
Abstract
Effective management of eutrophication in tidal ecosystems requires a thorough understanding of the dynamics of their responses to decreases in nutrient loading. We analyze a 34-year dataset on a shallow embayment of the tidal freshwater Potomac River, Gunston Cove, for long-term responses of ambient nutrient levels, light transparency measures, phytoplankton biomass, and coverage of submersed aquatic vegetation (SAV) to decreased nutrient loading. Point source loading of phosphorus, the nutrient most limiting primary production in this system, was greatly curtailed coincident with the study onset (1983/84) exhibiting a sharp decrease of 95% from peak loading levels. However, water column total phosphorus decreased much more slowly and gradually. Phytoplankton chlorophyll a did not show a distinctive decrease until 2000 and SAV responded strongly beginning in 2004. The habitat suitability model for SAV developed by Chesapeake Bay researchers was able to explain the recovery of SAV coverage based on data on light transparency and basin morphometry collected in this study. The study results were consistent with the alternative stable state theory with a sharp transition from a phytoplankton-dominated “turbid water” state to an SAV-dominated “clear water” state in a 2-year period from 2003 to 2005. The system eventually responded to nutrient load reductions, but the nonlinear and incomplete nature of this recovery and the two-decade delay illustrate the complexities of managing these systems.
Similar content being viewed by others


Introduction
Tidal freshwater systems occur in the landward reaches of estuaries where the rate of freshwater inflow is greater than the rate of mixing with brackish water from further down in the estuary, allowing salinities to remain less than 0.5 S. Drowned river valleys such as the tidal Potomac River (Schubel and Pritchard 1987) are conducive to substantial stretches of tidal freshwater due to their elongate nature. Odum et al. (1984) were among the first to highlight the distinctive nature and location of these systems. As they pointed out, these systems are found worldwide, typically associated with river mouths which provide the consistent input of freshwater necessary in an aquatic system at sea level to counter the intrusion of brackish water from the adjacent estuary.
In the absence of excessive nutrients associated with eutrophication, shallow freshwater ecosystems are generally dominated by submersed aquatic vegetation (SAV). Reviews of pre-eutrophication conditions in representative systems (Haramis and Carter 1983; Osborne and Moss 1977; Moss 1980) conclude that these systems contained lush and diverse growths of SAV. Specifically, tidal freshwater systems are notable for the taxonomic diversity of their vegetation when compared with shallow estuarine and marine systems (Odum 1988). Furthermore, SAV beds have great value as habitat for macroinvertebrates, fish, and waterfowl (Thorp et al. 1997; Stewart 1962). SAV promotes water clarity by increasing particle sedimentation including open-water–produced phytoplankton (Kemp et al. 1984). SAV and associated periphyton may also decrease dissolved nutrient concentrations during their growth period, although differing results have been found in various systems and as a function of season (Carpenter 1980; Landers 1982; Rorslett et al. 1986).
Shallow freshwater ecosystems are often subjected to increased nutrient loading and this is particularly true of tidal freshwater systems. Their location just downstream from the head of tide places them at a crucial nexus of navigation and industrial development. Cities have sprung up at this upstream point of oceanic navigation which corresponds to the Fall Line on the Atlantic coast of the USA. In the mid-Atlantic region, human population centers grew into major metropolitan areas including Richmond, Washington, Baltimore, and Philadelphia. Prior to the very recent advent of advanced sewage treatment, these urban areas contributed large amounts of pollutants including the plant nutrients phosphorus and nitrogen to tidal fresh receiving waters, resulting in extreme cultural eutrophication. Tidal freshwaters were also the loci at which point and nonpoint loadings from the watershed encountered aquatic ecosystems with longer residence times promoting eutrophication effects such as algal blooms.
The watershed of the tidal Potomac River was settled by European colonists during the late 1600s and early 1700s and its landscape and waterscape have been modified in various ways through several stages of development (Brush 2009). Early land clearing during the colonial and early national period brought sediments into the tidal river and rendered many formerly bustling colonial ports like Dumfries (Berkley 1924) and Port Tobacco (Gottschalk 1945) impassable, but these sediments apparently had only localized effects on SAV. Abundant evidence indicates that submersed macrophytes blanketed the shallow channel margins and embayments of the tidal Potomac in the late nineteenth century and the early part of the twentieth century (Carter et al. 1985; Carter and Rybicki 1986; Haramis and Carter 1983). Native species such as Vallisneria americana (water-celery), Ceratophyllum demersum (coontail), Najas flexilis (northern naiad), Potamogeton crispus (curly pondweed), and Anacharis alcinastrum (old world elodea) were among the species reported as abundant in this period. However, by the mid-twentieth century, these SAV beds had been drastically reduced and a 3-year study culminating in 1980 found aquatic macrophytes virtually absent from the tidal freshwater Potomac due to increasing eutrophication (Haramis and Carter 1983).
This eutrophication was the product of the burgeoning growth of human populations in the lands adjacent to the tidal freshwater Potomac. Population in the Washington, D.C., metropolitan area grew from about 670,000 in 1930 to over 2.5 million by 1970 (https://population.us/). Corresponding flows from wastewater treatment plants increased from 80 MGD in 1930 to 300 MG in 1970 (Jaworski et al. 2007). With this was a concomitant increase in human waste being discharged into sewers and making its way into the river after various levels of treatment. Early on, efforts were mainly directed at removing solids and dissolved organics (BOD) from the sewage and decreasing the possibility of waterborne fecal diseases with chlorination.
While these efforts resulted in major improvements in water quality and habitat in the river (Jaworski et al. 2007), it was clear that additional pollutants in the sewage were causing major problems. Studies elsewhere have documented that phosphorus inputs from municipal wastewater fuel the growth of suspended algae, the phytoplankton, and in the process increase the turbidity of the water (Larsen et al. 1975; Edmondson 1970). Data on inputs to the tidal freshwater Potomac River indicate a dramatic increase in phosphorus from wastewater treatment plants beginning in the late 1940s (Jaworski et al. 2007; Walker et al. 2000). By 1965, total phosphorus concentrations in the tidal freshwater Potomac regularly exceeded 500 μg/L indicating highly eutrophic conditions and phytoplankton responded with frequent chlorophyll a concentrations above 100 μg/L reflecting bloom conditions (Jaworski et al. 2007; U.S. EPA 1970a; 1970b). In addition, high concentrations of cyanobacteria were the summertime norm in the tidal freshwater Potomac. For 1970, when about 40 readings were available in Gunston Cove, median values reported were Secchi depth 0.29 m, Chl-a 110 μg/L, TP 0.77 mg/L, and TN 2.0 mg/L (U.S. EPA 1970a, 1970b).
Based on the strong evidence from these studies, EPA and the states went to work to reduce phosphorus loading. A phosphorus detergent ban was implemented and strict limits were placed on phosphorus concentrations in sewage effluent. These measures resulted in a dramatic decline in phosphorus inputs to the tidal Potomac to levels not seen since 1900 (Jaworski et al. 2007; Walker et al. 2000). Unfortunately, the blooms continued and a particularly strong bloom in 1983 resulted in a major effort to understand the cause the continued blooms despite a 90% reduction in phosphorus loading.
An expert panel was convened and studied possible explanations for the continued blooms. They determined that an additional source of phosphorus was needed to explain the blooms (Thomann et al. 1985). Previous analyses and models had discounted the possibility of sediment release of phosphorus into the water column of the tidal Potomac because the system does not stratify owing to strong tidal currents. Therefore, the anoxic release of phosphorus from sediments could not occur. However, managers had not been aware of studies that indicated enhanced phosphorus release under oxic conditions when pH rises above 9.5 (Andersen 1975). Studies were commissioned on tidal freshwater Potomac cores and the enhanced release under oxic conditions at high pH was validated for the tidal Potomac River (Seitzinger 1991).
Further reductions in phosphorus from wastewater treatment plants were not feasible and blooms continued to recur throughout the 1980s. These blooms, every year from 1985 to 1989, were similar to the 1983 bloom being characterized by Microcystis aeruginosa as the dominant cyanobacterial species, chlorophyll a concentrations well above 200 μg/L over substantial portions of the river, and pH levels well above 9.5 (Jones 1991). However, moving into the 1990s, bloom frequencies and intensities abated indicating that sediment phosphorus was becoming less accessible to the phytoplankton either due to depletion or burial.
Meanwhile, SAV recolonization began with an unexpected invasion of the exotic species Hydrilla verticillata which was accidentally introduced into the tidal Potomac River and the recovery of some native species and earlier invading species such that by 1985 most of the shallow (< 2 m) areas in the upper freshwater tidal Potomac (above Gunston Cove) had a modest cover of SAV (Carter and Rybicki 1986). As time progressed through the 1990s and 2000s, SAV colonization continued, and despite some setbacks, by the mid-2000s, much of the shallow areas of the tidal freshwater Potomac were well-colonized with SAV (https://www.vims.edu/bio/sav). While H. verticillata led the recolonization and continues to be an important contributor to the SAV community, other species such as Vallisneria americana, Ceratophyllum demersum, Najas minor, and Myriophyllum spicatum are important and sometimes quite abundant in the tidal freshwater Potomac in general and Gunston Cove in particular. A timeline of significant water quality events in the Potomac River basin is found at https://www.potomacriver.org/potomac-basin-facts/potomac-timeline/
Restoration measures attempt to reverse the degradation process with the goal of returning the ecosystem to its pre-impact configuration. Numerous studies have found that this process is often far from linear and that response lag, or hysteresis, is commonly observed. Two states have been recognized for shallow aquatic systems: one lower nutrient “clear water” state dominated by SAV and another higher nutrient “turbid water” state dominated by phytoplankton. Investigators have suggested that shallow water systems are not arrayed equally across the light availability spectrum, but are drawn to one of these two end states (Scheffer et al. 1993). Originally identified in shallow lakes, this phenomenon is now being observed in shallow estuarine systems as well and has been termed “alternative stable states” (Duarte et al. 2009). Investigators are addressing the question: “once the ecosystem has been degraded to the “turbid water” stable state, how long and under what conditions will it return to its original “clear water” stable state?”
Investigators have also questioned whether the basic underlying characteristics of the ecosystem have shifted so much that a return to identical pre-existing conditions is even possible (Duarte et al. 2009). This phenomenon is called “shifting base lines.” Thus, major considerations for evaluation of restoration programs include the time required for the system to recover and the extent to which recovery may be complete and allow return to the “clear water” state.
As part of the Chesapeake Bay restoration effort, bay scientists developed a habitat suitability model to allow prediction of SAV colonization depths based on light attenuation factors (Dennison et al. 1993; Kemp et al. 2004). The model incorporates light attenuation by phytoplankton, total suspended solids, color, and epiphytes as well as observed light requirements of SAV species in each salinity regime of the bay including tidal freshwater. The model was then calibrated to empirical observations of SAV occurrence to derive threshold values of light required to reach SAV to initiate and sustain growth (as percent of surface light). This model can then be used to calculate the depth to which SAV can successfully grow given observed light penetration.
Given the observed lags (hysteresis) in many systems recovering from pollution in general and eutrophication in particular, long-term data sets provide a unique opportunity to test these concepts of “multiple stable states” and “shifting baselines.” In this paper, we examine the process of recovery in Gunston Cove, an embayment of the tidal freshwater Potomac River in Fairfax County, Virginia. Beginning with laboratory analyses of nutrient data in 1983 and continuing with a full suite of water quality and biological data collected typically semi-monthly beginning in 1984, a long-term record of the Gunston Cove ecosystem has been compiled. Since this long-term study was initiated roughly one decade after the point of maximum P loading, we are unable to completely characterize onset of the fully degraded, turbid water state, but as this paper will show, the study area clearly remained in the “turbid-water” phytoplankton-dominated state for a number of years after the study began.
The goals of this paper are four-fold. One, I will subject the consistent data record from the early 1980s through 2017 to statistical and trend analysis to determine the magnitude, consistency, and statistical significance of long-term trends in water quality parameters and ascertain the magnitude and consistency of changes during that period. Two, we will analyze changes in the underwater light environment in the cove and make predictions of SAV recovery in the cove based on the Chesapeake Bay Program habitat suitability model which bases SAV recovery on improved water clarity. Three, we will test these predictions against interannual changes in the SAV distribution as observed by aerial mapping. And finally, we will interpret our results in context of the discussion on stable states, hysteresis, shifting baselines, and recovery/restoration targets for managers.
Study Site
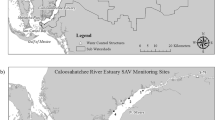
The study site is the Gunston Cove embayment of the tidal freshwater Potomac River, located about 20 km downstream of Washington, D.C., USA (Fig. 1). The tidal Potomac River is one of the main subestuaries of the Chesapeake Bay system. Due to substantial inflow of freshwater into the tidal Potomac, freshwater conditions (< 0.5 S) are maintained through Quantico, about 23 km downstream from the study site. Semidiurnal tidal amplitude is about 0.6 m. Bricker et al. (2014) provide a summary of the morphology and tidal characteristics of the tidal Potomac River and a thorough overview of the watershed.

Gunston Cove showing location of Station 7 (starred), the location of all data reported in this paper. Inset shows relationship of Gunston Cove to the tidal Potomac River
Gunston Cove is a shallow embayment (zmax ≈ 2 m) with a maximum length of 4 km and an average width of about 1.25 km resulting in an area of approximately 5 km2. The Cove is formed by the confluence of Pohick and Accotink creeks which are also tidal in their lower reaches. The two creeks together drain 21,940 ha of suburban Fairfax County, Virginia, with Accotink having a slightly larger drainage area. Average monthly residence time of water in the cove varies from 17 to 34 days based in gauged inflows and measured cove volume (Harper 1988). Further background information about Gunston Cove and its watershed may be found in Jones et al. (2008).
The Noman M. Cole, Jr. Pollution Control Plant discharges into Pohick Creek 1.8 km above the head of tide. The plant was originally placed in operation in 1970. The effluent discharge rate was initially about 20 megaliters per day and gradually increased to about 170 megaliters per day by 1998 and has remained fairly stable since (Fig. 2). Loading of phosphorus reached a maximum of 480 kg/day in 1974, but declined markedly in the late 1970s and early 1980s to less than 17 kg/day by 1984 where it remained through the rest of the study period. This strong reduction was accomplished by specific changes in treatment processes and a basin-wide phosphate detergent ban (https://www.potomacriver.org/potomac-basin-facts/potomac-timeline/). Nitrogen loads remained high through 2002; subsequent implementation of N removal at Noman Cole dropped loads to less than 100 kg/day by 2005 and they have continued to decline gradually reaching 30 kg/day by 2015.

Effluent flow (megaliters per day), total P loading (kilograms per day), and total N loading (tens of kilograms per day) at Noman M. Cole Jr. Pollution Control Plant since the plant came on line in 1970. Data courtesy of Fairfax County Department of Public Works and Environmental Services
This area of the tidal Potomac River has been the site of a long-term ecological monitoring study since 1984. While the river channel area receives large loads of nutrients upstream of the study site, previous studies have determined that little of this load penetrates beyond the outer cove area. During the period 1984 to 1990, a spatially and temporally extensive sampling program was carried out in the cove and nearby Potomac mainstem (Jones et al. 2008). Sampling was conducted at as many as 15 sites in the tidal river and nearby Gunston Cove during this period. A recent analysis of spatial characteristics of these water quality data found that sites grouped into two regions: (1) the mainstem area including the Potomac River channel and its flanks and (2) the embayment area consisting of Gunston Cove (Jones et al. 2008). Using both univariate and multivariate analysis of water quality, it was determined that data from a single station in each area could be used to characterize each region. Dye studies and hydrological modeling (Harper 1988) corroborated the distinct nature of the cove water mass.
In this paper, we examine long-term trends in the Gunston Cove embayment characterized by Station 7 located in the middle of Gunston Cove with a water depth of about 2 m (38.676° N, 77.157° W). Tidal current velocities at this site average 5–8 cm/s, much reduced from the average of 23–34 cm/s observed in the river mainstem area (Kelso et al. 1985). With negligible salinity and the shallow depth, stratification is minor and temporary. Gunston Cove has been the site of extensive scientific studies which are documented in annual reports, published journal articles, theses and dissertations, a list of which can be found at https://cos.gmu.edu/perec/our-research/gunston-cove-study/gunston-cove-publications/#.XU4dgvZFxhE
Methods
Two water quality data sets were available for the Gunston Cove site for the period 1983–2017. One data set (GMU) resulted from monthly to semimonthly collections of field parameters like temperature (T), specific conductance (SPC), dissolved oxygen (DOppm), percent saturation DO (DOsat), field pH, Secchi depth (SD), and light attenuation coefficient (Kd) as well as depth-integrated and surface water quality samples analyzed for chlorophyll a (Chl-a DI and Chl-a Sf, respectively) collected by George Mason University personnel. Total suspended solids (TSS) and volatile suspended solids (VSS) have been collected since 1993 by GMU and will be used in examination of light attenuation factors. A second data set (FC) resulted from monthly to semimonthly sampling of near surface (z = 0.3 m) and near bottom (z = 0.5 m above the bottom) waters for standard lab water quality parameters including chloride (Cl−), lab pH, alkalinity (ALK), TSS, VSS, biochemical oxygen demand (BOD), total phosphorus (TP), soluble reactive phosphorus (SRP), total Kjeldahl nitrogen (TKN), ammonia nitrogen (NH3N), nitrate nitrogen (NO3N), and nitrite nitrogen (NO2N). Organic nitrogen (ON) was calculated as the difference between TKN and NH3N. N to P ratio was calculated as (TKN+ NO3N+ NO2N)/TP. The FC samples were collected by either George Mason University or Fairfax County personnel and were analyzed by the Fairfax County Environmental Services Laboratory. Laboratory analysis followed standard methodologies and has been further documented in a previous publication (Jones et al. 2008).
Extensive depth profile sampling indicates that, in general, the water column in the cove undergoes minor diel stratification. This daytime stratification is mainly reflected in some of the GMU parameters such as T, DOsat, DOppm, and field pH. Due to its minimal nature, depth-integrated values of these parameters were used in all analyses. Preliminary analyses of the FC data set showed that while there was generally little consistent difference between surface and bottom samples in the FC parameters, on occasion the bottom samples showed substantially greater amounts of suspended solids relative to the surface samples, probably due to some resuspension of bottom sediments during sampling. This tended to skew some of the relationships and so the analyses on the FC parameters in this paper were done with the surface samples only.
Analysis of long-term trends focused on the months of May through September. These months have been subjected to the greatest frequency of sampling, typically semimonthly, through the course of the study. They are also the months of greatest productivity by both phytoplankton and submersed aquatic vegetation (SAV) in the study area.
Long-term trends were assessed both graphically and statistically. Data were aggregated by year and scatterplots were constructed graphing year vs. each water quality parameter. A LOWESS smoother was applied to create a trend line using a tension of 0.5. This aided in visualizing the data and discriminating nonlinear trends. Linear regressions were also performed for each parameter vs. year to ascertain any statistically significant linear patterns over the entire period of record. While some variables were more nearly log-normally distributed, untransformed data were used in the linear regressions to allow easier interpretation of significant regression coefficients.
As a second approach to temporal data analysis and one that may be more consistent with the theory of alternate stable states and the possibility of hysteresis, the 34-year data record was binned into two intervals that correspond roughly to trend breakpoints in chlorophyll and Secchi disk transparency: Period 1: 1984–2003; period 2: 2005–2017. Means were compiled for each parameter during these periods and tests were done to establish significant differences in individual parameters between the periods.
Multiple regression was used to analyze the relative contribution of TSS and Chl-a to Kd values. This analysis relied on the GMU data set as all three of these parameters were measured concurrently on GMU cruises.
Extensive research and synthesis efforts have resulted in establishing that in the tidal freshwater portions of the Chesapeake Bay (including the tidal freshwater Potomac River), 9–13% of incident light must reach the bottom during the growing season to allow growth of submerged aquatic vegetation at a particular site (Kemp et al. 2004). The extensive data set of Kd and SD measurements in this study facilitated calculation of the maximum depth of SAV colonization for a substantial part of the growing season during each year. Applying the Beer-Bouguer law, the maximum depth of SAV colonization (ZSAV) was obtained on each date from the equation ZSAV = ln 0.09/Kd = 2.41/Kd. This assumes that 9% of incident light is the minimum required for SAV growth. Kd was available for most sample dates for the period 1991–2017, but SD was available for the entire period and on virtually all sample dates. To create the most robust and temporally extensive dataset of ZSAV, a regression was conducted to relate Kd to 1/SD and then Kd was calculated for all sampling dates. The ZSAV results were compared with SAV areal coverage data from VIMS (http://web.vims.edu/bio/sav/) and cove bathymetry to determine the degree to which SAV recovery could be explained.
Results
Water Quality Trends and Relationships: Entire Period of Record
Linear regression analysis revealed a host of significant long-term trends in Gunston Cove water quality parameters (Table 1). In the GMU dataset SPC, field pH, SD, Kd, Chl-a DI, Chl-a Sf, TSS, VSS, and Chl-a/TSS showed statistically significant correlation coefficients and linear trends over the 34-year period (Table 1). Trends in light transparency, Chl-a, and suspended solid parameters were highly significant (P < 0.0001). SPC and field pH displayed weaker, but still significant patterns. Dissolved oxygen did not display a significant linear trend over time.
Examination of the scatterplots (Fig. 3) reinforced the relative strength of time trend trajectories among the parameters. SPC exhibited a steady, but very gradual rise, yielding an increase of 63 μS/cm over the period. SD and Kd were essentially constant or showed minor oscillations until about 1998. At that point, a marked change in both variables was observed indicating a clear improving trend in water clarity through the remainder of the study period. The trend line for SD showed a little more than a doubling over the period from about 0.4 m to about 0.8 m. For Kd, the trend line decreased from 3.7 to 2.0 m−1. The trend line for Chl-a levels followed trends similar to those in water clarity but shifted by a few years. The trend line was fairly flat from 1984 to 2002, although there were some years above and below that line which was generally in the 80–100 μg/L range. Then beginning in about 2002, after a 4-year dry period marked with elevated Chl-a concentrations, a strong decline in Chl-a was observed resulting in the trend line dropping to less than 20 μg/L in both surface and depth-integrated samples by 2017. Many readings were below 10 μg/L and only three above 60 μg/L. TSS from the GMU dataset showed a fairly steady decline over the period of measurement with the steepest drop from 1992 to 2005. The Chl-a/TSS ratio was fairly constant from 1992 to 1998, increased noticeably between 1999 and 2002, and then decreased strongly from 2003 to 2017 as Chl-a declined and TSS stabilized.

Scatterplots of GMU parameters that were significantly correlated with year
Correlation analysis of GMU data (Table 2) revealed that the highest correlations were for the SD-Kd pair and for the two slightly different Chl-a measurements. These were expected as in each case the two variables are just alternate measures of water clarity and algal density, respectively. The next highest group of correlations were between the water clarity indicators and Chl-a and TSS. These results are consistent with the expectation that phytoplankton and inorganic particles are major absorbers of light in the study area. Neither TSS nor VSS represents solely inorganic particles as algae also contribute to both. Also, the Chl-a/TSS ratio was more highly correlated with variations in chlorophyll a than those in TSS. Lesser, but still significant, correlations were found between chlorophyll measures and indicators of photosynthesis such as field pH and DO indicating that phytoplankton algae were major components of primary production affecting pH and DO levels.
Almost all of the water quality parameters measured by the Fairfax County Environmental Services Laboratory (FC) showed very significant temporal trends over the study period via linear regression (Table 3). Only soluble reactive phosphorus (SRP) showed a marginally significant trend (P = 0.0045). All of these regression coefficients were negative indicating a decrease in these parameters over the period.
Examination of the FC scatterplots revealed several distinct patterns (Fig. 4). One pattern was a gradual and fairly consistent decline which started slowly in the first decade of the study and then increased in slope from about 1995–2000 onward. This was the pattern observed for TSS, VSS, ON, and TP. Correlation analysis revealed that these parameters were very closely related to one another with correlation coefficients between 0.729 and 0.861 (Table 4). A second pattern was exhibited by BOD and ammonia nitrogen in which concentrations increased markedly during the first 6–7 years and then declined consistently through the remainder of the study (Fig. 5). BOD showed a strong correlation with the parameters in the first pattern (Table 4). NO3N showed a slow steady decline, but was not strongly correlated with any of the other parameters except N to P ratio and NO2N. N to P ratio showed little trend until 2000 after which it declined slowly but remained well within the range expected for P-limited conditions.

Scatterplots of Fairfax County Lab parameters that were significantly correlated with year. Parameters showing continuous decline

Scatterplots of Fairfax County Lab parameters that were significantly correlated with year. Parameters showing different patterns
Water Quality Trends and Relationships: Turbid Water State vs. Clear Water State
Following up on the hypothesis that shallow aquatic systems possess stable states which may show lagged step function switching responses, the data were divided into two periods with the break occurring in 2004 when the study area exhibited a rapid transition from a phytoplankton-dominated, “turbid water” state (period I) to an SAV-dominated “clear water” state (period II).
To assess this pattern, the measured parameters were binned by period; then means were calculated and parameter means by period were compared. All of the particle and light transparency parameters measured by GMU (Table 5) exhibited a strong pattern with each variable showing significant differences between periods. Average Chl-a decreased by 75% from about 90 μg/L to about 23 μg/L. Average TSS was down 33% from 23.2 to 15.2 mg/L and VSS was down 22%. The stronger decrease in chlorophyll compared with TSS is also shown by their ratio which declined by 53.4% between the two periods. SD increased from an average of 0.448 m in period I to 0.768 m in period II and Kd showed a similar change, both reflecting substantially greater water clarity. In contrast, there was little to no change in indicators of photosynthesis, pH, and DOsat. Perhaps the decrease in phytoplankton photosynthesis was offset by an increase in SAV photosynthesis.
All of the FC Lab water quality parameters exhibited significant differences between the periods (Table 6). Many of them showed a decline of just over 50% from period I to period II: TP, ON, NO3N, BOD, TSS, and VSS. Chl-a Sf and SD, which were assayed less frequently by Fairfax County than by GMU, showed very similar changes from period I to period II as that observed in GMU data. Two variables, NH3N and NO2N, showed very strong declines which can be explained by the onset of N removal from the Noman Cole plant effluent at about the time of the transition from period I to period II.
Light Environment and Particle Concentrations
A regression approach was used to further investigate the relationship between particles and light transparency properties (Table 7). Each particle parameter was taken as an independent variable and used to predict each of the two light transparency parameters. Kd is considered the fundamental light transparency property and SD is used as an empirical way to easily measure it. Work has shown that inverse SD is a better linear predictor of Kd than untransformed values (Table 2 vs. Table 7). There was a very high Pearson correlation between Kd and 1/SD (r = 0.896) and a highly significant linear regression coefficient (Fig. 6). Across the board, each of the particle parameters performed well in predicting the light transparency parameters. Fixed suspended solids (FSS, the ash weight of TSS) was less correlated with Kd or 1/SD than TSS. The predictions were slightly improved when both TSS and chlorophyll were employed in the regressions.

Relationship between inverse Secchi depth (1/SD) and light attenuation coefficient for measured data at Gunston Cove
The regression equation resulting from the May to September data was:
This equation allows the portion of Kd attributable to Chl-a and that attributable to TSS to be tracked independently. This can be done by multiplying the observed Chl-a by its coefficient (in this case, 0.0118) doing the same for TSS. By looking at a ratio between the calculated Kd (Chl-a) and Kd (TSS) over time, we can determine which has been most responsible for light attenuation over time. As Fig. 7 shows during the period 1992–1998, the contribution of Chl-a to light attenuation declined. This was followed by an increase in Kd (Chl)/Kd (TSS) through 2002 after which Chl-a declined strongly and TSS became the dominant factor in light attenuation.

Relative contribution of Chl-a DI and TSS to light attenuation as given by Kd (Chl)/Kd (TSS). Box plot of all data within each year. Upper and lower ends of box are the upper quartile and lower quartile values, respectively. The horizontal line in the middle of each box is the median. The whiskers extend out to the extent of adjacent values and the asterisks and circles indicate outlying values
Calculating Expected SAV Colonization Depth Based on Light Attenuation
Chesapeake Bay scientists have developed a method for predicting SAV colonization depth based on light attenuation (Kemp et al. 2004). This is most easily calculated using Kd values. Systematic and consistent light extinction measurements leading to calculation of Kd have been collected since 1991. However, SD values have been collected on a regular basis over the entire course of the study. Kd is generally inversely related to SD. A regression was calculated to determine the consistency of this relationship with the goal of being able to back cast light attenuation coefficient to the earlier years of the study.
A highly significant correlation between inverse SD and observed values of Kd was found for the period May–September (Fig. 6, Table 7). These data pairs were well-distributed through the months and years of the study with normally 1–4 measurements per month in every year from 1991 to 2017. Forcing the regression line through the origin (not including a constant in the equation) resulted in a slightly higher slope of 1.473 and a higher correlation coefficient (r) of 0.987. Kd was predicted using this equation.
These strong relationships demonstrate that SD can be effectively and reliably used to backcast Kd values. Thus, for consistency sake, Kd values derived from the measured SD values were used in the habitat suitability model to calculate expected SAV colonization depth for all dates even if measured Kd values were available.
The ZSAV predictions of the habitat suitability model using all available Kd values is shown in a box and whiskers plot by year (Fig. 8). Median SAV colonization depth varied from 0.5 m to 0.75 m over the period from 1984 to 2000. From 2000 to 2006 there was a sustained increase such that median SAV colonization depth reached 1.0 m, remaining above that level for the period from 2005 to 2017 with median values attaining almost 1.5 m in some years. There was a clear shift in the distribution of depths of SAV colonization between the two periods. This change constituted a virtual doubling in SAV colonization depth based on light availability which was expected to produce an increase in SAV areal coverage in the cove as more of the cove would be receiving this threshold light level.

Predicted maximum depth of SAV colonization (ZSAV). Horizontal lines show mean values for periods I and II, respectively from Table 5
Cove Bathymetry
Gunston Cove is a shallow, relatively flat-bottomed embayment of the tidal Potomac River. Harper (1988) mapped the bathymetry of Gunston Cove, dividing the cove into 20 segments with depth-profiled cross-sections at the edge of each of segments. The cove is a Y-shaped body of water (Fig. 1) with the two upper prongs corresponding to Pohick Bay (left prong looking upstream) and Accotink Bay (right prong). For purposes of summarizing the bathymetry of the cove, it was divided into four areal segments shown in Fig. 1 and characterized in Table 8. Note that the cove is pan-shaped with fairly steep sides and extensive very flat bottom and also that within each of the segments there is a gradual deepening moving toward the outer cove. Combining this bathymetry with the calculations of SAV colonization depth above suggests that sufficient light has been present in Pohick and Accotink Bays for SAV growth for most of the study period. The major increase SAV colonization depth would be expected to allow the area of SAV colonization to move out into the inner Gunston Cove segment, but not cover it completely as it gradually deepens beyond the 1.0–1.4-m maximal SAV colonization depth.
SAV Responses
Since 1989, consistent, standardized data have been available via the annual aerial survey of SAV throughout the Chesapeake Bay conducted by the Virginia Institute of Marine Science (Orth and Nowak 1990 to Orth et al. 2018). The data on aerial coverage of SAV in Gunston Cove have been extracted and are compiled in Table 9. The yearly coverage data is also plotted along with average yearly SAV colonization depth (Fig. 9). There is a clear relationship between predicted SAV colonization depth and areal coverage of SAV in Gunston Cove providing support to the hypothesis that light availability is controlling the restoration of SAV in the Cove. A plot of SAV coverage vs. ZSAV (Fig. 10) emphasizes the strong clustering of years in period I vs. years in period II. Note also that changes in the annual spatial pattern of SAV distribution in the cove further supports this linkage between colonization depth and areal coverage (Table 9). From 1989 to 2003, virtually all the SAV was confined to the shallow bays and there was little net change over this period. Beginning in 2004, SAV coverage in the bays became complete and the plants began to appear in the Cove itself consistent with the increase in predicted colonization depth. The expansion of SAV in Gunston Cove was broad-based across all of the species which had been present before 2005 without any obvious shift in relative abundance.

Comparison of observed SAV coverage in Gunston Cove to predicted maximum depth of SAV colonization (ZSAV)

Scatterplot of observed SAV coverage in Gunston Cove to predicted maximum depth of SAV colonization (ZSAV). Numbers beside each data point indicate the year
Discussion
Almost all water quality variables examined in this study exhibited significant changes over the study period. Major temporal changes were observed in nutrient concentrations, BOD, suspended solids, and Chl-a when analyzed by linear regression. Some of the patterns were consistently linear and others were clearly nonlinear, but all showed a strong net change over the period. Water transparency also improved markedly.
The trend line for TP was linear when plotted on a log axis over time indicating a constant rate of decrease over the entire period. This contrasts sharply with the pattern of loading from the Noman Cole Wastewater Treatment Plant which declined abruptly in the late 1970s and early 1980s and was relatively constant at low levels during the period covered by the samples reported in this study. This indicates a delayed response of ambient water column P to external point source loading. One explanation for the delayed response is P release from the sediments of the cove. Cove sediments were high in P in the 1980s (Oehrlein 1990; Kircher and Jones 1991). Strongly elevated pH (> 9.5) was observed on numerous dates during the mid-1980s which results in P release from nutrient-rich sediment in the Potomac (Seitzinger 1991). This mechanism could account for the lagged decline in P over the study period if one assumes that this source becomes gradually less effective at recharging water column P as the frequency of high pH’s declines (as it has) and sediment P becomes gradually depleted or covered by fresh low P sediment. Suspended solids (both total and volatile) and organic N also exhibited fairly consistent linear declines over the whole study period.
Two water quality characteristics exhibited a nonlinear unimodal pattern in which they increased during the 1980s and then declined over the rest of the period: NH3N and BOD. During the early years of the study, NH3N exhibited a steady increase, but with the onset of point source N removal in 1990, ambient levels declined steadily from a median of about 0.3 mg/L in 1990 to a median of < 0.1 mg/L by 2005. Median BOD peaked in 1990 at about 7 mg/L decreasing to 3 mg/L by 2005. Some of the decline in BOD can be ascribed to the decline in NH3N (since oxygen is required to degrade NH3N to NO3N) and the rest to declining decomposable organic matter as expressed by VSS and Chl-a.
Water transparency measures and chlorophyll exhibited a two-phase pattern. Values for SD, Kd, and Chl-a showed little temporal trend from the study onset through about 2003. Then Kd and Chl-a began a downward trend while SD began to increase, and within 2–3 years, all three had attained new ranges. Clearly, major directional changes in water quality had occurred during the study period which corresponded, although not linearly and with some lags, to the observed improvements in wastewater treatment at Noman Cole Wastewater Treatment Plant.
SAV was virtually extirpated in the tidal freshwater Potomac including Gunston Cove in the first half of the twentieth century. Carter et al. (1985) surveyed the tidal Potomac River during the years 1978–1981 and found no SAV in 13 transects sampled in Gunston Cove in spring and fall of 1979. In 1982, the exotic submersed macrophyte Hydrilla verticillata was first reported in the tidal Potomac and extensive surveys conducted in 1983 revealed that H. verticillata was expanding and several native species were also becoming more abundant. Further expansion of SAV was observed in the shallow areas over much of the tidal freshwater Potomac by 1985, but only very small patches were observed in Gunston Cove.
VIMS aerial coverage data indicate that by 1992, the very shallow inner parts of Pohick and Accotink Bays had fairly consistent coverage with SAV amounting to about 50–75 ha with a water depth of 0.6 m or less. Over a 2-year period from 2003 to 2005, total coverage increased to over 200 ha and into areas with a mean water depth of 1.0 to 1.5 m. To examine the extent to which improved water clarity could explain the rapid expansion of SAV in Gunston Cove, the light transparency-SAV maximum depth habitat suitability model developed for Chesapeake Bay was applied (Kemp et al. 2004). Using the 9% of I(0) threshold, the model not only predicted the observed trend in SAV coverage but also quantitatively matched the process well. For example, in the pre-2000 period, maximum colonization depth was predicted at about 0.75 m and SAV was restricted to Pohick and Accotink Bays where the mean depth was 0.6 to 0.75 m. For the 2005–2017 period, SAV colonization depth was about 1.3–1.5 m, and during this period, SAV spread out rapidly into the inner part of Gunston Cove which has a mean depth of about 1.5 m. Use of the 9% of incident light value to calculate expected SAV colonization depth was justified in two ways. This value was within the range specified for freshwater SAV colonization in the Chesapeake Bay region and the predictions produced a better fit of the observed SAV coverage data than somewhat higher values.
A study of a nearby embayment, Mattawoman Creek (Boynton et al. 2014), tracked Chl-a, SD, and SAV acreage from 1985 to 2010 with results similar to those found here. A general doubling in SD was observed from 2000 to 2010 accompanied by a major increase in SAV coverage. They associated the increase in SAV coverage predominantly to a decrease in Chl-a with a threshold of 18 μg/L which corresponded to a Secchi depth of 0.7 m. In the current study, the average Chl-a in the “clear water” phase was 22 μg/L and the average SD was 0.77 m indicating fairly close agreement.
An attempt was made in the current study to partition the relative contribution of suspended sediments and phytoplankton to the changes in light transparency which we hypothesize have driven the ecosystem level changes. The variables that were available to conduct this analysis were SD, Kd, TSS, Chl-a DI, and Chl-a Sf. It was recognized that TSS included not only inorganic sediments but also phytoplankton. Both phytoplankton and suspended sediments strongly correlated with SD and Kd indicating that both are contributing to light attenuation in the study area. Their individual correlations with the light attenuation factors were almost uniform across the board at about 0.75. A multiple regression with both TSS and Chl-a vs. SD and Kd each resulted in a slightly higher correlation coefficient of 0.83–0.84. No temporal trend was observed in the product of SD and Kd which has been observed in other parts of the Chesapeake Bay (Gallegos et al. 2011).
The trends in attenuation assignable to TSS and Chl-a (Fig. 7) suggest that TSS has become a more dominant factor in light attenuation since 1990. The gradual decline in the importance of Chl-a was interrupted by several years around 2000 when chlorophyll attenuation rebounded. This period corresponded to a major drought in the region from Fall 1998 through summer 2002 (Olson 2003). Under drought conditions, inputs of suspended solids would be expected to be at a minimum, and phytoplankton populations could more fully develop due to the absence of flushing and lack of storm-generated suspended sediment clouding the water. Examining the scatterplots for TSS and Chl-a (Fig. 3), we see that several parameters were indeed above the long-term trend line during these years, particularly Chl-a, TP, Org N, and VSS, but TSS values were evenly dispersed on either side of the trend line. So, it appears that the main factor in this temporary shift was an increase in Chl-a during the drought, not a decrease in TSS. Since TP and Org N values were elevated during this period, perhaps more nutrients were available under drought conditions either by external or internal loading and there was less flushing. When wet conditions returned in 2002, Chl-a levels resumed a steady and even expedited decrease and phytoplankton became a much reduced contributor to light attenuation.
The alternative stable state concept provides a useful framework for interpreting our results. Gunston Cove prior to 2003 fits the description of the “turbid water” state or regime dominated by phytoplankton which after several years of gradual decline of TP levels underwent a regime shift in the space of 2 years to a lower nutrient “clear water” state dominated by SAV which has persisted at least through 2017. The occurrence of this phenomenon in multiple shallow freshwater systems worldwide led investigators to assert that shallow water systems are not arrayed equally across the light availability spectrum, but are characterized by one of these two end states (Scheffer et al. 1993). The response of these degraded systems to a reversal of the nutrient loading which brought about the “turbid water” regime is not immediate, but delayed, a phenomenon termed hysteresis (Duarte et al. 2009). While nutrient data are not available for Gunston Cove for the period before degradation, it is clear from the data presented here that there was not a linear response and gradual return to the “clear water” state. Loading was reduced by over 95% by 1980 and TP gradually declined, but it was 25 years later that the system actually showed a decisive shift toward the “clear water” state.
The return to the fully formed “clear water” state with SAV abundant in the entire length of Gunston Cove has still not been achieved. The areal coverage of SAV in Gunston Cove has not increased substantially since 2005 and extends no further than mid-cove. This suggests that Gunston Cove is an example of another phenomenon identified by Duarte et al. (2009), which is “shifting baselines” meaning that fundamental changes in the system have occurred which make return to the original state difficult or impossible. In the case of Gunston Cove, phytoplankton have ceased to be the major driver of light attenuation in most years with suspended sediment becoming the most important factor. The watersheds draining into Gunston Cove have become more developed in the past century and routinely contribute more TSS to the Cove than when the watersheds were forested. Thus, somewhat elevated TSS may be a permanent fixture of the Cove and limit the return to the original even clearer “clear water” stage and resulting spread of SAV to the entire cove. Alternatively, this may be another instance of a lag in ecosystem recovery and enhanced recovery may still be possible.
References
Andersen, J.M. 1975. Influence of pH on release of phosphorus from lake sediments. Archiv für Hydrobiologie 76: 411–419.
Berkley, H.J. 1924. The Port of Dumfries. Prince William County, VA. The William and Mary Quarterly. Second Series 4 (2): 99–116.
Boynton, W.R., C.L.S. Hodgkins, C.A. O’Leary, E.M. Bailey, A.R. Bayard, and L.A. Wainger. 2014. Multi-decade responses of a tidal creek system to nutrient load reductions: Mattawoman Creek, Maryland USA. Estuaries and Coasts 17: S111–S127.
Bricker, S.R., K.C. Rice, and O.P. Bricker III. 2014. From headwaters to coast: Influence of human activities on water quality of the Potomac River estuary. Aquatic Geochemistry 20: 201–323.
Brush, G.S. 2009. Historical land use, nitrogen, and coastal eutrophication: A paleoecological prospective. Estuaries and Coasts 32: 18–28.
Carpenter, S.R. 1980. Enrichment of Lake Wingra, Wisconsin, by submersed macrophyte decay. 61: 1145–1155.
Carter, V., J.E. Paschal, Jr., and N. Bartow. 1985. Distribution and abundance of submersed aquatic vegetation in the tidal Potomac River and Estuary, Maryland and Virginia, May 1978 to November 1981. U.S. Geological Survey Water-Supply Paper 2234-A. 46pp.
Carter, V., and N. Rybicki. 1986. Resurgence of submersed aquatic macrophytes in the tidal Potomac River, Maryland, Virginia, and the District of Columbia. Estuaries 9: 368–375.
Dennison, W.C., R.J. Orth, K.A. Moore, J.C. Stevenson, V. Carter, S. Kollar, P.W. Bergstrom, and R.A. Batiuk. 1993. Assessing water quality with submersed aquatic vegetation. Habitat requirements as barometers of Chesapeake Bay health. Bioscience 43: 86–94.
Duarte, C.M., D.J. Conley, J. Carstensen, and M. Sánchez-Camacho. 2009. Return to Neverland: Shifting baselines affect eutrophication restoration targets. Estuaries and Coasts 32: 29–36.
Edmondson, W.T. 1970. Phosphorus, nitrogen, and algae in Lake Washington after diversion of sewage. Science 169: 690–691.
Gallegos, C.L., P.J. Werdell, and C.R. McClain. 2011. Long-term changes in light scattering in the Chesapeake Bay inferred from Secchi depth, light attenuation, and remote sensing measurements. Journal of Geophysical Research 116: C00H08. https://doi.org/10.1029/201IJC007160.
Gottschalk, L.C. 1945. Effects of soil erosion on navigation in upper Chesapeake Bay. Geographical Review 35: 219–238.
Haramis, G.M., and V. Carter. 1983. Distribution of submersed aquatic macrophytes in the tidal Potomac River. Aquatic Botany 15: 65–79.
Harper, J.D. 1988. Effects of summer storms on the phytoplankton of a tidal Potomac River embayment. Ph.D. Dissertation. Environmental Biology and Public Policy. George Mason University. 205 pp.
Jaworski, N.A., B. Romano, and C. Buchanan. 2007. The Potomac River basin and its estuary: landscape loadings and water quality trends, 1985–2005. https://www.potomacriver.org/the-potomac-river-basin-and-its-estuary-landscape-loadings-and-water-quality-trends-1895-2005/ . Accessed January 10, 2019.
Jones, R.C. 1991. Spatial and temporal patterns in a cyanobacterial phytoplankton bloom in the tidal freshwater Potomac River, USA. Verh. Internat. Verein. Limnol 24: 1698–1702.
Jones, R.C., D.P. Kelso, and E. Schaeffer. 2008. Spatial and seasonal patterns in water quality in an embayment-mainstem reach of the tidal freshwater Potomac River, USA: A multiyear study. Environmental Monitoring and Assessment 147 (1-3): 351–375. https://doi.org/10.1007/s10661-007-0126-0.
Kemp, W.M., W.R. Boynton, R.R. Twilley, J.C. Stevenson, and L.G. Ward. 1984. Influences of submersed vascular plants on ecological processes in upper Chesapeake Bay. Pages 367-394 in: The Estuary as a Filter. V.S. Kennedy (ed). New York: Academic press.
Kemp, W.M., R. Batiuk, R. Bartleson, P. Bergstrom, V. Carter, C.L. Gallegos, W. Hunley, L. Karrh, E.W. Koch, J.M. Landwehr, K.A. Moore, L. Murray, M. Naylor, N.B. Rybicki, J.C. Stevenson, and D.J. Wilcox. 2004. Habitat requirements for submerged aquatic vegetation in Chesapeake Bay: Water quality, light regime, and physical-chemical factors. Estuaries 27: 363–377.
Kircher, S.R. and R.C. Jones. 1991. The temporal and spatial distribution of sediment phosphorus and iron in Gunston Cove, Virginia. In: New perspectives in the Chesapeake system: A research and management partnership. Proceedings of a Conference. Chesapeake Research Consortium. Publication No. 137.
Landers, D.H. 1982. Effects of naturally senescing aquatic macrophyte on nutrient chemistry and chlorophyll a of surrounding waters. Limnology and Oceanography 27: 428–439.
Larsen, D.P., K.W. Malueg, D.W. Schults, and R.M. Brice. 1975. Response of eutrophic Shagawa Lake, Minnesota, U.S.A., to point-source, phosphorus reduction. Verh.Internat. Verein. Limnol. 19: 884–892.
Moss, B. 1980. Further studies on the palaeolimnology and changes in the phosphorus budget of Barton Broad, Norfolk. Freshwater Biology 10: 261–279.
Odum, W.E., T.J. Smith, J.K. Hoover, and C.C. McIvor. 1984. The ecology of tidal freshwater marshes of the United States east coast: a community profile. Fish and Wildlife Service. U.S. Department of Interior. FWS/085-83/17.
Odum, W.E. 1988. Comparative ecology of tidal freshwater and salt marshes. Annual Review of Ecology and Systematics 19: 147–176.
Oehrlein, W.L. 1990. Sediment phosphorus available to phytoplankton as a function of pH in Pohick Bay, Virginia. MS. Thesis. George Mason University.
Olson, M. 2003. Seasonal flow characterizations for the principal tributaries of Chesapeake Bay 1984–2002. Provided by Claire Buchanan.
Orth, R. J. and J. F. Nowak. 1990. Distribution of submerged aquatic vegetation in the Chesapeake Bay and tributaries and Chincoteague Bay - 1989. Final Report to U.S. EPA, Chesapeake Bay Program, Annapolis, MD. https://www.vims.edu/GreyLit/VIMS/sav1989.pdf
Orth, R. J., D. J. Wilcox, J. R. Whiting, A. K. Kenne, and E. R. Smith. 2018. 2017 distribution of submerged aquatic vegetation in the Chesapeake Bay and coastal bays. Virginia Institute of Marine Science. College of Willam and Mary. Gloucester Point, VA. http://www.vims.edu/bio/sav/sav17/index.html .
Osborne, P.L., and B. Moss. 1977. Paleolimnology and trends in the phosphorus and iron budgets of an old man-made lake, Barton Broad, Norfolk. Freshwater Biology 7: 213–233.
Rohlf, F.J., and R.R. Sokal. 1981. Statistical tables. 2nd ed. W.H. Freeman and Co..
Rorslett, B., D. Berge, and S.W. Johansen. 1986. Lake enrichment by submersed macrophytes: A Norwegian whole-lake experience with Elodea canadensis. Aquatic Botany 26: 325–340.
Scheffer, M., S.H. Hosper, M.-L. Meijer, B. Moss, and E. Jeppesen. 1993. Alternative equilibria in shallow lakes. TREE 8 (8): 275–279.
Schubel, J. and D. Pritchard. 1987. A brief physical description of the Chesapeake Bay. In “Contaminant Problems & Management of Living Chesapeake Bay Resources”. p. 1-32.
Seitzinger, S.P. 1991. The effect of pH on the release of phosphorus from Potomac estuary sediments: Implication for blue-green algal blooms. Estuarine, Coastal, and Shelf Science 33: 409–418.
Stewart, R.E. 1962. Waterfowl populations in the upper Chesapeake Bay region. Washington: Fish and Wildlife Service. U.S. Department of the Interior. Special Scientific Report – Wildlife No. 65.
Thomann, R.V., N.J. Jaworski, S.W. Nixon, H.W. Paerl, and J. Taft. 1985. The 1985 algal bloom in the Potomac estuary. Prepared for the Potomac Strategy State/EPA Management Committee. U.S. Environmental Protection Agency, Philadelphia, PA. USA.
Thorp, A.G., R.C. Jones, and D.P. Kelso. 1997. A comparison of water-column macroinvertebrate communities in beds of differing submersed aquatic vegetation in the tidal freshwater Potomac River. Estuaries 20: 86–95.
U.S. Environmental Protection Agency. 1970a. Water quality survey of the Potomac estuary embayments and transects. 1970 Data Report number 28. Region 3. 16 pp. https://nepis.epa.gov/Exe/ZyPURL.cgi?Dockey=2000V9TU.TXT
U.S. Environmental Protection Agency. 1970b. Water quality of the Potomac estuary. Gilbert Swamp and Allen’s Fresh and Gunston Cove. 1970 Data Report number 30. Region 3. 16 pp. https://nepis.epa.gov/Exe/ZyPURL.cgi?Dockey=2000V9TU.TXT
Walker, H.A., J.S. Latimer, and E.H. Dettmann. 2000. Assessing the effects of natural and anthropogenic stressors in the Potomac estuary: Implications for long-term monitoring. Environmental Monitoring and Assessment 63: 237–251.
Acknowledgments
The direct financial and in-kind lab analysis support by Fairfax County, Virginia, Department of Public Works and Environmental Services is gratefully acknowledged. While they focused on other aspects of this ecosystem study, my co-PI’s Donald Kelso, Richard Kraus, and Kim de Mutsert were essential to the overall study success. In addition, there were literally hundreds of graduate and undergraduate students who participated in this study over the 34 years and deserve acknowledgement as a group. Claire Buchanan was an outstanding sounding board and encouraging colleague. Finally, I must acknowledge those who personally supported me during this multidecadal enterprise: my parents, my late wife, Ann, and my friends June and Anne. Needless to say, this work would never have been possible without their support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Hans W. Paerl
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jones, R.C. Recovery of a Tidal Freshwater Embayment from Eutrophication: a Multidecadal Study. Estuaries and Coasts 43, 1318–1334 (2020). https://doi.org/10.1007/s12237-020-00730-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-020-00730-3