
Abstract
We examined the attraction of adults to six LEDs in the red flour beetles, Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae), which is widespread as a stored grain insect. In the experiment, bluish green and green were more attractive than the two UVs, blue, and red LEDs only in females. On the other hand, no difference was found in attraction among the six LEDs in males. Next, we investigated the relationship between light intensity and attractiveness. No sexual difference in attractiveness in different light intensities was found, but the strongest light intensity was more attractive than other light intensities. Finally, we investigated the relationship between light attraction and strains artificially selected for the duration of death feigning. Short-strain beetles were more attracted to UV lights than long-strain beetles.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The red flour beetle, Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae), is an important cosmopolitan pest species that damages stored wheat and rice flour (Campbell et al. 2010). Although many chemical insecticides and fumigants have been used for management, the beetles have developed resistance to those chemicals (Opit et al. 2012; Sokoloff 1977). Thus, new methods are necessary to manage this beetle species.
Recently, light has been used to trap and keep insects away from products, and recent studies have shown that insects respond differently to different light wavelengths (Shimoda and Honda 2013). The light response of stored product pests in the laboratory has been examined (Diaz-Montano et al. 2016; Hironaka et al. 2017; Katsuki et al. 2013) and also used to monitor the abundance of pests in a stored grain mill (see Miyatake et al. 2016).
On the other hand, behavioral responses to light may vary among insect populations, and different strains might respond differently to light, which is possible that all individuals are not equally attracted to light in the field. In fact, genetic variation in phototaxis has been reported in Drosophila flies (Dobzhansky and Spassky 1969; Hadler 1964). However, to our knowledge, no study has been conducted on the relationship between heritable variations of behavior and attraction to light sources using insect pest species.
Duehl et al. (2011) studied the attraction of T. castaneum to UV-LED lights. However, no studies have examined the effects of sex, and specific behavioral characteristics on the light-induced attraction of T. castaneum. Here, we measured (1) sexual differences in the distance of attraction to several different colors of LED light, (2) the relationships between the attractiveness and light intensities of LED light in male and female individuals, and (3) the difference in the distance of attraction to several different colors of LED lights of two strains artificially selected for the death-feigning duration, long and short, in T. castaneum adults (see Miyatake et al. 2004, 2008).
Materials and methods
Insects
Three strains of T. castaneum were used in the experiments: a “base”, which has been kept in the laboratory of Okayama University for more than 20 years, and “short” (S-strain) and “long” (L-strain) strains, which have been artificially selected for more than 10 years for the duration of death feigning (Matsumura and Miyatake 2018).
The beetles were reared with a mixture of whole-meal (Nisshin Seifun Group, Tokyo, Japan) enriched with brewer’s yeast (Asahi Beer, Tokyo, Japan) as the rearing medium and kept in a chamber (Model-470D Hitachi, Osaka, Japan) maintained at 28 ℃ under a photoperiod of 16L:8D.
Light stimuli for behavioral experiments
We used brand new LEDs (Nichia Corp. Tokushima, Japan) and examined the attraction of the base population of T. castaneum to different wavelengths using six LEDs: UV-L (NSHU590B 365 nm), UV-H (NSPU510CS 375 nm), blue (NSPB510 470 nm), bluish green (NSPB510DS 505 nm), green (NSPG510AS 520 nm), and red (NSPR510CS 640 nm). An LED was set on an optical bench equipped with a set of quartz ND filters, a quartz optical wedge, and a UV-transparent liquid fiber, which directed the light to the experimental arena (see Fig. 1A). In the present experiment, the numbers of photons were equalized to compare the attractiveness of LEDs accurately. First, we measured the photon flux of each LED at its maximal intensity with a spectrophotometer (HSU-100S; Asahi Spectra, Tokyo, Japan). The photon numbers were compared based on the integrated value in a range from 350 to 750 nm. The integral photon flux value was the lowest in UV-L LED (15.25 × 1015 photons cm−2 s−1), so we adjusted the photon flux of all other LEDs to these values as closely as possible using the ND filters and the optical wedge. The resulting photon fluxes (UV-L: 15.25 × 1015 photons cm−2 s−1, UV-H: 14.01 × 1015 photons cm−2 s−1, blue: 16.55 × 1015 photons cm−2 s−1, bluish green: 15.89 × 1015 photons cm−2 s−1, green: 14.06 × 1015 photons cm−2 s−1, and red: 16.66 × 1015 photons cm−2 s−1) were used as the “maximal equiquantal” intensities in this study.

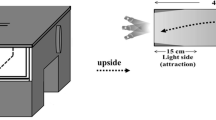
Equipment measuring insect attraction to different light intensities and wavelengths. A A set of quartz ND filters, a quartz optical wedge, and a UV-transparent liquid fiber, which led the light to the experimental arena, B the box used for the attraction experiment, and C the release point (1 cm × 1 cm) of the beetles, the position of the beetle after 1 h, and the distance to the beetle from the surface where an LED light is set
The behavioral experiments were conducted in a dark room (24 ± 1 ℃, humidity 45 ± 5%) using a styrene foam box (22 cm in height × 28 cm width × 35 cm in length). We set one LED on the optical bench at a time (Fig. 1A) and changed the LED to alter the stimulus wavelengths. We made a hole at the bottom center of a wall (wall 1, see Fig. 1B, C) of the box to attach the exit pupil of the optical fiber (Fig. 1A). When necessary, we systematically changed the light intensity using the ND filters and the optical wedge on the bench.
Attraction of beetles to six different LEDs
We set a release point (1 cm × 1 cm) on the floor at the bottom center of wall 2 (Fig. 1C). Twenty each male and female T. castaneum individuals of the base strain were randomly selected from 80 individuals each time, separately. The sexes were separated during the pupal period by the difference in genital organs and kept for 5 days after eclosion as virgins.
We put the lid on after we set the beetle in the release point, and then turned on the LED. 1 h after the light was turned on, we opened the box and photographed the inside from just above the box using a smartphone (iPhone 10, Apple, USA). In the photograph, we drew a line from each beetle perpendicular to the wall to measure the distance from wall 1 to each beetle (see dotted lines in Fig. 1C). Then, we compared the averages and standard errors of the distances of individuals in each wavelength.
We used the distances of each individual as the attraction index. The attraction index was defined as a reciprocal of the distance from wall 1 to each beetle that was attracted to an LED. If a beetle never moved, then we scored 0 for the beetle. If a beetle reaches wall 1, we noted it as 1. The graphs are presented using attraction indices rather than raw data to show them visually more clearly (see Figs. 2, 3, 4). We used the Steel–Dwass test for the distances to compare sexes and among all six LED colors using JMP version 12.2 (SAS Institute Inc. 2015). The Wilcoxon rank-sum test was used to compare the sexual difference overall, or in each wavelength by JMP version 12.2 (SAS Institute Inc. 2015). Statistical tests for distances, on the other hand, were performed on raw data.

Comparison of attraction of the red flour beetle to different light LEDs. The vertical axis is the attraction index. The letter on the graph is the result of the Steel–Dwass test. There was a significant difference between those with different letters. The asterisk on the graph shows the result of the Wilcoxon rank-sum test. There was a sex difference in wavelength with an asterisk. The white bar shows male results, and the black bar shows female results. The error bar is SE

Comparison of attraction of the red flour beetle to different light intensities in males and females. The horizontal axis shows the number of photons, and the vertical axis is the attraction index. The asterisk on the graph shows the result of the Steel test. The white bar shows male results, and the black bar shows female results. The data bar is SE

Comparison of attraction to different light LEDs of S- and L-strains of the red flour beetle: the results of a 1 h and b 3 h at the insect release. The vertical axis is the attraction index. The letter on the graph is the result of the Steel–Dwass test. There was a significant difference between those with different letters. The asterisk on the graph shows the result of the Wilcoxon rank-sum test. There was a significant difference between strains in wavelength with an asterisk. The white bar shows S-results, and the black bar shows L-results. The error bar is SE
Light intensity dependency of attraction
We tested the attraction at four different light intensities (1.96 × 1015 → 1.44 × 1016 → 1.35 × 1017 → 1.51 × 1018 [photons cm−2 s−1]) to the green LED using the base strain. The procedure was similar to the experiments described above. The Steel test was used to test the relationships between photon flux and attraction distance by JMP version 12.2 (SAS Institute Inc., 2015). The Wilcoxon rank-sum test was used to compare the sexual difference overall, or in each light intensity by JMP version 12.2 (SAS Institute Inc., 2015).
Comparison of attraction for short and long strains
We examined the attractiveness of six different LEDs to virgin beetles derived from short (S) and long (L) strains, which were artificially selected for the duration of death feigning. Each strain has two independent replicate selection lines (see Miyatake et al. 2004). The experimental procedure was the same as above except that we measured the distance twice at 1 h and 3 h after turning on the light to consider the effect of the long death-feigning duration of L-strains. The Steel–Dwass test was used to compare the attraction distance between the strains and the effects of LEDs by JMP version 12.2 (SAS Institute Inc., 2015). The Wilcoxon rank-sum test was used to compare the difference of strains overall, or in each wavelength by JMP version 12.2 (SAS Institute Inc., 2015). In the comparison between S- and L-strains, we used only females because only females had shown a wavelength preference in the previous experiment (see Fig. 2).
Results
Attraction of beetles to six different LEDs
Figure 2 shows the attractiveness of different LED colors to males and females. Statistical analysis showed that females were attracted to LEDs more strongly than males (Wilcoxon rank-sum test, male–female, p < 0.0001). A sexual comparison of each wavelength showed sexual differences in blue, bluish green, green, and red (Fig. 2, Wilcoxon rank-sum test, male–female, blue, p = 0.0015; bluish green, p < 0.0001; green, p < 0.0001; red, p < 0.0001). In males, no significant difference was found in attraction distances among the six LEDs (Fig. 2, Steel–Dwass, colors: p > 0.05). On the other hand, bluish green and green LEDs were more attractive to females than UV-L, UV-H, and blue LEDs (Fig. 2, Steel–Dwass, UV-L–bluish green, p = 0.0004; UV-H–bluish green, p = 0.0022; UV-L–green, p = 0.0019; UV-H–green, p = 0.0114). Moreover, red LED was more attractive compared to UV-L and UV-H LEDs (Fig. 2, Steel–Dwass, UV-L–red, p = 0.0012; UV-H–red, p = 0.0088).
Light intensity dependency of attraction
Figure 3 shows the effects of light intensity on males and females measured using the green LED. This is because the green LED was one of the most attractive colors in the previous experiment (see Fig. 2). We found no significant differences in attraction indices between sexes (Wilcoxon rank-sum, male–female, p > 0.05). In the comparison of light intensity, we found a significant difference between the weakest intensity and the strongest intensity (Fig. 3, Steel, 1.96 × 1015–1.51 × 1018, p = 0.0428). In the Steel test, 1.96 × 1015 was used as the criterion. In addition, the Steel test was performed on combined male and female data because no sex differences were found.
Comparison of attraction in S- and L-strains
We used only females, which showed the wavelength preference in the base strain (see Fig. 2).
Figure 4 shows the attractiveness of different LED colors to S- and L-strains. The S-strain individuals were attracted more strongly to LED other than those of L-strains at 1 h (Wilcoxon rank-sum test, S–L, p < 0.0001) and 3 h (Wilcoxon rank-sum test, S–L, p = 0.0005). A strain comparison of each wavelength showed significant differences in UV-L, blue, and green at 1 h (Fig. 4a, Wilcoxon rank-sum test, S–L, UV-L, p = 0.0101; blue, p = 0.0396; green, p = 0.0306). At 3 h, a strain comparison of each wavelength showed significant differences in UV-H, blue and green (Fig. 4b, Wilcoxon rank-sum test, S–L, UV-H, p = 0.0374; blue, p = 0.0093; green, p = 0.0316). In addition, UV-L was more attractive to the S-strains compared to UV-H and red at 1 h after the light was turned on (Fig. 4a, Steel–Dwass, UV-H–UV-L, p = 0.0362; red–UV-L, p = 0.0321). At 3 h after the light was turned on, UV-L and blue were more attractive than UV-H and red in the S-strain (Fig. 4b, Steel–Dwass, UV-H–UV-L, p = 0.0496; red–UV-L, p = 0.0240, UV-H–blue, p = 0.0118; red–blue, p = 0.0119). We detected no significant difference among LED color preferences in the L-strains at 1 h (Fig. 4a, Steel–Dwass, colors: p > 0.05). At 3 h, only UV-H was less attractive than UV-L (Fig. 4b, Steel–Dwass, UV-H–UV-L, p = 0.0129).
Discussion
In the present study, a significant sex difference was found in the attractiveness of LEDs (Fig. 2). In some species, sexual differences in responses to light were found (Imafuku et al. 2007; Nowinszky and Puskas 2015), so sex is possibly an important factor in light attraction. Furthermore, previous studies showed sex differences at the level of photoreceptor spectral sensitivity (van der Kooi et al. 2021). Further studies are required to understand the mechanism of sexual differences in attraction to LEDs. In terms of wavelength preferences, the attraction to bluish green (505 nm), green (520 nm), and red (640 nm) was higher in females, but not in males (Fig. 2). A few previous studies of other species show that wavelength preferences are different between the sexes (Endo et al. 2014; Katsuki et al. 2013; Ogino et al. 2015). Because the mechanism and function of such a sexual difference in attraction to LEDs are unknown, further detailed research is needed to determine these.
The results of the present experiment differ from those of previous studies on attractive wavelengths in T. castaneum. For example, Duehl et al. (2011) conducted experiments equalizing spectral irradiance (W/cm2) and showed that T. castaneum was attracted more to UV lights (390 nm) than other lights including 360, 380, 410, 555, 587, and 655 nm. On the other hand, Song et al. (2016) conducted experiments equalizing luminosity (lux) and concluded that T. castaneum was more attracted to red light (625 nm) than other lights, including 470, 520, 590, and 625 nm.
Note that the energy of a photon is inversely proportional to light wavelength. For example, the energy of one photon of 350 nm is equal to that of two photons of 700 nm (Arikawa et al. 2014). When spectral irradiance is equalized, the numbers of photons decrease as wavelengths become shorter. If Duehl et al. (2011) conducted their experiments with equiquantal stimuli, as we have done here, the attractiveness of 360 nm and 380 nm would have been a little enhanced, but the predominance of 390 nm would remain. Therefore, using irradiance instead of photon flux is completely acceptable because both values are convertible. However, using luminosity for studying vision in non-human organisms is inappropriate because luminosity is a measure of brightness for humans. Any illuminometer is equipped with a sensor whose sensitivity peaks at 500 nm and zeros at 400 nm and 700 nm. When the luminosity of LED (470, 520, 590, 625 nm) is equalized using this device as Song et al. (2016) did, 470 nm and 625 nm light would contain many more photons than 520 nm, for example. The high attractiveness of 625 nm lights that Song et al. (2016) observed is presumably attributed to the light measurement protocol. In any case, the high attractiveness of 390 nm obtained by Duehl et al. (2011) is not consistent with our result here (Fig. 2). The difference might be attributed to the paradigm difference in light conditions of both behavioral experiments. UV used in the present study were 365 nm and 375 nm, which were different from UV (390 nm) attractive to T. castaneum in Duehl et al. (2011). Then, Duehl et al. (2011) adjusted the spectral irradiance of all LED lights to 10mW/cm2. On the other hand, we adjusted the photon flux of all LEDs to about 15 × 1015 photons cm−2 s−1. The spectral irradiance of each LED we adjusted was about 0.5–0.8 μW/cm2. The light intensity of our experiments with six different LEDs was about 10,000 times lower than those of Duehl et al. (2011). The light intensity influences the light response of insects (Mellor and Hamilton 2003; Merling et al. 2020; Meyer 1978). It is possible that responsiveness to UV was lower than green in the light intensity we adjusted. In addition, we compared the average distance from the wall set light source in the box to each beetle for 1 h, while Duehl et al. (2011) compared the number of beetles that visited LED lights for 5 min. These differences might be related to the difference in the results of spectral preference between Duehl et al. (2011) and the present experiments.
No sexual differences in attraction by light intensity were found, although the attraction to green LED increased as the value of light intensity in males and females (Fig. 3). We found that the light intensity influences the light response of insects in T. castaneum. Moreover, the attraction of females rapidly tended to increase in a change from 1.96 × 1015 to 1.44 × 1016, while the attraction of males gradually tended to increase as the value of light intensity. Thus, in T. castaneum, the threshold of the optical response may differ by sex. These would provide a useful perspective on pest control by light attraction, as light intensity is related to phototaxis and wavelength preference in insects, and the optimum light intensity for attraction may differ between light wavelengths and sexes. In sandfly, Lutzomyia longipalpis, it has been observed that wavelength preference varies with light intensity in males and females (Mellor and Hamilton 2003). On the other hand, a previous study with the blowfly, Calliphora erythrocephala has confirmed that positive phototaxis is maximized at a particular light intensity in males (Meyer 1978). Therefore, it is necessary to verify in the future how wavelength preference changes in females and whether wavelength preference is observed in males when similar attraction experiments are conducted at higher reference light intensities.
In the experiment that compared short with long strains using adult females, the L-strain individuals were less attracted than those of S-strains. A previous study showed beetles derived from the L-strains had lower levels of activity than beetles from S-strains (Miyatake et al. 2008). These results suggest that there seems to be a relationship between activity and attraction to light. Therefore, it is possible that the colors such as UV, blue, and green preferred by the beetles (Fig. 2) differed significantly between S- and L-strains (Fig. 4).
On the other hand, the death-feigning duration of wild-caught T. castaneum is basically similar to that of the S-strain (Konishi et al. 2020). Therefore, a trapping and monitoring system using LED lights may be effective to trap wild beetles. However, the base and selected lines differed in their attractiveness to light wavelengths (See Figs. 2 and 4). The reason for this is unclear, but the attractiveness to light wavelengths may change during long periods of successive rearing or artificial selection. In addition, the wavelength preferences of the base and selected lines are possible to differ from that of the wild type. Therefore, we need to conduct future experiments in the wild type of T. castaneum.
In this study, the number of insects attracted was recorded 1 h after the insects were released. Previous studies using other species of beetles have set longer experimental durations than the present study, with some of the longer ones lasting more than 24 h (Jeon et al. 2012; Katsuki et al. 2013; Song et al. 2016). However, as the wavelength preference was confirmed even for as short a time as 1 h, the duration of the present experiment is considered sufficient for the assessment of phototaxis. Also, in the comparison of attraction in S- and L-strains, the time of the experiment was set up as 1 h and 3 h in consideration of the effect of the long death-feigning duration of L-strains. The fact that there is little difference between the data after 1 h and 3 h suggests that the results of this study adequately assess positive phototaxis, as it is likely that T. castaneum remains in that position after being attracted to some extent. However, we need to conduct more detailed experiments on the behavior of attraction to light over time in this insect in the future.
References
Arikawa K, Wakakuwa M, Kinoshita M (2014) Spectral sensitivity of insect photoreceptors. Jpn J Appl Entomol Zool 58:5–11 (in Japanese with English abstract)
Campbell JF, Toews MD, Arthur FH, Arbogast RT (2010) Long-term monitoring of Tribolium castaneum in two flour mills: seasonal patterns and impact of fumigation. J Econ Entomol 103:991–1001
Diaz-Montano J, Campbell JF, Phillips TW, Cohnstaedt LW, Throne JE (2016) Evaluation of light attraction for the stored-product psocid, Liposcelis bostrychophila. J Pest Sci 89:923–930
Dobzhansky T, Spassky B (1969) Artificial and natural selection for two behavioral traits in Drosophila pseudoobscura. Proc Nat Am Sci 62:75–78
Duehl AJ, Cohnstaedt LW, Arbogast RT, Teal PEA (2011) Evaluating light attraction to increase trap efficiency for Tribolium castaneum (Coleoptera: Tenebrionidae). J Econ Entomol 104:1430–1435
Endo N, Wakakuwa M, Arikawa K, Hironaka M (2014) Spectral preference in a free-flying condition of the southern green stink bug, Nezara viridula (Heteroptera: Pentatomidae). Jpn J Appl Entomol Zool 58:23–28 ((in Japanese with English abstract))
Hadler NM (1964) Heritability and phototaxis in Drosophila melanogaster. Genetics 50:1269–1277
Hironaka M, Kamura T, Osada M, Sasaki R, Shinoda K, Hariyama T, Miyatake T (2017) Adults of Lasioderma serricorne and Stegobium paniceum (Anobiidae: Coleoptera) are attracted to ultraviolet (UV) over blue light LEDs. J Econ Entomol 110:1911–1915
Imafuku M, Shimizu I, Imai H, Shichida Y (2007) Sexual difference in color sense in a lycaenid butterfly, Narathura japonica. Zool Sci 24:611–613
Jeon J, Oh M, Cho K, Lee H (2012) Phototactic response of the rice weevil, Sitophilus oryzae Linnaeus (Coleoptera: Curculionidae), to light-emitting diode. J Korean Soc Appl Biol Chem 55:35–39
Katsuki M, Arikawa K, Wakakuwa M, Omae Y, Okada K, Sasaki R, Shinoda K, Miyatake T (2013) Which wavelength does the cigarette beetle, Lasioderma serricorne (Coleoptera: Anobiidae), prefer? electrophysiological and behavioral studies using light-emitting diodes (LEDs). Appl Entomol Zool 48:547–551
Konishi K, Matsumura K, Sakuno W, Miyatake T (2020) Death feigning as an adaptive anti-predator behavior: further evidence for its evolution from artificial selection and natural populations. J Evol Biol 33:1120–1128
Matsumura K, Miyatake T (2018) Responses to relaxed and reverse selection in strains artificially selected for duration of death-feigning behavior in the red flour beetle, Tribolium castaneum. J Ethol 36:161–168
Mellor HE, Hamilton JGC (2003) Navigation of Lutzomyia longipalpis (Diptera: Psychodidae) under dusk or starlight conditions. Bull Entomol Res 93:315–322
Merling M, Eisenmann S, Bloch G (2020) Body size but not age influences phototaxis in bumble bee (Bombus terrestris, L.) workers. Apidologie 51:763–776
Meyer HW (1978) Phototaxis in the walking male and female fly (Calliphora erythrocephala Meig.). J Comp Physiol 123:307–314
Miyatake T, Katayama K, Takeda Y, Nakashima A, Sugita A, Mizumoto M (2004) Is death-feigning adaptive? Heritable variation in fitness difference of death-feigning behavior. Proc R Soc B 271:2293–2296
Miyatake T, Tabuchi K, Sasaki K, Okada K, Katayama K, Moriya S (2008) Pleiotropic anti-predator strategies, fleeing and feigning death, correlated with dopamine levels in Tribolium castaneum. Anim Behav 75:113–121
Miyatake T, Yokoi T, Fuchikawa T, Korehisa N, Kamura T, Nanba K, Ryouji S, Kamioka N, Hironaka K, Osada M, Hariyama T, Sasaki R, Shinoda K (2016) Monitoring and detecting the cigarette beetle (Coleoptera: Anobiidae) using ultraviolet (LED) direct and reflected lights and/or pheromone traps in a laboratory and a storehouse. J Econ Entomol 109:2551–2560
Nowinszky L, Puskas J (2015) Sex ratio analysis of some Macrolepidoptera species collected by Hungarian forestry light traps. Acta Silv Lign Hung 11:99–110
Ogino T, Uehara T, Yamaguchi T, Maeda T, Noro C, Shimoda M (2015) Spectral preference of the predatory bug Orius sauteri (Heteroptera: Anthocoridae). Jpn J Appl Entomol Zool 59:10–13 ((in Japanese with English abstract))
Opit GP, Phillips TW, Aikins MJ, Hasan MM (2012) Phosphine resistance in Tribolium castaneum and Rhyzopertha dominica from stored wheat in Oklahoma. J Econ Entomol 105:1107–1114
SAS Institute Inc (2015) JMP 12.2.0. SAS Institute Inc., Cary
Shimoda M, Honda K (2013) Insect reactions to light and its applications to pest management. Appl Entomol Zool 48:413–421
Sokoloff A (1977) The biology of Tribolium with special emphasis on genetic aspects, vol 3. Clarendon Press, Oxford
Song J, Jeong E, Lee H (2016) Phototactic behavior 9: phototactic behavioral response of Tribolium castaneum (Herbst) to light-emitting diodes of seven different wavelengths. J Appl Biol Chem 59:99–102
van der Kooi CJ, Stavenga DG, Arikawa K, Belušič G, Kelber A (2021) Evolution of insect color vision: from spectral sensitivity to visual ecology. Ann Rev Entomol 66:435–461
Acknowledgements
We thank Prof. Kentaro Arikawa (Research Center for Integrative Evolutionary Science, SOKENDAI) for advice on experiments and for the use of measuring equipment materials and Dr. Kentarou Matsumura (Okayama University) for advice on statistical methods. We also thank Eisuke Takada for helping us with the experiment. This study was funded by the Japan Society for the Promotion of Science, KAKENHI Grant Numbers 21H02568 and 21K19116 (to TM).
Funding
Open access funding provided by Okayama University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sone, S., Miyatake, T. Sex and strain-specific spectral attraction of Tribolium castaneum (Coleoptera: Tenebrionidae): behavioral studies. Appl Entomol Zool 58, 257–263 (2023). https://doi.org/10.1007/s13355-023-00829-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13355-023-00829-6