
Abstract
The pattern of occurrence and recent changes in the distribution of macrobenthic organisms in fjordic and coastal (nearshore) Arctic waters are reviewed and future changes are hypothesized. The biodiversity patterns observed are demonstrated to be contextual, depending on the specific region of the Arctic or habitat type. Two major areas of biotic advection are indicated (the North Atlantic Current along Scandinavia to Svalbard and the Bering Strait area) where larvae and adult animals are transported from the species-rich sub-Arctic areas to species-poor Arctic areas. In those Arctic areas, increased temperature associated with increased advection in recent decades brings more boreal-subarctic species, increasing the local biodiversity when local cold-water species may be suppressed. Two other large coastal areas are little influenced by advected waters; the Siberian shores and the coasts of the Canadian Archipelago. There, local Arctic fauna are exposed to increasing ocean temperature, decreasing salinity and a reduction in ice cover with unpredictable effect for biodiversity. One the one hand, benthic species in Arctic fjords are exposed to increased siltation (from glacial meltwater) and salinity decreases, which together may lead to habitat homogenization and a subsequent decrease in biodiversity. On the other hand, the innermost basins of Arctic fjords are able to maintain pockets of very cold, dense, saline water and thus may act as refugia for cold-water species.
Similar content being viewed by others

Introduction
Much of the information that has been published to date on the marine aspects of climate change in the Arctic addresses vertebrates, plankton and sea-ice biota (Gradinger et al. 1999; Walther et al. 2002; ACIA 2005; Stempniewicz et al. 2007). Compared to the pelagic and ice domains, information on climate change effects on Arctic benthic fauna and flora are only slowly beginning to appear in the international literature (e.g. Sirenko and Gagaev 2007, Renaud et al. 2008a, b; Wassmann et al. 2010). Nevertheless, several studies have documented substantial ecological changes as a result of climate change. For example, Beaugrand et al. (2002, 2008) described a major northward shift of pelagic assemblages from the southern to the northern North Sea and from the North Sea to the Norwegian Sea. Similar phenomena for the benthic fauna are presented by Renaud et al. (2008a, b). Grebmeier et al. (2006) described poleward shifts of pelagic and benthic communities and consequent food web changes in the Bering Strait region following the massive sea-ice retreat in recent years. Additionally, a major northward shift of boreal fish species was associated with the warming of the North Atlantic in the 1930s (Drinkwater 2006). Furthermore, it is predicted that, as the extent of summer sea-ice cover continues to decrease over the coming decades (ACIA 2005), the habitat for the sympagic community—a characteristic element of biodiversity in Arctic waters and an important source of energy for sea floor assemblages (McMahon et al. 2006; Sun et al. 2007)—will in turn decline. The loss of sea ice may be very rapid, with some models indicating that the Arctic could be ice-free during summer months by 2050 (Johannessen and Miles 2000; Comiso 2006).
The clearest indication of a changing Arctic climate is an increase in both air and ocean temperatures. Predictions of the extent of change vary with both the model used and the carbon usage scenario simulated, but it is reasonable to assume that by 2050 there will be a 2.5°C rise in mean air temperature, followed by a further increase of as much as 4.5°C by the end of the century (ACIA 2005). Increases in air temperature will lead to subsequent increases in ocean temperature. While ocean temperature plays a key role in controlling the distribution of pelagic species, in the case of the benthos it is only one element of a broad range of factors that define the ecological niche of a species. Under a changing climate, factors such as seafloor morphology and sediment type may be largely unaffected. Consequently, it is difficult to discuss regime change in the same way as has been done for the plankton, for which changes are related to water mass and biotic poleward advection (Beaugrand et al. 2002, 2008). Nevertheless, it is very clear from many studies in both Europe and the Pacific Arctic that boreal species at the northern edge of their ranges are shifting their distributions polewards (Helmuth et al. 2006; Mieszkowska et al. 2007; Hawkins et al. 2008). Each species will have its own characteristic rate of movement, which is mostly related to its mode of reproduction. Clearly, species with planktonic larvae have greater capacity to spread farther and faster, but having extended their range their persistence is likely to be less than that of species with either lecithotrophic eggs or direct development (Mileikovsky 1968; Berge et al. 2005). As each boreal species expands polewards, new competitive and predator-prey interactions will occur. The outcomes of these new interactions are difficult to predict. While it is reasonable to predict that benthic assemblages in polar regions will change as ocean temperatures rise, the rate at which they will do so and the composition and function of the future assemblages cannot yet be forecast.
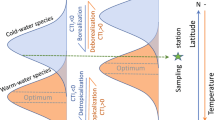
In this paper we address the changes that are likely to take place with the seafloor biota of Arctic coastal waters. For several of these discussed changes, evidence is already accumulating and references are provided accordingly, while other changes are hypothetical at present. Change in Arctic benthic ecosystems will depend on the extent to which they are connected to boreal ecosystems. In those areas that are open to the exchange of propagules, new species will be advected northwards and existing species may be displaced. In this scenario, biological interactions will play a dominant role in the change. In areas isolated from low-latitude immigrants, the balance of interactions between endemic species will change and new patterns of dominance may emerge as a result of a decline in salinity and increase in both temperature and turbidity. Here physical factors will play a dominant role (Fig. 1).

Two major modes by which biodiversity is changing in the Arctic. Arrows indicate inflow of warm water with its biota northward from the south. Ovals indicate generally the areas of warming of nearshore waters with local fauna
The first mechanism driving biological change is the direct transfer of propagules from south to north by strong current systems such as the North Atlantic-West Spitsbergen Current, the Bering Shelf and Anadyr waters and Alaska Coastal Current and, to a limited extent, the Baffin Bay Current. The strong interannual changes of Atlantic water inflow into the Arctic, with the 5°C isoline at 100 m depth moving up to 1,000 km in a decade (Fig. 2) show the variability of this advection. In such circumstances, the coastal benthic physical habitat properties are relatively unchanged but the habitats receive new species with the imported water. As the coastal Arctic waters are ecologically very young (only ice free for the last 10,000 years), their level of endemism is very low (Zenkevitch 1963, Dunton 1992). The number of benthic species currently living in Arctic waters decreases with increasing distance (and decreasing habitat age) from the boreal source species pool (Sirenko 2001).

Extreme positions of Atlantic water at Svalbard shelf, July 2001 and 2006. Temperature at 100 dbar depth, based on Walczowski and Piechura (2007), supplemented with data from IOPAS hydrography archive, courtesy W. Walczowski
The second mechanism is the warming and declining salinity of nearshore waters (Greene et al. 2008). This mechanism will operate over large coastal areas, including the Siberian coast and the Canadian Archipelago, that are not directly influenced by boreal water advection (Fig. 1). Such areas will be far slower to gain new species and will be largely influenced by local warming and strong discharge of fresh, turbid water flowing from the rivers and thawing tundra. In those coastal areas, the existing species pool is modest in size and more of the species are Arctic endemics, mainly brackish water-adapted amphipods (Gurjanova 1951). As the main characteristic of coastal fauna off Siberia is a high tolerance to low salinity, it is likely that the fauna will cope well with a certain degree of additional salinity decrease, but the already low diversity makes this assemblage vulnerable to species loss.
State of knowledge of coastal biota
The intensity of field and taxonomic studies varies greatly from place to place. A single Arctic fjord may contain anywhere from 200 to 1,000 benthic macrofaunal species (Table 1). Based on estimates by Sirenko (2001) that the macrobenthic fauna of the Arctic shelves number over 3,000 species and by Palerud et al. (2004) that there are about 1,500 species around the Svalbard archipelago. The current estimate for Hornsund Fjord alone is over 800 macro zoobenthic species (http://www.iopan.gda.pl/projects/ATBI/atbi.html).
More complete data are available for intertidal biodiversity. The Arctic intertidal is species poor due to ice scour, UV radiation, desiccation and wave exposure. Estimates suggest that the Arctic as a whole is home to no more than 100 intertidal macrobenthic species (Table 2). Although sampling effort has been unevenly distributed (Fig. 3), it appears that most intertidal species are circumpolar with fewer found off islands than are found off continental landmasses. Vast areas of Beaufort Sea and Siberian coasts are devoid of intertidal macro-organisms as beaches are predominantly gravel or stony, substrates that provide neither shelter nor a stable substratum for macrofauna.

Number of macrobenthic species reported from the intertidal zone (0-2 m) in the Arctic compared with Northern Norway (boreal/subarctic zone); data selected from references listed in Table 2. Note that sampling effort varies between regions
The physical diversity of Arctic coastal benthic habitats may be considered higher than that in sub-arctic environments. That is simply because ice is an additional physical factor that can be added to basic habitat determinants like depth, substrate, and exposure. As oceanic and atmospheric temperatures increase, occurrence of ice will be reduced and habitat complexity will decline. Furthermore, sea ice reduces the impact of wave action on shallow water assemblages and therefore its loss will effectively increase exposure, creating erosion and turbidity in coastal waters (Rachold et al. 2004).
Aim of this paper
The present review of the status and observed and predicted changes of biodiversity of Arctic coastal and fjordic waters is limited to the macrobenthos and focused on fjordic basins and open coastal waters to the depth of the offshore euphotic zone (approx. 30 m). Up to date analyses of the biodiversity of Arctic shelf benthos and the benthos of the central deep basin are presented by Piepenburg et al. (2010) and Bluhm et al. (2010). In addressing the ecological impacts of climate change, it is important to realise that many changes will take place and that each process will have its own intensity and operational time scale. For example, the retreat of glaciers occurring within the coming decades will result in increased turbidity in inshore waters, while at the same time opening up new areas of seabed for colonisation. However, in the longer term glaciers will either disappear completely or will be greatly diminished. In such circumstances turbidity will be massively reduced compared with present values and the chemistry of inshore waters substantially changed. Our review considers the shorter time scale only, and analyse processes that can be currently observed. As the most complete inventories exist for two Svalbard fjords (Hornsund and Kongsfjorden) and coastal bays in Greenland (Young Sound), Alaska (Prudhoe Bay) and Franz Josef Land (Table 1), we took these five locations as the basis for this review.
There is no single, simple way to predict the impacts of global climate change on the shallow-water biota of the Arctic seas; rather, they may result in the increase, decrease or stabilization of biodiversity in the Arctic marine nearshore realm. The assessment of Arctic biodiversity is dependent on the type of biota, taxon or geographic scale considered. Below, some prominent and the most likely cases are discussed for the fjordic and coastal macrobenthos.
Scenarios of change
Biodiversity increase
-
1.
Advection of boreal species into the Arctic.
Advection of species new to the Arctic happens directly in two areas. The main route is along the North Atlantic West Spitsbergen Current, which flows along the Norwegian coast and provides close to 70% of the present-day inflow into the Arctic Basin (Dickson et al 1999). Atlantic water flows over the Norwegian shelf and slope, from the surface to some 800 m depth, sinking below the surface (below 100 m) near Svalbard (Walczowski and Piechura 2006, 2007). The Norwegian Sea shelf is distinctly richer in species than the European Arctic shelf of Svalbard and its adjacent archipelagos. The faunal inventories of Brattegard and Holthe (1997) estimate over 3,400 macroinvertebrate species for the coast of Norway while Palerud et al. (2004) estimate about 1,500 species for Svalbard (in both cases macroinvertebrates only). Both regions are comparatively well-studied based on long-term research programs in several fjords of mainland Norway and two fjords, Hornsund Fjord and Kongsfjorden, in Svalbard. However, the coastal area of Norway is much larger than Svalbard, and this fact alone is linked with higher species richness. Among the most studied groups are the Amphipoda, represented by 740 species in Northern Norway and 270 species on Svalbard (Palerud et al. 2004).
The second area of advection is the Bering Strait, where waters from the Bering Sea enter the Beaufort and Chukchi Seas in the Arctic Ocean. Here, the shallow Bering Strait (50 m) prevents deeper water transport and the volume of water that enters the Arctic this way is estimated at some 20% of the inflow budget (Dickson et al. 1999; Maslowski et al. 2008). Pacific waters entering the Arctic Ocean through the Bering Sea are estimated to contain approximately 5,000 macrobenthic species (Grebmeier et al. 2006), more than double the 1,500 or so species recorded in the Chukchi and Beaufort seas (Sirenko 2001). In addition to natural transport by ocean currents, the introduction of species new to the Arctic through the expected increase in human activity, particularly shipping, could occur just as quickly as it happened in temperate locations (Gollasch 2006).
Major current systems can transport larvae, propagules and even adult organisms to and from the Arctic (Mileikovsky 1968), though there are surprisingly few recent records of such advection. Nevertheless, the boreal bivalve Mytilus edulis has recently reappeared on the Svalbard archipelago after more than 1,000 years of absence. Recolonisation was observed in two stages; it was recorded in Bjornoya in 1994 (Weslawski et al. 1997) and then in Spitsbergen 10 years later (Berge et al. 2005). Besides M. edulis, a crab (Geryon tridens) and a benthic amphipod (Schisturella sp.) have made similar northward shifts (Vader 1980; Vader and Johnsen 2000). An extensive survey of shelf and coastal waters by Blacker (1957) revealed within-Arctic shifts in the distribution of Decapoda and Echinodermata species with northern advancement of the boreal species Sclerocrangon boreas, Sabinea sarsi, Hippasteria phyrigiana, Gorgonocephalus eucnemis and Poliometra prolixa in comparison with nineteenth century surveys. However, no species new to the Arctic area were recorded, only northward shifts and increases in abundance of southerly species known previously from the area. Similar results appear in long-term surveys by Beuchel et al. (2006) and Kędra et al. (2010). Although both recorded benthos at a limited number of stations, biodiversity on soft and hard bottoms increased in the outer parts of a Svalbard fjord, but all species new to the fjord were already recorded elsewhere in the Svalbard archipelago. Similarly, studies in the Pacific Arctic showed changes in the diversity of Arctic fauna and structure of the food web, yet inflow and colonisation of the area with new benthic species was limited to single species (Grebmeier et al. 2006). Specifically, a few individuals of the Pacific bivalve Pododesmus macrochisma and the crabs Oregonia gracilis and Telmessus cheiragonus were recently recorded for the first time on the Alaska coast of the Chukchi Sea (Sirenko and Gagaev 2007). It is unlikely that large conspicuous organisms like these could have been overlooked in the previous, relatively intense sampling of the region, and hence a recent northward expansion of their range can be assumed. On the other hand, similar understanding of smaller and more difficult taxa is far from complete in the Arctic region; see biogeographic data base OBIS (http://www.iobis.org/).
-
2.
Effects of shrinking ice cover and duration
Another factor likely to increase coastal benthic diversity in the Arctic is the reduction in sea ice duration, thickness and extent. All forms of ice can be destructive to benthic animals; through freezing of body tissues, freezing of the sediment habitat, mechanical destruction by ice scouring and low temperatures. A number of studies on the effects of ice on shallow-water benthos have been performed in boreal areas of Europe (e.g. Beukema 1979; Reise et al. 2001) and reviews relevant to polar areas were published by Barnes (1999) and Gutt (2001). Unlike the situation in offshore waters where pack ice occurs, there are no specific species assemblages directly associated with the nearshore ice forms (fast ice, ice foot, cryolittoral). A warmer Arctic will lead to a reduction in ice-related pressures on the shallow water biota due to a shorter ice season, a smaller area of ice coverage and the reduction in thickness of the ice that does form. The relaxation of these stressors might lead to an enhancement of biodiversity in a number of ways. The disappearance of the ice foot and fast ice on rocky coasts will permit the survival of perennial macroalgae and their associated fauna in the littoral zone, resulting in an increase in biomass and diversity (Fig. 4, Weslawski et al. 2010). An exceptional case of a habitat that might be lost may be expected in those areas where the ice foot actually creates a favourable habitat for the biota. An example would be the creation of a stable ice roof over the water column that can maintain organisms at a temperature higher than on an exposed, ice-free shore (Scrosati and Eckersley 2007). A reduction in sea ice on soft sediment shores, particularly tidal flats, could have more dramatic effects than on hard bottom. Until recently, Arctic tidal flats were solidly frozen from October to June, with ice penetrating deep into the sediment. During the thaw, large chunks of ice, firmly frozen to the surface sediment, detached and floated away causing defaunation and facilitating a pattern of colonisation on an annual cycle. Such repeated cycles of defaunation and recovery lead to high biological variability in the biota of Arctic tidal flats (Weslawski and Szymelfenig 1999; Conlan and Kvitek 2005). As the ocean warms and ice cover thickness and duration decline, there is likely to be a development of a perennial, stable tidal flat biota, and an increase in the frequency of species interactions (Fig. 5). Such an increase in biotic interactions was already reported after colonization of Arctic shallow waters by Bryozoa (Barnes and Kuklinski 2003; Kuklinski and Barnes 2005). Another consequence of ice disappearance from tidal flats is a longer period of exposure to wave erosion, subsequently leading to a coarser sediment structure and a consequential change in the composition of the fauna. If these effects are not too severe, such disturbance at intermediate levels might increase biodiversity.

Observed environmental consequences of a disappearing ice foot in Arctic coastal waters. a Presence of ice foot: clear water and absence of macrophytes in the upper littoral. b Absence of ice foot: increased turbidity and erosion through waves, and occurrence of macrophytes higher up on shore

Environmental consequences of changing fast ice cover in sedimentary shores. a. Substantial fast ice cover: massive sediment displacement and effective defaunation during spring melt. b. Diminished fast ice cover: intermediate disturbance, increased patchiness, possibility of perennial community survival
Iceberg scour will become less frequent as glaciers thin and begin to melt (Rabus and Echelmeyer 2002; Motyka et al. 2003). The impact of ice scour in Arctic benthos is greatest in glaciated fjords at 20–100 m depth, causing sediment displacement and direct removal of organisms (Gutt 2001). The relaxation and eventual disappearance of this pressure will permit the development of more stable, biologically controlled communities.
The pattern of increasing biodiversity as one moves away from glacier fronts is broadly similar to that found as one moves away from a point source of organic matter load or pollution (Pearson and Rosenberg 1978; Rhoads et al. 1978). More diverse assemblages consisting of larger-bodied individuals can be found with increasing distance from glaciers (Włodarska-Kowalczuk et al. 2005). As a glacier retreats, the intensity of the ice impact on the seabed declines. According to the intermediate disturbance hypothesis, this could result in higher benthic biodiversity due to the moderating of the ice as a stressor (Huston 1979; Conlan and Kvitek 2005). For the negative impacts of ice retreat see the paragraph on siltation below. Today, the impact of ice scour varies greatly from place to place. Shallow waters are obviously impacted more strongly than offshore areas and locations close to a glacial front are more impacted than those farther away. In distant and deeper places, the frequency of moderate disturbance might decline and, if it does, diversity will also decline. In the future, in areas close to the glacial front, severe disturbance will abate and diversity will increase. The intermediate disturbance that promotes diversity will operate with varying intensities between these extremes.
The recession of ice and the warming of surface waters are predicted to result in a prolonged season of primary production (ACIA 2005; Wiktor and Wojciechowska 2005; Piwosz et al. 2008; Ellingsen et al. 2009). However, the overall impacts of a warmer climate on productivity and organic flux to the benthos are difficult to assess. On the one hand, warming may result in a loss of ice algal production (see model by Ellingsen et al. 2009) and, consequently, a loss of efficient sympagic-pelagic-benthic coupling. On the other hand, the transport of increased pelagic organic matter production to the seabed might be prolonged and thus increased, resulting in a higher overall POC content of the sediment (Kedra et al. 2010). A moderately elevated food supply may result in an increase in benthos biodiversity (Witman et al. 2008), although studies on productivity-diversity relationships have produced rather controversial results. However, very high organic matter load can be damaging for the benthos through extreme levels of microbial decay and subsequent oxygen depletion, as has repeatedly been shown for isolated fjord basins in temperate and warm climates (Syvitski et al. 1987).
Biodiversity loss
-
1.
Habitat homogenization and sedimentation
Habitat homogenization may occur as a result of the lack (or severe reduction) of ice in Arctic fjords and the increase of freshwater run-off with its associated sedimentation. Together these changes will lead to the loss of small geomorphological structures and physical barriers, and thereby contribute to larger and more uniform seascapes (Kedra et al. 2010). Currently, the size of faunistically homogeneous benthos patches increases from the diverse outer fjord to the more uniform inner fjord bottom (Kendall et al. 2003; Wlodarska-Kowalczuk and Weslawski 2008). Such habitat homogenization is regarded as one of the most important threats to biodiversity as the number of rare species can be related to the level of patchiness and habitat heterogeneity (Hewitt et al. 2009).
An increase in coastal siltation and a reduction in the depth of the euphotic layer will occur when melt-water discharge increases as a consequence of warming. As the volume of melt-water increases so will the volume of mineral particles discharged into glacial bays and surrounding coastal areas. Rapidly retreating glaciers already discharge substantial amounts of particulate material (over 500 mg/dm3 in some areas; Svendsen et al. 2002), leading to heavy sedimentation that may exceed 200 g/m2/day (op. cit.). Heavy sedimentation of mineral particles is a direct stressor for those benthic species that need to maintain contact with the sediment surface, and acts to effectively dilute organic material, making feeding more energetically expensive (Moore 1977). An additional impact on benthic productivity relates to the depression of phytoplankton production in the highly turbid surface water close to glacial fronts. The combination of these physical and biotic factors is responsible for the reduced biodiversity in areas of strong siltation (Syvitski et al. 1989; Wlodarska-Kowalczuk and Weslawski 2001; Jewett et al. 2009). As glacial retreat progresses the situation can be expected to intensify (Fig. 6).

Environmental consequences of increased glacial melt in fjords. a Moderate glacial melt. b Large glacial melt: increased turbidity and sedimentation, less winter cooled water, homogenisation of seabed habitats from three distinct zones of benthos distribution patterns in a towards two zones in b
-
2.
Decrease in the strength of the pelagic-benthic coupling
Petersen and Curtis (1980) noted that, functionally, the main difference between Arctic and temperate biological assemblages lies in the strength of pelagic-benthic coupling. In the Arctic, most of the primary production sinks to the seafloor ungrazed, creating favourable food conditions for benthic animals (McMahon et al. 2006; Sejr et al. 2007; Renaud et al. 2008a, b). The loss of ice biota to be expected with the shrinking ice cover and the change from the short, intense cycle of Arctic primary production towards a longer weaker period of productivity (ACIA 2005), coupled with the switch from ice diatoms to small pelagic flagellates (Keck et al. 1999; Piwosz et al. 2008) in a warmer ocean will likely lead to less food becoming available to the benthos (Hunt et al. 2002; Carroll and Carroll 2003). Besides quantity, a change in food quality can be expected from the PUFA-rich ice diatoms to less energy-rich microplankton (Hop et al. 2006; Sun et al. 2009). These changes are most likely to occur in productive shelf areas where the ice-pelagic-benthic coupling is most intense (Wassman et al. 2006). Coastal and fjord waters are dominated by first-year ice of relatively lower productivity (Keck et al. 1999) and have strong spring and summer production that reaches the seabed (Zajaczkowski et al. 2010).
Unchanged biodiversity
-
1.
Food supply.
The overall period of primary production in the Arctic is and will remain comparatively short, with a long period of darkness. The Polar Night at 77°N lasts from the beginning of November to late February, and the amount of light is insufficient for primary production by late September (Eilertsen et al. 1989). As a result, supply of fresh food from the water column is highly seasonal for the benthos and will remain that way to some extent. While taxa depending on fresh food undergo starvation periods during the winter (Weslawski et al 1991), other taxa appear to maintain their diets stable. An example is benthic predatory amphipods from the productive coastal areas of Svalbard that have a striking similarity in their diets in winter and summer (Legeżynska and Kedra pers. comm.). Stable feeding conditions for predatory and opportunistic benthos such as these amphipods seem to be related to the longevity of most Arctic invertebrates. In areas with perennial macroalgae such as the Boulder Patch in the Alaskan Beaufort Sea, some of the carbon within the benthic detrital food web is derived from perennial macroalgae, similarly providing a relatively stable food source for benthic organisms (Dunton and Schell 1987).
However, the high reliance on detrital deposits and an opportunistic feeding mode (Iken et al. 2010) and the existing adaptation to survive long periods of seasonally low food availability may make the benthic biota well adapted to some degree of change (Sun et al. 2009). Such adaptability may well act as stabilizing factor for biodiversity.
-
2.
Life history adaptations
Long life cycles contribute to functional, if not taxonomic biodiversity among long-lived, large species (Weslawski et al. 2010). For example, in Arctic amphipods, the juvenile, immature and adult stages are not only different in size but also have different habitat preferences, food spectra, patterns of mobility, etc. Thereby, an Arctic population of a species with a 5-year life span may contribute to five different functional groups, effectively playing the role of up to five different species in the ecosystem. At lower latitudes, in contrast, similar species are able to complete their life cycles in less than 1 year and juveniles are not functionally much different from adults (Weslawski et al. 2010). At lower latitudes differences in life history traits over the latitudinal range of species are common. Also, there is increasing evidence that some boreal species, in the northern limit of their range, are currently developing characteristics that 20 years ago were typical of individuals living much further south (Mieszkowska et al. 2007). If changes illustrated in this paragraph occurred as a consequence of warming, the Arctic might loose a considerable element of functional diversity.
-
3.
Cold water refuges
The annual autumn cooling and freezing of surface water leads to the sinking of cold, saline and oxygen-rich water to the bottom of Arctic fjords. This cold water is retained within the system because deep-water exchange is usually restricted by the sill at the entrance to the fjord (Syvitski et al. 1987). Thus, the main difference between Arctic and temperate/warm climate fjords is that the former always contains oxygen-rich bottom water, while seasonal oxygen depletion is typical for the near-bottom waters of temperate fjords. Observations from the Svalbard (Hornsund) fjord inner basin show that during the last 30 years the near-bottom temperature ranged between -1.8°C and –0.6°C (Fig. 7). Such stable conditions provide a refuge for cold-water stenothermic species and conserve local species inventories at a time when the outer fjord basins are influenced by warmer shelf waters, the temperatures of which are far more variable (Fig. 8). Relict populations of cold-water species are also known from boreal fjords (Drainville 1970; Brattegard 1980).

Stability of hydrological conditions in inner fjord basins. Near-bottom temperature (°C), expressed as the difference between annual maximum and minimum temperature in the inner fjord basin (Brepolen, Hornsund, 77°N) (black bars) and outer fjord basins (light bars). Archival data from IOPAS

Example of extreme stability of biodiversity (species richness) of soft bottom macrofauna in the inner fjord basin and changeable biodiversity in the outer fjord exposed to shelf water [50–100 m depth, Kongsfjorden, Svalbard; after Kedra (2008)]
On the western coast of Svalbard, the proportion of species of Arctic origin is higher in the inner fjord basins than in other fjord areas (Weslawski 1990). The more isolated the basin, the less exchange it has with shelf water and the greater hydrological stability that might be expected. For example, the benthos of the semi-enclosed Van Mijen Fjord on Svalbard has shown very little change between 1980 and 2002 (Renaud et al. 2007). Similar observations come from the inner basin of Kongsfjorden of the period 1996-2006 (Kedra et al. 2010). Even in a warming Arctic, seasonal ice is likely to be formed in the inner fjord basins in winter, providing cold, oxygen-rich bottom water. This situation may preserve Arctic conditions in small isolated pockets, providing a refuge, at least for some soft bottom deep sublittoral species.
-
4.
Direct species replacement
As the Arctic Ocean warms, new patterns of species distribution will emerge as boreal species and populations will be able to invade northern waters and compete with functionally similar resident species. In the short term we might expect a diversity increase, with patches of coexisting residents and competitors (Beuchel et al. 2006). Over time though, diversity will likely stabilise at some level as outcomes of the novel competitive and predator-prey interactions emerge. Since every species will shift its distribution at its own intrinsic rate, the emerging patterns of competition will not just be between immigrants and residents but also between immigrants themselves. Species with long-lived planktonic larvae will immigrate first but may not persist as their larvae are easily drifted away and lost for the area (Johanesson and Johanesson 1995). Species with direct development will be slower to arrive but once established are more likely to remain. For species living on unvegetated bottom with a continuous habitat between their current range and their potential future range in a warmer ocean, an extension of their distribution is straightforward. Taxa living in discontinuous habitats such as the rocky intertidal or in association with macrophytes need to bridge the gaps between suitable habitats, for example, between the shores of Norway and those of Svalbard, and expansion of the distribution ranges may take far longer, as observed, for example, in the slow Mytilus recolonization in Svalbard (Berge et al. 2005).
A number of Arctic species have close relatives in species of a more boreal distribution, yet their ecological characteristics are so similar that they compete directly for the same habitat and resources; replacement of one such species by the other has no consequence for the biodiversity level. With the increasing Arctic warming, cold water species are already retreating and are being replaced by boreal species (e.g. Weslawski et al. 2010). A number of such species pair replacements in the benthos was presented for Svalbard by Blacker (1957) and was linked to the large-scale hydrographic changes with the advance of warm waters to the European Arctic in 1930-1950 (op. cit). Recent observations of changes in the Spitsbergen intertidal zone show the result of the same process among intertidal amphipods, with boreal Gammarus oceanicus on Spitsbergen replacing the Arctic species G. setosus (Weslawski et al. 2010). A similar process is described in the pelagic domain with the boreal copepod Calanus finmarchicus replacing the Arctic C. glacialis (Hop et al. 2006).
Processes that affect nearshore biodiversity in unknown direction
-
1.
Fisheries
Fisheries impacts on the Arctic are likely to increase and move from the sub-Arctic into the high Arctic, directed more toward pelagic fish (e.g. capelin, Walleye Pollock, herring, mackerel) than demersal species (cod, halibut, sole) (ACIA 2005; Christiansen et al. 2008; Pope et al. 2009). The effects of increased fishing on the coastal Arctic benthos are unclear. On the sub-Arctic shelves of the Bering and Barents seas, there is direct impact through scouring of the seafloor by heavy fishing gear, particularly damaging species with shells or carapaces and causing the replacement of long-lived species by smaller opportunists (McConnaughey et al. 2000; Brown et al. 2005).
Studies on fishery effects in boreal waters indicate a decrease in bivalves and increase in mobile crustaceans such as crabs and shrimps (e.g. Rumohr and Kujawski 2000; Quijon and Snelgrove 2005). Potential future fisheries of Arctic long-living, slow growing species such as scallops will offer substantial short-term financial gains to fishermen and exploitation can be expected to be intense for a short time period. It is unlikely, however, that this expected catch will ever be replaced in the fished populations as long as fishing pressure persists as the individuals in many populations are very old [see example of Chlamys islandica exploitation on Iceland; Jonasson et al. (2007)].
-
2.
Predation by benthic-feeding top predators
The high benthic biomass on the arctic shelves is exploited by specialized predators, including grey whales and walrus. The grey whale, a benthic feeder, is an important predator in offshore shelf areas (Grebmeier et al. 2006). However, the extent of predation varies greatly seasonally and between regions, and potential influences of predation on benthic diversity are difficult to ascertain, but might be substantial (Coyle et al. 2007), though dependant on the stock fluctuations. Depending on the regions, walruses feed on nearshore and shelf benthos, often using ice floes as platforms for rest (Born et al. 2003). In areas like the north Greenland coast the access to fjords is seasonally restricted by fast ice and walrus predation is restricted to 3 months in summer during the open water season. A reduction in ice cover would thus increase benthic predation by walrus in those areas. In other regions of the Arctic such as the Chukchi Sea, walruses need areas of thin or broken pack ice to enable them to feed and rest. The recession of the summer ice edge will make such conditions increasingly rare. This will force walruses to haul out on shore (Freitas et al. 2009) and utilize shallow feeding grounds, where they will have a strong impact on shallow-water biota. Walrus feeding strongly influences productivity and ecological function in the Bering Sea benthos as the species’ activities cause benthic bioturbation and nutrient flux (Ray et al. 2006). In this area walruses annually consume an estimated 3 million metric tons of benthic biomass and feed over thousands of square kilometres of Bering Sea shelf per year. The absence of the walrus as a consequence of ice loss or its concentration in nearshore locations would, therefore, influence benthic biomass (Ray et al. 2006; see also Kovacs et al. this issue), and, though selective feeding with a preference of bivalves, community composition as well.
Important benthic invertebrate predators are reptant decapods (crabs). Currently, this group of crustaceans appears to be rare or absent in waters with permanent sub-zero temperatures due to poor cold tolerance associated with high magnesium levels in their body fluids (Frederich et al. 2001). A northward range expansion, as already observed with the snow crab in the Barents and Chukchi Seas (Alvsvåg et al. 2009; Bluhm et al. 2009), is expected to influence to the benthic food web and potentially the diversity as well.
-
3.
Ocean acidification
Although this paper is focused on the effects of warming on the Arctic, ocean acidification will also play an increasingly important role in shaping the biologic landscape of the Arctic in the future. An increase in atmospheric carbon dioxide has led to a decline in the pH of the ocean. Increasing levels of dissolved carbon dioxide will have their own effects on the biogeochemistry of Arctic ecosystems and the species they contain. Indeed, low Arctic water temperature will lead to more severe acidification effects in the polar regions than in any other part of the planet (ACIA 2005). Within 50 years, aragonite saturation in the Arctic will be too low for species such as molluscs and crustaceans to be able to produce their shells (Bellerby et al. 2005; Orr et al. 2005). Impacts will not simply be limited to calcifying animals and plants, but all species are expected to suffer some physiological impact, e.g. disruption in breeding and growth (Havas and Rosseland 2004; Widdicombe and Spicer 2008). In Svalbard, the impact of reduced pH on the barnacle Semibalanus predicted for the mid-century is expected to be greater than the effect of temperature (Findlay et al. 2009), but the relative impacts of these factors are reversed at the species’ southern limit. Such additive effects may also be expected for other species.
Conclusions
The coastal and fjordic diversity in the Arctic does not have a uniform pattern, and the changes that might be expected as a consequence of a warmer, more acidic ocean will differ from place to place and across ecosystem compartments. The realization that a diversity of ecosystem responses can be expected is important for setting standards of environmental quality and “naturalness”. This is particularly relevant when biodiversity is used as an indicator in national or international management or conservation efforts.
The predicted temperature rise will, in itself, be of minor direct importance for coastal benthos as nearshore taxa are often adapted to a wide temperature range. More important for the future of the coastal benthos are variables associated with temperature rise; an increase of coastal turbidity and sedimentation, changes in ice cover, an increase in storminess, increasing coastal erosion and the freshening of surface waters, all of which will occur at the same time that ocean acidification is also increasing. Such complex environmental changes will reshape coastal biocenoses and likely drive them to a new state, possibly close to or beyond a point of no return. Changes will not eradicate Arctic nearshore and fjordic benthos, but give a new shape and function to the familiar Arctic. Understanding the change in the Arctic ecosystem requires the knowledge of organisms at a taxonomic and species level and detailed knowledge of the individual species life histories. This is because a number of important changes may occur at the level of closely related, morphologically similar species that often play significantly different roles in the system.
References
ACIA (2005) Arctic climate impact assessment. Cambridge University Press, New York
Alvsvåg J, Agnalt AL, Jørstad KE (2009) Evidence for a permanent establishment of the snow crab (Chionoecetes opilio) in the Barents Sea. Biol Invasions 11:587–595
Barnes DKA (1999) The influence of ice on polar nearshore benthos. J Mar Biol Ass UK 79:401–407
Barnes DKA, Kuklinski P (2003) High polar spatial competition: extreme hierarchies at extreme latitude. Mar Ecol Prog Ser 259:17–28
Beaugrand G, Reid PC, Ibanez F, Lindley JA, Edwards M (2002) Reorganisation of North Atlantic marine copepod biodiversity and climate. Science 296:1692–1694
Beaugrand G, Edwards M, Brander K, Luczac C, Ibanez F (2008) Causes and projections of abrupt climate-driven ecosystem shifts in the North Atlantic. Ecol Lett 11:1157–1168
Bellerby RGJ, Olsen A, Furevik A, Furevik T, Anderson LG (2005) Response of the surface cean CO2 system in the Nordic Seas and northern North Atlantic to climate change. In: Drange H, Dokken TM, Furevik T, Gerdes R, Berger W (eds) Climate variability in the Nordic seas. Geophysical Monograph Series 158, AGU, pp 189-198
Berge J, Johnsen G, Nilsen F, Gulliksen B, Slagstad D (2005) Ocean temperature oscillations enable reappearance of blue mussel Mytilus edulis in Svalbard after 1000 years of absence. Mar Ecol Prog Ser 303:167–175
Beuchel F, Gulliksen B, Carroll ML (2006) Long-term patterns of rocky bottom macrobenthic community structure in an Arctic fjord (Kongsfjorden, Svalbard) in relation to climate variability (1980-2003). J Mar Sys 63:35–48
Beukema JJ (1979) Biomass and species richness of the macrobenthic animals living on a tidal flat area in the Dutch Wadden Sea: effects of a severe winter. Neth J Sea Res 13:203–223
Blacker RW (1957) Benthic animals as indicators of hydrographic conditions and climate change in Svalbard waters. Fisheries Investigations, London, Series 2 20:1–49
Bluhm BA, Iken K, Mincks Hardy S, Sirenko BI, Holladay BA (2009) Community structure of epibenthic megafauna in the Chukchi Sea. Aquat Biol 7:269–293
Bluhm BA, Ambrose Jr WG, Bergmann M, Clough LM, Gebruk AV, Hasemann C, Iken K, Klages M, MacDonald IR, Renaud PE, Schewe I, Soltwedel T, Wlodarska-Kowalczuk M (2010) Diversity of the Arctic deep-sea benthos. Mar Biodiv (in press)
Born EW, Rysgaard S, Ehlme G, Sejr M, Acquarone M, Levermann N (2003) Underwater observations of foraging free-living Atlantic walruses (Odobenus rosmarus rosmarus) and estimates of their food consumption. Polar Biol 26:348–357
Brattegard T (1980) Why biologists are interested in fjords. In: Freeland HJ, Farmer DM, Levings CE (eds) Fjords oceanography. Plenum Press, New York, pp 53–66
Brattegard T, Holthe T (1997) Distribution of marine, benthic macro-organisms in Norway. A tabulated catalogue. Preliminary edition. Research Report for DN, Nr. 1997-1, Trondheim
Brown EJ, Finney B, Dommise M, Hills S (2005) Effects of commercial otter trawling on the physical environment of the southeastern Bering Sea. Cont Shelf Res 25:1281–1301
Carroll ML, Carroll J (2003) The Arctic seas. In: Black KD, Shimmield GB (eds) Biogeochemistry of marine systems. CRC Press, Boca Raton, Florida, pp 127–156
Christiansen JS, Karamushko OV, Fevolden SE, Praebl K (2008) Marine fishes of NE Greenland—diversity and adaptation. ICES 2008/B:19, (http://www.ices.dk/products/CMdocs/CM-2008/B/B1908.pdf).
Christie HC, Jørgensen NM, Norderhaug KM, Waage-Nielsen E (2003) Species distribution and habitat exploitation of fauna associated with kelp (Laminaria hyperborea) along the Norwegian Coast. J Mar Biol Ass UK 83(4):687–699
Comiso JC (2006) Abrupt decline in the Arctic winter sea ice cover. Geophys Res Lett 33. doi:10.1029/2006GL027341.
Conlan KE, Kvitek RG (2005) Recolonization of soft-sediment ice scours on an exposed Arctic coast. Mar Ecol Prog Ser 286:21–42
Coyle KO, Bluhm BA, Konar B, Blanchard A, Highsmith RC (2007) Amphipod prey of gray whales in the northern Bering Sea: changes in biomass and distribution. Deep Sea Res II 54:2906–2918
Dahle S, Anisimova NA, Palerud R, Renaud PE, Pearson TH, Matishov GG (2009) Macrobenthic fauna of the Franz Josef Land archipelago. Pol Biol 32:169–180
Dickson RR, Meincke J, Vassie I, Jungclaus J, Østerhus S (1999) Possible predictability in overflow from the Denmark Strait. Nature 397:243–246
Drainville G (1970) The Saguenay fjord: the ichtyological fauna and ecological conditions. Nat Can 97:623–666
Drinkwater KF (2006) The regime shift of the 1920s and 1930s in the North Atlantic. Prog Oceanogr 68(2–4):134–151
Dunton K (1992) Arctic biogeography: the paradox of the marine benthic fauna and flora. Trends Ecol Evol 7:183–189
Dunton KH, Schell DM (1987) Dependence of consumers on macroalgal (Laminaria solidungula) carbon in an arctic kelp community: δ13C evidence. Mar Biol 93:615–625
Eilertsen HC, Taasen JP, Weslawski JM (1989) Phytoplankton studies in the fjords of West Spitsbergen: physical environment and production in spring and summer. J Plankton Res 11(6):1245–1260
Ellingsen I, Slagstad D, Sundfjord A (2009) Modification of water masses in the Barents Sea and its coupling to ice dynamics: a model study. Ocean Dyn 59:1095–1108
Ellis DV, Wilce RT (1961) Arctic and Subarctic examples of intertidal zonation. Arctic 14:224–235
Espinosa F, Guerra Garcia JM (2005) Algae, macrofaunal assemblages and temperature: a quantitative approach to intertidal ecosystems of Iceland. Helgol Mar Res 59:273–285
Findlay HS, Kendall MA, Spicer JI, Widdicombe S (2009) Future high CO2 in the intertidal may compromise adult barnacle Semibalanus balanoides survival and embryonic development rate. Mar Ecol Prog Ser 389:193–202
Frederich M, Sartoris F et al (2001) Distribution patterns of decapod crustaceans in polar areas: a result of magnesium regulation? Polar Biol 24:719–723
Freitas C, Kovacs KM, Ims RA, Fedak MA, Lydersen C (2009) Deep into the ice: over-wintering and habitat selection in male Atlantic walruses. Mar Ecol Prog Ser 375:247–261
Golikov AN, Averincev VG (1977) Biocenoses of the upper regions of the shelf of Franz Josef Land archipelago and some regularities of their distribution. Iss Fauny Moriej 14(22):5–54
Golikov AN, Scarlato OA, Averincev VG, Menshutkina TV, Novikov OK, Sheremetevsky AM (1990) Ecosystems of the New Siberian shoals their distribution and functioning. Issl Fauny Moriej 37(45):4–79
Gollasch S (2006) Overview on introduced aquatic species in European navigational and adjacent waters. Helgol Mar Res 60:84–89
Gradinger R, Friedrich C, Spindler M (1999) Abundance, biomass and composition of the sea ice biota of the Greenland Sea pack ice. Deep Sea Res 46:1457–1472
Grebmeier JM, Overland JE, Moore SE, Farley EV, Carmack EC, Cooper LW, Frey KE, Helle JH, McLaughlin FA, McNutt SL (2006) A major ecosystem shift in the northern Bering Sea. Science 311:1461–1464
Greene CH, Pershing AJ, Cronin TM, Ceci N (2008) Arctic climate change and its impacts on the ecology of the North. Ecology 89:S24–S38
Gurjanova EF (1951) Gammaridea of the seas of U.S.S.R. and adjacent waters. Fauna SSSR vol. 41:1–1031 [in Russian]
Gutt J (2001) On the direct impact of ice on marine benthic communities, a review. Polar Biol 24:553–564
Havas M, Rosseland BO (2004) Response of zooplankton, benthos and fish to acidification: an overview. Water Air Soil Poll 85:51–62
Hawkins SJ, Moore PJ, Burrows MT, Poloczanska E, Mieszkowska N, Herbert RJH, Jenkins SR, Thompson RC, Genner MJ, Southward AJ (2008) Complex interactions in a rapidly changing world: responses of rocky shore communities to recent climate change. Climate Res 37:123–133
Helmuth B, Mieszkowska N, Moore P, Hawkins SJ (2006) Living on the edge of two changing worlds: forecasting the responses of rocky intertidal ecosystems to climate change. Ann Rev Ecol Evol Syst 37:373–404
Hewitt J, Thrush S, Lohrer A, Townsend M (2009) A latent threat to biodiversity: consequences of a small-scale heterogeneity loss. Biodivers Conserv 19:1315–1323
Holte B, Gulliksen B (1998) Common macrofaunal dominant species in the sediments of some North Norwegian and Svalbard glacial fjords. Pol Biol 19:375–382
Hop H, Falk Petersen S, Svendsen H, Kwasniewski S, Pavlov V, Pavlova O, Soreide J (2006) Physical and biological characteristics of the pelagic system across Fram Strait to Kongsfjorden. Prog Oceanogr 71:182–231
Hunt GL, Stabeno P, Walters G, Sinclair E, Brodeur RD, Napp JM, Bond NA (2002) Climate change and control of the southeastern Bering Sea pelagic ecosystem. Deep Sea Res II 49:5821–5853
Huston M (1979) A general hypothesis of spceis diversity. Am Nat 113:81–101
Jewett SC, Clough LM, Blanchard AL, Ambrose WG, Feder HM, Hoberg MK, Whiting AV (2009) Nearshore macrobenthos of northern Kotzebue Sound Alaska, with reference to local sewage disposal. Polar Biol 32(11):1665–1680
Johanesson K, Johanesson B (1995) Dispersal and population expansion in a direct developing marine snail (Littorina saxatilis) following a severe population bottleneck. Hydrobiologia 309:173–180
Johannessen OM, Miles MW (2000) Arctic sea ice and climate change- will the ice disappear in this century? Sci Prog 83(3):209–222
Jonasson JP, Thorarinsdottir G, Eiriksson H, Solmundsson J, Marteinsdottir G (2007) Collapse of the fishery for Iceland scallop (Chlamys islandica) in Breidafjordur, West Iceland. ICES J Mar Sci 64:298–308
Kaczmarek H, Wlodarska- Kowalczuk M, Legezynska J, Zajaczkowski M (2005) Shallow sublittoral macrozoobenthos in Kongsfjord, West Spitsbergen, Svalbard. Pol Pol Res 26:137–155
Keck A, Wiktor J, Hapter R, Nilsen R (1999) Phytoplankton assemblages related to physical gradients in an Arctic glacier−fed fjord in summer. ICES J Mar Sci 56:194–202
Kedra M (2008) Interannual changes in the biodiversity of the Arctic soft bottom macrobenthos in Kongsfjorden, Spitsbergen. PhD thesis, Institute of Oceanology Polish Academy of Sciences, Sopot
Kedra M, Wlodarska-Kowalczuk M, Weslawski JM (2010) Decadal change in a macrobenthic soft bottom community structure in a high Arctic fjord (Kongsfjorden, Svalbard). Polar Biol 33:1–11
Kendall MA, Widdicombe S, Weslawski JM (2003) A multi-scale study of the biodiversity of the benthic infauna of the high latitude Kongsfjord, Svalbard. Polar Biol 26:383–388
Konar B, Iken K, Edwards M (2009) Depth stratified community zonation patterns on Gulf of Alaska rocky shores. Mar Ecol 30:63–73
Kuklinski P, Barnes DKA (2005) Microhabitat diversity of Svalbard Bryozoa. J Nat Hist 39:539–554
Maslowski W, Roman R, Kinney JC (2008) Effects of mesoscale eddies on the flow of the Alaskan Stream. J Geophys Res 113:C07036. doi:10.1029/2007JC004341
McConnaughey RA, Mier KL, Dew CB (2000) An examination of chronic trawling effects on soft-bottom benthos of the eastern Bering Sea. ICES J Mar Sci 57:1377–1388
McMahon KW, Ambrose WG, Johnson BJ, Sun MY, Lopez GR, Clough LM, Carroll ML (2006) Benthic community response to ice algae and phytoplankton in Ny Ålesund, Svalbard. Mar Ecol Prog Ser 310:1–14
Mieszkowska N, Hawkins SJ, Burrows MT, Kendall MA (2007) Long-term changes in the geographic distribution and population structures of Osilinus lineatus (Gastropoda: Trochidae) in Britain and Ireland. J Mar Biol Assoc UK 87:537–545
Mileikovsky SA (1968) Distribution of pelagic larvae of bottom invertebrates of the Norwegian and Barents Seas. Mar Biol 1:164–167
Moore PG (1977) Inorganic particulate suspensions in the sea and their efects on marine animals. Oceanogr Mar Biol Annu Rev 15:225–363
Motyka RJ, Hunter L, Echelmeyer K, Connor C (2003) Submarine melting at the terminus of a temperate tidewater glacier, LeConte Glacier, Alaska. Ann Glaciol 36:57–65
Orr JC, Fabry VJ, Aumont O, Bopp L, Doney SC, Feely RA, Gnanadesikan A, Gruber N, Ishida A, Joos F, Key RM, Lindsay K, Maier-Reimer E, Matear R, Monfray P, Mouchet A, Najjar RG, Plattner GK, Rodgers KB, Sabine CL, Sarmiento JL, Schlitzer R, Slater RD, Totterdell IJ, Weirig MF, Yamanaka Y, Yool A (2005) Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437:681–686
Palerud R, Gulliksen B, Brattegard T, Sneli A-J, Vader W (2004) The marine macro-organisms in Svalbard waters. In: Prestrud P, Strøm H, Goldman HV (eds) A catalogue of the terrestrial and marine animals of Svalbard. Norwegian Polar Institute, pp 1-137
Pearson TH, Rosenberg R (1978) Macrobenthic succession in relation to 615 organic enrichment and pollution of the marine environment. Oceanogr Mar Biol Ann Rev 16:229–311
Petersen GH (1962) The distribution of Balanus balanoides (L.) and Littorina saxatilis, Olivi var. Groenlandica, Mencke in Northern West Greenland. Meddel Gronl 159:8–39
Petersen GH, Curtis MA (1980) Differences in energy flow through major components of subarctic, temperate and tropical marine shelf ecosystems. Dana 1:53–64
Piepenburg D, Archambault P, Ambrose WG Jr, Blanchard A, Bluhm BA, Carroll ML, Conlan K, Cusson M, Feder HM, Grebmeier JM, Jewett SC, Lévesque M , Petryashev VV, Sejr MK, Sirenko B, Włodarska-Kowalczuk M (2010) Towards a pan-Arctic inventory of the species diversity of the macro- and megabenthic fauna of the Arctic shelf seas. Mar Biodiv. doi:10.1007/s12526-010-0059-7
Piwosz K, Walkusz W, Hapter R, Wieczorek P, Hop H, Wiktor J (2008) Comparison of productivity and phytoplankton in a warm (Kongsfjorden) and cold (Hornsund) Spitsbergen fjord in mid summer 2002. Polar Biol 32:549–559
Pope JG, Falk-Pedersen J, Jennings S, Rice JC, Gislason H, Daan N (2009) Honey, I cooled the cods: Modelling the effect of temperature on the structure of Boreal/Arctic fish ecosystems. Deep Sea Res II 56(21–22):2097–2107
Quijon PA, Snelgrove PVR (2005) Predation regulation of sedimentary faunal structure: potential effects of a fishery-induced switch in predators in a Newfoundland sub-Arctic fjord. Oecologia 144(1):125–136
Rabus BT, Echelmeyer KA (2002) Increase of 10 m ice temperature: climate warming or glacier thinning. J Glaciol 48:279–286
Rachold V, Are FE, Atkinson DE, Cherkasov G, Solomon SM (2004) Arctic coastal dynamics—an introduction. Geo Mar Lett 25:63–68
Ray GC, McCormick-Ray J, Berg P, Epstein HE (2006) Pacific walrus: benthic bioturbator of Beringia. J Exp Mar Biol Ecol 330:403–419
Reise K, Simon M, Herre E (2001) Density-dependent recruitment after winter disturbance on tidal flats by the lugworm Arenicola marina. Helgol Mar Res 55:161–165
Renaud PE, Wlodarska-Kowalczuk TH, Holte B, Weslawski JM, Cochrane S, Dahle S, Gulliksen B (2007) Multidecadal stability of benthic community structure in a high Arctic glacial fjord (van Mijenfjord, Spitsbergen). Polar Biol 30:295–305
Renaud PE, Carroll ML, Ambrose WG Jr (2008a) Effects of global warming on Arctic sea-floor communities and its consequences for higher trophic levels. In: Duarte C (ed) Impacts of global warming on polar ecosystems. Bilbao, Fundacion BBVA
Renaud PE, Morata N, Carroll ML, Denisenko SG, Reigstad M (2008b) Pelagic-benthic coupling in the western Barents Sea: processes and time scales. Deep Sea Res II 55(20–21):2372–2380
Rhoads DC, McCall PL, Yingst JY (1978) Disturbance and production on the estuarine seafloor. Am Sci 66:577–586
Rumohr H, Kujawski T (2000) The impact of trawl fishery on the epifauna of the southern North Sea. ICES J Mar Sci 57(5):1389–1394
Schmid MK, Piepenburg D (1993) The benthos zonation of the Disko Fjord, West Greenland. Medd Grønl Biosci 37:3–21
Scrosati R, Eckersley LK (2007) Thermal insulation of the intertidal zone by the ice foot. J Sea Res 58:331–334
Sejr M, Christensen PB (2007) Growth production and carbon demand of macrofauna in Young Sound with special emphasis on the bivalves Hiatella arctica and Mya truncata. Medd Grønl Biosci 58:122–135
Sejr MK, Nielsen TG, Rysgaard S, Sturluson M, Risgard-Petersen N, Blicher M (2007) Marine production and sedimentation during spring and autumn in Disko Bay, Greenland. Mar Ecol Prog Ser 341:75–88
Sejr MK, Blicher ME, Rysgaard S (2009) Sea ice cover affects inter-annual and geographic variation in growth of the Arctic cockle Clinocardium ciliatum (Bivalvia) in Greenland. Mar Ecol Prog Ser 389:149–158
Sejr MK, Włodarska-Kowalczuk M, Legeżyńska J, Blicher ME (2010) Macrobenthic species composition and diversity in the Godthaabsfjord system, SW Greenland. Polar Biol 33(4):421–431
Sirenko BI ed. (2001) List of species of free-living invertebrates of Eurasian Arctic Seas and adjacent deep waters. Explorations of the fauna of the seas (Zoological Institute RAS, St. Petersburg) 51(59): 1–76
Sirenko BI, Gagaev SY (2007) Unusual abundance of macrobenthos and Pacific species invasions into the Chukchi Sea. Biologya Morya 33(6):399–407
Sneli JA (1968) The intertidal distribution of polychaetes and molluscs on a muddy shore in Nord-Möre. Sarsia 31:63–68
Stempniewicz L, Blachowiak-Samolyk K, Weslawski JM (2007) Impact of climate change on zooplankton communities, seabird populations and Arctic terrestrial ecosystem—a scenario. Deep Sea Res II 54:2934–2433
Sun MY, Carroll ML, Ambrose WG, Clough LM, Zou L, Lopez GR (2007) Rapid consumption of phytoplankton and ice algae by Arctic soft-sediment benthic communities: Evidence using natural and C-13-labeled food materials. J Mar Res 65(4):561–588
Sun MY, Clough LM, Carroll ML, Dai JH, Ambrose WG, Lopez GR (2009) Different responses of two common Arctic macrobenthic species (Macoma balthica and Monoporeia affinis) to phytoplankton and ice algae: Will climate change impacts be species specific? J Exp Mar Bio Ecol 376(2):110–121
Svendsen H, Beszczynska-Moller A, Hagen JO, Lefauconnier B, Tverberg V, Gerland S, Orbaek JB, Bischof K, Papucci C, Zajaczkowski M, Azzolini R, Bruland O, Wiencke C, Winther JG, Dallmann W (2002) The physical environment of Kongsfjorden–Krossfjorden, an Arctic fjord system in Svalbard. Polar Res 21:133–166
Syvitski JPM, Burell CD, Skei JM (1987) Fjords: processes and products. Springer, New York
Syvitski JPM, Farrow GE, Atkinson RJA, Moore PG, Andrews JT (1989) Baffin Island fjord macrobenthos: bottom communities and environmental significance. Arctic 42:232–247
Vader W (1980) A record of Geryon tridens (Crustacea Decapoda) from the Barents Sea. Fauna Norv A 1(1):46
Vader W, Johnsen JR (2000) Schisturella pulchra (Hansen) (Lysianassoidea), a new amphipod for the Barents Sea area. Polskie Archiwum Hydrobiologii 47(3-4):489-494
Walczowski W, Piechura J (2006) New evidence of warming propagating toward the Arctic Ocean. Geoph Res Lett 33:L1 2601
Walczowski W, Piechura J (2007) Pathways of the Greenland Sea warming. Geoph Res Lett 34:L1 0608
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:389–395
Wassmann P, Reigstad M, Haug T, Rudels B, Carroll ML, Hop H, Gabrielsen GW, Falk-Petersen S, Denisenko SG, Arashkevich E, Slagstad D, Pavlova O (2006) Food webs andcarbon flux in the Barents Sea. Prog Oceanogr 71:232–287
Wassmann P, Duarte CM, Agusti S, Sejr MK (2010) Footprints of climate change in the Arctic marine ecosystem. Glob Change Biol. doi:10.1111/j.1365.2486.2010.02311.x
Weslawski JM (1990) Distribution and ecology of coastal waters amphipods from Southern Spitsbergen. Pol Arch Hydrobiologii 37:503–519
Weslawski JM, Zajaczkowski M(1992) Benthic fauna and its environment in Tikhaia Bay, Hooker Island. Norsk Polarinstitutt Medd 120, Oslo, pp 35- 42
Weslawski JM, Szymelfenig M (1999) Community composition of tidal flats on Spitsbergen: consequence of disturbance? In: Gray JS, Ambrose W, Szaniawska A (eds) Biogeochemical cycling and sediment ecology. Kluver, Dordrecht Boston London, pp 185–193
Weslawski JM, Kwaśniewski S, Wiktor J (1991) Winter in a Svalbard fjord ecosystem. Arctic 44:115–123
Weslawski JM, Zajaczkowski M, Wiktor J, Szymelfenig M (1997) Intertidal zone of Svalbard. 3 Littoral of a subarctic, oceanic island: Bjornoya. Polar Biol 18:45–52
Weslawski JM, Wiktor J Jr, Kotwicki L (2010) Increase in biodiversity in the arctic rocky littoral, Sorkappland, Svalbard, after 20 years of climate warming. Mar Biodiv 40:123–130
Widdicombe S, Spicer JI (2008) Predicting the impact of ocean acidification on benthic biodiversity: What can animal physiology tell us? J Exp Mar Biol Ecol 366(1–2):187–197
Wiktor J, Wojciechowska K (2005) Differences in taxonomic composition of summer phytoplankton in two fjords of West Spitsbergen, Svalbard. Pol Polar Res 26(4):259–268
Witman JD, Cusson M, Archambault P, Pershing AJ, Mieskowska N (2008) The relation between productivity and species diversity in temperate-arctic marine ecosystems. Ecology 89:S66–S80
Wlodarska M, Weslawski JM, Gromisz S (1996) A comparison of the macrofaunal community structure and diversity in two arctic glacial bays—a ‘cold’ one off Franz Josef Land and a ‘warm’ one off Spitsbergen. Oceanologia 38:251–283
Wlodarska-Kowalczuk M, Weslawski JM (2001) Impact of climate warming on Arctic benthic biodiversity: a case study of two Arctic glacial bays. Climate Res 18:127–132
Wlodarska-Kowalczuk M, Weslawski JM (2008) Mesoscale spatial structures of soft bottom macrozoobenthos communities: effects of physical control and impoverishment. Mar Ecol Prog Ser 35:215–224
Wlodarska-Kowalczuk M, Pearson T, Kendall MA (2005) Benthic response to chronic natural physical disturbance by glacial sedimentation in an Arctic fjord. Mar Ecol Prog Ser 303:31–41
Włodarska-Kowalczuk M, Kuklinski P, Ronowicz M, Legeżyńska J, Gromisz S (2009) Assessing species richness of macrofauna associated with macroalgae in Arctic kelp forests (Hornsund, Svalbard). Polar Biol 32:897–905
Zajaczkowski M, Nygard H, Hegseth EN, Berge J (2010) Vertical flux of particulate matter in an Arctic fjord: the case of lack of the sea ice cover in Adventfjorden 2006–2007. Polar Biol 33:223–239
Zenkevitch LA (1963) Biology of the seas of the U.S.S.R. Allen and Unwin, London
Acknowledgements
The authors are indebted to three anonymous reviewers that contributed greatly to the shaping of the text. We are also acknowledging the projects that supported our research: Arctic Ocean Diversity (CoML) and ArcEco (MARBEF EU network of excellence). This publication is part of the Census of Marine Life’s Arctic Ocean Diversity project synthesis and was originally presented at the Arctic Frontiers Conference in Tromsø, January 2010. The support and initiative of ARCTOS and Arctic Frontiers are gratefully acknowledged.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Węsławski, J., Kendall, M., Włodarska-Kowalczuk, M. et al. Climate change effects on Arctic fjord and coastal macrobenthic diversity—observations and predictions. Mar Biodiv 41, 71–85 (2011). https://doi.org/10.1007/s12526-010-0073-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12526-010-0073-9