
Abstract
Purpose of the Review
Weather and climate extremes substantially affect global- and regional-scale carbon (C) cycling, and thus spatially or temporally extended climatic extreme events jeopardize terrestrial ecosystem carbon sequestration. We illustrate the relevance of drought and/or heat events (“DHE”) for the carbon cycle and highlight underlying concepts and complex impact mechanisms. We review recent results, discuss current research needs and emerging research topics.
Recent Findings
Our review covers topics critical to understanding, attributing and predicting the effects of DHE on the terrestrial carbon cycle: (1) ecophysiological impact mechanisms and mediating factors, (2) the role of timing, duration and dynamical effects through which DHE impacts on regional-scale carbon cycling are either attenuated or enhanced, and (3) large-scale atmospheric conditions under which DHE are likely to unfold and to affect the terrestrial carbon cycle. Recent research thus shows the need to view these events in a broader spatial and temporal perspective that extends assessments beyond local and concurrent C cycle impacts of DHE.
Summary
Novel data streams, model (ensemble) simulations, and analyses allow to better understand carbon cycle impacts not only in response to their proximate drivers (drought, heat, etc.) but also attributing them to underlying changes in drivers and large-scale atmospheric conditions. These attribution-type analyses increasingly address and disentangle various sequences or dynamical interactions of events and their impacts, including compensating or amplifying effects on terrestrial carbon cycling.
Similar content being viewed by others

Drought, Heat, and the Carbon Cycle: an Introduction and Overview
The Earth’s climate is inherently variable on time scales from seconds to millennia [1], including extreme conditions on time scales of a few days to several years [2]. These extremes affect the biosphere, inducing substantial changes in the functioning of terrestrial ecosystems. These changes further affect regional- and global-scale variability in the C cycle via multiple, sometimes not yet understood, and often highly nonlinear processes [3,4,5], including lagged effects that may perturb ecosystem C cycling from years to decades [6, 7].
For example, during the European heat wave and drought 2003, losses of up to 0.5 PgC were reported ([8]), corresponding to 4 years of terrestrial net carbon uptake [8]. This would be equivalent to half of the annual anthropogenic CO2 emissions (2015 levels) of the 28 European Union member states (i.e., 0.99 PgCFootnote 1), implying that targets and policies for reducing anthropogenic CO2 emissions (e.g., 20% on an EU level by 2020 relative to 1990 levelsFootnote 2) could be simply offset by positive C cycle feedbacks in a more extreme climatic regime. However, Vetter et al. and Bastos et al. [9, 10] estimate lower C releases for the same event based on different baselines, spatio-temporal event definitions (including whether the event is regarded as a drought or heat wave) and datasets, illustrating high methodological uncertainties and sensitivity to baseline choice of such calculations. Hence, improved understanding of the effects of DHEs on the carbon cycle is essential to evaluate the carbon sequestration potential of the terrestrial biosphere.
Complexities of Drought and Heat Extreme Impacts in the Terrestrial Carbon Cycle
Seven years after the European heat wave, a heat event of similar magnitude hit Western Russia [11]. While events of such a magnitude have not been observed at least since the beginning of the instrumental record [11] and both induced reductions in net carbon uptake [8,9,10, 12], the primary pathways through which these events affected terrestrial carbon cycling were not identical: in the case of Europe 2003, direct plant physiological responses to drought stress led to a reduction in gross primary productivity (GPP, for conventions regarding carbon fluxes, please see [13]) that was accompanied by a significant but smaller reduction in respiration [12]. In the case of Russia 2010, forests responded mainly to heat rather than to drought [10], inducing a reduction in GPP but unchanged or even increased (autotrophic) respiration rates [10], even though these respiration estimates are modeling results. Moreover, and unlike Europe 2003, indirect carbon losses via forest and peat fires added an additional C loss equivalent of 78% of the reported GPP losses (i.e., fire emissions of around 70 Tg C [14] and GPP losses in the order of 90 Tg C [10]). These examples illustrate the complexity of pathways (even without considering indirect effects beyond fire, e.g., facilitation of pest and/or pathogen outbreaks) through which even seemingly analogue “drought and heat mega-events” may affect terrestrial carbon cycling.
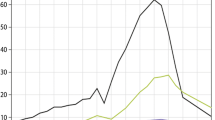
A crucial feature of carbon uptake by terrestrial ecosystems at the global scale is that it provides a negative feedback to anthropogenic climate change, as terrestrial ecosystems absorb around 3.1 PgC year−1 or a fraction of 30% of anthropogenic CO2 emissions from fossil fuel burning and land use changes (2006–2015 [15]). However, this fraction varies strongly from year to year, with coefficients of variation (i.e., year-to-year standard deviation divided by the long-term mean) that range from around 23% in 2006–2015 up to 61% in 1986–1995 (based on [15]). These global-scale differences are associated with large-scale modes of ocean-atmospheric variability (see “Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability” section) and are largely driven by climatic variability [16, 17] and extreme events such as in particular drought, heat, and fire [18]. On an ecosystem scale, the importance of relatively short (“extreme”) periods that cause disproportionate flux reductions can be illustrated by the fact that, for example, in a US Midwest deciduous broadleaf forest (Morgan Monroe State ForestFootnote 3), almost 50% of cumulative negative flux anomalies in GPP occur during less than 5% of the time (Supplementary Fig. 1). In the context of climatic changes in the twenty-first century, heat extremes are projected to increase widely, and even relatively modest changes in the magnitude of events can result in disproportionately large changes in the occurrence frequencies (e.g., [19, 20]). For example, climate changes have induced a roughly 1 °C temperature increase, but this relatively modest increase relative to the total magnitude of events such as Russian heat wave (+ 6° relative to long-term mean monthly temperature) resulted in a tripling of the occurrence frequencies of heat extremes [21]. Because ecosystem carbon cycling responses to DHE often depend on the exceedance of ecophysiological thresholds, e.g., photosynthesis is inhibited at very high temperature values, or heterotrophic respiration limited by water availability [3, 22], these insights on changes in extreme event characteristics might imply crucial consequences for the carbon cycle (see Fig. 1 for an illustrative example).

(Top) A small shift in the mean (PDF1; + 0.2 SD) or in the standard deviation (PDF2; + 15%) compared to a reference distribution (PDF0), for instance induced by large-scale atmospheric phenomena, can lead to (bottom) disproportionate changes in the occurrence frequencies (“return times”) or probabilities (“risk ratio”); here, illustrated for a 2-sigma extreme, the probability of which increases by + 58 or + 80% for PDF1 and PDF2, respectively
Changes in drought and its characteristics are more nuanced, however, and assessments of drought trends typically depend on the type of drought, metrics, and models used and their underlying assumptions and datasets [23,24,25]. Hence, the catchphrase “more frequent, intense, and extreme droughts are to be expected in the future,” stated in the introduction or motivation of many scientific studies, is oversimplified. While thermodynamic arguments indicate that increased heating might indeed amplify droughts via increased atmospheric water demand [25], potentially coinciding with a tendency towards more variable precipitation 26], negative feedbacks via relative humidity, soil moisture limits to further drying [19], and plant physiological responses to increased CO2 [27] can counteract drought. In addition changes in atmospheric circulation can be play an overriding role regionally. Thus, twenty-first century drought projections remain widely uncertain. Nonetheless, in some regions such as Southern Europe trends towards more intense and frequent droughts have been observed historically and are indeed projected with confidence according to the IPCC [19, 28].
Despite these expected changes in climate extremes in the twenty-first century, global land carbon uptake is thought to increase, mainly due to a longer temperate and boreal growing season, higher CO2 availability and N deposition, albeit uncertainties are still large [29]. These uncertainties are related to nitrogen or nutrient limitations [30], the time scales of carbon sequestration, for instance whether faster vegetation growth leads to only transitory or long-term carbon sequestration [31,32,33], and carbon cycle feedbacks via future climate extremes [4].
Concepts, Definitions, and Study Objectives
Weather and climate extremes are typically defined as the “occurrence of a value of a weather or climate variable above (or below) a threshold value near the upper (or lower) ends of the range of observed values of the variable” [19]—although a large suite of definitions exists. In the context of extreme events relevant to the carbon cycle, a strict climatological definition is not always useful. Carbon cycle impacts are not only determined or triggered by univariate climatological variables [3, 4], and thus, various constellations of climate variables that lead to an extreme impact would have to be considered. These issues are currently discussed under the term “compound events” [19, 34] and, although still in its infancy, corresponding suitable multivariate detection methods are being developed [35,36,37]. As an alternative, definitions starting from extreme ecosystem or carbon cycle responses have been proposed [3]. Moreover, ecosystem processes are subject to thresholds, and climatologically “extreme” conditions might be within a plant’s tolerance limits, or vice versa, an ecosystem might experience “extreme” conditions for periods that are sufficiently persistent to not be considered as “extreme” in a statistical sense any more. In summary, differences in the nature of extreme events, the affected ecosystems’ responses and associated time scales, and varying objectives of scientific enquiry might imply that a universally acceptable definition, let alone specific analysis metric or index of C cycle relevant weather and climate extremes might currently not exist.
In this review, we focus on drought and/or heat events (DHEs), thus following a broadly climatological definition, and—in addition—refer to “carbon cycle extremes” (CCEs) as an impact-oriented definition to illustrate global patterns of carbon cycle variability and extremes (in many cases, but not all, related to heat and drought, “Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability” section).
Uncertainties on future developments of DHEs become even more pronounced when considering the impacts on the global carbon cycle. Frank et al. [5] distinguish four categories of carbon cycle impacts: “concurrent” vs. “lagged” impacts according to their differences in response time (i.e., impacts during and after the DHE, respectively), and “direct” vs. “indirect” impacts, with “indirect impacts” being facilitated by the climate extreme but initialized by an external trigger. Here, we focus on direct impacts of DHEs, and obtain a wider spatial (i.e., large-scale atmospheric phenomena) and temporal (the role of “pre-onset” ecosystem conditions) perspective on these impacts. For indirect effects of DHEs, we refer the reader to specialized, in-depth literature: (1) fire following drought and other drivers [38], (2) insect outbreaks and pathogens [39], and (3) changes in litter quality and soil microbial communities following drought (e.g., [40]). Also, Seidl et al. [41] review in detail climate impacts on fire, drought, insect and pathogen disturbance, and their interactions in forests.
Achieving a comprehensive understanding or performing a quantitative meta-study on the effects of DHEs on the global carbon cycle is still a considerable challenge, despite several decades of active research and detailed insights on a plant physiological level [42]. This is because, in addition to issues of definition, the sample size of observed extreme events in situ is small by definition, and vastly different conceptual and methodological approaches are being pursued that reach from local plot-scale experiments to global-scale remote sensing and modeling, including direct/indirect and concurrent/lagged effects, which thus render syntheses difficult.
Here, we aim at an overview of recent developments and emerging research themes on the effects of DHEs on the terrestrial carbon cycle.Footnote 4
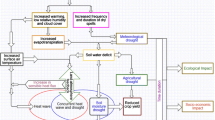
First, we focus on ecophysiological and phenological processes (“Physiological and Phenological Processes Through Which Heat and Drought Affect Ecosystem Carbon Cycling” section), biotic and abiotic factors that mediate carbon cycle impacts (“The Role of Mediating Factors: Ecosystem-Specific Characteristics” section), and effects of timing, duration and “pre-onset effects” that modulate the response of ecosystem carbon cycling to DHEs (“Timing, Duration, Dynamical, and Legacy Effects that Enhance or Dampen Heat/Drought Impacts on the Carbon Cycle” section). Second, we review recent insights on large-scale patterns of atmospheric circulation variability, and its link via DHEs to the terrestrial carbon cycle (“Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability” section). All in all, “Ecosystem-Scale Carbon Cycle Responses to Heat and Drought: Processes and the Role of Mediating Factors” and “Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability” sections indicate that a broader spatial and temporal perspective on DHE events, extending assessments beyond local and concurrent C cycle impacts might be important (Fig. 2). Finally, we summarize potential future research directions and needs for model-data evaluation and conclude with an outlook on the prospects of attributing carbon cycle extremes to underlying drivers of climatic changes and large-scale atmospheric circulation (“Summary and Research Needs” section).

a Conceptual illustration of spatial and temporal mechanisms that can affect ecosystem and regional-scale C-cycle responses to DHEs (as reviewed in the “Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability” and “Summary and Research Needs” sections), and also affect the intensity, probability and spatial patterns of DHEs. The regional carbon balance affected by a DHE event is thus ideally viewed in a broad spatio-temporal continuum (rather than a narrow focus on local and concurrent C-cycle effects of DHEs). b Recent studies address important effects of DHEs on the regional carbon balance that go beyond local and concurrent DHE effects, including both pre-event conditions, legacy effects, and spatial interactions
Ecosystem-Scale Carbon Cycle Responses to Heat and Drought: Processes and the Role of Mediating Factors
Basic plant physiological theory suggests that drought and/or heat adversely affect plant productivity [42] and thus terrestrial ecosystem C cycling. These effects have since been further quantified from ecosystem to global scales via case studies of observed DHE [8, 12], dedicated ecosystem manipulation experiments (e.g., [43]), synthesis studies based on several events across several sites [44, 45] and empirical or process-oriented models [46]. However, it often remains less clear how ecosystem carbon cycle responses to DHE are mediated by individual factors. In this section, we provide an overview of processes that trigger direct effects of DHE on ecosystem carbon cycling (“Physiological and Phenological Processes Through Which Heat and Drought Affect Ecosystem Carbon Cycling” section), ecosystem-specific and abiotic factors that mediate these responses (“The Role of Mediating Factors: Ecosystem-Specific Characteristics” section), and review the role of timing, duration, dynamical and legacy effects that enhance or dampen these impacts (“Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability” section).
Physiological and Phenological Processes Through which Heat and Drought Affect Ecosystem Carbon Cycling
DHE affect ecosystem productivity through both ecophysiological and phenological processes [47], and these events occur throughout all major biomes of the Earth because plants are adapted to thermal and water availability characteristics of their respective environments [48, 49]. DHE synergistically affect plant productivity: on one hand, given a constant specific humidity, with increasing temperature the vapor pressure deficit increases exponentially, which determines the gradient affecting the diffusion of water from the leaves to the atmosphere. On the other hand, soil drought or high VPD leads to stomatal closure and thus reduced photosynthesis and transpiration, reduced evaporative cooling of the leaf and consequently warmer leaves. This implies higher vapor pressure within the leaves and a stronger gradient exacerbating the drought stress.
Extreme heat affects plant physiological processes at cell, leaf, and plant level, including changes in leaf area and leaf development, decreases in photosynthesis and growth, and increased oxidative stress [50]. In the absence of severe drought, a decoupling of photosynthesis (reduced under heat) from transpiration (sustained under heat) was observed recently for Eucalyptus trees in conjunction with rapidly increased leaf thermal tolerance, leading to a vegetation-induced negative feedback to heatwave intensity [51].
At ecosystem level, via physiological and biophysical interactions [52], drought and heat often co-occur, and combined have a stronger effect on vegetation photosynthesis than each of the factors alone, as recently detected at the ecosystem level [45]. For respiration, in particular soil respiration, drought and heat rather have compensating effects: high temperature increases microbial respiration and drought decreases it. Hence, the immediate effect of drought and heat on the overall carbon balance is rather negative as confirmed in many studies [12, 44, 45]. However, the relative role of heat vs. drought can vary. For instance, in a mesic C4 grassland, soil drought was dominant and additional heat effects only had relatively small effects [53], while in the absence of soil drought, extreme heat directly reduced photosynthesis. Current research focuses on the question how respiration components such as autotrophic and heterotrophic respiration are affected [54, 55].
Moreover, DHEs alter the diurnal pattern of ecosystem carbon uptake and release, where the peak of maximum C uptake occurs earlier in the day, followed by an intensified noon and afternoon C uptake depression [56]. This pattern is illustrated in Fig. 3 for a deciduous broadleaf forest in the US Midwest (Morgan Monroe State Forest, Indiana) in the drought and heat year 2012. This “fingerprint” of flux anomalies also demonstrates a “prototypical” reduction of net carbon uptake under DHE and an associated suppression of nighttime respiratory fluxes (Fig. 3). When drought ceases, rewetting of soils typically induces a soil respiration pulse leading to further C losses [57].

a, b Daily temperature and vapor pressure deficit in the year 2012 that featured a very warm spring followed by a hot and dry summer compared to the long-term average at the deciduous broadleaf Morgan Monroe State Forest site (Fluxnet site “US-MMS”). c “Footprint” of half-hourly net ecosystem productivity (NEP) anomalies in the extreme year 2012 relative to long-term averages. The red and gray lines in the footprint illustrate the time of the daily NEP peak in the year 2012 and in the long-term average, respectively
The above processes operate on time scales of minutes to several days. Yet, one has to consider longer timescales to address long-term effects. At a seasonal time scale, phenology, i.e., the development of morphologically visible and functionally relevant characteristic stages of vegetation, such as budburst, leaf unfolding, and leaf coloring is affected [58, 59], leading to altered productivity but also to regional climate feedback via albedo changes for instance [60]. After the DHE, lagged effects on the carbon cycle are induced via changes in plant or soil structural characteristics (e.g., defoliation, reduction in carbohydrate reserves or changes in soil microbial communities), and constitute a crucial topic of present research (see Section 3.3).
The Role of Mediating Factors: Ecosystem-Specific Characteristics
Several ecosystem-specific factors mediate direct physiological and phenological carbon cycle DHE impacts; here, we focus on vegetation type and the role of elevated CO2, but note that for instance nutrient interactions [40], and ecosystem management can also be important.
Vegetation Type
Different vegetation types imply different growth patterns, and accordingly, a fundamentally different C allocation response to water stress and heat between species and ecosystems [42]. For instance on an ecosystem-scale, forests did not show obvious changes in canopy optical characteristics during the European heat wave 2003, but GPP reduction via a distinct physiological response, while non-forests responded fast through canopy changes [61]. These different mechanisms are also consistently reflected by contrasting forest vs. grass responses of transpiration to heat and drought [62], with different response times (grasses respond faster than trees) and feedbacks to local temperature via altered partitioning of sensible vs. latent heat [52]. Deeper tree roots enable access to deeper soil layers, which thus can sustain transpiration, and accordingly photosynthesis, for a longer period, but depending on the availability of fine roots in deeper soil layers [63]. A similar grass-tree dichotomy was observed for ecosystem productivity during the Russian heat wave 2010 [10] and the US drought 2012 [64]. Yet, it remains unclear, if in remote sensing based studies the drought effects in forests are simply not detected, because forests do not change their spectral characteristics and absorption of light as quickly as grasses but rather react physiologically (see, e.g., [61]). Figure 4 illustrates these different response patterns of forests vs. nonirrigated crop/grassland ecosystems to temperature over the contiguous USA, and shows that in summer, forests reach their temperature optimum earlier (at 15–20 °C summer mean temperature) than crops/grasslands (approx. 20–25 °C), but the reduction of GPP after exceeding optimum temperatures is much stronger in the latter. Furthermore, even among grassland sites within a single biome, ecosystem sensitivity to precipitation can vary several fold [66], thus highlighting considerable variation within grasslands. For instance, C4 grasses are adapted to higher temperatures and are typically more drought resistant than C3 grasses, with a slower decline of stomatal conductance, carbon uptake, and transpiration under drought, due to photosynthetic and hydraulic advantages and a deeper root system [67].

Temperature and soil moisture response of 8-daily GPP fluxes from forested and crop/grassland ecosystems in the contiguous USA (25.75–48.75° N, 66.75–106.25° W, 0.5° spatial resolution) in spring and summer. Figure modified after Fig. 7 in Flach et al. (2018, under review in Biogeosciences Discussions, https://doi.org/10.5194/bg-2018-130); GPP data based on the FLUXCOM initiative (http://www.fluxcom.org/) [65]
Within forests, isohydric tree species such as tulip poplar or sugar maple have been shown to strongly regulate stomatal conductance, thus minimizing the risk of cavitation but leading to proportionally high C losses [68, 69]. In contrast, anisohydric species such as oak tended to regulate their stomates only moderately, indicating a greater risk of xylem cavitation and stronger reliance on nighttime refilling of water storage, but only minor reductions in GPP even under severe drought such as in the US 2012 [68].
In addition to inter-specific variation in hydraulic strategies and drought sensitivity, Plant physiological knowledge suggests that plants adapt C allocation strategies to overcome resource limitations most efficiently [70]. In grasslands, experiments indicate a proportionally increased belowground C allocation during drought to optimize supply of available soil water [71] at the expense of aboveground NPP. For trees under moderate drought, similar responses have been hypothesized [72] and observed [73]. In contrast, however, under severe DHE, trees are expected to reduce belowground NPP and to shift to aboveground allocation either due to transport or sink limitations [72], or perhaps because trees might prioritize growth to avoid competitive disadvantages [74]. The latter had been hypothesized based on the observation that the Amazon drought 2010 reduced overall GPP and autotrophic respiration, but not growth (i.e., NPP); trees thus reduced investments in roots and defense components and shifted C allocation aboveground after drought [74]. Conversely, precipitation reduction experiments suggest that Norway spruce shows plasticity of root growth under drought, resulting in enhanced growth of deeper roots under drought at the expense of shallow roots [75]. Hence, plants adapt to drought stress via adjustments in C allocation, which might be key to understanding often differential, sometimes contrasting ecosystem C cycling responses to DHE, but mechanisms vary depending on species type, nutrients, DHE timing, magnitude and duration, among others [72].
Because of these different strategies and susceptibilities of species and plant types to drought and heat stress, changing frequency and intensity of these stressors will also affect vegetation dynamics. For instance, Gherardi and Sala [76] found that under increased precipitation variability shrubs are favored over grasses because of their deeper rooting system, which can catch deeply drained water and thus cope with longer dry spells. In summary, a systematic regional-scale quantification of differences in C cycling responses to DHEs due to different vegetation types is widely lacking, and effects and interactions with local topography, soil and nutrient characteristics [40], or biotic effects such as stand age or species richness [77] are rarely considered.
Effects of Elevated CO2
There is empirical evidence that elevated CO2 can mitigate effects of meteorological drought indirectly because of water savings via reduced stomatal opening under elevated CO2 [78], in addition to anticipated direct “fertilization” effects of elevated CO2 on plant photosynthesis and leaf area in the absence of extreme conditions [79, 80]. Hence, there is potential that increasing CO2 levels alleviate the impacts of meteorological dryness over the coming century indirectly via reduced transpiration [27, 81], but can both increase and reduce temperatures [82]. This is an important example where extreme events interfere with slowly changing conditions. State-of-the-art models in CMIP5 exhibit indeed a relative decrease of extreme drought impacts globally [83]. Yet, both experimental and modeling approaches have their limitations. The experimental approach ignores coupling with the boundary layer, i.e., stomatal closure will lead to less moistening of the air and thus atmospheric demand and potential drought stress. The modeling approaches have certain assumption on stomatal behavior, partly lack the effects of the energy balance on leaf physiology and biological processes with are important for the response to drought.
Further, under high temperatures or very dry conditions, benefits of elevated CO2 might be limited as shown in a mesic grassland experiment [80] and cotton growth experiments [84]. Moreover, combined effects, e.g., of temperature and elevated CO2 are often not simply additive [85]—and thus constitute a considerable challenge for model development and evaluation. In summary, while elevated CO2 is generally expected to alleviate DHE impacts on the carbon cycle via both direct and indirect effects, with anticipated indirect effects most pronounced in water-limited ecosystems [86], accurate regional-scale quantifications of these effects are still lacking, and interactions between variables often remain elusive.
Timing, Duration, Dynamical, and Legacy Effects that Enhance or Dampen Heat/Drought Impacts on the Carbon Cycle
Ecosystem carbon cycle responses depend not only on the intensity of DHEs or mediating factors. Biotic processes follow a distinct phenological cycle, and thus, carbon cycle impacts depend on the initial ecosystem state at the onset of a DHE, its timing [71, 87], and duration. Furthermore, sequences of events [43], drought-heat interactions [52], possibly interacting with transient climatic trends [88], mediate carbon cycle responses to DHE.
The Role of Timing and Duration
Grassland experiments show that spring and summer drought effects can range from being detrimental to growth to almost no impact only due to shifts in spring drought timing of a few weeks—sensitivity to drought appears to be highest at very early development stages and in summer [71, 89]. The impact of high temperatures on vegetation greenness and productivity also depends on its timing with contrasting sensitivities across the year in mid-latitudes [64, 90]. Furthermore, heat and drought effects on carbon fluxes are affected by the duration of events [45], thus lending support to conceptual dose-response relationships as the product of stress intensity and stress duration [5]. Nonetheless, more complex patterns emerge as well: for instance, von Buttlar [45] shows that heat triggers initially enhanced soil respiration, and the latter is reduced only after several weeks of enduring heat, which thus highlights the role of antagonistic mechanisms linked to heat but operating on different time scales.
The Role of Antecedent Conditions and Event Sequences
Consideration of early-season or previous season(s) effects and event sequences on the carbon cycle response to DHE has emerged as a cross-cutting theme in recent studies. Long-term monitoring sites show strong positive autocorrelation of carbon flux anomaly time series, i.e., previous ecosystem states and variability (not necessarily “extremes”) decisively influence carbon flux anomalies today [91]. This link might come about through biotic feedback loops, e.g., via carbohydrate reserves, changes in ecosystem, or canopy structure [6], but also via abiotic pathways such as longer-term effects of previous season soil moisture or snow anomalies [92, 93]. The importance of antecedent effects on vegetation is highest in regions of low total annual precipitation [94], and for instance, moisture conditions in spring can be indeed decisive for summer net carbon exchange in arid ecosystems [95].
Nonetheless, few studies have targeted sequences of events within or across seasons (but see [43]). For instance, a synthesis of a flux tower network and remotely sensed products revealed that losses in net carbon uptake induced by the US drought in 2012 were compensated by high spring temperatures that triggered exceptionally high spring carbon uptake [64]. But, early plant activity in spring might have induced a “carry-over” soil moisture deficit in early summer that exacerbated drought impacts in summer (ibid.). Models suggest that prior to summer drought, increases in spring carbon uptake due to higher temperature and elevated CO2 indeed compensate up to 20% of summer losses in European regions [96], and in the longer term, perhaps, spring water savings due to elevated CO2 and reduced transpiration could alleviate summer drought to some extent [82]. However, whether warmer springs can compensate for carbon losses in summer in the longer term, all other things held constant, also depends on sufficient winter chilling [97] and the absence of late spring frosts during sensitive plant development stages [98], with potential risks associated with both factors depending on individual species and future climate characteristics.
Legacy Effects
After DHE occurred, carbon cycle effects might persist through plant phenological or plant structural changes, e.g., reductions in carbohydrate reserves [91] or defoliation, and mortality [99], and changes in soil structure, communities, and nutrients [40]. Rewetting of soils after drought typically induces a soil respiration peak [100]. According to recent studies legacy effects span 1–2 years in shrubs and grasses [101] and up to four [59, 102] or more years [6] in forests. Recovery times from drought are thought to be longer in tropical and boreal biomes [103] and correlate positively with climatic anomalies post-drought (temperature, precipitation) and GPP amplitude, and negatively with CO2 concentration, among other secondary factors [103]. On a process level, root architecture and C allocation belowground are thought to be important factors for post-drought recovery [100], along with eco-hydrological properties [101] and plant water use strategies [104]. Recently, several mechanisms that could lead to post-drought compensatory C dynamics have been hypothesized: Possibly, (1) increased C allocation belowground might lead to increased C uptake post-drought due to sink control [100], and (2) more diverse plant communities might dampen drought losses through post-drought compensation [105].
Legacy effects may interact with other factors such as insect attacks, e.g., previously less produced defense components may facilitate post-drought insect attacks, but depending on tree species [106]. Legacy effects of moderate drought or warming on soil organismic communities appear rather limited [107]. However, when stress induced by drought or heat exceeds a threshold, mortality can happen, which plays an important role in particular in forests [99] due to their longevity, high C storage and decades to centuries for recovery. Simplified, it has been hypothesized that under these conditions, plants have to choose between carbon starvation and disruption of their water transport system [108], and recent research indicates that carbon starvation occurs rather rarely [109, 110].
In summary, DHE legacies, dynamical mechanisms due to temporal sequences of events, or interactions with long-term trends (e.g., in temperature, CO2, or snow), might enhance or dampen carbon cycle impacts of DHEs, and might require a broader view on DHEs that includes conditions prior to the onset of the actual DHE. Hence, it is essential that ecosystem experiments and data syntheses consider effects of timing, duration, interactions of events, and legacy effects. Because the “sample size” of event constellations in observations or experiments is limited, model ensembles or various sets of simulations might provide a tool to address different spatial or temporal patterns, sequences or interactions of events, and the role of individual factors (e.g., [82, 86, 96]).
Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability
In this section, we provide a brief overview of large-scale patterns of extremes in the carbon cycle and their proximate climatic drivers and discuss the links between different modes of ocean-atmosphere variability and carbon cycle responses.
Year-to-year variability in terrestrial carbon uptake is largely, yet not solely, driven by variations in the interplay of photosynthesis and carbon release processes in tropical semi-arid regions [16, 111]. Accordingly, the largest spatio-temporally integrated extreme anomalies in GPP occur in semi-arid steppe, savanna, or cropland regions at the southern and eastern edges of the Amazon forest, East and South Africa, Eurasian steppe, and Central North America [112]. In Fig. 5, we illustrate the importance of semi-arid regions for extreme reductions in GPP in the hydrologically motivated “Budyko space” [113]. Assuming long-term stationary climate conditions, this framework separates regions of “energy-limited” and “water-limited” evapotranspiration regimes.Footnote 5 A random sample of mean land GPPFootnote 6 shown in Fig. 5 spans a wide range of energy-limited (low dryness and low evaporative index) and water-limited regimes (high dryness and evaporative index approaching unity). However, if these subsampled grid cells are weighted by the occurrence of total GPP losses induced by the largest 1000 negative spatio-temporal extreme events (following [115] for an in-depth methodological description), the bivariate distribution in the Budyko framework is confined to regions that are transitional between water-limited and energy-limited regimes (Fig. 5). This example thus illustrates the importance of semi-arid regions for extreme reductions in the terrestrial carbon cycle. Besides the mere location of GPP reductions in geographical or climatological space, the majority of these events have been linked to water scarcity, fire, or heat [18], thus emphasizing the role of DHEs in these regions that are subject to pronounced land-atmosphere interactions [52].

For any given location, the Budyko framework relates, climatological dryness (expressed as the ratio of annual potential evapotranspiration to actual annual evapotranspiration, PET/ET) to the ratio of annual evapotranspiration to precipitation (i.e., the evaporative index, ET/P). Overview of (1) land grid cells weighted by the GPP losses imposed by the 1000 largest negative extremes (orange lines) and (2) all land grid cells (black lines) in the bivariate Budyko space (i.e., dryness index (PET/P) vs. evaporative index (ET/P)); marginal distributions of the dryness index and evaporative index are shown at top panel and right panel, respectively. Background colors show the difference between bivariate kernel density estimates of (1) GPPextremes weighted grid cells and (2) all land grid cells
Do Large-Scale Patterns of Ocean-Atmosphere Variability Propagate into the Terrestrial Carbon Cycle via Drought, Heat, and Fire?
Large-scale patterns of ocean-atmosphere (or atmosphere-only) variability such as the El Niño-Southern Oscillation (ENSO) phenomenon or the North Atlantic Oscillation (NAO) have been long recognized as dominant sources of interannual climatic variability and extremes (e.g., [116,117,118]). The ENSO phenomenon consists of two opposite extreme phases, El Niño and La Niña [119]. During El Niño, large-scale warming of sea surface temperatures (SSTs) occurs in the Eastern tropical Pacific, which reduces the zonal (east-west) gradient in SSTs and surface pressure (as reflected in the Southern Oscillation Index) in the Pacific and accordingly reduces the strength of the (east-west) Walker circulation, which weakens trade winds and displaces the main convective center from the western to the central Pacific [120]. These conditions are typically associated with a large area of the Southern hemisphere under drought [117] and the occurrence of heat waves regionally [121] with particularly dry conditions over western Pacific regions and wet conditions over the northwest coast of South America. Moreover, El Niño and the opposite La Niña phases, potentially interacting with other ocean-atmosphere modes of variability [122], induce manifold ramifications and “teleconnections” of anomalous, spatio-temporally coherent weather patterns across many regions of the globe [116], thus affecting multiple weather and climate variables simultaneously at any particular location as shown below.
It is widely recognized that ENSO induced climatic variability and extremes propagate into hydrological [123] and ecological [124, 125] systems. ENSO is intimately related to global terrestrial carbon cycling, with El Niño years and associated widespread dryness acting to reduce the magnitude of the residual land sink [15, 111] likely via reductions in NPP, as seen in remote sensing based, diagnostically modeled (i.e., about 40% of global NPP explained by ENSO dynamics [126]) and simulated [127] datasets. The imprint of ENSO on productivity dynamics is also reflected in a significant, but not particularly strong, relationship between changes in tropical GPP due to large-scale carbon cycle extremes and the Southern Oscillation Index (Fig. 6; Pearson correlation on annual aggregates R = 0.56; however, note that the empirically upscaled GPP dataset used here is known to underestimate interannual variability). Consequently, several recent case studies have highlighted patterns and mechanisms that draw a more complex picture, including regional peculiarities:

Southern Oscillation index (SOI) and changes in carbon uptake due to large spatio-temporal extremes following [115], but without detrending, aggregated over the tropical regions of the Earth (30° N–30° S) in the 1982–2011 time period (both time series smoothed using a 12-month centered running mean). Negative SOI values correspond to El Niño episodes
For instance, Cleverly et al. [128] highlight that rather than ENSO alone, a synchronization of different Southern hemisphere climate modes controls drought and precipitation patterns in Australia, reducing primary productivity and providing a basis for wildfires due to hot and dry conditions [129]. Moreover, combinations of different ocean-atmosphere climate modes provide predictive skill of fire activity globally [130]. While variations and extremes in water availability are generally considered as the dominant mechanistic link between ENSO and the terrestrial carbon cycle [65, 126], Liu et al. [131] report different carbon cycle mechanisms in response to the strong 2015/16 El Niño year, which include dryness-driven GPP reductions in South America, heat-induced increases in TER in Africa, and indirect, fire-induced C losses in tropical Asia.
The North Atlantic Oscillation is a meridional dipole in surface pressure that controls the position of the jet stream and winter circulation patterns regionally over Eastern North America and Eurasia [132] with implications for regional-scale carbon cycling: the European net carbon balance is jointly affected by the NAO and a southerly displaced pattern, the East Atlantic oscillation, i.e., for instance with both in their negative phases wet and cool summers tend to increase photosynthesis [133]. These conditions also tend to increase winter precipitation in Southern Europe [133] with positive effects on the C-cycle in these water-limited ecosystems. Negative ENSO phases (La Niña) in conjunction with a negative NAO can induce large losses of carbon uptake over Texas via seasonal-scale drought and heat [134]. Regionally, winter circulation patterns associated with the NAO and Arctic Oscillation that drive winter warm southerly winds are associated with enhancing carbon uptake via reduced snow cover and a longer growing season in the Northern Alps [135].
Despite these insights, a systematic understanding and assessment of large-scale circulation-induced climate extremes, their spatio-temporal variation, and the mechanistic pathways that directly and indirectly affect carbon cycle components, is still lacking. Developing such an understanding is crucial for (at least) two reasons:
First, understanding these patterns might pave the way towards short-term predictive capacity of carbon cycle responses [136] useful for management or planning and towards process understanding of atmospherically coherent spatio-temporal anomaly patterns that can “compensate” each other from a carbon cycle perspective and thus might lead to misinterpretation of continental- to global-scale correlations: for example, Jung et al. [65] showed that on a local scale, water availability is the main driver of carbon uptake. However, spatially compensating patterns and compensation between TER and GPP lead to a weak continental-scale correlation between net carbon uptake and temperature, which thus should not be interpreted in terms of ecosystem-scale processes. Second, projections of changes in circulation characteristics are still uncertain [120, 137]. This is due to partly counteracting processes and feedbacks in a warming climate (e.g., [120] for details regarding ENSO changes), although interactions between large-scale warming and ENSO might lead to more frequent strong El Niño due to reduced zonal temperature gradients [138]. Further, reduced meridional temperature gradients in the Northern hemisphere might contribute to a weakening of the summer circulation in the mid-latitudes, thus favoring persistent heat waves and atmospheric blocking [139], consistent with enhanced interannual variability of the North Atlantic Jet stream latitudinal position [140]. Nonetheless, unavoidable uncertainties related to large-scale circulation are implicit in twenty-first century carbon cycle projections and feedbacks, including the potential of simulated DHE to trigger self-amplifying feedbacks that lead to forest dieback responses [141]. In general, how distortions of atmospheric moisture recycling affects water supply, especially at remote locations, has not been studied extensively. Keys et al. [142] showed that the continental evaporation recycling ratio over land is highly variable, with clear zonal tendencies towards a less oceanic influences, e.g., in the Amazon basin, Eastern Africa, and central Asia. These large-scale patterns bring another aspect into play: the fact that any land use change may also have a feedback to the regional water recycling with obvious consequences for drought occurrence probabilities. However, the effects of such lateral transport issues that are, of course, always triggered by atmospheric transport, and hence, circulation patterns, are not yet studied in detail (but, see [143] for an analysis of remote GPP changes via eco-climatic teleconnections induced through Amazon and Western North America forest loss). In particular, we miss global assessments for the impacts on the C cycle.
Potential future research might disentangle the effects of atmospheric circulation, changes, land use effects (through moisture transport), and direct thermodynamic changes (due to anthropogenic greenhouse gases), all of which affect the carbon cycle via DHEs. This could be achieved by conducting and analyzing dedicated ensemble simulation experiments conditioned to specific atmospheric circulation types, hypothesis about anthropogenic changes, or land-use scenarios (e.g., [96, 144, 145]).
Summary and Research Needs
In this review, we have provided an overview of recent research regarding the ecosystem-scale mechanistic links between DHE and the terrestrial carbon cycle. These mechanisms include direct physiological and phenological responses (“Physiological and Phenological Processes Through Which Heat and Drought Affect Ecosystem Carbon Cycling” section), which are mediated by ecosystem-specific factors such as ecosystem type and CO2 availability (“The Role of Mediating Factors: Ecosystem-Specific Characteristics” section). Furthermore, DHE timing and duration, “pre-onset” ecosystem state and legacy effects play an important role in shaping carbon cycle responses to DHE, as these can directly or via various dynamical effects enhance or dampen DHE impacts (“Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability” section). Moreover, recent research highlights the need to view DHE that affect ecosystem carbon cycling in the context of large-scale atmospheric phenomena (e.g., circulation regimes, long-term trends, remote moisture transport, see the “Large-Scale Carbon Cycle Extremes and Their Link to Ocean-Atmosphere Variability” section), which alter spatial patterns and occurrence probabilities of DHE.
Overall, research on the link between DHE and terrestrial carbon cycling has moved beyond individual “case study type” approaches, enabled by the availability of various multidecadal in situ observations, remote sensing, and experiments (Box 1). Now, systematic syntheses based on monitoring networks [44, 45] are possible, and increasingly allow to pinpoint the role of various contributing and mediating factors in experiments [146] and models [86, 96], and a more clear-sighted view on spatial patterns of DHE events and their link to large-scale atmospheric phenomena [133]. Furthermore, we find that compensatory dynamics that might dampen adverse physiological or phenological DHE impacts were observed recently: these include (1) temporal compensation due to previous events, or interactions with long-term trends [64, 96]; (2) compensating recovery dynamics due to increased rhizosphere C allocation [100] or community diversity effects [105], and (3) spatial compensatory effects in relation to large-scale atmospheric patterns [65]. However, these effects have not been systematically quantified, and dynamics that amplify drought/heat impacts need to be considered as well, e.g., via soil moisture [64], atmospheric boundary layer dynamics [147] or albedo dynamics [60] in the short term, and via tree mortality in the long term [99].
Box 1 The data basis for understanding the impact of climate extremes on the carbon cycle
(a) Ecosystem manipulation experiments improve process understanding Experiments are a natural approach to study the impacts of extreme events as they allow to simulate key attributes of extreme events such as frequency, intensity, timing or duration and at levels of interest from single leaves or plants to entire ecosystems [148]. Controlled experiments have substantially added to our understanding of the effect of climate extremes on ecosystem functioning [149] and CCE mechanisms specifically [150]. For instance, Shi et al. [151] synthesized the effect of extreme drought on net primary productivity (NPP) and heterotrophic respiration (Rh) and found a consistently greater reduction in NPP than Rh across grasslands, and Denton et al. [71] showed that summer drought shifts C allocation towards belowground NPP to secure soil water. Laboratory or field manipulation experiments also include combinations of extreme climatic variables such as coinciding extreme drought and heat [152], but also interactions between long-term trends such as increasing CO2 with extreme drought or heat [51]. However, because laboratory or single-site experiments are often hard to compare and generalize, coordinated distributed experiments [153] are used to systematically investigate the consequences of a specific impact (e.g., drought), or multifactorial experiments across multiple ecosystem types [85]. However, designing these experiments can be practically challenging, as the number of event characteristics (e.g., frequency, intensity, duration, pre-conditioning, multiple extreme variables, etc.) to test for, and co-variates to control for (species composition, plant development stage, nutrient status, “background” meteorology and capturing feedbacks, etc.) are limited. For instance, precipitation manipulation and resulting soil moisture drought often leaves VPD unchanged, and thus possibly induce a bias towards soil moisture as a driver of C-cycle impacts [154]. Moreover, experiments involve often unrealistic step changes in treatment plots [155] and controls plots can be affected by interannual variability in ambient meteorological conditions (see [155] for a detailed overview). Nonetheless, in summary, manipulation experiments are crucial for realizing rigorous statistical testing of the effects of, e.g., drought on productivity or mortality, but also to evaluate process based models under extreme conditions [156]. (b) In situ flux measurements allow to quantify CCE at the ecosystem level An alternative to controlled experiments is to rely on continuous measurements that may coincidentally capture extreme events as “natural experiments.” Contemporary in situ networks experience extreme events because most droughts and heatwaves have a substantial spatial extent and duration [157]. Measurements of net carbon exchanges with the eddy covariance technique have proven to be highly relevant to elucidate the impacts of extremes on the C-cycle, and the derived GPP and terrestrial ecosystem respiration (TER) can further reveal direct impacts of DHE [44, 151]. In the past, many singular events were studied, such as the European summer 2003 [8], the extreme DHE year 2012 in the USA [64] or even multiyear droughts [158]. A recent synthesis study across 11 ecosystem types revealed that integral quantities, such as the duration of DHEs, mainly determine the size of the reductions in gross fluxes [45], and relationships that involve interannual variability and extremes in climate drivers and ecosystem C-cycle responses might be useful to determine C-cycling under climate change [159]. However, several ecosystems that are highly relevant for the global-scale C balance, such as subtropical or tropical forests [160] that have not been so well sampled in the past, but coverage is improving. Nonetheless, incomplete coverage still reduces detection probabilities of regional extremes [157], and in combination with methodological issues such as incomplete energy balance closure [161] especially under nocturnal conditions, and the often “slow-in fast-out” characteristics of carbon exchange under extreme meteorological conditions [162] still challenges improved understanding and interpretation of regional-scale DHE impacts in some ecosystems. (c) Remote sensing and derived data quantify CCE across scales Space-borne remote sensing offers a unique vantage point from which to monitor Earth’s ecosystems and to directly detect and interpret climate variability and extremes and associated CCEs [94]. These global data streams offer the opportunity to assess canopy phenology, stress, and fire at the regional and larger scales [163]. However, remote sensing data are a strong abstraction from the effective carbon cycle impacts, because they measure interaction with electromagnetic radiation and no direct fluxes or stocks. In particular, widely used remotely sensed canopy “greenness” indicators based on differences in spectral reflectances (NDVI, EVI, etc.) are often unable to detect physiological reductions in C uptake under DHE in forest ecosystems if structural properties of the canopy remain unchanged [164, 165], and products derived from such indices such as the MODIS GPP algorithm might fail in precisely detecting drought reductions at specific sites [166]. Nonetheless, recent refinements towards capturing drought responses include the incorporation of information from spatially adjacent pixels in vegetation mosaics [164], or targeting species-specific water use strategies in response to droughts via the use of differences between shaded and sunlit canopy portions [167]. Moreover, a surge of methodological innovations and applications have emerged that have the potential for direct inferences on the C-cycle. In particular, the sun-induced chlorophyll fluorescence (SIF) measurements [168,169,170] provide a promising way for estimating photosynthesis from space, which have already been applied in studying drought-CCEs links, e.g., in the context of regional-scale stress responses [171, 172] or fire [173]. In addition, satellite retrievals of atmospheric CO2 concentrations, such as those from Japan’s GOSAT, NASA’s OCO-2, and China’s TanSat, provide a unique top-down view of the Earth’s carbon cycle and the CO2 concentration retrieval from these spaceborne sensors are particularly suitable to study the CCEs over broad scales [172, 174, 175]. In summary, process understanding of DHE impacts on the C-cycle emerges through multiple lines of evidence, including (a) manipulation experiments, (b) C-cycle monitoring, and (c) remote sensing. Each of these methods involves its unique advantages but also design challenges, which can be partly overcome by integrating and synthesizing data obtained through different methods or scales. |
Opportunities for Model-Data Evaluation and Model Development
Taken together, drought and heat triggers a hierarchy of ecosystem-physiological and thus carbon-cycle responses which occur across a range of time scales, and partly depend on the intensity and partly on the duration of extreme conditions. Modeling this interconnected hierarchy remains a crucial challenge for describing carbon cycle responses to extreme heat and drought. At short time scales (minutes to hours), modeling of energy balance, stomatal responses, internal leaf physiology, and plant-hydraulics are relevant, including an accurate causal representation of vegetation responses to co-limiting stressors such as low soil moisture and high VPD, which can become decoupled at short time scales and in the long term under climate change [154, 176] At seasonal time scale in particular phenology, growth, acclimation, allocation, and repair mechanisms are crucial. On longer time scales (years to decades), still allocation remains very important while competition, adaptation, demographic processes, and thus changes in vegetation composition come into play. It is evident that at longer time scales, biological processes rather than bio-physical processes become decisive, which is backed up by empirical studies (e.g., [177]). Current land-surface models tend to emphasize the physical side, while biological processes are rather modeled ad hoc or empirically. Consequently, many effects seen in experiments are not represented in models or models fail to reproduce these. These include, for instance, C allocation or phenological sensitivity to soil water deficits [178], including variation across soil layers [179], phenology in response to warming treatments, nutrient cycling, and competition dynamics, as recently identified in a grassland experiment [178], photoperiodic controls [179], and leaf dynamics in tropical forests [59]. Furthermore, trait-based approaches have great promise in more realistically modeling biological responses and incorporating diversity of hydraulic traits has already been shown to improve simulated plant responses to water stress [180]. In the long term, plant trait diversity might decrease the sensitivity of vegetation carbon cycling to climate change in models [181]. Yet, it will be important to understand the effect of gradual climate changes versus extreme events in this context.
The wealth of available data allows to integrate models and data using, e.g., pattern-oriented model evaluation strategies [182] to avoid scale mismatches and thus potential misinterpretations of drivers. The latter is crucial, as for instance continental-scale correlations cannot be interpreted on a process level [65]. Model-data integration with a focus on temporal or spatial patterns of drivers of carbon cycle extremes remains rare (but see, e.g., [46, 183]), as large model intercomparison projects are primarily focused on means, seasonal quantities and/or interannual variability [184], and biases in (simulated) climate forcing data affect the magnitude of simulated CCE severely [185, 186]. New experiments are designed to evaluate ecosystem responses to precipitation manipulation that compare model-data responses at the ecosystem scale [148, 187]. Pattern-oriented model-data comparisons on larger scales that use metrics designed for evaluating extremes could yield further insights into drivers and ecosystem responses. For example, present-day models appear to overestimate drought responses [188], perhaps due to compensatory dynamics (“Ecosystem-Scale Carbon Cycle Responses to Heat and Drought: Processes and the Role of Mediating Factors” section), or biological adaptation [189]. Such model-data intercomparisons not only reveal current limitations in models, but also pinpoint possible directions by which models can improve structure and parameterization.
Outlook: Towards an Attribution of Carbon Cycle Extremes to Large-Scale Atmospheric Conditions and Changing Drivers
Improvements in large-scale monitoring abilities offer unprecedented opportunities for cross-scale detection and attribution of the link between DHE and the carbon cycle. These data streams will serve as the basis for designing model-data evaluation and integration approaches targeted to evaluate the DHE-carbon cycle link. However, beyond detecting and evaluating CCEs, attributing these events to their respective proximate drivers (e.g., water availability, temperature, radiation), or linking these to large-scale atmospheric events such as El Niño is a rapidly evolving practice in the carbon cycle community [115, 131]. Nonetheless, in IPCC terminology, “attribution” is defined in a broader way as “the process of evaluating the relative contributions of multiple causal factors to a change or event with an assignment of statistical confidence” [190]. Hence, C-cycle attribution approaches might be extended towards understanding the drivers behind changes in occurrence probabilities of CCE, thus extending attribution of weather and climate extremes (see [191] for an overview) towards C-cycle impacts. For example, anthropogenic climate changes might have contributed both through thermodynamical and dynamical changes to the odds of precipitation extremes, which are typically disentangled via dedicated model simulations ([145], see also [96] for an attribution test case using climate-ecosystem model simulations). Given unprecedented monitoring capabilities and data products that allow careful carbon cycle model evaluation from local to global scales, and the availability of atmospheric model ensembles suitable for these purposes,Footnote 7 we argue that attributing carbon cycle extremes not only to their proximate drivers, but also to the underlying global and local drivers of change might reveal new insights into C-cycle risk imposed by DHEs, their various spatial and/or temporal characteristics and interactions, and thus broader climatic changes. However, this requires that models are evaluated carefully against observations and that uncertainties are clearly stated, and as such attribution of carbon cycle extremes can be seen as an analogous problem to future prediction of carbon cycle extremes.
Notes
Most of the manuscript content is based on a Web of Science search in October 2017 and updated in February 2018 for the terms: (Climate Extremes AND carbon cycle) OR (heat wave AND carbon cycle) OR (drought AND carbon cycle). The resulting 1040 research papers were filtered manually and classified into topics based on the structure of the manuscript, and complemented with classical references during writing.
For any given location of the world, the Budyko framework essentially relates, climatological dryness (expressed as the ratio of annual potential evapotranspiration to actual annual evapotranspiration, PET/ET) to the ratio of annual evapotranspiration to precipitation (i.e., the evaporative index, ET/P).
Here, 5000 randomly subsampled grid cells, weighted by area, from [114]
References
Huybers P, Curry W. Links between annual, Milankovitch and continuum temperature variability. Nature. 2006;441(7091):329–32. https://doi.org/10.1038/nature04745.
Ghil M, Yiou P, Hallegatte S, Malamud BD, Naveau P, Soloviev A, et al. Extreme events: dynamics, statistics and prediction. Nonlinear Proc Geoph. 2011;18(3):295–350. https://doi.org/10.5194/npg-18-295-2011.
Smith MD. An ecological perspective on extreme climatic events: a synthetic definition and framework to guide future research. J Ecol. 2011;99(3):656–63. https://doi.org/10.1111/j.1365-2745.2011.01798.x.
Reichstein M, Bahn M, Ciais P, Frank D, Mahecha MD, Seneviratne SI, et al. Climate extremes and the carbon cycle. Nature. 2013;500(7462):287–95. https://doi.org/10.1038/nature12350.
Frank DA, Reichstein M, Bahn M, Thonicke K, Frank D, Mahecha MD, et al. Effects of climate extremes on the terrestrial carbon cycle: concepts, processes and potential future impacts. Glob Chang Biol. 2015;21(8):2861–80. https://doi.org/10.1111/gcb.12916.
Saatchi S, Asefi-Najafabady S, Malhi Y, Aragao LEOC, Anderson LO, Myneni RB, et al. Persistent effects of a severe drought on Amazonian forest canopy. Proc Natl Acad Sci U S A. 2013;110(2):565–70. https://doi.org/10.1073/pnas.1204651110.
Anderegg WRL, Schwalm C, Biondi F, Camarero JJ, Koch G, Litvak M, et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science. 2015;349(6247):528–32. https://doi.org/10.1126/science.aab1833.
Ciais P, Reichstein M, Viovy N, Granier A, Ogee J, Allard V, et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature. 2005;437(7058):529–33. https://doi.org/10.1038/nature03972.
Vetter M, Churkina G, Jung M, Reichstein M, Zaehle S, Bondeau A, et al. Analyzing the causes and spatial pattern of the European 2003 carbon flux anomaly using seven models. Biogeosciences. 2008;5(2):561–83. https://doi.org/10.5194/bg-5-561-2008.
Bastos A, Gouveia CM, Trigo RM, Running SW. Analysing the spatio-temporal impacts of the 2003 and 2010 extreme heatwaves on plant productivity in Europe. Biogeosciences. 2014;11(13):3421–35. https://doi.org/10.5194/bg-11-3421-2014.
Barriopedro D, Fischer EM, Luterbacher J, Trigo R, Garcia-Herrera R. The hot summer of 2010: redrawing the temperature record map of Europe. Science. 2011;332(6026):220–4. https://doi.org/10.1126/science.1201224.
Reichstein M, Ciais P, Papale D, Valentini R, Running S, Viovy N, et al. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: a joint flux tower, remote sensing and modelling analysis. Glob Chang Biol. 2007;13(3):634–51. https://doi.org/10.1111/j.1365-2486.2006.01224.x.
Chapin FS, Woodwell GM, Randerson JT, Rastetter EB, Lovett GM, Baldocchi DD, et al. Reconciling carbon-cycle concepts, terminology, and methods. Ecosystems. 2006;9(7):1041–50. https://doi.org/10.1007/s10021-005-0105-7.
Guo M, Li J, Xu JW, Wang XF, He HS, Wu L. CO2 emissions from the 2010 Russian wildfires using GOSAT data. Environ Pollut. 2017;226:60–8. https://doi.org/10.1016/j.envpol.2017.04.014.
Le Quere C, Andrew RM, Canadell JG, Sitch S, Korsbakken JI, Peters GP, et al. Global carbon budget 2016. Earth Syst Sci Data. 2016;8(2):605–49. https://doi.org/10.5194/essd-8-605-2016.
Ahlstrom A, Raupach MR, Schurgers G, Smith B, Arneth A, Jung M, et al. The dominant role of semi-arid ecosystems in the trend and variability of the land CO2 sink. Science. 2015;348(6237):895–9. https://doi.org/10.1126/science.aaa1668.
Fang YY, Michalak AM, Schwalm CR, Huntzinger DN, Berry JA, Ciais P, et al. Global land carbon sink response to temperature and precipitation varies with ENSO phase. Environ Res Lett. 2017;12(6):064007. https://doi.org/10.1088/1748-9326/aa6e8e.
Zscheischler J, Mahecha MD, von Buttlar J, Harmeling S, Jung M, Rammig A, et al. A few extreme events dominate global interannual variability in gross primary production. Environ Res Lett. 2014;9(3):035001. https://doi.org/10.1088/1748-9326/9/3/035001.
Seneviratne SI, Nicholls N, Easterling D, Goodess CM, Kanae S, Kossin J, et al. Changes in climate extremes and their impacts on the natural physical environmenty. In: Field CB, Barros V, Stocker TF, et al., editors. Managing the risks of extreme events and disasters to advance climate change adaptation. A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change (IPCC). Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press; 2012. p. 109–230.
Fischer EM, Knutti R. Anthropogenic contribution to global occurrence of heavy-precipitation and high-temperature extremes. Nature Climate Change. 2015;5(6):560. https://doi.org/10.1038/nclimate2617.
Otto FEL, Massey N, van Oldenborgh GJ, Jones RG, Allen MR. Reconciling two approaches to attribution of the 2010 Russian heat wave. Geophys Res Lett. 2012;39:L04702. https://doi.org/10.1029/2011gl050422.
Bahn M, Reichstein M, Dukes JS, Smith MD, McDowell NG. Climate-biosphere interactions in a more extreme world. New Phytol. 2014;202(2):356–9. https://doi.org/10.1111/nph.12662.
Sheffield J, Wood EF, Roderick ML. Little change in global drought over the past 60 years. Nature. 2012;491(7424):435. https://doi.org/10.1038/nature11575.
Orlowsky B, Seneviratne SI. Elusive drought: uncertainty in observed trends and short- and long-term CMIP5 projections. Hydrol Earth Syst Sc. 2013;17(5):1765–81. https://doi.org/10.5194/hess-17-1765-2013.
Trenberth KE, Dai AG, van der Schrier G, Jones PD, Barichivich J, Briffa KR, et al. Global warming and changes in drought. Nat Clim Chang. 2014;4(1):17–22. https://doi.org/10.1038/Nclimate2067.
Pendergrass AG, Knutti R, Lehner F, Deser C, Sanderson BM. Precipitation variability increases in a warmer climate. 2017;7:17966. https://doi.org/10.1038/s41598-017-17966-y.
Swann ALS, Hoffman FM, Koven CD, Randerson JT. Plant responses to increasing CO2 reduce estimates of climate impacts on drought severity. Proc Natl Acad Sci U S A. 2016;113(36):10019–24. https://doi.org/10.1073/pnas.1604581113.
Stahl K, Hisdal H, Hannaford J, Tallaksen LM, van Lanen HAJ, Sauquet E, et al. Streamflow trends in Europe: evidence from a dataset of near-natural catchments. Hydrol Earth Syst Sc. 2010;14(12):2367–82. https://doi.org/10.5194/hess-14-2367-2010.
Friedlingstein P, Meinshausen M, Arora VK, Jones CD, Anav A, Liddicoat SK, et al. Uncertainties in CMIP5 climate projections due to carbon cycle feedbacks. J Clim. 2014;27(2):511–26. https://doi.org/10.1175/Jcli-D-12-00579.1.
Zaehle S, Dalmonech D. Carbon-nitrogen interactions on land at global scales: current understanding in modelling climate biosphere feedbacks. Curr Opin Env Sust. 2011;3(5):311–20. https://doi.org/10.1016/j.cosust.2011.08.008.
Körner C. Biosphere responses to CO2 enrichment. Ecol Appl. 2000;10(6):1590–619. https://doi.org/10.2307/2641226.
Friend AD, Lucht W, Rademacher TT, Keribin R, Betts R, Cadule P, et al. Carbon residence time dominates uncertainty in terrestrial vegetation responses to future climate and atmospheric CO2. Proc Natl Acad Sci U S A. 2014;111(9):3280–5. https://doi.org/10.1073/pnas.1222477110.
Körner C. A matter of tree longevity. Science. 2017;355(6321):130–1. https://doi.org/10.1126/science.aaal2449.
Leonard M, Westra S, Phatak A, Lambert M, van den Hurk B, McInnes K, et al. A compound event framework for understanding extreme impacts. Wires Clim Change. 2014;5(1):113–28. https://doi.org/10.1002/wcc.252.
Mahony CR, Cannon AJ, Wang TL, Aitken SN. A closer look at novel climates: new methods and insights at continental to landscape scales. Glob Chang Biol. 2017;23(9):3934–55. https://doi.org/10.1111/gcb.13645.
Bevacqua E, Maraun D, Haff IH, Widmann M, Vrac M. Multivariate statistical modelling of compound events via pair-copula constructions: analysis of floods in Ravenna (Italy). Hydrol Earth Syst Sc. 2017;21(6):2701–23. https://doi.org/10.5194/hess-21-2701-2017.
Flach M, Gans F, Brenning A, Denzler J, Reichstein M, Rodner E, et al. Multivariate anomaly detection for earth observations: a comparison of algorithms and feature extraction techniques. Earth Syst Dynam. 2017;8(3):677–96. https://doi.org/10.5194/esd-8-677-2017.
Williams AP, Abatzoglou JT. Recent advances and remaining uncertainties in resolving past and future climate effects on global fire activity. Curr Climate Change Rep. 2016;2(1):1–14. https://doi.org/10.1007/s40641-016-0031-0
Jactel H, Petit J, Desprez-Loustau ML, Delzon S, Piou D, Battisti A, et al. Drought effects on damage by forest insects and pathogens: a meta-analysis. Glob Chang Biol. 2012;18(1):267–76. https://doi.org/10.1111/j.1365-2486.2011.02512.x.
Schlesinger WH, Dietze MC, Jackson RB, Phillips RP, Rhoades CC, Rustad LE, et al. Forest biogeochemistry in response to drought. Glob Chang Biol. 2016;22(7):2318–28. https://doi.org/10.1111/gcb.13105.
Seidl R, Thom D, Kautz M, Martin-Benito D, Peltoniemi M, Vacchiano G, et al. Forest disturbances under climate change. Nat Clim Chang. 2017;7(6):395–402. https://doi.org/10.1038/Nclimate3303.
Larcher W. Physiological plant ecology : ecophysiology and stress physiology of functional groups. 4th edn. Berlin; New York: Springer; 2003.
Jentsch A, Kreyling J, Elmer M, Gellesch E, Glaser B, Grant K, et al. Climate extremes initiate ecosystem-regulating functions while maintaining productivity. J Ecol. 2011;99(3):689–702. https://doi.org/10.1111/j.1365-2745.2011.01817.x.
Schwalm CR, Williams CA, Schaefer K, Arneth A, Bonal D, Buchmann N, et al. Assimilation exceeds respiration sensitivity to drought: a FLUXNET synthesis. Glob Chang Biol. 2010;16(2):657–70. https://doi.org/10.1111/j.1365-2486.2009.01991.x.
von Buttlar J, Zscheischler J, Rammig A, Sippel S, Reichstein M, Knohl A, et al. Impacts of droughts and extreme temperature events on gross primary production and ecosystem respiration: a systematic assessment across ecosystems and climate zones. Biogeosci Discuss. 2017;2017:1–39. https://doi.org/10.5194/bg-2017-393.
Zscheischler J, Michalak AM, Schwalm C, Mahecha MD, Huntzinger DN, Reichstein M, et al. Impact of large-scale climate extremes on biospheric carbon fluxes: an intercomparison based on MsTMIP data. Global Biogeochem Cy. 2014;28(6):585–600. https://doi.org/10.1002/2014gb004826.
Xia JY, Niu SL, Ciais P, Janssens IA, Chen JQ, Ammann C, et al. Joint control of terrestrial gross primary productivity by plant phenology and physiology. Proc Natl Acad Sci U S A. 2015;112(9):2788–93. https://doi.org/10.1073/pnas.1413090112.
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, et al. Global convergence in the vulnerability of forests to drought. Nature. 2012;491(7426):752. https://doi.org/10.1038/nature11688.
O'Sullivan OS, Heskel MA, Reich PB, Tjoelker MG, Weerasinghe LK, Penillard A, et al. Thermal limits of leaf metabolism across biomes. Glob Chang Biol. 2017;23(1):209–23. https://doi.org/10.1111/gcb.13477.
Teskey R, Wertin T, Bauweraerts I, Ameye M, McGuire MA, Steppe K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015;38(9):1699–712. https://doi.org/10.1111/pce.12417.
Drake JE, Tjoelker MG, Vårhammar A, Medlyn Belinda E, Reich PB, Leigh A, et al. Trees tolerate an extreme heatwave via sustained transpirational cooling and increased leaf thermal tolerance. Global Change Biol. 2018;24:2390–402. https://doi.org/10.1111/gcb.14037.
Seneviratne SI, Corti T, Davin EL, Hirschi M, Jaeger EB, Lehner I, et al. Investigating soil moisture-climate interactions in a changing climate: a review. Earth-Sci Rev. 2010;99(3–4):125–61. https://doi.org/10.1016/j.earscirev.2010.02.004.
Hoover DL, Knapp AK, Smith MD. The immediate and prolonged effects of climate extremes on soil respiration in a mesic grassland. J Geophys Res-Biogeo. 2016;121(4):1034–44. https://doi.org/10.1002/2015jg003256.
Balogh J, Papp M, Pinter K, Foti S, Posta K, Eugster W, et al. Autotrophic component of soil respiration is repressed by drought more than the heterotrophic one in dry grasslands. Biogeosciences. 2016;13(18):5171–82. https://doi.org/10.5194/bg-13-5171-2016.
Wang X, Liu LL, Piao SL, Janssens IA, Tang JW, Liu WX, et al. Soil respiration under climate warming: differential response of heterotrophic and autotrophic respiration. Glob Chang Biol. 2014;20(10):3229–37. https://doi.org/10.1111/gcb.12620.
Baldocchi D. Measuring and modelling carbon dioxide and water vapour exchange over a temperate broad-leaved forest during the 1995 summer drought. Plant Cell Environ. 1997;20(9):1108–22. https://doi.org/10.1046/j.1365-3040.1997.d01-147.x.
Jarvis P, Rey A, Petsikos C, Wingate L, Rayment M, Pereira J, et al. Drying and wetting of Mediterranean soils stimulates decomposition and carbon dioxide emission: the “Birch effect”. Tree Physiol. 2007;27(7):929–40. https://doi.org/10.1093/treephys/27.7.929.
Ma XL, Huete A, Moran S, Ponce-Campos G, Eamus D. Abrupt shifts in phenology and vegetation productivity under climate extremes. J Geophys Res-Biogeo. 2015;120(10):2036–52. https://doi.org/10.1002/2015jg003144.
Wu J, Albert LP, Lopes AP, Restrepo-Coupe N, Hayek M, Wiedemann KT, et al. Leaf development and demography explain photosynthetic seasonality in Amazon evergreen forests. Science. 2016;351(6276):972–6. https://doi.org/10.1126/science.aad5068.
Ma SX, Pitman AJ, Lorenz R, Kala J, Srbinovsky J. Earlier green-up and spring warming amplification over Europe. Geophys Res Lett. 2016;43(5):2011–8. https://doi.org/10.1002/2016gl068062.
Zhang Y, Xiao XM, Zhou S, Ciais P, McCarthy H, Luo YQ. Canopy and physiological controls of GPP during drought and heat wave. Geophys Res Lett. 2016;43(7):3325–33. https://doi.org/10.1002/2016gl068501.
Teuling AJ, Seneviratne SI, Stockli R, Reichstein M, Moors E, Ciais P, et al. Contrasting response of European forest and grassland energy exchange to heatwaves. Nat Geosci. 2010;3(10):722–7. https://doi.org/10.1038/NGEO950.
Breda N, Huc R, Granier A, Dreyer E. Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Ann Forest Sci. 2006;63(6):625–44. https://doi.org/10.1051/forest:2006042.
Wolf S, Keenan TF, Fisher JB, Baldocchi DD, Desai AR, Richardson AD, et al. Warm spring reduced carbon cycle impact of the 2012 US summer drought. Proc Natl Acad Sci. 2016;113(21):5880–85. https://doi.org/10.1073/pnas.1519620113.
Jung M, Reichstein M, Schwalm CR, Huntingford C, Sitch S, Ahlstrom A, et al. Compensatory water effects link yearly global land CO2 sink changes to temperature. Nature. 2017;541(7638) https://doi.org/10.1038/nature20780.
Knapp AK, Carroll CJW, Denton EM, La Pierre KJ, Collins SL, Smith MD. Differential sensitivity to regional-scale drought in six central US grasslands. Oecologia. 2015;177(4):949–57. https://doi.org/10.1007/s00442-015-3233-6.
Taylor SH, Ripley BS, Martin T, De-Wet LA, Woodward FI, Osborne CP. Physiological advantages of C-4 grasses in the field: a comparative experiment demonstrating the importance of drought. Glob Chang Biol. 2014;20(6):1992–2003. https://doi.org/10.1111/gcb.12498.
Roman DT, Novick KA, Brzostek ER, Dragoni D, Rahman F, Phillips RP. The role of isohydric and anisohydric species in determining ecosystem-scale response to severe drought. Oecologia. 2015;179(3):641–54. https://doi.org/10.1007/s00442-015-3380-9.
Yi K, Dragoni D, Phillips RP, Roman DT, Novick KA. Dynamics of stem water uptake among isohydric and anisohydric species experiencing a severe drought. Tree Physiol. 2017;37(10):1379–92. https://doi.org/10.1093/treephys/tpw126.
Bloom AJ, Chapin FS, Mooney HA. Resource limitation in plants—an economic analogy. Annu Rev Ecol Syst. 1985;16:363–92. https://doi.org/10.1146/annurev.es.16.110185.002051.
Denton EM, Dietrich JD, Smith MD, Knapp AK. Drought timing differentially affects above- and belowground productivity in a mesic grassland. Plant Ecol. 2017;218(3):317–28. https://doi.org/10.1007/s11258-016-0690-x.
Sevanto S, Dickman LT. Where does the carbon go?-Plant carbon allocation under climate change. Tree Physiol. 2015;35(6):581–4. https://doi.org/10.1093/treephys/tpv059.
Blessing CH, Werner RA, Siegwolf R, Buchmann N. Allocation dynamics of recently fixed carbon in beech saplings in response to increased temperatures and drought. Tree Physiol. 2015;35(6):585–98. https://doi.org/10.1093/treephys/tpv024.
Doughty CE, Metcalfe DB, Girardin CAJ, Amezquita FF, Cabrera DG, Huasco WH, et al. Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature. 2015;519(7541):78–U140. https://doi.org/10.1038/nature14213.
Børja I, Godbold DL, Světlík J, Nagy NE, Gebauer R, Urban J, et al. Norway spruce fine roots and fungal hyphae grow deeper in forest soils after extended drought. In: Soil Biological Communities and Ecosystem Resilience. Berlin: Springer; 2017. p. 123–42.
Gherardi LA, Sala OE. Enhanced precipitation variability decreases grass- and increases shrub-productivity. Proc Natl Acad Sci U S A. 2015;112(41):12735–40. https://doi.org/10.1073/pnas.1506433112.
Musavi T, Migliavacca M, Reichstein M, Kattge J, Wirth C, Black TA, et al. Stand age and species richness dampen interannual variation of ecosystem-level photosynthetic capacity. Nat Ecol Evol. 2017;1:0048. https://doi.org/10.1038/s41559-016-0048.
Roy J, Picon-Cochard C, Augusti A, Benot ML, Thiery L, Darsonville O, et al. Elevated CO2 maintains grassland net carbon uptake under a future heat and drought extreme. Proc Natl Acad Sci U S A. 2016;113(22):6224–9. https://doi.org/10.1073/pnas.1524527113.
Zhu ZC, Piao SL, Myneni RB, Huang MT, Zeng ZZ, Canadell JG, et al. Greening of the earth and its drivers. Nature Climate Change. 2016;6(8):791. https://doi.org/10.1038/Nclimate3004.
Obermeier WA, Lehnert LW, Kammann CI, Muller C, Grunhage L, Luterbacher J, et al. Reduced CO2 fertilization effect in temperate C3 grasslands under more extreme weather conditions. Nature Climate Change. 2017;7(2):137. https://doi.org/10.1038/Nclimate3191.
Leuzinger S, Zotz G, Asshoff R, Körner C. Responses of deciduous forest trees to severe drought in Central Europe. Tree Physiol. 2005;25(6):641–50. https://doi.org/10.1093/treephys/25.6.641.
Lemordant L, Gentine P, Stefanon M, Drobinski P, Fatichi S. Modification of land-atmosphere interactions by CO2 effects: implications for summer dryness and heat wave amplitude. Geophys Res Lett. 2016;43(19):10240–8. https://doi.org/10.1002/2016gl069896.
Zscheischler J, Reichstein M, von Buttlar J, Mu MQ, Randerson JT, Mahecha MD. Carbon cycle extremes during the 21st century in CMIP 5 models: future evolution and attribution to climatic drivers. Geophys Res Lett. 2014;41(24):8853–61. https://doi.org/10.1002/2014gl062409.
Broughton KJ, Smith RA, Duursma RA, Tan DKY, Payton P, Bange MP, et al. Warming alters the positive impact of elevated CO2 concentration on cotton growth and physiology during soil water deficit. Funct Plant Biol. 2017;44(2):267–78. https://doi.org/10.1071/Fp16189.
Dieleman WIJ, Vicca S, Dijkstra FA, Hagedorn F, Hovenden MJ, Larsen KS, et al. Simple additive effects are rare: a quantitative review of plant biomass and soil process responses to combined manipulations of CO2 and temperature. Glob Chang Biol. 2012;18(9):2681–93. https://doi.org/10.1111/j.1365-2486.2012.02745.x.
Fatichi S, Leuzinger S, Paschalis A, Langley JA, Barraclough AD, Hovenden MJ. Partitioning direct and indirect effects reveals the response of water-limited ecosystems to elevated CO2. Proc Natl Acad Sci U S A. 2016;113(45):12757–62. https://doi.org/10.1073/pnas.1605036113.
De Boeck HJ, Dreesen FE, Janssens IA, Nijs I. Whole-system responses of experimental plant communities to climate extremes imposed in different seasons. New Phytol. 2011;189(3):806–17. https://doi.org/10.1111/j.1469-8137.2010.03515.x.
Beierkuhnlein C, Thiel D, Jentsch A, Willner E, Kreyling J. Ecotypes of European grass species respond differently to warming and extreme drought. J Ecol. 2011;99(3):703–13. https://doi.org/10.1111/j.1365-2745.2011.01809.x.
Darenova E, Holub P, Krupkova L, Pavelka M. Effect of repeated spring drought and summer heavy rain on managed grassland biomass production and CO2 efflux. J Plant Ecol. 2017;10(3):476–85. https://doi.org/10.1093/jpe/rtw058.
Sippel S, Zscheischler J, Reichstein M. Ecosystem impacts of climate extremes crucially depend on the timing. Proc Natl Acad Sci U S A. 2016;113(21):5768–70. https://doi.org/10.1073/pnas.1605667113.
Desai AR. Influence and predictive capacity of climate anomalies on daily to decadal extremes in canopy photosynthesis. Photosynth Res. 2014;119(1–2):31–47. https://doi.org/10.1007/s11120-013-9925-z.
Buermann W, Bikash PR, Jung M, Burn DH, Reichstein M. Earlier springs decrease peak summer productivity in North American boreal forests. Environ Res Lett. 2013;8(2):024027. https://doi.org/10.1088/1748-9326/8/2/024027.
Papagiannopoulou C, Miralles DG, Dorigo WA, Verhoest NEC, Depoorter M, Waegeman W. Vegetation anomalies caused by antecedent precipitation in most of the world. Environ Res Lett. 2017;12(7):074016. https://doi.org/10.1088/1748-9326/aa7145.
Seddon AWR, Macias-Fauria M, Long PR, Benz D, Willis KJ. Sensitivity of global terrestrial ecosystems to climate variability. Nature. 2016;531(7593):229. https://doi.org/10.1038/nature16986.
Kwon H, Pendall E, Ewers BE, Cleary M, Naithani K. Spring drought regulates summer net ecosystem CO2 exchange in a sagebrush-steppe ecosystem. Agric For Meteorol. 2008;148(3):381–91. https://doi.org/10.1016/j.agrformet.2007.09.010.
Sippel S, Forkel M, Rammig A, Thonicke K, Flach M, Heimann M, et al. Contrasting and interacting changes in simulated spring and summer carbon cycle extremes in European ecosystems. Environ Res Lett. 2017;12(7):075006. https://doi.org/10.1088/1748-9326/aa7398.
Körner C, Basler D. Phenology under global warming. Science. 2010;327(5972):1461–2. https://doi.org/10.1126/science.1186473.
Hufkens K, Friedl MA, Keenan TF, Sonnentag O, Bailey A, O'Keefe J, et al. Ecological impacts of a widespread frost event following early spring leaf-out. Glob Chang Biol. 2012;18(7):2365–77. https://doi.org/10.1111/j.1365-2486.2012.02712.x.
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecol Manag. 2010;259(4):660–84. https://doi.org/10.1016/j.foreco.2009.09.001.
Hagedorn F, Joseph J, Peter M, Luster J, Pritsch K, Geppert U, et al. Recovery of trees from drought depends on belowground sink control. Nat Plants. 2016;2(8):16111. https://doi.org/10.1038/Nplants.2016.111.
Wu X, Liu H, Li X, Ciais P, Babst F, Guo W, et al. Differentiating drought legacy effects on vegetation growth over the temperate Northern Hemisphere. Global Change Biol:n/a-n/a. https://doi.org/10.1111/gcb.13920.
Anderegg WRL, Plavcova L, Anderegg LDL, Hacke UG, Berry JA, Field CB. Drought's legacy: multiyear hydraulic deterioration underlies widespread aspen forest die-off and portends increased future risk. Glob Chang Biol. 2013;19(4):1188–96. https://doi.org/10.1111/gcb.12100.
Schwalm CR, Anderegg WRL, Michalak AM, Fisher JB, Biondi F, Koch G, et al. Global patterns of drought recovery. Nature. 2017;548(7666):202. https://doi.org/10.1038/nature23021.
Peltier DMP, Fell M, Ogle K. Legacy effects of drought in the southwestern United States: a multi-species synthesis. Ecol Monogr. 2016;86(3):312–26. https://doi.org/10.1002/ecm.1219.
Wagg C, O'Brien MJ, Vogel A, Scherer-Lorenzen M, Eisenhauer N, Schmid B, et al. Plant diversity maintains long-term ecosystem productivity under frequent drought by increasing short-term variation. Ecology. 2017;98(11):2952–61.
Rouault G, Candau JN, Lieutier F, Nageleisen LM, Martin JC, Warzee N. Effects of drought and heat on forest insect populations in relation to the 2003 drought in Western Europe. Ann Forest Sci. 2006;63(6):613–24. https://doi.org/10.1051/forest:2006044.
Rousk J, Smith AR, Jones DL. Investigating the long-term legacy of drought and warming on the soil microbial community across five European shrubland ecosystems. Glob Chang Biol. 2013;19(12):3872–84. https://doi.org/10.1111/gcb.12338.
McDowell NG, Beerling DJ, Breshears DD, Fisher RA, Raffa KF, Stitt M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol Evol. 2011;26(10):523–32. https://doi.org/10.1016/j.tree.2011.06.003.
Hartmann H, Ziegler W, Kolle O, Trumbore S. Thirst beats hunger - declining hydration during drought prevents carbon starvation in Norway spruce saplings. New Phytol. 2013;200(2):340–9. https://doi.org/10.1111/nph.12331.
Rowland L, da Costa ACL, Galbraith DR, Oliveira RS, Binks OJ, Oliveira AAR, et al. Death from drought in tropical forests is triggered by hydraulics not carbon starvation. Nature. 2015;528(7580):119. https://doi.org/10.1038/nature15539.
Zeng N, Mariotti A, Wetzel P. Terrestrial mechanisms of interannual CO(2) variability. Global Biogeochem Cy. 2005;19(1):Gb1016. https://doi.org/10.1029/2004gb0022763.
Zscheischler J, Reichstein M, Harmeling S, Rammig A, Tomelleri E, Mahecha MD. Extreme events in gross primary production: a characterization across continents. Biogeosciences. 2014;11(11):2909–24. https://doi.org/10.5194/bg-11-2909-2014.
Budyko M. Climate and life. New York: Academic Press; 1974.
Jung M, Reichstein M, Margolis HA, Cescatti A, Richardson AD, Arain MA, et al. Global patterns of land-atmosphere fluxes of carbon dioxide, latent heat, and sensible heat derived from eddy covariance, satellite, and meteorological observations. J Geophys Res-Biogeo. 2011;116:G00j07. https://doi.org/10.1029/2010jg001566.
Zscheischler J, Mahecha MD, Harmeling S, Reichstein M. Detection and attribution of large spatiotemporal extreme events in Earth observation data. Ecol Inform. 2013;15:66–73. https://doi.org/10.1016/j.ecoinf.2013.03.004.
Dai A, Wigley TML. Global patterns of ENSO-induced precipitation. Geophys Res Lett. 2000;27(9):1283–6. https://doi.org/10.1029/1999gl011140.
Lyon B, Barnston AG. ENSO and the spatial extent of interannual precipitation extremes in tropical land areas. J Clim. 2005;18(23):5095–109. https://doi.org/10.1175/Jcli3598.1.
Marshall J, Kushner Y, Battisti D, Chang P, Czaja A, Dickson R, et al. North Atlantic climate variability: phenomena, impacts and mechanisms. Int J Climatol. 2001;21(15):1863–98. https://doi.org/10.1002/joc.693.
Trenberth KE. The definition of El Nino. Bull Am Meteorological Soc. 1997;78(12):2771–7. https://doi.org/10.1175/1520-0477(1997)078<2771:TDOENO>2.0.CO;2.
Collins M, An SI, Cai WJ, Ganachaud A, Guilyardi E, Jin FF, et al. The impact of global warming on the tropical Pacific ocean and El Nino. Nat Geosci. 2010;3(6):391–7. https://doi.org/10.1038/Ngeo868.
Gibson PB, Pitman AJ, Lorenz R, Perkins-Kirkpatrick SE. The role of circulation and land surface conditions in current and future Australian heat waves. J Clim. 2017;30:9933-47. https://doi.org/10.1175/JCLI-D-17-0265.1.
Ummenhofer CC, Sen Gupta A, Briggs PR, England MH, McIntosh PC, Meyers GA, et al. Indian and Pacific Ocean influences on southeast Australian drought and soil moisture (vol 24, pg 1313, 2011). J Clim. 2011;24(14):3796–6. https://doi.org/10.1175/Jcli-D-11-00229.1.
Richey JE, Nobre C, Deser C. Amazon River discharge and climate variability—1903 to 1985. Science. 1989;246(4926):101–3. https://doi.org/10.1126/science.246.4926.101.
Holmgren M, Scheffer M, Ezcurra E, Gutierrez JR, Mohren GMJ. El Nino effects on the dynamics of terrestrial ecosystems. Trends Ecol Evol. 2001;16(2):89–94. https://doi.org/10.1016/S0169-5347(00)02052-8.
Hallett TB, Coulson T, Pilkington JG, Clutton-Brock TH, Pemberton JM, Grenfell BT. Why large-scale climate indices seem to predict ecological processes better than local weather. Nature. 2004;430(6995):71–5. https://doi.org/10.1038/nature02708.
Bastos A, Running SW, Gouveia C, Trigo RM. The global NPP dependence on ENSO: La Nina and the extraordinary year of 2011. J Geophys Res-Biogeo. 2013;118(3):1247–55. https://doi.org/10.1002/jgrg.20100.
Kim JS, Kug JS, Yoon JH, Jeong SJ. Increased atmospheric CO2 growth rate during El Nino driven by reduced terrestrial productivity in the CMIP5 ESMs. J Clim. 2016;29(24):8783–805. https://doi.org/10.1175/Jcli-D-14-00672.1.
Cleverly J, Eamus D, Luo QY, Coupe NR, Kljun N, Ma XL, et al. The importance of interacting climate modes on Australia’s contribution to global carbon cycle extremes. Sci Rep-Uk. 2016;6:23113. https://doi.org/10.1038/srep23113.
Cai W, Cowan T, Raupach M. Positive Indian Ocean Dipole events precondition southeast Australia bushfires. Geophys Res Lett. 2009;36:L19710. https://doi.org/10.1029/2009gl039902.
Chen Y, Morton DC, Andela N, Giglio L, Randerson JT. How much global burned area can be forecast on seasonal time scales using sea surface temperatures? Environ Res Lett. 2016;11(4):045001. https://doi.org/10.1088/1748-9326/11/4/045001.
Liu JJ, Bowman KW, Schimel DS, Parazoo NC, Jiang Z, Lee M, et al. Contrasting carbon cycle responses of the tropical continents to the 2015-2016 El Nino. Science. 2017;358(6360):191. https://doi.org/10.1126/science.aam5690.
Belmecheri S, Babst F, Hudson AR, Betancourt J, Trouet V. Northern Hemisphere jet stream position indices as diagnostic tools for climate and ecosystem dynamics. Earth Interact. 2017;21:1–23. https://doi.org/10.1175/Ei-D-16-0023.1.
Bastos A, Janssens IA, Gouveia CM, Trigo RM, Ciais P, Chevallier F, et al. European land CO2 sink influenced by NAO and East-Atlantic Pattern coupling. Nat Commun. 2016;7:10315. https://doi.org/10.1038/ncomms10315.
Parazoo NC, Barnes E, Worden J, Harper AB, Bowman KB, Frankenberg C, et al. Influence of ENSO and the NAO on terrestrial carbon uptake in the Texas-northern Mexico region. Global Biogeochem Cy. 2015;29(8):1247–65. https://doi.org/10.1002/2015gb005125.
Desai AR, Wohlfahrt G, Zeeman MJ, Katata G, Eugster W, Montagnani L, et al. Montane ecosystem productivity responds more to global circulation patterns than climatic trends. Environ Res Lett. 2016;11(2):024013. https://doi.org/10.1088/1748-9326/11/2/024013.
Bastos A, Ciais P, Park T, Zscheischler J, Yue C, Barichivich J, et al. Was the extreme Northern Hemisphere greening in 2015 predictable? Environ Res Lett. 2017;12(4):044016. https://doi.org/10.1088/1748-9326/aa67b5.
Shepherd TG. Atmospheric circulation as a source of uncertainty in climate change projections. Nat Geosci. 2014;7(10):703–8. https://doi.org/10.1038/Ngeo2253.
Cai WJ, Borlace S, Lengaigne M, van Rensch P, Collins M, Vecchi G, et al. Increasing frequency of extreme El Nino events due to greenhouse warming. Nat Clim Chang. 2014;4(2):111–6. https://doi.org/10.1038/Nclimate2100.
Coumou D, Lehmann J, Beckmann J. The weakening summer circulation in the Northern Hemisphere mid-latitudes. Science. 2015;348(6232):324–7. https://doi.org/10.1126/science.1261768.
Trouet V, Babst F, Meko M. Recent enhanced high-summer North Atlantic Jet variability emerges from three-century context. Nat Commun. 2018;9:180. https://doi.org/10.1038/s41467-017-02699-3.
Zemp DC, Schleussner CF, Barbosa HMJ, Hirota M, Montade V, Sampaio G, et al. Self-amplified Amazon forest loss due to vegetation-atmosphere feedbacks. Nat Commun. 2017;8:14681. https://doi.org/10.1038/ncomms14681.
Keys PW, van der Ent RJ, Gordon LJ, Hoff H, Nikoli R, Savenije HHG. Analyzing precipitationsheds to understand the vulnerability of rainfall dependent regions. Biogeosciences. 2012;9(2):733–46. https://doi.org/10.5194/bg-9-733-2012.
Garcia ES, Swann ALS, Villegas JC, Breshears DD, Law DJ, Saleska SR, et al. Synergistic ecoclimate teleconnections from forest loss in different regions structure global ecological responses. Plos One. 2016;11(11):e0165042. https://doi.org/10.1371/journal.pone.0165042.
Shepherd TG. A common framework for approaches to extreme event attribution. Curr Clim Change Rep. 2016;2(1):28–38. https://doi.org/10.1007/s40641-016-0033-y.
Vautard R, Yiou P, Otto F, Stott P, Christidis N, Van Oldenborgh GJ, et al. Attribution of human-induced dynamical and thermodynamical contributions in extreme weather events. Environ Res Lett. 2016;11(11):114009. https://doi.org/10.1088/1748-9326/11/11/114009.
Hoover DL, Rogers BM. Not all droughts are created equal: the impacts of interannual drought pattern and magnitude on grassland carbon cycling. Glob Chang Biol. 2016;22(5):1809–20. https://doi.org/10.1111/gcb.13161.
Miralles DG, Teuling AJ, van Heerwaarden CC, de Arellano JVG. Mega-heatwave temperatures due to combined soil desiccation and atmospheric heat accumulation. Nat Geosci. 2014;7(5):345–9. https://doi.org/10.1038/NGEO2141.
Knapp AK, Avolio ML, Beier C, Carroll CJW, Collins SL, Dukes JS, et al. Pushing precipitation to the extremes in distributed experiments: recommendations for simulating wet and dry years. Glob Chang Biol. 2017;23(5):1774–82. https://doi.org/10.1111/gcb.13504.
Jentsch A, Kreyling J, Beierkuhnlein C. A new generation of climate-change experiments: events, not trends. Front Ecol Environ. 2007;5(7):365–74. https://doi.org/10.1890/1540-9295(2007)5%5B365:Angoce%5D2.0.Co;2.
Wilcox KR, Shi Z, Gherardi LA, Lemoine NP, Koerner SE, Hoover DL, et al. Asymmetric responses of primary productivity to precipitation extremes: a synthesis of grassland precipitation manipulation experiments. Glob Chang Biol. 2017;23(10):4376–85. https://doi.org/10.1111/gcb.13706.
Shi Z, Thomey ML, Mowll W, Litvak M, Brunsell NA, Collins SL, et al. Differential effects of extreme drought on production and respiration: synthesis and modeling analysis. Biogeosciences. 2014;11(3):621–33. https://doi.org/10.5194/bg-11-621-2014.
Ruehr NK, Gast A, Weber C, Daub B, Arneth A. Water availability as dominant control of heat stress responses in two contrasting tree species. Tree Physiol. 2016;36(2):164–78. https://doi.org/10.1093/treephys/tpv102.
Fraser LH, Henry HAL, Carlyle CN, White SR, Beierkuhnlein C, Cahill JF, et al. Coordinated distributed experiments: an emerging tool for testing global hypotheses in ecology and environmental science. Front Ecol Environ. 2013;11(3):147–55. https://doi.org/10.1890/110279.
Novick KA, Ficklin DL, Stoy PC, Williams CA, Bohrer G, Oishi AC, et al. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat Clim Chang. 2016;6(11):1023–7. https://doi.org/10.1038/Nclimate3114.
Beier C, Beierkuhnlein C, Wohlgemuth T, Penuelas J, Emmett B, Korner C, et al. Precipitation manipulation experiments - challenges and recommendations for the future. Ecol Lett. 2012;15(8):899–911. https://doi.org/10.1111/j.1461-0248.2012.01793.x.
Wu D, Ciais P, Viovy N, Knapp AK, Wilcox K, Bahn M, et al. Asymmetric responses of primary productivity to altered precipitation simulated by ecosystem models across three longterm grassland sites. Biogeosci Discuss. 2018;2018:1–27. https://doi.org/10.5194/bg-2018-53.
Mahecha MD, Gans F, Sippel S, Donges JF, Kaminski T, Metzger S, et al. Detecting impacts of extreme events with ecological in situ monitoring networks. Biogeosciences. 2017;14(18):4255–77. https://doi.org/10.5194/bg-14-4255-2017.
Schwalm CR, Williams CA, Schaefer K, Baldocchi D, Black TA, Goldstein AH, et al. Reduction in carbon uptake during turn of the century drought in western North America. Nat Geosci. 2012;5(8):551–6. https://doi.org/10.1038/Ngeo1529.
Biederman JA, Scott RL, Goulden ML, Vargas R, Litvak ME, Kolb TE, et al. Terrestrial carbon balance in a drier world: the effects of water availability in southwestern North America. Glob Chang Biol. 2016;22(5):1867–79. https://doi.org/10.1111/gcb.13222.
Yu GR, Chen Z, Piao SL, Peng CH, Ciais P, Wang QF, et al. High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region. Proc Natl Acad Sci U S A. 2014;111(13):4910–5. https://doi.org/10.1073/pnas.1317065111.
Foken T. The energy balance closure problem: an overview. Ecol Appl. 2008;18(6):1351–67. https://doi.org/10.1890/06-0922.1.
Korner C. Slow in, rapid out—carbon flux studies and Kyoto targets. Science. 2003;300(5623):1242–3. https://doi.org/10.1126/science.1084460.
Asner GP, Alencar A. Drought impacts on the Amazon forest: the remote sensing perspective. New Phytol. 2010;187(3):569–78. https://doi.org/10.1111/j.1469-8137.2010.03310.x.
Sims DA, Brzostek ER, Rahman AF, Dragoni D, Phillips RP. An improved approach for remotely sensing water stress impacts on forest C uptake. Glob Chang Biol. 2014;20(9):2856–66. https://doi.org/10.1111/gcb.12537.
Vicca S, Balzarolo M, Filella I, Granier A, Herbst M, Knohl A, et al. Remotely-sensed detection of effects of extreme droughts on gross primary production. Sci Rep-Uk. 2016;6:28269. https://doi.org/10.1038/srep28269.
Leuning R, Cleugh HA, Zegelin SJ, Hughes D. Carbon and water fluxes over a temperate Eucalyptus forest and a tropical wet/dry savanna in Australia: measurements and comparison with MODIS remote sensing estimates. Agric For Meteorol. 2005;129(3–4):151–73. https://doi.org/10.1016/j.agrformet.2004.12.004.
Hwang T, Gholizadeh H, Sims DA, Novick KA, Brzostek ER, Phillips RP, et al. Capturing species-level drought responses in a temperate deciduous forest using ratios of photochemical reflectance indices between sunlit and shaded canopies. Remote Sens Environ. 2017;199:350–9. https://doi.org/10.1016/j.rse.2017.07.033.
Rascher U, Pieruschka R. Spatio-temporal variations of photosynthesis: the potential of optical remote sensing to better understand and scale light use efficiency and stresses of plant ecosystems. Precis Agric. 2008;9(6):355–66. https://doi.org/10.1007/s11119-008-9074-0.
Guanter L, Zhang YG, Jung M, Joiner J, Voigt M, Berry JA, et al. Global and time-resolved monitoring of crop photosynthesis with chlorophyll fluorescence. Proc Natl Acad Sci U S A. 2014;111(14):E1327–33. https://doi.org/10.1073/pnas.1320008111.
Sun Y, Frankenberg C, Wood JD, Schimel DS, Jung M, Guanter L, et al. OCO-2 advances photosynthesis observation from space via solar-induced chlorophyll fluorescence. Science. 2017;358(6360):189. https://doi.org/10.1126/science.aam5747.
Lee JE, Frankenberg C, van der Tol C, Berry JA, Guanter L, Boyce CK, et al. Forest productivity and water stress in Amazonia: observations from GOSAT chlorophyll fluorescence. P Roy Soc B-Biol Sci. 2013;280(1761):20130171. https://doi.org/10.1098/rspb.2013.0171.
Ma XL, Huete A, Cleverly J, Eamus D, Chevallier F, Joiner J, et al. Drought rapidly diminishes the large net CO2 uptake in 2011 over semi-arid Australia. Sci Rep-Uk. 2016;6:37747. https://doi.org/10.1038/srep37747.
Yoshida Y, Joiner J, Tucker C, Berry J, Lee JE, Walker G, et al. The 2010 Russian drought impact on satellite measurements of solar-induced chlorophyll fluorescence: insights from modeling and comparisons with parameters derived from satellite reflectances. Remote Sens Environ. 2015;166:163–77. https://doi.org/10.1016/j.rse.2015.06.008.
Chevallier F, Maksyutov S, Bousquet P, Breon FM, Saito R, Yoshida Y, et al. On the accuracy of the CO2 surface fluxes to be estimated from the GOSAT observations. Geophys Res Lett. 2009;36:L19807. https://doi.org/10.1029/2009gl040108.
Detmers RG, Hasekamp O, Aben I, Houweling S, van Leeuwen TT, Butz A, et al. Anomalous carbon uptake in Australia as seen by GOSAT. Geophys Res Lett. 2015;42(19):8177–84. https://doi.org/10.1002/2015gl065161.
Sulman BN, Roman DT, Yi K, Wang LX, Phillips RP, Novick KA. High atmospheric demand for water can limit forest carbon uptake and transpiration as severely as dry soil. Geophys Res Lett. 2016;43(18):9686–95. https://doi.org/10.1002/2016gl069416.
Stoy PC, Richardson AD, Baldocchi DD, Katul GG, Stanovick J, Mahecha MD, et al. Biosphere-atmosphere exchange of CO2 in relation to climate: a cross-biome analysis across multiple time scales. Biogeosciences. 2009;6(10):2297–312. https://doi.org/10.5194/bg-6-2297-2009.
De Kauwe MG, Medlyn BE, Walker AP, Zaehle S, Asao S, Guenet B, et al. Challenging terrestrial biosphere models with data from the long-term multifactor prairie heating and CO2 enrichment experiment. Glob Chang Biol. 2017;23(9):3623–45. https://doi.org/10.1111/gcb.13643.
Rogers A, Medlyn BE, Dukes JS, Bonan G, von Caemmerer S, Dietze MC, et al. A roadmap for improving the representation of photosynthesis in Earth system models. New Phytol. 2017;213(1):22–42. https://doi.org/10.1111/nph.14283.
Xu XT, Medvigy D, Powers JS, Becknell JM, Guan KY. Diversity in plant hydraulic traits explains seasonal and inter-annual variations of vegetation dynamics in seasonally dry tropical forests. New Phytol. 2016;212(1):80–95. https://doi.org/10.1111/nph.14009.
Sakschewski B, von Bloh W, Boit A, Poorter L, Pena-Claros M, Heinke J, et al. Resilience of Amazon forests emerges from plant trait diversity. Nat Clim Change. 2016;6(11):1032. https://doi.org/10.1038/Nclimate3109.
Reichstein M, Mahecha MD, Ciais P, Seneviratne SI, Blyth EM, Carvalhais N, et al. Elk–testing climate–carbon cycle models: a case for pattern–oriented system analysis. iLEAPS Newsletter. 2011;11:14–21.
Rammig A, Wiedermann M, Donges JF, Babst F, von Bloh W, Frank D, et al. Coincidences of climate extremes and anomalous vegetation responses: comparing tree ring patterns to simulated productivity. Biogeosciences. 2015;12(2):373–85. https://doi.org/10.5194/bg-12-373-2015.
Luo YQ, Randerson JT, Abramowitz G, Bacour C, Blyth E, Carvalhais N, et al. A framework for benchmarking land models. Biogeosciences. 2012;9(10):3857–74. https://doi.org/10.5194/bg-9-3857-2012.
Sippel S, Otto FEL, Forkel M, Allen MR, Guillod BP, Heimann M, et al. A novel bias correction methodology for climate impact simulations. Earth Syst Dynam. 2016;7(1):71–88. https://doi.org/10.5194/esd-7-71-2016.
Ahlstrom A, Canadell JG, Schurgers G, Wu MC, Berry JA, Guan KY, et al. Hydrologic resilience and Amazon productivity. Nat Commun. 2017;8:387. https://doi.org/10.1038/s41467-017-00306-z.
Powell TL, Galbraith DR, Christoffersen BO, Harper A, Imbuzeiro HMA, Rowland L, et al. Confronting model predictions of carbon fluxes with measurements of Amazon forests subjected to experimental drought. New Phytol. 2013;200(2):350–64. https://doi.org/10.1111/nph.12390.
Huang YY, Gerber S, Huang TY, Lichstein JW. Evaluating the drought response of CMIP5 models using global gross primary productivity, leaf area, precipitation, and soil moisture data. Global Biogeochem Cy. 2016;30(12):1827–46. https://doi.org/10.1002/2016gb005480.
Restrepo-Coupe N, Levine NM, Christoffersen BO, Albert LP, Wu J, Costa MH, et al. Do dynamic global vegetation models capture the seasonality of carbon fluxes in the Amazon basin? A data-model intercomparison. Glob Chang Biol. 2017;23(1):191–208. https://doi.org/10.1111/gcb.13442.
Hegerl GC, Hoegh-Guldberg O, Casassa G, Hoerling MP, Kovats RS, Parmesan C, et al. Good practice guidance paper on detection and attribution related to anthropogenic climate change. In Meeting Report of the Intergovernmental Panel on Climate Change Expert Meeting on Detection and Attribution of Anthropogenic Climate Change. IPCC Working Group I Technical Support Unit, University of Bern, Bern, Switzerland; 2010.
Stott PA, Christidis N, Otto FEL, Sun Y, Vanderlinden JP, van Oldenborgh GJ, et al. Attribution of extreme weather and climate-related events. Wires Clim Change. 2016;7(1):23–41. https://doi.org/10.1002/wcc.380.
Acknowledgements
We thank Prof. Kimberly A. Novick, Prof. Richard P. Phillips, and the Morgan Monroe State Forest (US-MMS) flux tower team for openly sharing their data. The authors are grateful to the FLUXCOM initiative (http://www.fluxcom.org) for providing the data used in Fig. 4. We also acknowledge valuable and insightful comments on a previous version of the manuscript provided by two anonymous reviewers. Authors affiliated with the MPI for Biogeochemistry acknowledge the European Union for funding via the H2020 project BACI (grant agreement no.: 640176).
Funding
Open access funding provided by Max Planck Society.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
This article is part of the Topical Collection on Carbon Cycle and Climate
Electronic Supplementary Material
ESM 1
(DOCX 112 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Sippel, S., Reichstein, M., Ma, X. et al. Drought, Heat, and the Carbon Cycle: a Review. Curr Clim Change Rep 4, 266–286 (2018). https://doi.org/10.1007/s40641-018-0103-4
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40641-018-0103-4