
Abstract
A relatively complete succession of the Bashkirian to Moscovian rocks is exposed in the Hadim Nappe, central Taurides, where rich and diverse fusulinid assemblages have been analysed in three sections (Yassıpınar, Gölbelen and Bademli). The Bashkirian–Moscovian successions mainly consist of medium-bedded sandstones, medium–thick-bedded oolitic limestone and thin–medium bedded fossiliferous limestones with medium–thick-bedded dolomite levels (Bademli section). Eight first occurrence fusulinid biozones are identified in three sections in the Bashkirian to Moscovian strata. The Bashkirian Stage comprises in ascending order the Plectostaffella jakhensis-Plectostaffella bogdanovkensis, Pseudostaffella antiqua-Pseudostaffella sofronizkyi, Staffellaeformes staffellaeformis-Staffellaeformes parva parva, and Tikhonovichiella tikhonovichi-Verella spicata zones, the Moscovian Stage contains in ascending order the Aljutovella aljutovica-Solovievaia ovata ovata, Aljutovella priscoidea-Eofusulina triangula, Fusulinella vozhgalensis devexa-Beedeina schellwieni, and Fusulinella bocki bocki zones. These fusulinid zones, and their characteristic index species, allow to correlate the Bashkirian–Moscovian strata in the Hadim Nappe with coeval strata in the Donets Basin, southern China, southern Urals, and especially, with the Russian Platform.
Similar content being viewed by others


Introduction
The Taurides, part of the Alpine-Himalayan orogenic belt, extend approximately 200 km parallel to the Mediterranean Sea coast in Anatolia and consist of a number of allochthonous and autochthonous sequences with different depositional features that were assigned to nappes (Blumenthal 1947, 1951; Brunn et al. 1971, 1973; Monod 1977; Gutnic et al. 1979) or tectonostratigraphic units (Özgül 1976, 1984, 1997). The Palaeozoic units of the Taurides consist of well-exposed and mainly continuous successions containing significant fossils that enable detailed biostratigraphical implications. The fusulinid biostratigraphy of the Hadim Nappe, which is one of the allochthonous units of the Taurides, has been intensively studied (Güvenç 1965; Monod 1977; Altıner 1981; Okuyucu 1997, 1999, 2002, 2008, 2009, 2013; Peynircioğlu 2005; Atakul-Özdemir 2006; Dzhenchuraeva and Okuyucu 2007; Kobayashi and Altıner 2008a, 2008b; Atakul-Ozdemir et al. 2011; Kobayashi 2011; Demirel 2012; Demirel and Altıner 2016). However, in spite of the many studies so far carried out, a complete and detailed local fusulinid zonation for the Lower–Middle Pennsylvanian successions of this area is still not available.
The aim of this study is to describe the fusulinid zones of the Bashkirian–Moscovian successions from the Hadim Nappe and to correlate them with coeval successions in the Tethyan Realm. For this purpose, three selected sections (Yassıpınar, Gölbelen and Bademli) have been studied in the Central Taurides.
Geological settings
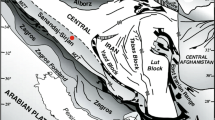
The Taurides are geographically subdivided into three parts, namely Eastern, Central and Western Taurides (Özgül 1976, 1984) (Fig. 1a). They consist of Beydağları-Anamas-Akseki Autochthonous, Beyşehir-Hoyran Nappes, Bolkar Dağ Nappe, Hadim Nappe, Antalya Nappes and Alanya Nappes (Fig. 1b) which are characterized by distinct stratigraphical, structural and metamorphic features (Monod 1977; Özgül 1971, 1997; Güvenç 1965, 1977; Turan 1990). The Hadim Nappe is composed of mainly carbonate-dominated sedimentary rocks, indicating shallow marine platform deposits from Mid(?) Devonian to Late Cretaceous time interval (Fig. 2).

a The broad geographical subdivision of the Tauride Belt (Özgül, 1984). b Schematic map showing the distribution of autochthonous and allochthonous sequences in the area between Western and Central Taurides (simplified and revised after Özgül, 1984) and location of the studied sections (Yassıpınar, Gölbelen and Bademli)

Investigated sections
The studied three sections are located in the Central Taurides (southern Turkey) and expose Upper Mississippian–Upper Pennsylvanian successions of the Hadim Nappe (Fig. 1b).
Yassıpınar section
The Yassıpınar section is located about 20 km southwest of the Hadim Town in Konya Province, Turkey (UTM coordinates on 1/25.000 scaled Alanya O28–b2 topographical map: between 36S 444875E/4085434N and 36S 444997E/4085048N). The total thickness of the section is about 75 m, and 37 samples were collected from this section for fusulinid biostratigraphy.
The upper Serpukhovian–lower Bashkirian interval of the section is represented by oolitic limestone and quartz arenite sandstone intercalation. The base of this interval is dominated by medium–thick-bedded grey/beige-coloured oolitic limestone while the top is mainly represented by thick-bedded yellow-coloured sandstone layers. The upper Bashkirian–Moscovian part of the section is characterized by thick-bedded grey-coloured fusulinid-rich limestones. The uppermost part of the section is represented by medium–thick-bedded sandstones at the base and medium-bedded, yellow-coloured limestones at the top (Fig. 3).

Lithostratigraphic characteristics, biostratigraphic zonation and fusulinid distribution of the Yassıpınar stratigraphic section
Gölbelen section
The Gölbelen section is located about 15 km southwest of the Bozkır Town in Konya Province, Turkey (UTM coordinates on 1/25.000 scaled Konya N28–d4 topographical map: between 36S 419453E/4100306N and 36S 419809E/4100450N). The total thickness of the section is about 108 m, and 47 samples were collected from this section.
The base of the section corresponding to the lower Bashkirian is represented by an alternation of medium-bedded sandstones, medium–thick-bedded and grey/beige-coloured oolitic limestones, and grey/beige thin-medium bedded fossiliferous limestones. The upper Bashkirian–Moscovian interval of the section is characterized by medium–thick-bedded grey/beige-coloured limestones at the lower part, thick-bedded grey/beige-coloured and fossil-rich limestones at the middle part, and medium-bedded yellow-pink-coloured at the upper part. The uppermost part (lower Kasimovian) of the section is composed of thick sandstone layers that are topped by thin–medium-bedded, yellow-coloured limestones (Fig. 4).

Lithostratigraphic characteristics, biostratigraphic zonation and fusulinid distribution of the Gölbelen stratigraphic section
Bademli section
The Bademli section is located about 30 km northeast of the Akseki Town in Antalya Province, Turkey (UTM coordinates on 1/25.000 scaled Konya N27–a3 topographical map: between 36S 388776E/4130367N and 36S 388897E/4130590N). The total thickness of the section is about 64 m, and 27 samples were collected from this section.
The lower part of the section, from upper Serpukhovian to the basal Moscovian (Vereian), is represented by medium–thick-bedded and grey-coloured dolostones, while the Moscovian interval is characterized by fusulinid-rich limestones. The lower Moscovian (Kashirian) is composed of medium–thick-bedded grey-coloured limestones with rare fusulinids. The upper Moscovian (Podolskian–Myachkovian) is represented by medium–thick-bedded grey/rarely pink-coloured limestones (particularly Myachkovian) that contain a rich and diverse fusulinid fauna. The lower Kasimovian part of the section is characterized by thick-bedded sandstones at the base, which are overlain by medium-bedded grey/yellow-coloured limestones (Fig. 5).

Lithostratigraphic characteristics, biostratigraphic zonation and fusulinid distribution of the Bademli stratigraphic section
Biostratigraphy
The Bashkirian–Moscovian carbonate-dominated succession of the Hadim Nappe is very suitable for establishing a detailed biostratigraphy in terms of fusulinid assemblages (Akbaş and Okuyucu 2021; Akbaş and Okuyucu 2022, in press). In this study, only the most significant index fusulinid taxa are illustrated. The described fusulinid assemblages allow a subdivision of the studied strata into eight first occurrence fusulinid zones for the Bashkirian–Moscovian successions of the Hadim Nappe (Figs. 3, 4, 5, 6). These are, Plectostaffella jakhensis-Plectostaffella bogdanovkensis First Occurrence Zone, Pseudostaffella antiqua-Pseudostaffella sofronizkyi First Occurrence Zone, Staffellaeformes staffellaeformis-Staffellaeformes parva parva First Occurrence Zone, Tikhonovichiella tikhonovichi-Verella spicata First Occurrence Zone, Aljutovella aljutovica-Solovievaia ovata ovata First Occurrence Zone, Aljutovella priscoidea-Eofusulina triangula First Occurrence Zone, Fusulinella vozhgalensis devexa-Beedeina schellwieni First Occurrence Zone and Fusulinella bocki bocki First Occurrence Zone, in ascending order. These first occurrence zones are described between first the occurrence datum (FOD) of different index taxa, and are almost identical to those previously identified in Russian Platform (Kagarmanov and Donakova 1990) and Urals (Kulagina et al. 2001).

Total stratigraphic distribution of fusulinids and biostratigraphic zonation from the Bashkirian to Moscovian in this study
Plectostaffella jakhensis-Plectostaffella bogdanovkensis First Occurrence Zone
Description
The base of this zone is marked by the FOD of Plectostaffella jakhensis (Reitlinger, 1971), Plectostaffella bogdanovkensis Reitlinger, 1980 and Plectostaffella varvariensis (Brazhnikova and Potievskaya, 1948), and its top by the FOD of Pseudostaffella antiqua (Dutkevich, 1934b). In addition to the index taxa, this zone comprises Eostaffella designata (Zeller, 1953), E. ovoidea (Rauzer-Chernousova, 1948), E. postmosquensis acutiformis Kireeva in Rauzer-Chernousova et al., 1951, E. postmosquensis postmosquensis Kireeva in Rauzer-Chernousova et al., 1951, E. cf. pseudoovoidea Reitlinger, 1980, E. pseudostruvei angusta Kireeva in Rauzer-Chernousova et al., 1951, E. ex gr. pseudostruvei pseudostruvei (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., 1936), Plectostaffella cf. evolutica (Rumyantseva, 1970), Semistaffella minuscilaria (Reitlinger, 1971), S. primitiva (Reitlinger, 1961), S. variabilis (Reitlinger, 1961), Varistaffella eostaffellaeformis (Rumyantseva, 1970), V. ziganica (Sinitsyna, 1975), and Pseudostaffella uralica Kireeva in Rauzer-Chernousova et al., 1951. The zone is approximately equivalent to the Syuranian of the early Bashkirian in Russia (Figs. 3, 4, 6, 7a–s).

Thin–section photomicrographs of the fusulinid assemblages of the Plectostaffella jakhensis-Plectostaffella bogdanovkensis Zone (a–s) and Pseudostaffella antiqua-Pseudostaffella sofronizkyi Zone (t–ac). a Eostaffella designata, axial section, 15Y06.04.02, YS; b–c Eostaffella postmosquensis postmosquensis, b axial section, 15Y05.04.02, YS, c subaxial section, 18G21.04.03, GS; d Eostaffella postmosquensis acutiformis, subaxial section, 15Y06.03.01, YS; e Eostaffella pseudostruvei angusta, subaxial section, 15Y10.03.02, YS; f Plectostaffella cf. evolutica, subaxial section, 15Y05.04.01, YS; g–h Plectostaffella jakhensis, g axial section, 15Y05.03.01, YS, h subaxial section, 15Y06.03.02, YS; i Eostaffella ovoidea, axial section, 15Y12.08, YS; j Eostaffella ex gr. pseudostruvei pseudostruvei, subaxial section, 15Y08.06.01, YS; k–l Plectostaffella bogdanovkensis k axial section, 15Y07.04.01, YS, l axial section, 15Y08.02, YS; m Plectostaffella varvariensis, subaxial section, 15Y08.06.02, YS; n Semistaffella minuscilaria, subaxial section, 18G13.05, GS; o Semistaffella primitiva, axial section, 15Y13.03, YS; p Semistaffella variabilis, subaxial section, 15Y13.08.01, YS; q Varistaffella eostaffellaeformis, axial section, 18G15.03, GS; r Varistaffella ziganica, subaxial section, 15Y10.04.02, YS; s Pseudostaffella uralica, axial section, 15Y12.06, YS; t Eostaffella mirifica, axial section, 15Y15.07, YS; u–v Pseudostaffella antiqua, u axial section, 18G18.04, GS, v axial section, 18G18.08, GS; w Pseudostaffella composita, axial section, 18G18.03, GS; x Pseudostaffella compressa, axial section, 18G17.04, GS; y Pseudostaffella grandis, axial section, 18G18.10, GS; z Pseudostaffella cf. proozawai, axial section, 15Y15.03, YS; aa-ab Pseudostaffella sofronizkyi, aa axial section, 15Y14.06, YS, ab subaxial, 18G18.09, GS; ac Schubertella obscura mosquensis, subaxial section, 15Y14.04, YS. Key: YS: Yassıpınar section, GS: Gölbelen section. The scale bar is 125 µm for (a–m) and 250 µm for (n–ac)
Reference section
Yassıpınar section, where it is 16 m thick.
Comparison
Plectostaffella jakhensis was described for the first time by Reitlinger (1971) in the lower Bashkirian successions of the Donets Basin (Ukraine). Later, it was found in coeval successions of the southern Urals (Russia) (Groves 1988; Kulagina and Sinitsyna 1997, 2003; Kulagina et al. 2000, 2001), North Ellesmere Island (Canada) (Groves et al. 1994), Iran (Leven et al. 2006; Leven and Gorgij 2011), Pamir Mountains (Tajikistan) (Leven 2012), Hadim Nappe (Turkey) (Atakul-Özdemir 2006), and North Caspian (Gibshman and Akhmetshina 1991).
One of the characteristic species of this zone is Plectostaffella bogdanovkensis, which was originally described by Reitlinger (1980) from the lower Bashkirian (Bogdanovskian) successions. Later, it was determined in the Syuranian successions of the southern Urals (Russia) (Kulagina and Sinitsyna 1997; Kulagina et al. 2000, 2001, 2013) and Iran (Leven and Gorgij 2011), as well as in Akavasian strata of the Hadim Nappe (Atakul-Özdemir 2006; Dzhenchuraeva and Okuyucu 2007), and in Askynbashian successions of the Pamir Mountains (Tajikistan) (Leven 2012).
Another significant species is Plectostaffella varvariensis. It has a wide geographical distribution and was described for the first time from lower Bashkirian (Syuranian) successions of the Donbas, Ukraine (Brazhnikova and Potievskaya 1948). Subsequent studies detected this species in the southern Urals strata, from the lower Bashkirian (Syuranian) (Kulagina and Sinitsyna 1997; Kulagina et al. 2000, 2001) up to the Askynbashian (Kulagina and Sinitsyna 2003). Plectostaffella varvariensis is also known from the Syuranian–Askynbashian successions of Iran, Pamir Mountains (Tajikistan) and Hadim Nappe (Turkey) (Leven et al. 2006; Dzhenchuraeva and Okuyucu 2007; Atakul-Özdemir et al. 2011; Leven and Gorgij 2011; Leven 2012).
The rest of the fusulinid species collected from this interval were also originally described from the upper Serpukhovian–lower Bashkirian (Rauzer-Chernousova et al. 1936, 1951; Zeller 1953; Rauzer-Chernousova 1948; Reitlinger 1961; Rumyantseva 1970; Grozdilova et al. 1975).
The Plectostaffella jakhensis-Plectostaffella bogdanovkensis First Occurrence Zone correlates with the lower Bashkirian (Syuranian) successions of the Russian Platform and South Urals.
Pseudostaffella antiqua-Pseudostaffella sofronizkyi First Occurrence Zone
Description
This zone is determined between the FOD of Pseudostaffella antiqua (Dutkevich, 1934b) and Pseudostaffella sofronizkyi Safonova in Rauzer-Chernousova et al., 1951, and the FOD of Staffellaeformes staffellaeformis (Kireeva in Rauzer-Chernousova et al., 1951). This stratal interval, in which Pseudostaffella Thompson, 1942 species are dominant, have yielded Eostaffella mirifica Brazhnikova, 1967, Pseudostaffella antiqua (Dutkevich, 1934), P. composita Grozdilova and Lebedeva, 1950, P. compressa (Rauzer-Chernousova, 1938), P. grandis (Shlykova in Grozdilova & Lebedeva, 1950), P. cf. proozawai Kireeva in Rauzer-Chernousova et al., 1951, P. sofronizkyi and Schubertella obscura mosquensis Rauzer-Chernousova in Rauzer-Chernousova et al., 1951. Based on its fusulinid assemblages, the Pseudostaffella antiqua-Pseudostaffella sofronizkyi First Occurrence Zone is correlated with the Akavasian of the early Bashkirian in Russia (Figs. 3, 4, 6, 7t–ac).
Reference section
Gölbelen section. Here, it reaches a thickness of 5 m.
Comparison
The most characteristic species of this interval, Pseudostaffella antiqua, was firstly described by Dutkevich (1934b) from lower Moscovian (C21) successions of the Urals. Later, this taxon was identified in strata of Bashkirian to early Moscovian age from different areas of the world, such as the Russian Platform (Rauzer-Chernousova et al. 1951), the Kolva-Vishera Region, Russia (Grozdilova and Lebedeva 1954), the Cantabrian Mountains, Spain (Ginkel 1965) and the Kitakami Mountains, Japan (Kobayashi 1973).
The other fossil marker of this zone, Pseudostaffella sofronizkyi, was described for the first time (Safonova in Rauzer-Chernousova et al. 1951) from the lower Bashkirian–lower Moscovian successions of the Russian Platform. Later, it was determined from the lower Bashkirian successions of the Alai Mountains (Turkmenistan) and southern Urals (Russia) (Dzhenchuraeva 1979; Kulagina et al. 2001; Kulagina 2003). Other zonal fossils of this interval, Eostaffella mirifica, Pseudostaffella composita, P. grandis, P. compressa and P. cf. proozawai, were originally described from coeval successions of the Donets Basin (Ukraine), Trans-Volga Mountains, southern Urals (Russia) and Russian Platform (Rauzer-Chernousova 1938; Shlykova 1948; Grozdilova and Lebedeva 1950; Rauzer-Chernousova et al. 1951; Brazhnikova et al. 1967).
The composition of the fusulinid assemblages present in the Pseudostaffella antiqua-Pseudostaffella sofronizkyi First Occurrence Zone is similar to that of the lower Bashkirian (Akavasian) assemblages observed in successions of Iran, Pamir, Russian Platform, and southern Urals (Kulagina and Sinitsyna 1997, 2003; Kulagina et al. 2000, 2001; Kulagina 2003; Leven et al. 2006; Leven and Gorgij 2011; Leven 2012).
Staffellaeformes staffellaeformis-Staffellaeformes parva parva First Occurrence Zone
Description
This zone is established between the FOD of Staffellaeformes staffellaeformis (Kireeva in Rauzer-Chernousova et al., 1951) and Staffellaeformes parva parva (Lee and Chen in Lee et al., 1930) at its base, and the FOD of Tikhonovichiella tikhonovichi (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951) at the top. According to the recovered microfauna, which includes Eostaffella acuta Grozdilova and Lebedeva, 1950, Ozawainella pararhomboidalis Manukalova, 1950, Pseudostaffella aff. paracompressa Safonova in Rauzer-Chernousova et al., 1951, Schubertella ex gr. multiforme Villa in Villa and Merino-Tomé, 2016, Depratina convoluta (Lee and Chen in Lee et al., 1930), Depratina prisca prisca (Deprat, 1912), D. prisca sphaeroidea (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), Staffellaeformes bona (Grozdilova and Lebedeva, 1954), S. parva parva, S. parva robusta (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., 1936) and S. staffellaeformis, this zone correlates with the Askynbashian of the early Bashkirian in Russia (Figs. 3, 4, 6, 8).

Thin–section photomicrographs of the fusulinid assemblages of the Staffellaeformes staffellaeformis-Staffellaeformes parva parva Zone (a–m) and Tikhonovichiella tikhonovichi-Verella spicata Zone (n–aa). a Eostaffella acuta, axial section, 15Y16A.29.02, YS; b Ozawainella pararhomboidalis, axial section, 18G20.04, GS; c Pseudostaffella aff. paracompressa, axial section, 15Y16A.08.01, YS; d Schubertella ex gr. multiforme, axial section, 15Y16A.07, YS; e Depratina convoluta, axial section, 15Y16B.11, YS; f Depratina prisca prisca, axial section, 18G27.08.03, GS; g Depratina prisca sphaeroidea, axial section, 15Y16A.12, YS; h Staffellaeformes bona, axial section, 15Y16A.19.02, YS; i–j Staffellaeformes parva parva i axial section, 15Y16A.05, YS, j axial section, 18G19.04.01, GS; k Staffellaeformes parva robusta, axial section, 15Y16A.06, YS; l–m Staffellaeformes staffellaeformis l subaxial section, 18G19.08.01, GS, m axial section, 18G23.07, GS; n Eostaffella kashirica, axial section 18G25.04.02, GS; o Pseudostaffella ex gr. gorskyii, axial section, 18G23.10, GS; p Schubertella obscura obscura, axial section, 15Y22.03.02, YS; q Profusulinella rhomboides, axial section, 15Y19.06, YS; r Solovievaia ex gr. nuratavensis, axial section, 18G26.07, GS; s Depratina beppensis, axial section, 15Y18.24, YS; t Depratina prisca timanica, axial section, 15Y18.11, YS; u Depratina turani, axial section, 15Y17.03, YS; v Aljutovella cf. pseudoaljutovica, axial section, 18G26.05, GS; w Tikhonovichiella praetikhonovichi, axial section, 15Y18.05, YS; x–y Tikhonovichiella tikhonovichi x axial section, 15Y18.09, YS, y subaxial section, 18G26.10, GS; z Verella normalis, axial section, 15Y17.10, YS; aa Verella spicata, axial section, 18G25.05, GS. Key: YS: Yassıpınar section, GS: Gölbelen section. The scale bar is 125 µm for a, n, 250 µm for (b–d), (o–p), 500 µm for (e–m), (q–y) and 1 mm for (z–aa)
Reference section
Gölbelen section, where it is 8 m thick.
Comparison
Staffellaeformes staffellaeformis and Staffellaeformes parva parva are the most significant markers of this zone. The former was described originally by Kireeva (in Rauzer-Chernousova et al. 1951) from the lower Moscovian (Vereian–Kashirian) successions of the Russian Platform; it is also known from the upper Bashkirian to the lower Moscovian of the Alai Mountains, Turkmenistan (Dzhenchuraeva 1979) and the Cantabrian Mountains, Spain (Villa 1995). The restriction of Staffellaeformes staffellaeformis in the Hadim Nappe to the upper Bashkirian (Askynbashian–Arkhangelskian) successions is similar to that in regions such as southern Urals (Russia), Iran, Pamir Mountains (Tajikistan) (Kulagina et al. 2001; Kulagina 2003, 2008, 2009; Dzhenchuraeva and Okuyucu 2007; Leven and Gorgij 2011; Leven 2012). Staffellaeformes parva parva was described for first time by Lee and Chen (in Lee et al. 1930) from the Bashkirian succession of south China. Later, this species has also been determined from the Bashkirian (Askynbashian–Arkhangelskian) successions of South Urals (Russia), Iran and Pamir Mountains (Tajikistan) (Kulagina et al. 2001; Kulagina 2008, 2009; Leven and Gorgij 2011; Leven 2012). Staffellaeformes parva parva ranges from the upper Bashkirian to the lower Moscovian, in the Russian Platform, Kolva-Vishera (Russia), Cantabrian Mountains (Spain), Alai (Turkmenistan), Iran and Hadim Nappe (Turkey) (Rauzer-Chernousova et al. 1951; Grozdilova and Lebedeva 1954; Dzhenchuraeva 1979; Ginkel 1987; Villa 1995; Dzhenchuraeva and Okuyucu 2007).
According to the fusulinid assemblages, the Pseudostaffella antiqua-Pseudostaffella sofronizkyi First Occurrence Zone can be approximately correlated with the Bashkirian (Askynbashian) successions of the southern Urals and Iran.
Tikhonovichiella tikhonovichi-Verella spicata First Occurrence Zone
Description
The base of this zone is marked by the FOD of Tikhonovichiella tikhonovichi (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951) and Verella spicata Dalmatskaya, 1951, whereas its top is indicated by the FOD of Aljutovella aljutovica (Rauzer-Chernousova, 1938). The microfauna of this zone includes Eostaffella kashirica Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, Pseudostaffella ex gr. gorskyii Dutkevich in in Grozdilova & Lebedeva, 1950, Schubertella obscura obscura Lee and Chen in Lee et al., 1930, Profusulinella pararhomboides Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., 1936, P. rhomboides (Lee and Chen in Lee et al., 1930), Solovievaia ex gr. nuratavensis (Solovieva, 1977), Depratina beppensis (Toriyama, 1958), D. prisca timanica (Kireeva in Rauzer-Chernousova et al., 1951), D. turani Akbaş in Akbaş and Okuyucu, 2021, Aljutovella cf. pseudoaljutovica Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, Tikhonovichiella praetikhonovichi Akbaş in Akbaş and Okuyucu, 2021, T. tikhonovichi, Verella normalis Rumyantseva, 1962 and V. spicata. The Tikhonovichiella tikhonovichi-Verella spicata First Occurrence Zone corresponds to the upper Bashkirian (Arkhangelskian) (Figs. 3, 4, 6, 8).
Reference section
Yassıpınar and Gölbelen sections. In the Yassıpınar section it represents 5 m strata, in the Gölbelen section 12 m.
Comparison
Tikhonovichiella tikhonovichi is one of the important index taxa occurring around the Bashkirian–Moscovian boundary. It was originally described by Rauzer-Chernousova (in Rauzer-Chernousova et al. 1951) from the Vereian Regional Substage of the lower Moscovian of the Russian Platform. Later, its occurrence was observed in the upper Bashkirian (Arkhangelskian)–lower Moscovian (Vereian) successions of the Urals (Russia), Pamir Mountains (Tajikistan) and Hadim Nappe (Turkey) (Grozdilova et al. 1975; Kulagina et al. 2001; Kulagina 2003, 2008; Dzhenchuraeva and Okuyucu 2007; Leven 2012).
Verella spicata is another characteristic taxon for establishing the base of this zone. It was described for the first time by Dalmatskaya (1951) from the base of the Moscovian stage of the Russian Platform. Later, it was identified in the upper Bashkirian successions of the southern Gissar and Zerafshan (Tajikistan), Alai (Turkmenistan), Cantabrian Mountains (Spain), Pamir Mountains (Tajikistan) and Hadim Nappe (Bensh 1969; Saltovskaya 1974; Dzhenchuraeva 1979; Villa 1995; Dzhenchuraeva and Okuyucu 2007; Leven 2012).
Considering the defined fusulinid assemblages and their stratigraphic ranges, the Tikhonovichiella tikhonovichi-Verella spicata First Occurrence Zone can be correlated with the upper Bashkirian (Arkhangelskian) successions of the Russian Platform and southern Urals.
Aljutovella aljutovica-Solovievaia ovata ovata First Occurrence Zone
Description
This zone is established from the FOD of Aljutovella aljutovica (Rauzer-Chernousova, 1938) and Solovievaia ovata ovata (Rauzer-Chernousova, 1938) at its base, to the FOD of Eofusulina triangula (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., 1936) at the top. The Aljutovella aljutovica-Solovievaia ovata ovata First Occurrence Zone shows a very abundant and diverse content of Aljutovella Rauzer-Chernousova, 1951 species, comprising Aljutovella aljutovica, A. arrisionis arrisionis (Leontovich in Rauzer-Chernousova et al., 1951), A. arrisionis molotovensis Safonova in Rauzer-Chernousova et al. 1951, A. artificialis Leontovich in Rauzer-Chernousova et al., 1951, A. cf. cafirniganica Bensh, 1969, A. conspecta Leontovich in Rauzer-Chernousova et al., 1951, A. lepida novoburasiensis Leontovich in Rauzer-Chernousova et al., 1951, A. postaljutovica Safonova in Rauzer-Chernousova et al., 1951, A. splendida Leontovich in Rauzer-Chernousova et al., 1951, A. cf. stocklini Davydov and Gorgij in Leven et al., 2006, A. subaljutovica Safonova in Rauzer-Chernousova et al., 1951, A. tumida Bensh, 1969, A. skelnevatica (Putrya and Leontovich, 1948) and A. priscoidea (Rauzer-Chernousova, 1938). Other genera present in this zone are represented by the following species: Profusulinella pseudorhomboides Putrya and Leontovich, 1948, Solovievaia ovata ovata, S. ovata nytvica (Safonova in Rauzer-Chernousova et al., 1951), S. subovata (Safonova in Rauzer-Chernousova et al., 1951), Depratina chernovi (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), D. paratimanica (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), D. posadai (Villa, 1995), D. sitteri (van Ginkel, 1987), and Staffellaeformes tashliensis (Lebedeva in Grozdilova et al., 1975). This zone is early Moscovian (Vereian) in age based on fusulinid assemblages and the presence of Aljutovella aljutovica index taxon (Figs. 3, 4, 6, 9a–q).

Thin–section photomicrographs of the fusulinid assemblages of the Aljutovella aljutovica-Solovievaia ovata ovata Zone (a-q) and Aljutovella priscoidea-Eofusulina triangula Zone (r-s). a-b Solovievaia ovata ovata a axial section, 15Y21.23, YS, b axial section, 18G28.21, GS; c Solovievaia subovata, axial section, 15Y21.34, YS; d Depratina chernovi, axial section, 15Y23.06, YS; e Depratina paratimanica, subaxial section, 15Y21.16.02, YS; f Depratina posadai, axial section, 15Y21.37, YS; g Depratina sitteri, axial section, 15Y21.22, YS; h Staffellaeformes tashliensis, axial section, 15Y21.04.02, YS; i-j Aljutovella aljutovica, i subaxial section, 15Y21.16.01, YS, j axial section, 18G27.03, GS; k Aljutovella arrisionis arrisionis, axial section, 18G28.07.02, GS; l Aljutovella conspecta, axial section, 18G28.23, GS; m Aljutovella lepida novoburasiensis, axial section, 18G29.15, GS; n Aljutovella postaljutovica, axial section, 18G29.04, GS; o Aljutovella skelnevatica, axial section, 18G27.11, GS; p Aljutovella tumida, axial section, 18G27.21, GS; q Neostaffella cf. formosa, axial section, 18G34.03, GS; r Neostaffella larionovae polasnensis, axial section, 18G34.07.02, GS; s Neostaffella nibelensis, axial section, 15Y31.03, YS. Key: YS: Yassıpınar section, GS: Gölbelen section. The scale bar is 250 µm for (r–s) and 500 µm for (a–q)
Reference section
Yassıpınar and Gölbelen sections. In both sections, its thickness is 4 m.
Comparison
The most significant marker for the base of the Moscovian Stage is Aljutovella aljutovica, which was described for the first time by Rauzer-Chernousova (1938) from the lower Moscovian beds of the Samara region and the Moscow Basin (Russia). Later, this species was determined in other lower Moscovian (Vereian–lower Kashirian?) localities of the Russian Platform (Rauzer-Chernousova et al. 1951) and in the C2K suite of the Donbas (Ukraine) (Putrya 1956). Its presence has also been established in the lower Moscovian (Vereian) successions of the Urals (Russia), Alai (Turkmenistan), Pamir Mountains (Tajikistan) and Hadim Nappe (Grozdilova et al. 1975; Dzhenchuraeva 1979; Leven 1998; Kulagina 2003, 2008; Dzhenchuraeva and Okuyucu 2007; Kobayashi 2011).
Solovievaia ovata ovata, one of the characteristic species of the Aljutovella aljutovica-Solovievaia ovata ovata First Occurrence Zone, was originally described by Rauzer-Chernousova (1938) from the Kashirian Substage of the lower Moscovian of the Russian Platform; later, it was noticed that it ranges from the Vereian to the lower Kashirian (Rauzer-Chernousova et al. 1951). This species also has been determined in the lower Moscovian (Vereian–Kashirian) successions of the southern Gissar, Tajikistan (Bensh 1969), Urals, Russia (Grozdilova et al. 1975), Alai, Turkmenistan (Dzhenchuraeva 1979) and Cantabrian Mountains, Spain (Villa 1995).
Most of the Aljutovella species occurring in this zone are widely distributed in the lower Moscovian (Vereian–Kashirian) successions of the Russian Platform (Rauzer-Chernousova et al. 1951). Aljutovella cafirniganica and A. tumida are known from southern Gissar, Tajikistan (Bensh 1969), and A. stocklini was originally described from eastern and central Iran (Leven et al. 2006). All of them are typical lower Moscovian forms.
The rest of the fusulinids of this zone belong to species that are common in coeval successions of the Tethys Realm (Rauzer-Chernousova 1938; Putrya and Leontovich 1948; Rauzer-Chernousova et al. 1951; Grozdilova et al. 1975; Ginkel 1987; Villa 1995).
The occurrence in the Aljutovella aljutovica-Solovievaia ovata ovata First Occurrence Zone of so diverse and abundant Aljutovella species of early Moscovian age correlates this zone with the lower Moscovian (Vereian) successions of the Russian Platform.
Aljutovella priscoidea-Eofusulina triangula First Occurrence Zone
Description
This zone is characterized by the FOD of Aljutovella priscoidea (Rauzer-Chernousova, 1938) and Eofusulina triangula (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., 1936) at its base, and the FOD of the Fusulinella vozhgalensis devexa Rauzer-Chernousova in Rauzer-Chernousova et al., 1951 at the top. The fusulinids collected from this interval consist of Ozawainella crassiformis Putrya, 1956, O. leei (Putrya, 1939), Neostaffella cf. formosa (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), N. larionovae polasnensis (Rauzer-Chernousova and Safonova in Rauzer-Chernousova et al., 1951), N. nibelensis (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), N. sphaeroidea sphaeroidea (Ehrenberg, 1842), N. sphaeroidea cuboides (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), N. subquadrata (Grozdilova and Lebedeva, 1950), Profusulinella parafittsi Rauzer-Chernousova and Safonova in Rauzer-Chernousova et al., 1951, P. rhombiformis nibelensis Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, Solovievaia arta (Leontovich in Rauzer-Chernousova et al., 1951), S. simplex (Safonova in Rauzer-Chernousova et al., 1951), S. syzranica (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), Depratina dubia (Villa, 1995), Taitzehoella librovitchi (Dutkevich, 1934a), T. mutabilis (Safonova in Rauzer-Chernousova et al., 1951), Aljutovella cybaea Leontovich in Rauzer-Chernousova et al., 1951), A. isvarica Putrya, 1956, A. priscoidea, Eofusulina binominata Putrya, 1956, E. triangula, Fusulinella sp. A, F. ex gr. schubertellinoides Putrya, 1938, and F. praecolaniae Safonova in Rauzer-Chernousova et al., 1951. This zone corresponds to the Kashirian of the lower Moscovian in Russia based on the occurrence of the index zone fossils (Figs. 3, 4, 5, 6, 9r–s, 10).

Thin–section photomicrographs of the fusulinid assemblages of the Aljutovella priscoidea-Eofusulina triangula Zone (continue). a Neostaffella sphaeroidea sphaeroidea, subaxial section, 15Y29.06, YS; b Neostaffella sphaeroidea cuboides, subaxial section, 18G32.09, GS; c Neostaffella subquadrata, axial section, 18G32.10, GS; d Solovievaia arta, axial section, 15Y28.03, YS; e Solovievaia simplex, axial section, 18G33.04, GS; f Solovievaia syzranica, axial section, 18G33.12, GS; g Depratina dubia, axial section, 15Y30.10, YS; h Taitzehoella librovitchi, subaxial section, 16B36.03, BS; i Taitzehoella mutabilis, subaxial section, 18G32.20, GS; j–k Aljutovella priscoidea, j axial section, 15Y24.19, YS, k axial section, 16B35.04, BS; l Eofusulina binominata, subaxial section, 18G31.06, GS; m–n Eofusulina triangula, m axial section, 15Y27.04, YS, n axial section, 15Y25.06, YS; o Fusulinella ex gr. schubertellinoides, axial section, 16B33.03, BS; p Fusulinella praecolaniae, axial section, 16B38.07, BS. Key: YS: Yassıpınar section, GS: Gölbelen section, BS: Bademli section. The scale bar is 250 µm for (a–c), 500 µm for (d–k) and 1 mm for (l–p)
Reference section
Yassıpınar, Bademli and Gölbelen sections. Its thickness is 9.5 m in the Yassıpınar section, 8 m in the Bademli section, and 16 m in the Gölbelen section.
Comparison
Aljutovella priscoidea, one important taxon within the described zone, was originally established by Rauzer-Chernousova (1938) in the lower Moscovian (Kashirian) successions of the Samara Region (Russia), and later it was also described from the equivalent successions of the Russian Platform by Rauzer-Chernousova and Safonova (in Rauzer-Chernousova et al. 1951). Aljutovella priscoidea has also been determined in the Kashirian of the lower Moscovian of the southern Gissar (Tajikistan), Urals (Russia), Alai (Turkmenistan), Cantabrian Mountains (Spain) and Iran (Bensh 1969; Grozdilova et al. 1975; Dzhenchuraeva 1979; Villa 1995; Leven et al. 2006; Leven and Gorgij 2011).
Eofusulina triangula, another index taxon of this zone, was described for the first time from the lower Moscovian (Vereian–Kashirian) of the Pechora Region, Russia (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al. 1936). Later, it was determined and widely known in the lower Moscovian successions of the Russian Platform (Safanova in Rauzer-Chernousova et al. 1951), Donets Basin, Ukraine (Putrya 1956; Davydov 2009), Uzbekistan (Rumyantseva 1962), Cantabrian Mountains, Spain (Ginkel 1965; Villa 1995), Zerafshan-Gissar, Tajikistan (Saltovskaya 1974), Urals, Russia (Grozdilova et al. 1975), and Alai, Turkmenistan (Dzhenchuraeva 1979).
In addition to the index taxa, many species that are present in this zone were also originally described by Rauzer-Chernousova et al. (1951) from the lower Moscovian (Vereian–Kashirian) successions of the Russian Platform, including Neostaffella cf. formosa, N. larionovae polasnensis, N. nibelensis, N. sphaeroidea cuboides, Profusulinella parafittsi, P. rhombiformis nibelensis, Solovievaia arta, S. simplex, S. syzranica, Taitzehoella mutabilis, Aljutovella cybaea, and Fusulinella praecolaniae. Other species, as Ozawainella crassiformis, O. leei, Aljutovella isvarica, Eofusulina binominata and Fusulinella ex gr. schubertellinoides, were also originally described from the lower Moscovian successions of the Donets Basin in Ukraine (Putrya 1938, 1939, 1956) while Taitzehoella librovitchi and Neostaffella subquadrata were originally designated in the Urals (Dutkevich 1934a; Grozdilova and Lebedeva 1950).
Based on the above-mentioned fusulinid content, the Aljutovella priscoidea-Eofusulina triangula First Occurrence Zone is approximately correlated with the Kashirian Substage of the Russian Platform.
Fusulinella vozhgalensis devexa-Beedeina schellwieni First Occurrence Zone
Description
This zone, which comprises a rich fusulinid assemblage mainly belonging to the Fusulinella Möller, 1877 and Beedeina Galloway, 1933 genera, is determined between the FOD of Fusulinella vozhgalensis devexa Rauzer-Chernousova in Chernousova et al., 1951 and Beedeina schellwieni (Staff in Schellwien and Staff, 1912) at its base, and the FOD of Fusulinella bocki bocki Möller, 1878 at the top. The abundant and diverse fusulinid assemblages of this zone includes Ozawainella angulata (Colani, 1924), O. kumpani Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, O. kurachovensis Manukalova, 1950, O. mosquensis Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, O. tingi (Lee, 1937), O. quadrata Kireeva, 1949, Neostaffella ozawai compacta (Manukalova, 1950), N. umbilicata (Putrya and Leontovich, 1948), Taitzehoella praelibrovichi (Safonova in Rauzer-Chernousova et al., 1951), Aljutovella pojarkovie Dzhenchuraeva, 1979, Beedeina elegans (Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., 1940), B. paradistenta (Safonova in Rauzer-Chernousova et al., 1951), B. pseudoelegans pseudoelegans (Chernova in Rauzer-Chernousova et al., 1951), B. pseudoelegans keltmensis (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), B. samarica (Rauzer-Chernousova and Belyaev, 1940), B. schellwieni, B. timanica (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951), Fusulinella cf. eopulchra Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, and F. vozhgalensis devexa. According to this microfauna, this zone corresponds to the upper Moscovian (Podolskian) (Figs. 3, 4, 5, 6, 11).

Thin–section photomicrographs of the fusulinid assemblages of the Fusulinella vozhgalensis devexa-Beedeina schellwieni Zone. a Ozawainella angulata, subaxial section, 15Y36.18, YS; b Ozawainella kumpani, axial section, 18G41.12, GS; c Ozawainella kurachovensis, axial section, 16B39.04.02, BS; d Ozawainella mosquensis, axial section, 18G41.16, GS; e Ozawainella quadrata, axial section, 15Y37.05, YS; f Ozawainella tingi, subaxial section, 18G40.08, GS; g Neostaffella ozawai compacta, axial section, 15Y34.03, YS; h Aljutovella pojarkovie, axial section, 15Y33.08, YS; i Beedeina paradistenta, axial section, 16B41.16, BS; j Beedeina elegans, subaxial section, 16B40B.08, BS; k Beedeina pseudoelegans pseudoelegans, axial section, 15Y36.09, YS; l Beedeina samarica, axial section, 16B40B.05, BS; m–n Beedeina schellwieni, m axial section, 16B40A.05, BS, n axial section, 18G41.15, GS; o Fusulinella cf. eopulchra, axial section, 15Y37.03, YS; p–q Fusulinella vozhgalensis devexa, p subaxial section, 16B39.07, BS, q axial section, 18G41.11, GS. Key: YS: Yassıpınar section, GS: Gölbelen section, BS: Bademli section. The scale bar is 250 µm for (a–g) and 1 mm for (h–q)
Reference section
Bademli and Gölbelen sections. It is 6 m thick in the Bademli section, and 20.5 m in the Gölbelen section.
Comparison
Fusulinella vozhgalensis devexa, one of the characteristic species of this zone, was described for the first time by Rauzer-Chernousova (in Rauzer-Chernousova et al. 1951) from the Podolskian strata (upper Moscovian) of the Russian Platform, and later it was determined in coeval successions of the northern Timan by Lebedeva (1966). Beedeina schellwieni, one of the zone fossils of this zone, was originally described from the Donets Basin (Ukraine) by Staff (in Schellwien and Staff 1912). This species is widely spread in upper Moscovian strata, more rarely in the Kashirian, of the Russian Platform, Donets Basin (Ukraine), Cantabrian Mountains (Spain), Pamir Mountains (Tajikistan) and Hadim Nappe (Turkey) (Rauzer-Chernousova et al. 1951; Putrya 1956; Ginkel 1965; Villa 1995; Leven 1998; Okuyucu 2002, 2009).
Regarding the Ozawainella species found within this zone, Ozawainella angulata, O. kumpani and O. mosquensis were originally described from the upper Moscovian of the Russian Platform, while O. kurachovensis, O. tingi, and O. quadrata were described from the Donets Basin, Ukraine (Colani 1924; Lee 1937; Kireeva 1949; Manukalova 1950; Rauzer-Chernousova et al. 1951). As to the Neostaffella Miklukho-Maklay, 1959 species, N. ozawai compacta was described originally from the Donets Basin, Ukraine (Manukalova 1950), and N. umbilicata from the Saratov Volga Region, Russia (Putrya and Leontovich 1948), both from Moscovian strata.
In spite of the rare occurrence of the species belonging to Aljutovella in the upper Moscovian strata, one form belonging to this genus, A. pojarkovie, originally described from the Alai Mountains, Turkmenistan (Dzhenchuraeva 1979), was determined in this study.
Several Beedeina and one Fusulinella species collected from this zone are forms well-known from the Russian Platform and other Eurasian areas, e.g. Beedeina elegans, B. samarica, B. paradistenta, B. pseudoelegans keltmensis, B. timanica, and Fusulinella cf. eopulchra. All of them were originally described from the upper Moscovian strata (Podolskian–Myachkovian) of the Russian Platform (Rauzer-Chernousova et al. 1940, 1951).
Based on the composition of the fusulinid assemblages found in the Hadim Nappe, the Fusulinella vozhgalensis devexa-Beedeina schellwieni First Occurrence Zone can be correlated with the upper Moscovian (Podolskian) successions of the Russian Platform.
Fusulinella bocki bocki First Occurrence Zone
Description
Fusulinella species are dominant in this zone, which is established from the FOD of Fusulinella bocki bocki Möller, 1878, at its base, to the FOD of Quasifusulinoides quasifusulinoides (Rauzer-Chernousova in Rauzer-Chernousova et al., 1951) at the top. The diverse microfauna of this zone includes Ozawainella krasnokamski Safonova in Rauzer-Chernousova et al., 1951, O. paratingi Manukalova, 1950, O. pseudorhomboidalis Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, Neostaffella ozawai ozawai (Lee and Chen in Lee et al., 1930), Beedeina cf. nytvica callosa (Safonova in Rauzer-Chernousova et al., 1951), Fusulinella bocki bocki, F. bocki pauciseptata Rauzer-Chernousova and Belyaev in Rauzer-Chernousova et al., 1936, F. cf. bocki timanica Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, F. cf. colaniae colaniae Lee and Chen in Lee et al., 1930, F. colaniae meridionalis Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, F. helenae Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, F. paracolaniae Safonova in Rauzer-Chernousova et al., 1951, F. praebocki Rauzer-Chernousova in Rauzer-Chernousova et al., 1951, F. pseudobocki Lee and Chen in Lee et al., 1930, Fusulinella ? sp. B, and Fusulinella ? sp. C. According to the composition of the fusulinid assemblages collected from these strata, the Fusulinella bocki bocki First Occurrence Zone corresponds to the upper Moscovian (Myachkovian) (Figs. 3, 4, 5, 6, 12).

Thin–section photomicrographs of the fusulinid assemblages of the Fusulinella bocki bocki Zone. a Ozawainella krasnokamski, axial section, 15Y39.16, YS; b Ozawainella paratingi, axial section, 15Y39.15, YS; c Ozawainella pseudorhomboidalis, subaxial section, 16B42.20, BS; d Neostaffella ozawai ozawai, axial section, 18G49.03, GS; e Beedeina cf. nytvica callosa, axial section, 18G55.03, GS; f–g Fusulinella bocki bocki, f axial section, 16B42.03, BS, g axial section, 18G48.06, GS; h Fusulinella bocki pauciseptata, axial section, 16B43.22, BS; i Fusulinella cf. bocki timanica, axial section, 15Y39.05, YS; j Fusulinella cf. colaniae colaniae, axial section, 18G56.04, GS; k Fusulinella colaniae meridionalis, axial section, 15Y39.11, YS; l Fusulinella paracolaniae, axial section, 16B43.05, BS; m Fusulinella praebocki, axial section, 16B42.18.01, BS; n Fusulinella pseudobocki, axial section, 18G56.05, GS. Key: YS: Yassıpınar section, GS: Gölbelen section, BS: Bademli section. The scale bar is 250 µm for (a–d) and 1 mm for (e–n)
Reference section
Bademli and Gölbelen sections. Its thickness is 8 m in the Bademli section, and 19 m in the Gölbelen section.
Comparison
Fusulinella bocki bocki was described for the first time by Möller (1878) from the Upper Coal Beds in Novoarks Region, Kresty (Russia). Later, it was determined in the upper Moscovian successions of the Russian Platform, Cantabrian Mountains (Spain), northern Timan, Urals, Pamir Mountains and Hadim Nappe (Rauzer-Chernousova et al. 1951; Ginkel 1965; Lebedeva 1966; Grozdilova et al. 1975; Leven 1998; Okuyucu 2013).
Beedeina and Fusulinella species are dominant in these strata, from which we have identified Beedeina cf. nytvica callosa, Fusulinella cf. bocki timanica, F. colaniae meridionalis, F. helenae, F. paracolaniae, F. praebocki (originally described from the Russian Platform; Rauzer-Chernousova et al. 1951), F. bocki pauciseptata (originally described from the Pechora Region; Rauzer-Chernousova et al. 1936), and F. cf. colaniae colaniae, F. pseudobocki (described from southern China; Lee et al. 1930).
With respect to other taxa, Ozawainella krasnokamski and O. pseudorhomboidalis were described by Rauzer-Chernousova et al. (1951) from coeval successions from the Russian Platform, while O. paratingi was described from the upper Moscovian of the Donets Basin (Manukalova 1950), and Neostaffella ozawai ozawai was described from southern China (Lee et al. 1930).
Based on the fusulinids yielded by these strata, the Fusulinella bocki bocki First Occurrence Zone can be correlated with the upper Moscovian (Myachkovian) of the Moscow Basin.
Correlation
Most of the fusulinids identified in the Hadim Nappe sections are species common in the Bashkirian to Moscovian successions of the Russian Platform, southern Urals, Donets Basin, Cantabrian Mountains (Spain), Iran, Pamir Mountains (Tajikistan), southern China, and Tien-Shan (Kagarmanov and Donakova 1990; Villa 1995; Kulagina et al. 2001; Orlov-Labkovsky et al. 2007; Davydov et al. 2010). In particular, the Bashkirian fusulinid zones established in the Hadim Nappe sections show very close similarity to the southern Urals biozonation (Kulagina et al. 2001) while the Moscovian fusulinid zones are close to the fusulinid biozones described by Kagarmanov and Donakova (1990) for the Russian Platform (Fig. 13).

Correlation of the fusulinid zones established in this study with the fusulinid zones of the adjacent regions
The Syuranian and Akavasian in the southern Urals are subdivided into several fusulinid zones (Kulagina et al. 2001). While there is a similar Syuranian and Akavasian fusulinid fauna in the Hadim Nappe sections, these intervals are represented by a single fusulinid zone due to the diachronic occurrences of some zonal fusulinid species in the Hadim Nappe sections (Fig. 13). The uppermost fusulinid zones of the Bashkirian in the Russian Platform which correspond to the Arkhangelskian, are congruent to a single zone in the Hadim Nappe sections related to some faunal differences. The base of Moscovian (Vereian) across the Bashkirian-Moscovian boundary is marked by the occurrence of Aljutovella aljutovica in Russian Platform, Tien-Shan and Donets Basin (Kagarmanov and Donakova 1990; Kulagina et al. 2001; Orlov-Labkovsky et al. 2007; Davydov et al. 2010), while in the Hadim Nappe sections the first occurrence of Aljutovella aljutovica is in basal Moscovian strata; Solovievaia ovata ovata additionally first occurs and is a good indicator for the base of Moscovian. Although the fusulinid zones described in overlying successions in Moscovian of the Hadim Nappe show some faunal differences and diachronism with adjacent regions in Tethyan Realm, most of the zonal markers are the same and allow a reasonable correlation to these regions.
Conclusion
The fusulinid-based biostratigraphy of the three stratigraphic sections studied in the Hadim Nappe, southern Turkey, has resulted in the definition of eight local biozones ranging from the Bashkirian to Moscovian. In ascending order, they are: (1) Plectostaffella jakhensis-Plectostaffella bogdanovkensis Zone (Bashkirian, correlatable with the Syuranian); (2) Pseudostaffella antiqua-Pseudostaffella sofronizkyi Zone (Bashkirian, correlatable with the Akavasian); (3) Staffellaeformes staffellaeformis-Staffellaeformes parva parva Zone (Bashkirian, correlatable with the Askynbashian); (4) Tikhonovichiella tikhonovichi-Verella spicata Zone (Bashkirian, correlatable with the Arkhangelskian); (5) Aljutovella aljutovica-Solovievaia ovata ovata Zone (Moscovian, correlatable with the Vereian); (6) Aljutovella priscoidea-Eofusulina triangula Zone (Moscovian, correlatable with the Kashirian); (7) Fusulinella vozhgalensis devexa-Beedeina schellwieni Zone (Moscovian, correlatable with the Podolskian); (8) Fusulinella bocki bocki Zone (Moscovian, correlatable with the Myachkovian).
The identified fusulinid zones and their faunal composition in the studied sections show similarities with adjacent areas within the Tethyan Realm (e.g., Moscow Basin, Donets Basin, southern Urals, Cantabrian Mountains, and southern China), but especially to the Russian Platform and the substage/horizons of the Russian regional chronostratigraphic subdivisions. The fusulinid similarity between the Hadim Nappe sections and the Russian Platform during the studied time interval reveals a close connection between these regions and support a same palaeolatitudinal position with a probable faunal exchange.
Data availability
All materials (thin-sections) are reposited in the collection of the MA–(HY15) at the Faculty of Engineering and Natural Sciences, Department of Geological Engineering, Konya Technical University.
Code availability
Not applicable.
References
Altıner, Demir. 1981. Recherches stratigraphiques et micropaléontologiques dans le Taurus Oriental au NW de Pinarbasi (Turquie). Unpublished PhD Thesis, 1–450. Geneve: Universite de Geneve.
Akbaş, M., and C. Okuyucu. 2021. Biostratigraphy and taxonomy of fusulinid Foraminifera across the Upper Mississippian (upper Serpukhovian)–lower Pennsylvanian (Bashkirian) successions from the Hadim Nappe, Central Taurides, southern Turkey. Journal of Paleontology 95(3): 476–496.
Akbaş, M., and C. Okuyucu. 2022. Fusulinid biostratigraphy of the Moscovian—Lower Kasimovian of Hadim Nappe. Central Taurides, southern Turkey. Geodiversitas (in press).
Atakul-Özdemir, A. 2006. Lower–Middle Carboniferous boundary in central Taurides, Turkey (Hadim area): paleontological and sequence stratigraphic approach. Unpublished MSc Thesis, 1–202. Ankara: Middle East Technical University, Graduate School of Natural and Applied Sciences.
Atakul-Özdemir, A., D. Altıner, S. Özkan-Altıner, and İÖ. Yılmaz. 2011. Foraminiferal biostratigraphy and sequence stratigraphy across the mid–Carboniferous boundary in the Central Taurides, Turkey. Facies 57(4): 705–730.
Bensh, F. R. 1969. Stratigraphy and foraminifers of the Carboniferous beds of the southwestern spurs and southern slope of the Gissar Mountains. Uzbekistan, Tashkent: Academy of Sciences of Uzbekistan SSR, Institute of Geology and Geophysics, "Fan" of Uzbekistan. (in Russian).
Blumenthal, M.M. 1947. Beyşehir-Seydişehir Hinterlandındaki Toros Dağlarının Jeolojisi [Geology of the Taurus Mountains in the Beyşehir-Seydişehir Hinterland]. Mineral Research and Exploration Institute Publications Series D 2: 1–242.
Blumenthal, M.M. 1951. Recherches géologiques dans le Taurus occidental dans l’arrière-pays d’Alanya. Mineral Research and Exploration Institute Publications Series D 5: 1–194.
Brazhnikova, N.E., and P.D. Potievskaya. 1948. Results of the foraminiferal study of materials from wells of the western outskirts of Donbass. Institut Geologichnikh Nauk, Zbirnik Prats’ z Paleonyologiy Ta Stratigrafiy 1(2): 76–101. (in Ukrainian).
Brazhnikova, N.E., G.I. Vakarchuk, M.V. Vdovenko, L.B. Vinnichenko, M.A. Karpova, Y.A.I. Kolomiets, P.D. Potievskaya, L.F. Rostovtseva, and G.D. Shevchenko. 1967. Microfaunal reference horizons of Carboniferous and Permian deposits from the Dnieper-Donets basin. Kiev: Naukova Dumka. (in Russian).
Brunn, J.H., J.F. Dumont, P.C. de Graciansky, M. Gutnic, Th. Juteau, J. Marcoux, O. Monod, and A. Poisson. 1971. Outline of the geology of the western Taurids. In Geology and history of Turkey, ed. A.S. Campbell, 225–255. Tripoli: Petroleum Exploration Society of Libya.
Brunn, J.H., I. Argyriadis, J. Marcoux, O. Monod, A. Poisson, and L. Ricou. 1973. Antalya’nin ofiyolit naplarinin orijini lehinde ve aleyhinde kanitlar [Evidence of the origin of the Ophiolite Nappes of Antalya]. Cumhuriyetin 50. Yili Yerbilimleri Kongresi, Mineral Research and Exploration Publications, Ankara, 58–69.
Colani, M. 1924. Nouvelle contribution à l’étude des fusulinidés de l’Extrême-Orient. Mémoires du Service Géologique de l’Indochine 11(1): 9–199.
Dalmatskaya, I.I. 1951. A new genus of fusulinid from the lower part of the Middle Carboniferous deposits of the Russian Platform. Proceedings of the Moscow Society of Naturalists, Section of Geology 1: 194–196. (in Russian).
Davydov, V.I. 2009. Bashkirian–Moscovian transition in Donets Basin: the key for Tethyan-Boreal correlation. In International Field Meeting of the I.U.G.S. Subcommission on Carboniferous Stratigraphy: The Carboniferous type sections in Russia, potential and proposed stratotypes, ed. V. N. Puchov Proceedings of the International Conference, South Urals Session. Ufa-Sibai, 13–18 August, 2009: 188–192. Ufa: Institute of Geology. Bashkirian Academy of Sciences.
Davydov, V.I., J.L. Crowley, M.D. Schmitz, and V.I. Poletaev. 2010. High-precision U-Pb zircon age calibration of the global Carboniferous time scale and Milankovitch band cyclicity in the Donets basin, eastern Ukraine. Geochemistry, Geophysics, Geosystems 11(1): 1–22.
Demirel, S. 2012. Foraminiferal paleontology and sequence stratigraphy in the Upper Viséan–Serpukhovian deposits (Aladağ Unit, Eastern Taurides, Turkey). Unpublished MSc Thesis, 1–360. Ankara: Middle East Technical University, The Graduate School of Natural and Applied Sciences.
Demirel, S., and D. Altıner. 2016. Foraminiferal biostratigraphy and glacioeustatic control on cyclic carbonate microfacies in the Viséan-Serpukhovian boundary beds (Aladağ Unit, Eastern Taurides, Turkey). Facies 62(1): 1–32.
Deprat, J. 1912. Étude géologique du Yun-Nan oriental; III: Étude des Fusulinidés de Chine et d’Indochine et classification des calcaires à Fusulines. Mémoires du Service Géologique de l’Indochine 1(3): 1–76.
Dutkevich, G.A. 1934a. Some new species of Fusulinidae from the Upper and Middle Carboniferous of Verkhne-Chussovskye Gorodki on the Chussovaya River (western slope of the Middle Ural). Vsesoyuzniyi Neftianoi nauchno-issledovatelskyi Geologorazvedochnyi Institute, Trudy Serii A (36): 3–98. (in Russian).
Dutkevich, G.A. 1934b. Stratigraphy of the Middle Carboniferous in the Urals. Vsesoyuzniyi Neftianoi nauchno-issledovatelskyi Geologorazvedochnyi Institute, Trudy Serii A 55: 3–41. (in Russian).
Dzhenchuraeva, A.V. 1979. Stratigraphy and foraminifera of the middle Carboniferous deposits of the northern slopes of Turkestan-Alai, 1–184. Frunze: Academy of Sciences of Kirghizistan SSR. (in Russian).
Dzhenchuraeva, A.V., and C. Okuyucu. 2007. Fusulinid Foraminifera of the Bashkirian-Moscovian boundary in the eastern Taurides, southern Turkey. Journal of Micropalaeontology 26(1): 73–85.
Ehrenberg, C.G. 1842. Mitteilung über die Polythalamen des Bergkalks am Onega-See in Russland. Berichte und Verhandlungen der Königlich Preussischen Akademie der Wissenschaften Berlin 1842: 273–275.
Galloway, J.J. 1933. A manual of Foraminifera. 483 pp. Bloomington, Indiana: Principia Press.
Gibshman, N.B., and L.Z. Akhmetshina. 1991. Micropaleontologic basis for determination of the Mid-Carboniferous boundary in the North Caspian syneclise, USSR. Courier Forschungsinstitut Senckenberg 130: 273–295.
Groves, J.R. 1988. Calcareous foraminifers from the Bashkirian stratotype (Midle Carboniferous, South Urals) and their significance for intercontinental correlations and the evolution of the Fusulinidae. Journal of Palaeontology 62(3): 368–399.
Groves, J.R., R. Lin, W.W. Nassichuk, and S. Pinard. 1994. Middle Carboniferous fusulinacean biostratigraphy, northern Ellesmere Island (Sverdrup Basin, Canadian Arctic Archipelago). Geological Survey of Canada Bulletin 469: 1–60.
Grozdilova, L.P., and N.S. Lebedeva. 1950. Some species of Staffella of Middle Carboniferous deposits of the western slope of the Urals. Microfauna of USSR, Proceedings of the Oil Research Institute Leningrad, new series 50: 5–45. (in Russian).
Grozdilova, L.P., and N.S. Lebedeva. 1954. Lower Carboniferous, and Middle Carboniferous Bashkirian Foraminifera from Kolva-Vishera Region. Microfauna of USSR, Proceedings of the Oil Research Institute Leningrad, new series 7: 4–236. (in Russian).
Grozdilova, L.P., N.S. Lebedeva, O.A. Lipina, N.P. Malakhova, Z.P. Mikhailova, V.A. Chermnykh, M.V. Postoyalko, Z.G. Simonova, Z.A. Sinitsyna, D.L. Krylova, A.K. Grozdilova, L.P. Pozner, and A.A. Sultanaev. 1975. Foraminifera. In Stepanov, D.L., A.K. Krylova, L.P. Grozdilova, V.M. Pozner, and A.A. Sultanaev, eds. 1975. Paleontological Atlas of the Carboniferous deposits from the Urals. Trudy VNIGRI 383: 27-64. Moscow: Ministry of Geology of the USSR. (in Russian).
Gutnic, M, O. Monod, A. Poisson, and F. Dumont. 1979. Géologie des Taurides Occidentales (Turquie). Mémoires de la Société Géologique de France 137: 1–109.
Güvenç, T. 1965. Etude stratigraphique et micropaléontologique du Carbonifère et du Permien des Taurus occidentaux dans l’arrière-pays d’Alanya (Turquie). Unpublished PhD Thesis, 1–273. Paris: Université de Paris.
Güvenç, T. 1977. Stratigraphie du Carbonifère et du Permien de la Nappe de Hadim. 6th Colloquium of the Geology of Aegean Regions, Aegean University, İzmir 263–282.
Kagarmanov, A.Kh., and L.M. Donakova. 1990. Carboniferous System. In Decision of the Interdepartmental Regional Stratigraphic Meeting on Middle and Upper Paleozoic of the Russian Platform, ed. L.M. Donakova, 3–40. Leningrad. (in Russian).
Kireeva, G.D. 1949. Some new species of fusulinids from the Carboniferous limestones of the central regions of Donbass. Geological and Research Work of the Main Directorate of Coal Exploration 6: 25–54. (in Russian).
Kobayashi, F. 1973. Fusulinids of the Nagaiwa formation. Transactions and Proceedings of the Paleontological Society of Japan New Series 92: 200–219.
Kobayashi, F. 2011. Two species of Profusulinella (P. aljutovica and P. ovata), early Moscovian (Pennsylvanian) fusulines from southern Turkey and subdivision of primitive groups of the family Fusulinidae. Rivista Italiana di Paleontologia e Stratigrafia 117(1): 29–37.
Kobayashi, F., and D. Altıner. 2008a. Late Carboniferous and Early Permian fusulinoideans in the central Taurides, Turkey: Biostratigraphy, faunal composition and comparison. The Journal of Foraminiferal Research 38(1): 59–73.
Kobayashi, F., and D. Altıner. 2008b. Fusulinoidean faunas from the upper Carboniferous and lower Permian platform limestone in the Hadim Area, central Taurides, Turkey. Rivista Italiana di Paleontologia e Stratigrafia 114(2): 191–232.
Kulagina, E.I. 2003. Stratigraphy and foraminifers of the Middle Carboniferous of the Belaya River, South Urals. Ufa Scientific Center, Academy of Sciences of Russia, Geological Collection 3: 57–62. (in Russian).
Kulagina, E.I. 2008. Boundary of Bashkirian and Moscovian stages (Middle Carboniferous) in South Urals on fusulinid evolution. Bulletin of the Moscow Society of Naturalists, Department of Geology 83: 33–43.
Kulagina, E.I. 2009. Evolution of the fusulinid Depratina in the Bashkirian-Moscovian interval. Palaeoworld 18(2–3): 94–101.
Kulagina, E.I., and Z.A. Sinitsyna. 1997. Foraminiferal zonation of the Lower Bashkirian in the Askyn section, south Urals, Russia. In Late Paleozoic Foraminifera. Their biostratigraphy, evolution and paleoecology and the Mid–Carboniferous boundary, eds. C.A. Ross, J.R.P. Ross, and P.L.Baccelle. Cushman Foundation for Foraminiferal Research Special Publication 36: 83–88.
Kulagina, E.I., and Z.A. Sinitsyna. 2003. Evolution of the Pseudostaffellidae in the Bashkirian Stage (Middle Carboniferous). Rivista Italiana di Paleontologia e Stratigrafia 109(2): 213–224.
Kulagina, E.I., V.N. Pazukhin, S.V. Nikolaeva, and N.N. Kochetova. 2000. Biozonation of the Syuran horizon of the Bashkirian Stage in the South Urals as indicated by ammonoids, conodonts, foraminifers and ostracodes. Stratigraphy and Geological Correlation 8(2): 137–156.
Kulagina, E.I., V.N. Pazukhin, N.M. Kotschetkova, Z.A. Sinitsyna, and N.N. Kochetova. 2001. Stratotype and reference sections of the Bashkirian Stage of the Carboniferous of the Southern Urals. 1–139. Ufa Scientific Center, Academy of Sciences of Russia: Gilem. (in Russian).
Kulagina, E.L., S. Nikolaeva, V.N. Pazukhin, and N.N. Kochetova. 2013. Biostratigraphy and lithostratigraphy of the Mid-Carboniferous boundary beds in the Muradymovo section (South Urals, Russia). Geological Magazine 151(2): 269–298.
Lebedeva, N.S. 1966. Collection XIV, Foraminifera from the Middle Carboniferous of the North Timan. Microfauna of USSR, Proceedings of the Oil Research Institute Leningrad 250: 176–253. (in Russian).
Lee, J.S. 1937. Foraminifera from the Donetz Basin and their stratigraphical significance. Bulletin of the Geological Society of China 16(1): 57–107.
Lee, J.S., S. Chen, and S. Chu. 1930. The Huanglung Limestone and its fauna. Memoirs National Research Institute of Geology 9: 85–143.
Leven, E.J. 1998. Stratigraphy and fusulinids of the Moscovian Stage (Middle Carboniferous) in the southwestern Darvaz (Pamir). Rivista Italiana di Paleontologia e Stratigrafia 104(1): 3–42.
Leven, E.J. 2012. The Bashkirian stage of southwestern Darvaz, the Pamir Mountains: Stratigraphy and paleotectonics. Stratigraphy and Geological Correlation 20(3): 240–260.
Leven, E.J., and M.N. Gorgij. 2011. Fusulinids and stratigraphy of the Carboniferous and Permian in Iran. Stratigraphy and Geological Correlation 19(7): 687–776.
Leven, E.J., V.I. Davydov, and M.N. Gorgij. 2006. Pennsylvanian stratigraphy and fusulinids of central and eastern Iran. Paleontologica Electronica 9: 1–36.
Manukalova, M.F. 1950. Description of some new fusulinids from the Middle Carboniferous of the Donets basin. Geological and research work of the Main Directorate of Coal Exploration, Materials on Stratigraphy and Paleontology of the Donetsk basin: Moscow, Kharkov, Ugletekhizdat 175–192. (in Russian).
Miklukho-Maklay, A.D. 1959. About the stratigraphic significance, taxonomy and phylogeny of staffelloid foraminifera. Academy of Sciences of USSR, Nauka 125: 628–631. (in Russian).
Möller, V. von. 1877. Über Fusulinen und ähnliche Foraminiferen-Formen des russischen Kohlenkalkes. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1877: 139–146.
Möller, V. von. 1878. Die spiral-gewundenen Foraminiferen des russischen Kohlenkalks. Mémoires de l'Académie impériale des Sciences de St. Pétersbourg 7(25/9): 1–147.
Monod, O. 1977. Recherches géologigues dans Ie Taurus Occidental au Sud de Beyşehir (Turquie). Unpublished PhD Thesis, 1–571. Paris: l'Université de Paris Sud.
Okuyucu, C. 1997. Hadim Napı Karbonifer–Permiyen geçişi biyostratigrafisi [Biostratigraphy of Permian–Carboniferous passage of Hadim Nappe]. Unpublished MSc Thesis 1–155. Ankara: Hacettepe University, The Graduate School of Natural and Applied Sciences.
Okuyucu, C. 1999. A new Multidiscus? species (Foraminifera) from a fusulinacean-rich succession encompassing the Carboniferous-Permian boundary in the Hadim Nappe (Central Taurus, Turkey). Rivista Italiana di Paleontologia e Stratigrafia 105(3): 439–444.
Okuyucu, C. 2002. Toroslarda Anadolu Platformu Karbonifer–Permiyen geçisinin mikropaleontolojik incelemesi [Micropaleontological study of the Carboniferous–Permian passage of Anatolian Platform in Taurus Mountains]. Unpublished PhD Thesis, 1–207. Ankara: Hacettepe University, The Graduate School of Natural and Applied Sciences.
Okuyucu, C. 2008. Biostratigraphy and systematics of late Asselian–early Sakmarian (Early Permian) fusulinids (Foraminifera) from southern Turkey. Geological Magazine 145(3): 413–434.
Okuyucu, C. 2009. Systematics and biostratigraphic notes of the upper Moscovian–upper Gzhelian fusulinid foraminifers from the Anatolian platform in the Southern Turkey. Geologica Balcanica 38 (1–3): 35–51.
Okuyucu, C. 2013. Fusulinid zonation of the Late Moscovian-Early Sakmarian sequences from the Taurides, southern Turkey. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 268(3): 237–258.
Orlov-Labkovsky, O., F.R. Bensh, and N.M. Mikhno. 2007. Revision of the Carboniferous foraminiferal zonation of the Middle and South Tien-Shan, Central Asia. In XV International Congress on Carboniferous and Permian Stratigraphy, ed. Th.E. Wong, 305–316. Utrecht: Royal Netherlands Academy of Arts and Sciences.
Özgül, N. 1971. Orta Toroslarin kuzey kesiminin yapısal gelişiminde blok hareketlerinin önemi [The importance of block movements in structural evolution of the northern part of central Taurus]. Bulletin of the Geological Society of Turkey 14(1): 85–101.
Özgül, N. 1976. Toroslar’ın bazı temel jeoloji özellikleri [Some geological aspects of the Taurus orogenic belt]. Bulletin of the Geological Society of Turkey 19: 65–78.
Özgül, N. 1984. Stratigraphy and tectonic evolution of the Central Taurides. In Proceedings, International Symposium on Geology of the Taurus Belt, eds. O. Tekeli, and M.C. Göncüoğlu, 77–90. Ankara: Mineral Research and Exploration Institute of Turkey.
Özgül, N. 1997. Bozkır-Hadim-Taşkent (Orta Toroslar’ın kuzey kesimi) dolayında yer alan tektono-stratigrafik birliklerin stratigrafisi [Stratigraphy of the tectono-stratigraphic units in the region Bozkir-Hadim-Taskent (north central Taurides)]. Bulletin of the Mineral Research and Exploration 119: 113–174.
Peynircioğlu, A.A. 2005. Micropaleontological analysis and facies evolution across the Tournaisian-Viséan Boundary in Aladağ Unit (Central Taurides, Turkey). Unpublished MSc Thesis, 1–76. Ankara: Middle East Technical University, The Graduate School of Natural and Applied Sciences.
Putrya, F.S. 1938. On the stratigraphy of the Middle Carboniferous of the southeast part of the greater Donbass. Azov-Black Sea Geological Administration, Rostov 6: 48–72. (in Russian).
Putrya, F.S. 1939. To the study of medium-level foraminifera in the Donetsk basin. Azov-Black Sea Geological Administration 8: 175–188. (in Russian).
Putrya, F.S. 1956. Stratigraphy and Foraminifera from Middle Carboniferous deposits of the Eastern Donbass. In Microfauna USSR. Collection VIII. Foraminifera, bryozoans and ostracods of the Russian platform, Donbass, Tengiz basin and Kuzbass, ed. A.K. Grozdilova, Trudy VNIGRI 8(98): 333–485. Moscow: Russia State Scientific and Technical Publishing House, Oil and Mining and Fuel Literature-Leningrad. (in Russian).
Putrya, F.S., and G.E. Leontovich. 1948. To the study of Middle Carboniferous fusulinids from the Saratov Volga region. Bulletin of the Moscow Society of Naturalists 23(4): 11–45. (in Russian).
Rauzer-Chernousova, D.M. 1938. The Upper Paleozoic Foraminifera of Samara Bend and the Trans-Volga region. Academy of Sciences of USSR, Proceedings of Institute of Geology, Leningrad 7: 69–168. (in Russian).
Rauzer-Chernousova, D.M. 1948. Contributions to the foraminiferal fauna of the Carboniferous deposits of central Kazakhstan. Academy of Sciences of USSR, Proceedings of Insitute of Geology 62: 1–263. (in Russian).
Rauzer-Chernousova, D.M., G.M. Belyaev, and E.A. Reitlinger. 1936. Upper Paleozoic Foraminifera of the Pechora region. Proceedings of the Polar Commission AS USSR 28: 159–232. (in Russian).
Rauzer-Chernousova, D.M., G.M. Belyaev, and E.A. Reitlinger. 1940. About Carboniferous Foraminifera from the Samara Bend. Academy of Sciences of USSR, Proceedings of Institute of Petroleum and Geological Exploration 7: 1–88. (in Russian).
Rauzer-Chernousova, D.M., N.D. Gryzlova, G.D. Kireeva, G.E. Leontovich, T.P. Safonova, and E.I. Chernova. 1951. Middle Carboniferous fusulinids of the Russian Platform and neighboring regions. Moscow: Academy of Sciences of USSR, Proceedings of Insitute of Geology, Nauka. (in Russian).
Reitlinger, E.A. 1961. Stratigraphy of the Middle Carboniferous (Krasnaja Poljana, boring well no 1) of the Trans Volga. Academy of Sciences of USSR, Regional Stratigraphy of USSR 5: 218–260. (in Russian).
Reitlinger, E.A. 1971. Some problems of systematics in the light of evolutionary stage of Upper Paleozoic foraminifers. In Systematics, paleobiogeography and stratigraphic value of foraminifers, ed. D.M. Rauzer-Chernousova, Voprosy Mikropaleontologii 14: 3–16. Moscow: Academy of Sciences of USSR, Institute of Geology, Nauka. (in Russian).
Reitlinger, E.A. 1980. On boundary between the Bogdanovsky and Krasnopolyansky horizons (foraminifers of the Homoceras Zone). Voprosy Mikropaleontologii 23: 23–38. Moscow: Academy of Sciences of USSR, Institute of Geology, Nauka. (in Russian).
Rumyantseva, Z.S. 1962. New species of fusulinids from the Middle Carboniferous of the the Central Kyzylkums. In Stratigraphy and paleontology of Uzbekistan and paleontology of Uzbekistan and contiguous areas, vol. 1, ed. V.I. Verkhov, 169–185. Tashkent: Publishing House of the Academy of Sciences of Uzbekistan SSR. (in Russian).
Rumyantseva, Z.S. 1970. Stratigraphy and foraminifera of the Nayur and Lower Bashkir deposits of the Chatkal mountains system. In Biostratigraphy of sedimentary deposits of Uzbekistan, vol. 9, ed. T. Shayakubov, 138–184. Publishing House of the Academy of Sciences of Uzbekistan SSR. (in Russian).
Saltovskaya, V.D. 1974. Stratigraphy of Carboniferous deposits of the Zeravshan-Hissar mountain region. Dushanbe, Tajikistan: Izdateľstvo "Donish", Academy of Sciences of Tajikistan SSR, Institute of Geology. (in Russian).
Schellwien, E., and H. von Staff. 1912. Monographie der Fusulinen. (Geplant und begonnen von E. Schellwien) Teil III: Die Fusulinen (Schellwienien) Nordamerikas. Palaeontographica 59: 157–192.
Shlykova, T.I. 1948. Upper Carboniferous fusulinids of Samarskaya Luka. Trudy VNIGRI, New Series 31: 109–146. (in Russian).
Solovieva, M.N. 1977. Zonal stratigraphy of Middle Carboniferous deposits based on the fauna of fusulinids. Voprosy Mikropaleontology 19: 43–67. (in Russian).
Thompson, M.L. 1942. New genera of Pennsylvanian fusulinids. American Journal of Science 240(6): 403–420.
Toriyama, R. 1958. Geology of Akiyoshi, Part 3. Fusulinids of Akiyoshi. Memoirs of the Faculty of Science, Kyushu University Series D 7: 1–264.
Turan, A. 1990. Toroslar'da Hadim (Konya) ve güneybatısının jeolojisi, stratigrafisi ve tektonik gelişimi [Geology, stratigraphy and tectonic evolotion of Hadim (Konya) and its southwest in the Taurus]. Unpublished PhD Thesis, 1–229. Konya: Selçuk University, The Graduate School of Natural and Applied Sciences.
van Ginkel, A.C. 1965. Carboniferous fusulinids from the Cantabrian Mountains (Spain). Leidse Geologische Mededelingen 34(1): 1–225.
van Ginkel, A.C. 1987. Systematics and biostratigraphy of fusulinids of the Lena Formation (Carboniferous) near Puebla de Lillo (León, NW Spain). Proceeding Koniklijke Nederlandse Akademie van Wetenschappen 90(3): 189–276.
Villa, E. 1995. Fusulináceos carboníferos del Este de Asturias (N de España). Lyon: Biostratigraphie du Paléozoique 13: 1–261. Lyon: Université Claude Bernard.
Villa, E., and Ó. Merino-Tomé. 2016. Fusulines from the Bashkirian/Moscovian transition in the Carboniferous of the Cantabrian Zone (NW Spain). Journal of Foraminiferal Research 46(3): 237–270.
Zeller, D.E.N. 1953. Endothyroid Foraminifera and ancestral fusulinids from the type Chesteran (Upper Mississippian). Journal of Paleontology 27(2): 183–199.
Acknowledgements
This study was financially supported by the Scientific Research Projects Coordination Unit of the Selcuk University, Konya, Turkey (Project no: 17101003). Many thanks to the reviewers E. Villa, H. Forke and M. Aretz for their constructive remarks and very useful comments that significantly improved the manuscript, and H.-G. Herbig and M. Reich for the editorial handling.
Funding
This study was funded by the Scientific Research Projects Coordination Unit of the Selcuk University, Konya, Turkey (Project no: 17101003).
Author information
Authors and Affiliations
Contributions
Not applicable.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling Editor: Hans-Georg Herbig.
Rights and permissions
About this article

Cite this article
Akbaş, M., Okuyucu, C. Fusulinid biozonation of the Bashkirian–Moscovian successions from the Hadim Nappe, central Taurides, southern Turkey. PalZ 96, 517–541 (2022). https://doi.org/10.1007/s12542-022-00622-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-022-00622-w