
Abstract
Competition for local and shared resources is widespread. For example, colonial waterbirds consume local prey in the immediate vicinity of their colony, as well as shared prey across multiple colonies. However, there is little understanding of conditions facilitating coexistence vs. displacement in such systems. Extending traditional models based on type I and type II functional responses, we simulate consumer-resource systems in which resources are “substitutable,” “essential,” or “complementary.” It is shown that when resources are complementary or essential, a small increase in carrying capacity or decrease in handling time of a local resource may displace a spatially separate consumer species, even when the effect on shared resources is small. This work underscores the importance of determining both the nature of resource competition (substitutable, essential, or complementary) and appropriate scale-dependencies when studying metacommunities. We discuss model applicability to complex systems, e.g., urban wildlife that consume natural and anthropogenic resources which may displace rural competitors by depleting shared prey.





Similar content being viewed by others
References
Amarasekare P (2003) Competitive coexistence in spatially structured environments: a synthesis: spatial coexistence mechanisms. Ecol Lett 6:1109–1122. https://doi.org/10.1046/j.1461-0248.2003.00530.x
Amarasekare P (2008) Spatial dynamics of keystone predation. J Anim Ecol 77:1306–1315. https://doi.org/10.1111/j.1365-2656.2008.01439.x
Brown JS (2000) Foraging ecology of animals. In: Hutchings MJ, John LA, Stewart AJA (eds) The ecological consequences of environmental heterogeneity: the 40th Symposium of the British Ecological Society held at University of Sussex. Cambridge University Press, Cambridge, p 184
Brown DG, Robinson DT (2006) Effects of heterogeneity in residential preferences on an agent-based model of urban sprawl. Ecol Soc 11:46
Cobb CW, Douglas PH (1928) A theory of production. Am Econ Rev 18:139–165
Cooper SD, Diehl S, Kratz K, Sarnelle O (1998) Implications of scale for patterns and processes in stream ecology. Aust J Ecol 23:27–40. https://doi.org/10.1111/j.1442-9993.1998.tb00703.x
Denny MW, Gaines SD (2007) Encyclopedia of tidepools and rocky shores. University of California Press, Oakland
Franco AC, Rossatto DR, de Ramos Silva LC, da Ferreira CS (2014) Cerrado vegetation and global change: the role of functional types, resource availability and disturbance in regulating plant community responses to rising CO2 levels and climate warming. Theor Exp Plant Physiol 26:19–38. https://doi.org/10.1007/s40626-014-0002-6
Grant JA, Quinn MS (2007) Factors influencing transboundary wildlife management in the North American “crown of the continent”. J Environ Plan Manage 50:765–782. https://doi.org/10.1080/09640560701609323
Graven HD, Keeling RF, Piper SC et al (2013) Enhanced seasonal exchange of CO2 by northern ecosystems since 1960. Science 341:1085–1089. https://doi.org/10.1126/science.1239207
Haegeman B, Loreau M (2015) A graphical-mechanistic approach to spatial resource competition. Am Nat 185:E1–E13. https://doi.org/10.1086/679066
Henny CJ (1972) An analysis of the population dynamics of selected avian species—with special references to changes during the modern pesticide era. U.S. Fish and Wildlife Service, Patuxent Wildlife Research Center, Laurel
Holling CS (1959) Some Characteristics of Simple Types of Predation and Parasitism. Can Entomol 91:385–398. https://doi.org/10.4039/Ent91385-7
Hothem RL, Hatch D (2004) Reproductive success of the black-crowned night heron at Alcatraz Island, San Francisco Bay, California, 1990-2002. Waterbirds 27:112–125
Hunt VM (2016) Reproductive success and habitat selection in black-crowned night-herons (Nycticorax nycticorax) in a city park. Am Midl Nat 175:168–182. https://doi.org/10.1674/0003-0031-175.2.168
Jorge MLSP, Brown JS, van der Merwe M (2012) Handling time and the evolution of caching behavior. Behav Ecol 23:410–417. https://doi.org/10.1093/beheco/arr205
Kneitel JM, Chase JM (2004) Trade-offs in community ecology: linking spatial scales and species coexistence. Ecol Lett 7:69–80. https://doi.org/10.1046/j.1461-0248.2003.00551.x
Levin S (1992) The problem of pattern and scale in ecology. Ecology 73:1943–1967. https://doi.org/10.2307/1941447
McNickle GG, Gonzalez-Meler MA, Lynch DJ et al (2016) The world’s biomes and primary production as a triple tragedy of the commons foraging game played among plants. Proc R Soc B 283:20161993. https://doi.org/10.1098/rspb.2016.1993
Monod J (1949) The growth of bacterial cultures. Annu Rev Microbiol 3:371–394. https://doi.org/10.1146/annurev.mi.03.100149.002103
R Development Core Team (2009) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Soetaert K, Petzoldt T, Setzer RW (2010) Solving differential equations in R: package deSolve. Journal of Statistical Software 33: 1-25
Trouwborst A (2012) Transboundary wildlife conservation in a changing climate: adaptation of the Bonn convention and its daughter instruments to climate change. Social Science Research Network, Rochester
Vickery WL, Brown JS (2009) Spite, egotism, population stability, and resource conservation. Evolutionary Ecology Research 11:253–263.
Vincent TLS, Scheel D, Brown JS, Vincent TL (1996) Trade-offs and coexistence in consumer-resource models: it all depends on what and where you eat. Am Nat 148:1038–1058
Whelan CJ, Schmidt KA, Steele BB et al (1998) Are bird-consumed fruits complementary resources? Oikos 83:195–205. https://doi.org/10.2307/3546561
Acknowledgments
The authors thank Rosalyn Rael, Paul Orlando, and two anonymous reviewers for providing very helpful comments and insights that improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(R 75 kb)
Appendices
Appendix 1: Complementary resources and partially selective diet
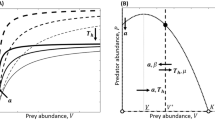
This appendix describes the derivation of rules governing consumption of complementary resources when consumers use an optimal foraging strategy and have a type II functional response. Table 1 of the main text shows equations for isolegs, lines in the resource state space where opportunistic and optimal foraging strategies would yield equal gain, which are based on these rules. Figure 6 provides a visual of the isolegs in the resource state space, and the shape of zero net growth isoclines (ZNGIs) for optimal vs. opportunistic foraging.

Zero net growth isoclines (ZNGIs) and isolegs in a type II system with complementary resources. The shape of the ZNGI depends on the foraging strategy employed by consumers: optimal foraging (dotted red line) vs. opportunistic foraging (solid red line). Isolegs divide the resource state space into three regions in which the optimal forager will have different probabilities of accepting resources. The probability of accepting an item of the local resource is p 1 and the probability of accepting an item of the shared resource is p 2
Under a given level of consumption (C i , where i ∈ {1, 2}), the fitness value of consuming more of the local resource, R 1, or shared resource, R 2, is given by
Equations (9) and (10) are derived from Eq. (3) of the main text. An encountered item of the shared resource should be rejected if greater fitness can be had by searching for and handling an item of the local resource than by simply handling an item of the shared resource. Therefore, consumers foraging optimally will reject an item of the shared resource if
Equation (11) can be rewritten as
Thus, the likelihood of being partially selective increases with α, a 1, R 1, h 2, and C 2. The likelihood of being partially selective decreases with β, h 1, and C 1. A similar equation can be derived for when an encountered item of the local resource should be rejected. In the event that the condition in Eq. (12) is satisfied, then the forager should modulate its consumption of the shared resource so that the condition is met with equality. Therefore, C 2 should be determined by C 1 according to the following relationship (found by setting the above equation equal and solving for C 2 as a function of C 1):
Conditions governing consumption in a system with optimal foraging can be visualized as isolegs, lines that divide the resource state space into regions wherein consumers use different foraging strategies. Substituting the consumption rates when feeding is opportunistic, and rearranging Eq. (13) yields the isolegs separating when the forager should be an opportunist, and when it should be partially selective (Table 1). Between the isolegs, consumers forage opportunistically, consuming resources as encountered. At high values of one resource, consumers reject consumption of some of the more abundant resource to increase consumption on the more limiting resource (Fig. 6). We observe that the main differences between optimal and opportunistic foraging in a type II system is that the shape of the ZNGIs is such that more of the limiting resource is required at high values of the non-limiting resource when foraging is opportunistic (Fig. 6).
Appendix 2: Simulation parameters
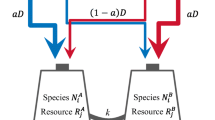
This appendix explains the rationale for choosing simulation parameters. The average death rate of the two populations (0.7) is based on the estimated death rate of 0.7 for black-crowned night herons (Nycticorax nycticorax) in their first year (Henny 1972). Conversion efficiency rates of turning resources into consumers (0.1 or 0.2) are also based on life history parameters of black-crowned night herons. For Fig. 5 with complementary resources, d A = 0.4 and d B = 0.9. In this case, more extreme parameter values were used to illustrate scenarios that would result in consumer displacement.
Consumption multiplied by conversion efficiency yields per capita increase in population size. Therefore, maximum birth rate will be conversion efficiency multiplied by the carrying capacity of the resources. For example, for essential resources in Table 2, conversion efficiency (0.1) multiplied by the maximum resource carrying capacity (16) yields per capita birth rate 1.6 (3.2 per pair). Black-crowned night herons have one to five eggs per clutch, with average 2.9 eggs per clutch, and average annual mean hatching success of nests of 56% (Hothem and Hatch 2004). From these life history values, we determine that a maximum birth rate of 3.2 per pair is reasonable.
Rights and permissions
About this article

Cite this article
Hunt, V.M., Brown, J.S. Coexistence and displacement in consumer-resource systems with local and shared resources. Theor Ecol 11, 83–93 (2018). https://doi.org/10.1007/s12080-017-0350-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12080-017-0350-3