquenie JM (2008) Green light for nocturnally migrating birds. Ecol Soc 13(2):47
Pounds JA, Bustamante MR, Coloma LA, Consuegra JA, Fogden MPL et al (2006) Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 439:161–167
Qin K (2015) Birds suffer from air pollution, just like we do. Audubon
Ritchie MG, Halsey EJ, Gleason JM (1999) Drosophila song as a species-specific mating signal and the behavioural importance of Kyriacou & Hall cycles in D. melanogaster song. Anim Behav 58:1–9
Rodríguez A, Rodríguez B (2009) Attraction of petrels to artificial lights in the Canary Islands: effects of the moon phase and age class. Ibis 151:299–310
Rodríguez A, García D, Rodríguez B, Cardona E, Parpal L, Pons P (2015) Artificial lights and seabirds: is light pollution a threat for the threatened Balearic petrels? J Ornithol 156(4):893–902
Rojas LM, McNeil R, Cabana T, Lachapelle P (1999) Diurnal and nocturnal visual capabilities in shorebirds as a function of their feeding strategies. Brain Behav Evol 53:29–43
RSPB (The Royal Society for the Protection of Birds) (2008) Europe’s birds face crisis from climate change. http://www.rspb.org.uk/news/details.aspx?id=tcm:9-180304. Accessed 09 May 2017
Ryan MJ, Brenowitz EA (1985) The role of body size, phylogeny, and ambient noise in the evolution of bird song. Am Nat 126:87–100
Saha DC, Panday PK (2011) Effect of air and noise pollution on species diversity and population density of forest birds at Lalpahari, West Bengal, India. Sci Total Environ 409(24):5328–5336
Sahu SC, Sharma J, Ravindranath NH (2015) Carbon stocks and fluxes for forests in Odisha (India). Trop Ecol 56(1):77–85
Salmon M (2006) Protecting sea turtles from artificial night lighting at florida’s oceanic beaches. In: Rich C, Longcore T (eds) Ecological consequences of artificial night lighting. Island Press, Washington, pp 141–168
Sehgal RNM (2010) Deforestation and avian infectious diseases. J Exp Biol 213(6):955–960
Sekercioglu CH, Schneider SH, Fay JP, Loarie SR (2008) Climate change, elevational range shifts, and bird extinctions. Conserv Biol 22(1):140–150
Sekercioğlu CH, Primack RB, Wormworth J (2012) The effects of climate change on tropical birds. Biol Conserv 148:1–18
Seneviratne SI, Donat MG, Mueller B, Alexander LV (2014) No pause in the increase of hot temperature extremes. Nat Clim Change 4:161–163
Sheikh MA, Kumar M, Bussman RW, Todaria NP (2011) Forest carbon stocks and fluxes in physiographic zones of India. Carbon Balance Manag 6:15
Sillanpää S (2010) Consequences of pollution-related carotenoid limitation in birds. Department of Biology, University of Turku, Finland, Turku
Silvestrini RA, Soares-Filho BS, Nepstad D, Coe M, Rodrigues HO, Assuncao R (2011) Simulating fire regimes in the Amazon in response to climate change and deforestation. Ecol Appl 21(5):1573–1590
Singh BK, Bardgett RD, Smith P, Reay DS (2010) Microorganisms and climate change: terrestrial feedbacks and mitigation options. Nat Rev Microbiol 8:779–790
Slabbekoorn H, Ripmeester EAP (2008) Birdsong and anthropogenic noise: implications and applications for conservation. Mol Ecol 17:72–83
Spellerberg I (1998) Ecological effects of roads and traffic: a literature review. Global Ecol Biogeogr 7:317–333
Stolton S, Dudley N, Randall J (2008) Natural security: protected areas and hazard mitigation. WWF International, Gland
Summers PD, Cunnington GM, Fahrig L (2011) Are the negative effects of roads on breeding birds caused by traffic noise? J Appl Ecol 48:1527–1534
Tang S, Ong P, Woon H (1986) Monte-Carlo simulation of sound-propagation through leafy foliage using experimentally obtained leaf resonance parameters. J Acoustical Soc Am 80:1740–1744
Tennessen JB, Parks SE, Langkilde T (2014) Traffic noise causes physiological stress and impairs breeding migration behaviour in frogs. Conserv Physiol. doi:10.1093/conphys/cou032
The Wildlife Trusts Website. http://www.wildlifetrusts.org/living-landscape/water-and-wetlands/drought-and-wildlife. Accessed 09 May 2017
The National Audubon Society Website (2014) Audubon’s birds and climate change report. http://climate.audubon.org/. Accessed 09 May 2017
Thomas CD, Cameron A, Green RE, Bakkenes M, Beaumont L et al (2004) Extinction risk from climate change. Nature 427:145
Tilman D, Fargione J, Wolff B et al (2001) Forecasting agriculturally driven global environmental change. Science 292:281–284
Tittensor DP, Walpole M, Hill SLL, Boyce DG, Britten GL, Burgess ND (2014) A mid-term analysis of progress toward international biodiversity targets. Science 346(6206):241–244
Tucker MA, Gerhardt HC (2011) Parallel changes in mate-attracting calls and female preferences in auto-triploid tree frogs. Proc R Soc B. doi:10.1098/rspb.2011.1968
UNEP (2015) Sustainable development goals. http://www.undp.org/content/dam/undp/library/corporate/brochure/SDGs_Booklet_Web_En.pdf. Accessed 09 May 2017
US Department of Transportation and Federal Administration (2004) Synthesis of noise effects on wildlife populations. https://www.fhwa.dot.gov/environment/noise/noise_effect_on_wildlife/effects/effects.pdf. Accessed 09 May 2017
US EPA (United States Environmental Protection Agency) (2015) Climate change: basic information. http://www3.epa.gov/climatechange/basics/. Accessed 09 May 2017
US EPA (2016) Climate change: greenhouse gas emissions: greenhouse gases overview https://www3.epa.gov/climatechange/ghgemissions/gases.html2016. Accessed 09 May 2017
US Geological Survey 2014. A few winners but many more losers: South-western bird and reptile distributions to shift as climate changes. http://www.usgs.gov/newsroom/article.asp?ID=3861#.ViPKwNIrJkg. Accessed 09 May 2017
van Langevelde F, Ettema JA, Donners M, Wallis DeVries MF, Groenendijk D (2011) Effect of spectral composition of artificial light on the attraction of moths. Biol Conserv 144:2274–2281
Verheijen FJ (1985) Photopollution: artificial light optic spatial control systems fail to cope with incidents, causations, remedies. Exp Biol 44:1–18
Visser ME, Vannoordwijk AJ, Tinbergen JM, Lessells CM (1998) Warmer springs lead to mistimed reproduction in Great Tits (Parus major). Proc R Soc B 265:1867–1870
Visser ME, Both C, Lambrechts MM (2004) Global climate change leads to mistimed avian reproduction. Adv Ecol Res 35:89–110
Watson MJ, Wilson DR, Mennill DJ (2016) Anthropogenic light is associated with increased vocal activity by nocturnally migrating birds. Condor 118(2):338–344
Whelan CJ, Wenny DG, Marquis RJ (2008) Ecosystem services provided by birds. Ann NY Acad Sci 1134:25–60
Wolf BO, Walsberg GE (1996) Respiratory and cutaneous evaporative water loss at high environmental temperatures in a small bird. J Exp Biol 199:451–457
Wood WE, Yezerinac SM (2006) Song sparrow (Melospiza melodia) song varies with urban noise. Auk 123:650–659
World Bank (2003) Cornerstones for conservation: World Bank assistance for protected areas. World Bank, Washington
World Meteorological Organization Website. Causes of climate change. https://www.wmo.int/pages/themes/climate/causes_of_climate_change.php. Accessed 15 March 2016
Wormworth J, Mallon K (2006) Bird species and climate change. Global status report version 1.0 submitted to World Wide Fund for Nature. Climate Risk Pty Limited, Australia and Climate Risk Europe Limited, London
WWF (World Wide Fund for Nature) (2006) Bird species and climate change: global status report. World Wide Fund for Nature, Gland
WWF (World Wide Fund for Nature) (2015) Climate change impacts on bird species. http://wwf.panda.org/about_our_earth/aboutcc/problems/impacts/species/cc_and_birds/. Accessed 09 May 2017
WWF (World Wide Fund for Nature) (2016) Living Planet Report 2016. Risk and resilience in a new era. WWF International, Gland
WWF-India (World Wide Fund for Nature, India) (2006) Climate change has birds out on a limb. http://www.wwfindia.org/news_facts/?1560. Accessed 09 May 2017
Yorzinski JL, Chisholm S, Byerley SD, Coy JR, Aziz A, Wolf JA, Gnerlich AC (2015) Artificial light pollution increases nocturnal vigilance in peahens. Peer J 3:e1174
Zalakevicius M (2000) Global climate change, bird migration and bird strike problems. International Bird Strike Committee, Amsterdam
Acknowledgements
The help received from Dr. Dean Rea from the UK in improving the standard of English is highly acknowledged.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
There is no conflict of interest.
Rights and permissions
About this article

Cite this article
Dutta, H. Insights into the impacts of four current environmental problems on flying birds. Energ. Ecol. Environ. 2, 329–349 (2017). https://doi.org/10.1007/s40974-017-0075-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40974-017-0075-6