
Abstract
Charles Darwin is well known for his studies on the expression of emotions in animals and humans and as founding father of the concept of sexual selection. Yet it is commonly believed that the various arguments Darwin developed about behavior were usually illustrated only by anecdotes and observations recounted by explorers, naturalists, or zookeepers, and lacking any experimental approach. Here we show that this is not true. In his last book, The Formation of Vegetable Mould Through the Action of Worms (1881), Darwin mentions a series of meticulous experiments he ran to test his hypotheses about why earthworms plug their burrows and comes to the conclusion that earthworms seem to act in an intelligent way. His study can still function as a prime example of how to design an experiment for testing hypotheses. Only one part was missing in Darwin’s research: statistical analyses. We retrieved his data and analyzed them statistically. Based on these results, we cannot reject his conclusion as the statistical analyses confirmed Darwin was right. This shows that Charles Darwin already used a hypothetico-deductive approach, and he can thus be seen as the first true behavioral ecologist—a representative of a discipline that has been recognized for only about a hundred years.
Similar content being viewed by others


Introduction
The study of animal behavior probably dates back to the earliest history of humankind. To survive and reproduce successfully, it was perhaps crucial to know how a lion hunts or where and when to ambush prey. Yet the scientific analysis of behavior is relatively recent. It evolved gradually via several lineages, two of the most prominent being behaviorism, mainly in North America, and ethology in Europe (Dewsbury 1999). In the 1970s, its most recent branch emerged, behavioral ecology, which is strongly founded in evolutionary biology and population genetics (Dewsbury 1999). Two hallmarks of behavioral ecology, in contrast to classical ethology, for example, are an essentially hypothetico-deductive formulation of questions with hypotheses often preceding observation and an experimental approach to test these hypotheses (Danchin et al. 2008).
In The Origin of Species (Darwin 1859), Charles Darwin already recognized that behavior, like any other trait, is liable to evolve by natural selection, and he later outlined the idea of sexual selection (Darwin 1871), which became one of the most prominent topics of behavioral biology. Despite this impact on the studies of animal behavior, it is generally thought that the various arguments Darwin developed about behavior were usually illustrated only by anecdotes and observations recounted by explorers, naturalists, or zookeepers (Danchin et al. 2008). This attitude is, for instance, expressed in the notion that “There is no trace in Darwin’s work of any truly experimental approach in support of his theories of behaviour” (Danchin et al. 2008, p. 11).
Here we show that this is not the case. In his last book, The Formation of Vegetable Mould Through the Action of Worms (Darwin 1881), he recounts a series of systematic observations of earthworms (Lumbricus terrestris; see Butt et al. 2008) which still today could function as a guideline on experimental design for students in behavioral biology. Based on an initial observation and reasoning about intelligence, he explicitly formulated hypotheses predicting how earthworms draw leaves inside their burrows to block them. Then he rigorously tested his hypotheses with various independent experiments in the field and under “laboratory” conditions. Finally, he critically discussed his results. Only one part essential to current behavioral studies was missing from his analyses: statistics. Here, we analyze his data set on earthworm behavior from 1881, and as a result, we can confirm Darwin’s (1881) conclusions even after rigorous testing.
Darwin’s Experiments
Reasoning about the intelligence of earthworms in plugging of their burrows Darwin (1881) stated: “If a man had to plug up a small cylindrical hole with such objects as leaves, petioles or twigs, he would drag or push them in by their pointed ends; but if these objects were very thin relatively to the size of the hole, he would probably insert some by their thicker and broader ends. The guide in this case would be intelligence” (Darwin 1881, p. 64).
We retrieved the original data of Darwin’s most important experiments that tested the hypothesis that earthworms pull leaves into burrows in an intelligent way. Most experiments were done in Charles Darwin’s garden or in his house (Down House, Kent, England). For more details, see Darwin’s results (1881). We analyzed Darwin’s data with statistical methods. All percentage values were transformed into numbers before each test. Chi-square tests were done without the application of Yate’s correction with the program R, version 2.5.1 (R Development Core Team 2007).
Reanalyses of Darwin’s Results
-
Experiment 1:
Natural assemblage of leaves from the garden (native and non-native trees, generally pointed tip, broad base; Darwin 1881, pp. 65–66).
Darwin collected 227 leaves from worm burrows in his garden. One hundred and eighty-one were drawn into the burrows by or near their tips, 20 by their bases, and 26 near the middle. This means that significantly more leaves were drawn in by the easiest site (the tip) than expected by chance (χ²2 = 220.19, p < 0.0001).
Seventy of the 227 leaves were from the common lime tree (Tilia spec.), a species supposed to be non-native to England (Fig. 1a). Also here, significantly more leaves were drawn into burrows by their tips than base or middle (χ²2 = 66.20, p < 0.0001; Fig. 1a).
Fig. 1 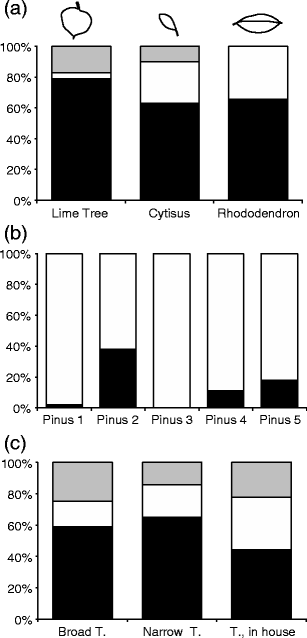
Results of Darwin’s experiments. Shown are the percentage of “leaves” dragged into burrows by the tip (black), the base (white), or the middle section (gray). a Experiments with differently shaped leaves from plants non-native to England; the shape of the leaves is shown above. b Experiments with pine needles: Pinus 1 Needles collected from the garden, Pinus 2 Pot experiments in the house, Pinus 3 Pot experiments in the garden, Pinus 4 Shellac experiment, Pinus 5 Thread experiment. c Experiments with paper triangles of different shapes (broad, narrow) and in the house. For more information, see text
-
Experiment 2:
Leaves from a non-native species, the blades of which are not more pointed towards the apex than towards the base (Darwin 1881, pp. 67–68).
He collected 73 leaves of a hybrid between Cytisus alpinus and Cytisus laburnum from burrows in the field. In this species, the terminal half generally fitted exactly the basal half (Fig. 1a). Significantly more leaves were drawn into the burrows by their tips than expected by chance (χ²2 = 32.41, p < 0.0001), yet the proportion of leaves drawn into the burrow by their bases was significantly higher than in the case of the common lime tree (χ²2 = 14.63, p < 0.0001; Fig. 1a).
-
Experiment 3:
Non-native plant with variable leaf shapes—Rhododendron (Fig. 1a) (Darwin 1881, pp. 69–70).
Two hundred thirty-seven leaves were collected from the garden. Based on their shape, 65% would be most easily dragged into burrows by the base, 27% by the tip, and 8% by either end. Out of 91 leaves collected from burrows, 66% were drawn by the base and 34% by the tip. Here the observed dragging pattern fitted very well with those expected based on leaf shape. There was no significant difference between the observed and expected frequencies (χ²1 = 0.67, p = 0.414; Fig. 1a).
-
Experiment 4:
Non-native plants under different conditions, Pinus needles (Darwin 1881, pp. 70–77).
The sharp, pointed leaves of pine trees also connected at the base (Fig. 1b) present a challenge for earthworms. After observing how earthworms drag these needles into their burrows, Darwin did a series of experiments under different environmental conditions. In the garden, in an area without pine trees, earthworms unfamiliar with these leaves dragged a significantly higher proportion (90 out of 92 needles) in the correct way than expected by chance, i.e., by their base rather than their two tips (χ²1 = 84.17, p < 0.0001; Fig. 1b; Pinus 1). He then did the same experiment in a warm room of his house with earthworms placed in pots. Here significantly more needles (16 out of 42 needles) were dragged by their tip (χ²1 = 32.00, p < 0.0001; Fig. 1b; Pinus 2) and the burrows were not closed properly. He interpreted this difference as a consequence of the warmer room conditions and tested this new hypothesis. He placed pots with earthworms outside where they were exposed to colder ambient temperatures. Now all 72 needles were dragged inside the burrows by their base (Fig. 1b, Pinus 3).
In the next experiment, he glued the tips of needle pairs together with shellac to test whether the earthworm’s behavior resulted because they “gain a general notion of the shape or structure of the pine leaves and perceive that it is necessary for them to seize the base where the two needles are conjoined.” After evaporation of the shellac, he deposited the leaves in an area without pine trees. Still 108 from 121 leaves were dragged into the burrows by their base (χ²1 = 74.59, p < 0.0001; Fig. 1b, Pinus 4). To test for a side effect of the shellac, he did a second experiment in which he tied the tips of the needles together with a fine thread. Also here, 123 out of 150 leaves were dragged by their base (χ²1 = 61.44, p < 0.0001; Fig. 1b, Pinus 5). These experiments led Darwin to conclude that the base of pine tree leaves has something attractive, although other leaves are rarely dragged by their base.
-
Experiment 5:
Artificial leaves: triangles of paper (Darwin 1881, pp. 82–90).
In a last set of experiments, Darwin used artificial “leaves” of two different shapes. He cut elongated triangles out of “moderately stiff writing-paper, which was rubbed with raw fat on both sides, so as to prevent their becoming excessively limp when exposed at night to rain and dew.” The sides of all triangles were three inches in length, so that they could be partitioned into a tip (first upper inch), mid- (second inch), and base section (third inch). The base of the triangles was either 1 inch (broad triangles, N = 120) or 0.5 inch (narrow triangles, N = 183). After the triangles were placed outside, 59% from the broad triangles were dragged into burrows by the tip, 25% by the mid-section, and 16% by the base (Fig. 1c). The corresponding values for the narrow triangles were 65%, 14%, and 21% (Fig. 1c). Thus in both cases, significantly more triangles were dragged by their tips than the 33% expected by chance (broad triangles: χ²2 = 37.55, p < 0.0001; narrow triangles: χ²2 = 82.90, p < 0.0001; Fig. 1c), and this despite the fact that the tip section had a smaller surface, smaller circumference, and fewer edges than the base section, as Darwin (1881) discussed in detail. He also could reject the hypothesis that the earthworms tried several sides first, before they finally dragged the triangles with the most appropriate side, i.e., the tip; the triangles were not covered with dirt and they were not crumpled, as would have been the case were the dragging process a result of trial and error.
As with the pine needles, he did a similar experiment in a warm room in his house using 63 triangles. Here again, earthworms acted more “careless,” and they used each section not significantly differently from a one in three chance (tip, 44%; middle, 22%; base, 33%; χ²2 = 4.67, p < 0.097; Fig. 1c). Thus again, the earthworm’s behavior between the warm room and the garden differed significantly (χ²2 = 6.69, p < 0.035).

Conclusions
Which conclusions did Darwin draw from his meticulous experiments? “No doubt worms are led by instinct to plug their burrows; and it might have been expected that they would have been led by instinct how best to act in each particular case, independently of intelligence” (Darwin 1881, pp. 91–92). Yet his results led to the rejection of the latter hypothesis. As Darwin discussed in detail (pp. 92–95), the dragging behavior of earthworms differed from random expectation, and it depended upon the object being presented. As several of these objects were new for earthworms (foreign plants, paper triangles), evolved instinct could not explain their dragging behavior. The next option would be trial-and-error learning, which according to Romanes (in Darwin 1881), involves intelligence (p. 95). As the last experiments with the paper triangles showed, the earthworms drag leaves in the right way even without trial and error. In fact, they drag leaves as humans would do: get an impression (notion) about the shape of an object, judge which way would be the best, and then drag the object accordingly. According to Darwin (1881), this is what earthworms seem to do: they get an “image” of the leaf shape by touching it with their prostomium. This left Darwin to conclude that earthworms, despite their “little brains,” seem to be intelligent. Based on the reanalysis of his data, we cannot reject this concluding hypothesis of Darwin.
References
Butt KR, Lowe CN, Beasley T, Hanson I, Keynes R. Darwin’s earthworms revisited. Eur J Soil Biol. 2008;44:255–9.
Danchin E, Giraldeau LA, Cézilly F. Behavioural ecology. Oxford: Oxford University Press; 2008.
Darwin C. On the origin of species by means of natural selection. London: John Murray; 1859.
Darwin C. The descent of man and selection in relation to sex. New York: Appleton; 1871.
Darwin C. The formation of vegetable mould through the action of worms, with observations on their habits. London: John Murray; 1881.
R Development Core Team (2007) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org
Dewsbury DA. The proximate and the ultimate: past, present, and future. Behav Processes. 1999;46:189–99.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Korb, J., Salewski, V. Unearthening Old Data: Darwin was Indeed Correct About Earthworm Behavior. Evo Edu Outreach 4, 133–136 (2011). https://doi.org/10.1007/s12052-010-0295-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12052-010-0295-0