
Abstract
The internal, multi-element skeleton of echinoderms can differentiate almost as much as that of the vertebrates. The implied probability of post-mortem disarticulation was bypassed in pelagic species, whose carcasses could sink directly onto toxic sea bottoms. Among them were isocrinids attached to driftwood. In the Toarcian Posidonia Shales they are represented by two easily distinguishable forms: (1) the giant and long-stemmed Seirocrinus was probably a passive tow-net filtrator; (2) the smaller and heavily cirrated Pentacrinites may have produced its own filter current, even though this assumption is in conflict with the distribution of muscles in present-day benthic Isocrinida. In a newly discovered colony of the smaller Early Liassic Pentacrinites fossilis, both strategies are found together. This leads to the hypothesis that the two Toarcian forms may also represent developmental stages of the same species. After the Toarcian, Seirocrinus (as either a taxon or a developmental stage) became extinct, probably due to shipworms reducing the drifting time of logs. In contrast, the proposed feeding style of Pentacrinites allowed it to return to the benthic realm, but in an inverted attitude. During this complex history, most adaptational changes required only heterochronic shifts in the development of tegmen, arms, pinnules, stem, and cirri. Unrelated pseudoplanktonic crinoids of earlier times show convergent adaptations with different pathways.
Similar content being viewed by others



Introduction
Two kinds of Isocrinida found on driftwood in the Toarcian (L. Jurassic) Konservat-Lagerstaette of Holzmaden (Hess 1999b) can be easily distinguished, because they represent radically different feeding strategies: (1) The larger Seirocrinus subangularis has an enlarged crown and an extremely long stem with widely spaced and partly dysfunctional cirri. It was probably a townet filtrator. (2) Pentacrinites dichotomus has short, densely cirrated stems and moderately sized crowns (Fig. 1). It possibly used its compressed (“winged”) cirri to produce an own filter current. In a colony of the Sinemurian Pentacrinites fossilis (Seilacher et al. 2007), both strategies are represented. This raises the question: do the two Toarcian forms really belong to different taxa or rather represent stages in the life history of one and the same animal?

In these two Toarcian driftwood dwellers, peramorphic development of the arms was combined with a switch to heterotomous, but unlimited, branching at the 20-arm stage. Branches on the inner side (ramuli) remain thinner, run parallel, and do not bifurcate any more, while those on the outer side keep branching at regular intervals. This allows neighboring arms to be coupled by pinnular adhesion and to act like the radial spokes of an umbrella. Differences between the two forms regard overall size and the distancing of nodal cirri, which became “winged” in Pentacrinites. If one deals with developmental stages rather than different taxa, the distal root cirri of Seirocrinus should also be winged (see Fig. 6), while its proximal cirri became reduced to a “mini-skirt” protecting only the non-filtrating base of the crown. (modified from Seilacher and Hauff (2004)
Answers to this question come from the morphogenetic outfit, from taphonomic behavior, and from earlier lineages with similar life styles.
Morphogenetic outfit
The basic construction of Isocrinida (including species surviving on present deep-sea bottoms) is a rigid body capsule (calyx) with a fixed number of sutured plates and a flexible tegmen, plus three kinds of flexible appendages:
-
(1)
The arms support rows of pinnules and tube feet for passive filter feeding in a current coming from the aboral side. In order to increase the filter area, the original five arms can multiply by subsequent bifurcations.
-
(2)
On the aboral side, an articulated stem elevates the crown from the substrate into zones of higher current velocity. It grew longer by generations of nodals formed at the base of the calyx and by intercalation of internodals between them.
-
(3)
The radiating cirri articulating with the nodals can be used for various purposes. They become longer by addition of new cirrals in the root tuft. These could alter shapes during subsequent accretionary growth.
Originally, all three kinds of appendages may have grown by introducing new ossicles at their tips; but with the specialization of the distal ossicles for particular functions (holdfasts in the stem; hooks in the cirri), growth zones had to be displaced. This basic outfit was modified in adaptation to a pseudoplanktonic life style.
In the following chapters, observed changes are interpreted as heterochronic shifts in a basic morphogenetic program.
Arms and ramuli
On driftwood, crinoids faced two problems: (1) the crowns had to hang down rather than standing up and the stem was exposed to tensional rather than compressional stress; (2) as a log moves with the water, there is no velocity gradient near the substrate other than by wave action. Therefore, all modern, but smaller, driftwood-dwellers (bryozoans, bivalves, barnacles, tunicates) create their own filter current.
Another tendency of pseudoplanktonic crinoids is to enlarge the filter fan relative to the calyx. This is facilitated by the absence of coarse sediment suspended during storms. So the arms could remain splayed all the time, held together by ciliary (?) adhesion between opposed pinnules. To this end, driftwood dwellers increased the number of arm branchings beyond the two or three usual in benthic forms—only that the following branchings are heterotomous. This means that in each axillary one branch continues to bifurcate at regular distances. The other branch (ramulus) is not only thinner; in pentacrinids it also grows-on without further branchings.
The determination of axillaries appears to differ as well. Isotomous branching probably started at the growing tip of the initial arms. As there was, at this stage, no left/right difference between the resulting branches, the number of interaxillaries could be even or uneven. In heterotomous arms, however, the tip sections are in all stages longer than the interaxillar ones. So these axillaries were probably established at some distance behind the tips. As the numbers of interaxillaries are always even (Simms 1989), new heterotomous axillaries were determined once the pinnules of the previous ramulus had grown sufficently. This is probably why axillaries bear no pinnules (Simms 1989, p. 18). After this, interaxillar sections could grow only by accretionary enlargement of ossicles, while the number of ossicles remained unchanged.
In this peramorphic process, some pecularities require special explanations: (1). The regular distancing of branching points. After the first two branchings (Fig. 1), one always counts 8, 10, 12, 14, or 16 brachial ossicles between branching points. The distance they cover corresponds to the length of the pinnules along the adjacent ramuli. That the intervening brachials always come in even numbers has its reason in an asymmetry that in Isocrinida cannot be recognized in aboral view: adorally, each brachial bears a single pinnule that is alternatingly positioned on the right or left side of the arm. In a one-sided system based on heterotomous bifurcations, a new ramulus can form only in phase with the proper step. (2) The switch from equal to endotomous dichotomy happening after the first two isotomous branchings (i.e. at the 20-arm-stage) means that the branch on the one side continued to grow as an arm with the potential for further branching, whereas the other was downgraded and attenuated into a long ramulus that in pentacrinids could not branch any further.
One effect of this transformation was that the ramuli run parallel to each other, rather than radiating, i.e., a banana-road system is established in a radially symmetric regime. As a second effect, the branch that grew-on as an arm became straight and thereby fit for a coupling function, which can be inferred from a strange taphonomic behavior. During landing, dragging, and compaction, the arms became passively splashed. Nevertheless, adjacent arms tend to keep in touch. This suggests that the tips of the pinnules adhered to each other.
Pinnules
These simple rows of minute ossicles serve various purposes:
-
(1)
They equally spread the ultimate food catchers (tube “feet”). For this purpose, they line the arms and ramuli on either side. But as in all echinoderms (except in the compound plates of cidaroid echinids) one ossicle can bear only a single tube foot or triplet, pinnules alternate. This is why in side view of uniserial arms and ramuli, pinnules appear only in half the number of brachials and ramulars.
-
(2)
They border the sides of the ambulacral grooves, in which mucus-bound food particles are transported to the mouth. Therefore, basal pinnulars tend to expand into polygonal plates suturing with their neighbors. That the corresponding tube feet helped in the food transport can be inferred.
-
(3)
In addition, pinnules along the basal parts of the arms may protect the vulnerable tegmen of the body capsule. For this purpose, almost all pinnulars may give up their flexible connections and become sutured. Still this armor did probably not fuse with the tegmen (there were still tube feet on the oral side!). Such additional protection became important when the tegmen disproportionally expanded, as happened in adult Seirocrinus. Because the transformation relates to scaling and could be easily accomplished (but only in this direction!), “incorporation” of pinnules (Simms 1989, p. 23) is unsuited as a feature separating the two genera.
-
(4)
Although it made pseudoplanktonic crinoid fossils so visually attractive, the adhesive function of pinnules received less attention. It is expressed by the fact that overall deformation during landing did never lead to entanglement of the pinnules and that torn-apart arms rarely lost the connection between their pinnules. Yet, this function (probably working like velcro strips in modern textiles, shoes, and camera boxes) was essential for pelagic crinoids, because it held the filter fan open without energy expenditure.
-
(5)
The alternative view (proposed by reviewers) that this effect is due to mutable collagen in the arms is unlikely, because it does not explain the stronger adherence towards the arm tips.
Constructionally, the adhesive function explains why pinnules are longest along the margins of endotomous arms facing the opposite ones—even though they probably formed aboral folds in the splayed filter fan.
This leads to the question about the twin rows of pinnules on arms and ramules. In aboral views of fossil crowns they cannot be seen. Were they reduced to adhesive tubercles?
As some degree of adhesiveness may also exist in modern benthic crinoids, the mechanism involved will hopefully be elucidated in the future.
At the same time, adhesive connections must have been spotty enough to let water flow through. In this way the velcro couplings may have served not only for stabilizing the splayed crown, but they also formed a sieve keeping oversized objects away from the food-catching tube feet on the downcurrent side.
As there was no limit to arm growth and the introduction of new ramuli, the food-catching apparatus became enormously enlarged relative to the size of the body capsule. This is particularly true for Seirocrinus, where the crown may reach 80 cm in diameter, but with a calyx of only about 3 cm. This transformation, however, was possible only in the pelagic realm, where crowns had no more to be closed against sediment stirred up during a storm or against predators (regenerated arms are rare in pelagic crinoids).
Stem
As mentioned above, the introduction of primary columnals (nodals) had switched early on from the tip of the stem to its connection with the body capsule. Because this connection grew larger with age, an unmodified stem would have tapered away from the crown. Benthic crinoids overcome distal tapering by accretionary growth of the columnals, in diameter as well as length (Fig. 2). In order to maintain the necessary flexibility of the stem, the columnals of townet filtrators remained very thin and enlarged mainly in diameter. At the same time, their outlines became rounded (Fig. 1). Nevertheless, radial growth of the columnals was insufficient to completely offset distal tapering of the stem in pseudoplanktonic species.

In all Articulata, stem growth proceeds in a double mode: Nodals (black) form serially at the base of the calyx, followed by the intercalation of internodals in hierarchical order. In the benthic Encrinus, the latter process stoped after three generations of internodals and accretionary growth led to drum-shaped columnals and a solid stem that thickens distally. In the pseudoplanktonic Pentacrinites, no more internodals were added after two generations, with little (mainly transversal) secondary accretion. Seirocrinus had no limit to either mode of stem-lengthening. It also shows how driftwood dwellers accelerated growth by “brooding” several generations of columnals simultaneously. The “factory” for nodals is protected by spines extending from the radial plates of the calyx (black), while the ones for internodals are protected by the thickened rims of already mineralized neighbors. (Modified from Seilacher and Hauff (2004)
The optimal flexibility distribution along the stem depends on life styles. The stems of benthic crinoids resemble the stems of trees: they should be most flexible near the top and become stiffer towards the base. Accordingly, nodals as well as internodals keep growing thicker and wider until an almost square axial section is reached. Proximal flexibility is maintained by the continuous addition of relatively thin nodals and a limited number of internodals. As observed in living benthic forms, crowns are passively bent downcurrent, so that they become flushed from the aboral side.
In claiming a pseudoplanktonic life style for Seirocrinus (Seilacher et al. 1968), stem flexibility was a major argument. It differs from benthic crinoids in a fabricational sense by the unlimited intercalation of internodals (more than 1,000 between two nodals). Only in the most distal part of the stem is this intercalation halted in order to develop a cirral root tuft. However, as Baumiller and Ausich (1996) found out, the aspect ratio of columnals is no reliable predictor of stem flexibility. Therefore, it is important to judge it by also measuring the minimal turning radii at accidental bends in many specimens relative to their distance from the crown. In Seirocrinus this method shows that stem flexibility increased away from the crown, opposite to what one observes in most benthic crinoids. One reason for this is the remnant tapering of the Seirocrinus stem away from the crown.
Crinoids also evolved the intercalation of internodals between preexisting columnals as an additional method of secondarily lengthening the stem. In proximal stem sections this process is expressed by a nested hierarchy of columnals in numbers of 1, 3, 7, 15,…..between the primary nodals. In Isocrinida, the difference is accentuated by the fact that only the nodals bear cirri and their corresponding sockets (Fig. 2).
By heterochronic shifts in the three growth modes (nodal, internodal, and accretional), the stem could be locally modified for specific functional requirements, such as a higher flexibility in the root section to prevent breakage during storms, as well as additional (but flexible) anchorage by crowded cirri (root tuft). This is in contrast to mud-dwelling crinoids, where rootlets are rigid (Seilacher and MacClintock 2005).
Cirri
The nodal cirri of Isocrinida are a derived feature. The transformation of their distalmost ossicle into a hook (which required new cirrals to be introduced at the base) indicates an original holdfast function of these organs. It is preserved in modern representatives, where a particular kind of connection (cryptosymplexy) on the distal side of each nodal allows older parts of the stem to be shed off. Thereby cirri at the broken end can readily switch into function by grasping onto firm substrates or by actively sinking the stem into soft mud (Isselicrinus Fujiwara et al. 2005; Seilacher and MacClintock 2005).
Secondary transformation for additional functions is possible as the cirrals grow larger farther away from their point of origin. It can result in non-cylindrical cross sections, as shown by the laterally compressed cirri that free-living comatulids use for grasping to the substrate (Donovan 1993) or by the alate cirrals in Pentacrinites (Fig. 1). Intercalation, analogous to the internodals of the stem, is unknown in cirri.
Obviously, the most critical question in the present case is the function of the cirri in Pentacrinites, because
-
(1)
By their distal hook they provide flexible attachment to a substrate bouncing in the waves (anchoring function).
-
(2)
Nevertheless, cirral development is not retarded. Rather it is completed next to the crown, thereby obstructing the passive aboral filter current of ordinary crinoids.
-
(3)
Rhombic (“alate”) cross sections do not improve the anchoring function; but theoretically they would allow undulation in order to swim Hauff (1984 ) or, more likely, to replace the ordinary passive filter current by an active one in the same (aboral to oral) direction. This would make sense for early stages attached to a substrate that drifts with the ambiental current.
-
(4)
Densing of cirri by reducing the intercalation of none-cirrate internodals added to the blocking effect, but enhanced the power of the hypothesized active ventilation.
-
(5)
This model, however, is in conflict with the fact that the cirri of modern isocrinoids are not equipped with muscles, which would be necessary to make such a machinery work.
Functional divergence in Pentacrinites versus Seirocrinus
As mentioned above, the two Toarcian forms appear to have evolved by approaching different adaptive peaks, or functional paradigms. In the absence of modern analogs for either strategy, evolutionary pathways must be reconstructed from fossil evidence. This is facilitated by the fact that heterochronic shifts in the organs concerned suffice to explain differences between the two morphologies. Our task, then, is to follow evolutionary trends in the two lineages through geologic time.
Crucial in this investigation has been a large slab from lowermost Jurassic (Sinemurian) bituminous shales on the Sumerset coast of England (Seilacher et al. 2007). After having been found and meticulously prepared by M. Wolf, it now shows about 20 individuals still attached to remains of a jettized driftwood (Figs. 3, 4). Surprisingly, morphologies are not intermediate to benthic ancestors, but resemble the Toarcian Pentacrinites to such a degree that separation at the species level (P. fossilis vs. P. dichotomus) has been questioned. Nevertheless, larger specimens from the same colony and from other localities approach Seirocrinus: they introduce more than the two generations of internodals required for proper distancing of subsequent cirri in the stem and enlarge the filter area by developing additional ramuli in the crown (Fig. 5).

Slab collected and prepared by Jettized remains of the driftwood are preserved along lower edge. Two individuals in the upper half broke off as the crinoids touched bottom. Because their crowns got entangled during final transport they ended up in opposite current orientation

Some crinoids drawn in life position from the colony on the Wolf slab. They show that the whole stem is densely cirrated in the early stages (a, b). Larger individuals, however (c, d), stretch the stem by intercalating more than two generations of internodals between the cirrated nodals. In d, one of the arms stopped growing at a juvenile stage; but neighboring arms closed the gap in the filter fan by clinging together with the tips of their pinnnules

Crowns grew by accretionary enlargement of ossicles and by the introduction of new ones at the arm tips (axillaria drawn in black). The switch from iso-to-heterotomous branching was not sudden: Arm sections following the last isotomies are longer in the outer versus the inner branches. The finer pinnulae (not drawn) filled spaces between the parallel ramuli. As indicated by numbers, new ramuli were constantly introduced in proportion to the length of the stem
On the other hand, the Sinemurian form (Pentacrinites fossilis; see Simms 1989, 1999) is clearly offset from benthic Isocrinida by endotomously branching arms and ramuli that end almost flush with the arm tips, as well as by its long, alate, and closely distanced cirri. Their successive usage for active filtration, as well as for strengthening anchorage, is suggested by an individual in the Wolf colony, in which the distal cirri do not turn towards the crown, but towards the root. Even the aboral spines of the calyx (which in Fig. 2 have been interpreted as a strengthening device allowing to accelerate nodalia production) were already present.
It is probably no coincidence that individuals with the characteristics of Pentacrinites (dense, alate cirri) and of Seirocrinus (widely distanced cirri) also co-occur in the Norian of Northern Siberia (Hagdorn, personal communication 2010). As available specimens are relatively large, both forms are united under the name Seirocrinus klikushini (see Hagdorn 1996). In any case, the essential steps from benthic to pseudoplanktonic Isocrinida had already been made by Late Triassic times.
Are Toarcian Pentacrinites and Seirocrinus conspecific growth stages?
As selection pressure in pseudoplanktonic crinoids shifts with body size, it is tempting to think of a kind of metamorphosis, in addition to the one happening after settlement of the larva. It would be comparable to that in advanced insects, but probably not connected with sexual maturation.
If Seirocrinus passed through an actively filtrating juvenile stage, one should expect cirrals with a rhombic cross section in the oldest part of its stem—more exactly in tufts attached to the wood, because piggybackers on older stems could bypass the Pentacrinites stage and start with the townet strategy right away. Unfortunately, to prove this is a difficult task. In colonies, root regions are so entangled that details are difficult to prepare and to analyse. Eventually, an individual Seirocrinus with a stem of 120 cm was found that had broken off the driftwood together with its root (Fig. 6a). As predicted by the two-stage model, root cirri are compressed and branch-off towards the crown before they bend back to act as roots (Fig. 6b).

a This medium-sized Seirocrinus from the Posidonia Shale is the hoped-for Rosetta Stone, because it broke off a driftwood and sank to the bottom with the heavy stem ahead. It shows the “miniskirt” of small cylindrical cirri protecting the nodal factory. Further away from the crown, cirri (arrows) became atrophied and distanced by up to 31 internodals. Near the distal end they are again larger and crowded into a root tuft. Note that breaks happened commonly on the proximal side of a nodal. b Close-up showing that root cirri are compressed and branch-off towards the crown, as in Pentacrinites, before turning towards the substrate for a subsequent root function. The three distancing internodals, however, grew to equal thickness in the more proximal parts of the stem. (GPIT 1918/2; phot. W. Gerber)
Another indication comes from Pentacrinites dichotomus itself. Its winged cirri are usually so long that they cover most of the crown (see fig. 202 in Hess 1999a). The specimen in which this is not the case (his fig. 201) is also larger. Was it just switching to the Seirocrinus strategy?
Earlier independent evolutionary experiments
M.Triassic Traumatocrinus (Fig. 7a)
Serendipitously, the fossil record provides us with parallel case histories for comparison. As shown by Hagdorn et al. (2007), the Middle Triassic (Ladinian/Carnian) encrinid crinoid Traumatocrinus shows an intriguingly parallel adaptation to a pseudoplanktonic life style on driftwood. Even though this genus does not quite reach the size of Seirocrinus (stems up to 11 m, crown up to 40 cm, it also shows an endotomous increase of the filter area and pinnular couplings including the arms. However, being a member of the Encrinida, it had no cirri except in the root. How did it filterfeed as a juvenile?

Convergent lineages reflect similar functional adaptations (e.g., peramorphic enlargement and adhesive integration of filter fans) as well as specific fabricational constraints. The encrinid Traumatocrinus resembles isocrinid driftwood dwellers; but it had branching ramuli and non-cirrated stems. In Ctenocrinus, ramuli are ectotomous and radial support is enhanced by fusion, rather than adhesive coupling, of arms. In Scyphocrinites, suspended from a self-made root buoy, arms continued to branch isotomously throughout. (Modified from Seilacher and Hauff 2004; all scales 5 mm)
Taxonomically important differences between Traumatocrinus and Seirocrinus do not help to answer this question. The biseriality of brachial and ramular ossicles merely emphasizes the alternating positions of the pinnules, as expressed in pentacrinids by the even numbers of distancing arm ossicles (see above). The fact that in Traumatocrinus ramules continue to branch heterotomously like the arms (but alternating and with aboral spines on the axials) only compensates for the distal tapering of the ramules. Neither relevant is the common bend of the proximal stem in the shape of a walking stick. It is too common to be only a taphonomic artifact, and it is mostly observed in adult individuals. So the question regarding the life style of Traumatocrinus juveniles remains open. Perhaps the waves on windy days were sufficient to flush their small filter fans in the traditional way?
L. Devonian Ctenocrinus (Fig. 7b)
Traumatocrinus was an independent evolutionary experiment, but as a member of the Articulata it had at least a similar morphogenetic outfit as the driftwood Isocrinida. More of an outgroup is Ctenocrinus (formerly Melocrinites; Brett 1999). Nevertheless, it also enlarged the filter fan by heterotomous arm branching. However, the pinnulated ramuli are exotomous, while two firmly connected arms take over the radial support function that is provided by more loosely coupled arms in pseudoplanktonic Articulata.
The life style of Ctenocrinus is still uncertain, because stems found attached to a driftwood had no crowns and most of the known crowns are detached from their stems (Brett 1999). But this very fact could mean that the connection between stem and crown had been weakened by a “nodal factory” for faster growth. Columnals are also short and amphicoelic, presumably providing space for as yet unmineralized generations of internodals (Seilacher and Hauff 2004).
U. Silurian Scyphocrinites (Fig. 7c)
At a time when suitable driftwood was not yet available, Scyphocrinites became pelagic by transforming the root part of the stem into an air-filled lobolith (Haude 1972). The stems hanging down from this self-made buoy (which is mostly broken off in fossils; Hess 1999a) consist of short, amphicoelic columnals, while the crowns developed into a dense filter fan that probably remained splayed all the time. This trend is typical for tow-net filtrators. But in this case, the filtering area was not increased by heterotomous ramulation; rather the arms continued to bifurcate isotomously and together with their adhesive pinnules approached the same paradigm.
The end of driftwood crinoids
Driftwood dwellers have the advantage of reduced perturbations by sedimentational events and predators. Therefore, pseudoplanktonic crinoids could afford large and permanently open filter fans. On the other hand, they faced the risk of being torn-off, or stranded, during storms and to sink to the bottom as soon as their float became water-logged and overloaded. To delay this ultimate fate, they should (1) speed up their development to sexual maturity; and (2) reduce their own weight to minimize the ballast effect.
Seirocrinus accelerated growth in various ways. In the crown, new brachials were simultaneously introduced at a growing number of arm tips (Fig. 1). In the stem, several generations of new columnals matured simultaneously—be it as nodals in a chamber protected by spines and a “miniskirt” of small and cylindrical cirri, or as internodals between already mineralized amphicoelic colummals (Seilacher and Hauff 2004).
All these measures failed, however, when the advent of shipworms dramatically reduced the drifting time of wood in marine waters. The disappearance of Seirocrinus in the Upper Toarcian may be ascribed to this biological event. Pentacrinites possibly remained somewhat longer on board, but instead of becoming extinct, it returned to the benthic realm, as documented by its occurrence in the Swiss Hauptrogenstein (Bajocian). As illustrated by Hess (1999c) this is an oolithic Lagerstaette, in which articulated benthic echinoderms of various kinds are beautifully preserved. Between the rare obrutional events importing the ooids from more agitated environments during extreme storms, waters in this environment may have been rather quiet, so that an active filtrator would have had an advantage over its passively filtrating kins.
What transformations other than re-orientation should one expect in crinoids returning from a pseudoplanktonic to the benthic life style? According to “Dollo’s law” they could not easily get rid of the adaptations acquired on the drift wood. But at least they should speed up intercalational stem growth and reduce cirri above the root sector, where they would only be in the way of an ambiental current flushing the filter fan from the aboral side. As nothing like this is observed in Pentacrinites dargniesi, there are only two alternative explanations:
-
(1)
This species still lived on drift wood and happened to sink down into a lithotope that also favored the preservation of other, but benthic, echinoderms. This model requires that the sinking of Pentacrinites coincided with the influx of oolitic sediment into an otherwise quiet basin, which was responsible for the perfect echinoderm preservation (Obrution Lagerstätte).
-
(2)
P. dargniesi had again become benthic. Nothing, except the orientation, had to change, because the proposed active filtration, acquired on drift woods, turned out to be advantageous also on quiet bottoms, nor did an active filtrator have to increase elevation. The entanglement of cirri is probably a taphonomic effect and does not mean that the animals formed current-reducing thickets in life.
Conclusions
Although our information largely depends on spotty taphonomic windows, the history of pseudoplanktonic Isocrinida during late Triassic and Early Jurassic times is fairly well documented. The fossil record suggests that:
-
1.
Pentacrinites and Seirocrinus can be interpreted to represent alternative strategies (active filtration versus townet filtration) of pseudoplanktonic driftwood dwellers in response to scaling effects.
-
2.
The larger number of ossicles in interaxillary sections (Fig. 8), and other claimed differences between the two genera could be the result of secondary heterochronic shifts in the morphogenetic program.
Fig. 8 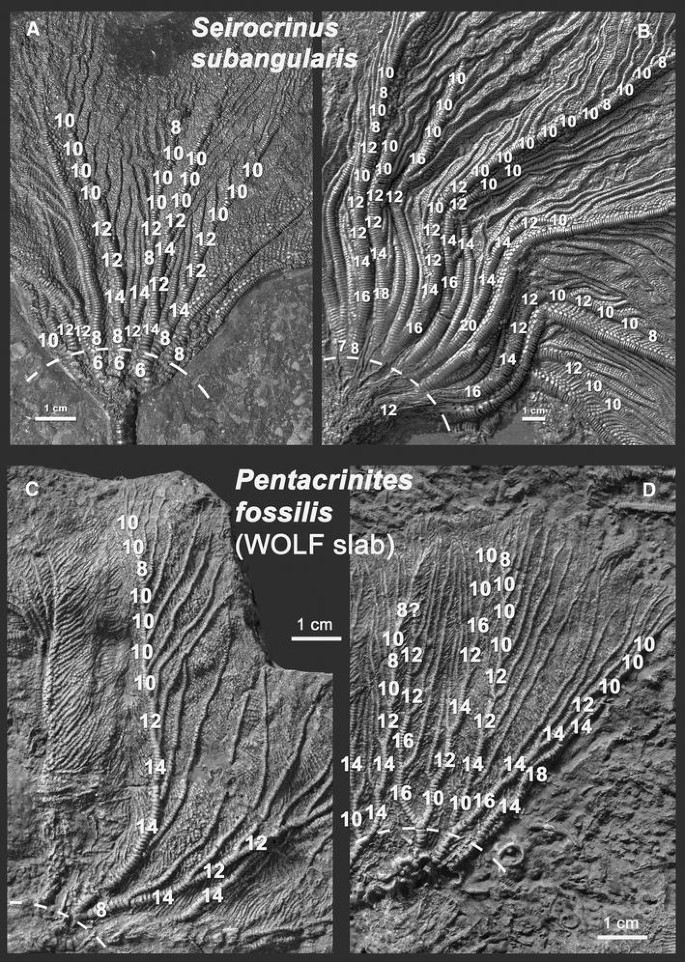
Numbers of brachials between axillaries could not secondarily change. They show certain trends along the arms and are always even in the ramulated zone. Still, the variability of these numbers and the difficulty to count them near the base of the arms (due to disarticulations and covering cirri) weakens their value as distinguishing character between Pentacrinites and Seirocrinus. (a, b, photos courtesy of Rolf Hauff, Holzmaden; c, d, photographed by Wolfgang Gerber from GPIT 1918/1)
-
3.
The two forms co-occur in a colony of Sinemurian P. fossilis and as isolated specimens in other horizons since the Upper Triassic (Norian). This supports the hypothesis that they represent stages in developmental history rather than different taxa also in later times.
-
4.
A two-stage development is also indicated by Pentacrinites-like root cirri of Seirocrinus and transitional stages of Pentacrinites in the Toarcian.
-
5.
The timing of this switch, however, varies. The Pentacrinites stage is absent in individuals attached to the proximal stem of an older Seirocrinus. In contrast, driftwood colonies from a slightly younger horizon stuck longer to active filtration and never became townet filtrators. They may still be distinguished at the species level.
-
6.
When the drifting time of logs became reduced (probably by the coming ot teredinid shipworms), first the larger Seirocrinus stage and then all crinoid driftwood dwellers disappeared.
-
7.
The Pentacrinites stage survived this biogenic event until Bathonian time by returning to the benthic realm, where its proposed feeding style would have been advantageous under quiet-water conditions, but again in an upright attitude.

In the future, this thypothetical scenario could be further tested by biometric studies (including inter-axillary ossicle numbers) in a larger material.
Throughout the present study, principles of constructional morphology (Seilacher 1971, 1991) have been applied. In this approach, the resulting functional reconstructions are often in conflict with what is known from the softpart anatomy of more or less closely related modern counterparts. In a way, the situation is comparable to the use of the living Nautilus as a model for the extinct ammonites: it should not be taken too literally (Seilacher and LaBarbera 1995). Models gained from the constructional morphology approach are necessarily hypothetical, but they can serve as guidelines for future research, for instance the analysis of articulation surfaces, which were not considered in the present paper.
References
Baumiller, T. K., & Ausich, W. I. (1996). Crinoid stalk flexibility: theoretical predictions and fossil stalk postures. Lethaia, 29, 47–59.
Brett, C. E. (1999). Lower Devonian Manlius/Coeymans Formation of Central New York, USA. In: Hess, H., Ausich, W. I., Brett, C. E., & Simms, M. J. (Eds.), Fossil crinoids (pp. 103–110).
Donovan, S. K. (1993). Contractile tissues in the cirri of ancient crinoids: critieria for recognition. Lethaia, 26, 169–183.
Fujiwara, S. I., Oji, T., Tanaka, Y., & Kondo, Y. (2005). Relay strategy and adaptation to a muddy environment in Isselicrinus (Isselicrinidae, Crinoidea). Palaios 20.
Hagdorn, H. (1996). Trias-Seelilien. Geologisch, Paläontologische Mitteilungen, Innsbruck, 21, 1–17.
Hagdorn, H., Wang, X., & Wang, C. (2007). Palaeoecology of the pseudoplanktonic Triassic crinoid Traumatocrinus from Southwest China. Palaeogeography, Plaeoclimatology, Palaeecology, 247, 191–196.
Haude, R. (1972). Bau und Funktion der Scyphocrinites-Lobolithen. Lethaia, 5, 95–125.
Hauff, R. B. (1984). Pentacrinus quenstedti (Oppel) aus dem oberen Untertoarcium (Lias Epsilon) von Ohmden bei Holzmaden (SW-Deutschland). Paläontologische Zeitschrift, 58, 255–263.
Hess, H. (1999a). Scyphocrinitids from the Silurian—Devonian boundary of Marocco. In: Hess, H., Ausich, W., Brett, C., & Simms, M. (Eds.), Fossil crinoids (pp. 93–102).
Hess, H. (1999b). Lower Jurassic Posidonia shale of Southern Germany. In: Hess, H., Ausich, W., Brett, C., & Simms, M. (Eds.), Fossil crinoids (pp. 183–196).
Hess, H. (1999c). Middle Jurassic of Northern Switzerland. In: Hess, H., Ausich, W., Brett, C., & Simms, M. (Eds.), Fossil crinoids (pp. 203–215).
Hess, H., Ausich, W., Brett, C., & Simms, M. (1999). Fossil crinoids (pp. 275). Cambridge University Press: Cambridge.
Rasskin-Gutman, D. & De Renzi, M. (2009). Pere Alberch, The creative trajectory of an Evo-Devo Biologist. Universitat de Valencia, Institut d’Estudis Catalans.
Seilacher, A. (1971). Arbeitskonzept zur Konstruktions-Morphologie. Lethaia, 3, 393–396.
Seilacher, A. (1991). Self-organizing mechanisms in morphogenesis and evolution. In: N. Schmidt-Kittler, K. Vogel (Eds.), Constructional morphology and evolution (pp. 251–271). Springer: London.
Seilacher, A., Drozdzewski, H., Haude, R. (1968). Form and function of the stem in a pseudoplanctonic crinoid (Seirocrinus). Palaeontology, 11, 275–282
Seilacher, A., & Hauff, R. B. (2004). Constructional morphology of pelagic crinoids. Palaios, 19, 3–16.
Seilacher, A., Hauff, R. B., & Wolf, M. (2007). Fest verwurzelt in schwimmendem Grund: Treibholz-Seelilien in Lias-Ölschiefern. Fossilien, 6, 349–355. Quelle und Mayer: Wiebelsheim.
Seilacher, A., & LaBarbera, M. (1995). Ammonites as Cartesian divers. Palaios, 10, 493–506.
Seilacher, A., & MacClintock, C. (2005). Crinoid anchoring strategies for soft-bottom dwelling. Palaios, 20, 224–240.
Simms, M. J. (1989). British Lower Jurassic crinoids. Monograph of the Palaeontographical Society, 142, 1–102. 15 Pls. London.
Simms, M. J. (1999). Pentacrinites from the Lower Jurassic of the Dorset coast of Southern England. In: Hess, H., Ausich, W., Brett, C., & Simms, M. (Eds.), Fossil crinoids (pp. 177–182).
Acknowledgments
This essay was written as a tribute to Dr. Dr.h.c. Hans Hess on the occasion of his 80th birthday. He is a model example for the role that private researchers have always played in paleontology. As a more knowledgeable expert (Hess et al. 1999 was a major source) he will no doubt disagree with some of my models. Another devoted hobby scientist, Dr. h.c Hans Hagdorn (Ingelfingen) critically read the manuscript at an earlier stage and made me aware of important publications. Thomas Baumiller and an anonymous reviewer helped with thoughfull critical remarks. This paper was prompted by the slab that M. Wolf (Lenningen-Gutenberg) inadvertently collected at the British coast. He carefully prepared it during months of hard work and allowed Hans Luginsland (preparator in the Geological Institut, Tübingen) to make a cast (GPIT 1918/1) before the original was sold to a private collector in England. I thank all these members of a family united by love for a most fascinating kind of fossils, crinoids. In addition I am obliged to Prof. Miguel de Renzi (University Valencia), who at the right moment provided me with a book about the principles of evo-devo research (Rasskin—Gutman and de Renzi, 2009).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Seilacher, A. Developmental transformations in Jurassic driftwood crinoids. Swiss J Palaeontol 130, 129–141 (2011). https://doi.org/10.1007/s13358-010-0012-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-010-0012-1