
Abstract
Background
Proliferative therapy, or prolotherapy, is a controversial treatment method for many connective tissue injuries and disorders. It involves the injection of a proliferant, or irritant solution, into the site of injury, which causes small-scale cell death. This therapeutic trauma is theorized to initiate the body’s wound-healing cascade, perhaps leading to tissue repair. The immediate effects of many of these proliferants are poorly characterized, as are the cellular responses to them; here, we sought to evaluate the immediate effects of two common proliferants (dextrose and P2G, a combination of phenol, glucose, and glycerin) on the cellular response of human tenocytes, and begin to explicate the mechanisms with which each proliferant functions.
Questions/purposes
We asked: What are the effects of treating cultured tenocytes with proliferative treatment agents on their (1) cellular metabolic activity, (2) RNA expression, (3) protein secretion, and (4) cell migration?
Methods
Using human hamstring and Achilles tendon cells, we attempted to answer our research questions. We used a colorimetric metabolic assay to assess the effect of dextrose and P2G proliferant treatment on cell mitochondrial activity compared with nontreated tenocytes. Next, using quantitative PCR, ELISA, and a reporter cell line, we assessed the expression of several key markers involved in tendon development and inflammation. In addition, we used a scratch wound-healing assay to evaluate the effect of proliferant treatment on tenocyte migration.
Results
Results showed that exposure to both solutions led to decreased metabolic activity of tenocytes, with P2G having the more pronounced effect (75% ± 7% versus 95% ± 7% of untreated control cell metabolic levels) (ANOVA; p < 0.01; mean difference, 0.202; 95% CI, 0.052–0.35). Next, gene expression analysis confirmed that treatment led to the upregulation of key proinflammatory markers including interleukin-8 and cyclooxygenase-2 and downregulation of the matrix marker collagen type I. Furthermore, using a reporter cell line for transforming growth factor-β (TGF-β), a prominent antiinflammatory marker, we showed that treatments led to decreased TGF-β bioactivity. Analysis of soluble proteins using ELISA revealed elevated levels of soluble prostaglandin E2 (PGE2), a prominent inducer of inflammation. Finally, both solutions led to decreased cellular migration in the tenocytes.
Conclusions
Taken together, these results suggest that prolotherapy, more so with P2G, may work by decreasing cellular function and eliciting an inflammatory response in tenocytes. Additional studies are needed to confirm the cellular signaling mechanisms involved and the resulting immediate response in vivo.
Clinical Relevance
If these preliminary in vitro findings can be confirmed in an in vivo model, they may provide clues for a possible cellular mechanism of a common alternative treatment method currently used for certain soft tissue injuries.
Similar content being viewed by others

Introduction
Ligament and tendon healing from injury often is hampered by incomplete healing and chronic tissue instability [15, 27]. Both tissues are strong collagenous structures that operate primarily in tension and are integral in body locomotion. Because of their structure, function, and composition, these tissues often are regarded as mostly avascular and therefore lack robust healing capabilities [8, 18, 32]. To address this, several approaches to repair that seek to decrease healing time and increase tissue biomechanical properties have been investigated. Conventional modalities including drug therapy (NSAIDs, corticosteroid injections, analgesics) and rest, ice, compression, and elevation have been met with their associated drawbacks [15, 23]. Because of this, alternative approaches have garnered increasing attention.
Proliferative therapy or prolotherapy is a controversial, nonsurgical approach that has been proposed for ligament and tendon injury [9]. The treatment involves injection of a proliferant (or irritant solution) into the damaged or lax connective tissue [10, 11, 28]. Originally used in the 1950s in the clinic by Dr. George Hackett as a method to tighten loose ligaments [11, 12], it is theorized that the proliferant initiates a local inflammatory response, accompanied by cell death and the release of chemotactic factors and inflammatory mediators to initiate the body’s wound-healing cascade. During progression of this cascade, it is presumed by some [2] that cell death leads to the release of factors such as prostaglandins, thromboxanes, and leukotreines, which recruit inflammatory cells. Granulocytes and macrophages from the initial stage of inflammation may release factors that attract and activate fibroblasts. These fibroblasts deposit new collagen at the wound site, which eventually matures and contracts, and may cause the ligament or tendon to tighten and strengthen. Commonly used proliferant agents include dextrose, phenol, glucose, glycerin, and morrhuate sodium used either alone or in combination (one common combination is P2G, which is comprised of phenol, glycerin, and glucose). Systematic reviews and meta-analyses on the clinical efficacy of prolotherapy have found very few rigorous studies, with a majority of studies having a high risk of bias, small treatment effect sizes, and short study durations [3, 17, 29, 31].
Prolotherapy has been reported to be used for various musculoskeletal ailments, but is most commonly used in the clinic to tighten lax joints and treat overuse tendon injuries. The therapy is practiced worldwide, especially in primary care, although investigations into the mechanisms behind the treatment’s healing have yielded inconsistent results [10, 14, 15, 28]. Unfortunately, very little to virtually no research exists regarding the in vitro cellular response of proliferants in clinical use [10]. Previous in vitro work by our group determining the effect of P2G on prominent extracellular matrix-building cells showed that the local response to prolotherapy of resident fibroblasts and osteoblasts in the tissue may not be adequate for complete healing, making assistance from the body’s wound-healing cascade necessary [10]. More specifically, P2G treatments can lead to recoverable cell death and collagen deposition. However, the response from treatment groups never surpassed controls, suggesting that the local response of resident cells may be insufficient for complete tissue healing in vivo. These initial results warranted further investigation into understanding the proliferant-induced cell response with a special focus on the immediate cascade of events (within 1 day). Considering such, it would be valuable to ascertain the theorized underlying molecular mechanisms leading to the prolotherapy-induced inflammatory response in vitro.
The objective of our study was to assess the immediate cellular response of prolotherapy. We investigated the effects of two common proliferant drugs, dextrose (or d-glucose) and P2G, on the cellular response to prolotherapy. We theorized that both proliferants will induce considerable changes in cellular response that ultimately lead to cell death and induction of inflammation in vitro. We asked the following questions: What are the effects of treating cultured tenocytes with proliferative treatment agents on (1) cellular metabolic activity of cultured tenocytes, (2) mRNA expression, (3) protein secretion, and (4) cell migration?
Materials and Methods
Study Overview
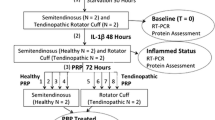
The goal of our study was to characterize the immediate cellular response to prolotherapy in vitro. To answer our research questions, we designed an in vitro experimental protocol and used a human tendon cell culture model (Fig. 1). Experimental groups included cells treated with two different proliferant agents and an untreated control population. For our first question regarding the effects of prolotherapy treatment on tenocyte metabolic activity, we used a fluorometric assay, which measures the mitochondrial activity of cultured cells. To answer our next question regarding the effect of prolotherapy on the immediate expression and production of key markers involved in tendon development and inflammation, we used quantitative PCR (qPCR), ELISA, and a transforming growth factor-beta (TGF-β) reporter cell line. We began by examining the mRNA expression of collagen types I and III after treatment because they are the two main collagens present in tendon tissue and are two appropriate markers of tenogenic differentiation and identity. Next we evaluated the mRNA expression of a small panel of inflammatory mediators including cyclooxygenase-2 (COX-2), interleukin (IL)-6, IL-8, and NFκβ. The initial results were further validated by assessing secreted levels of prostaglandin E2 (PGE2) and TGF-β1 using ELISA and the bioactivity of the secretome using a TGF-β reporter cell line. Finally, to answer our question concerning the effect of prolotherapy treatment on the cellular migration of tendon cells, we used a tenocyte scratch wound-healing assay.

The experimental overview of the study is shown.
Proliferants
Two commonly used proliferants, 50% (v/v) dextrose and P2G (a combination of 2% phenol, 25% glucose, and 25% glycerin), in sterile water were obtained (Buderer Drug Company, Perrysburg, OH, USA) and used for cellular studies.
Cell Isolation and Culture
All procedures and studies were conducted with approval from the Rutgers Environmental Health and Safety Office, the Medical Ethical Research Committee at the Utrecht Medical Center (The Netherlands), and Ludwig-Maximilians-University (Munich, Germany). Following standard written informed consent, human hamstring tendon samples were harvested from an adult patient undergoing ACL reconstruction. After isolation, the tendons were rinsed with sterile phosphate-buffered saline (PBS), excess muscle tissue was carefully removed, and the tendon was cut into smaller pieces. Next, the tendon pieces were cultured in growth medium consisting of Dulbecco’s modified Eagle’s medium (DMEM) (PAA Laboratories, Morningside, Queensland Australia) supplemented with 10% fetal bovine serum (FBS) (Lonza, Basel, Switzerland), 100 U/mL penicillin and 100 µg/mL streptomycin (Gibco, Grand Island, NY, USA), and 0.2 mmol/L ascorbic acid (Sigma Aldrich, St Louis, MO, USA) for the cells to migrate from the tissue pieces. Primary cells between passages 2 and 4 were used for experiments. In addition to using a primary human hamstring tendon isolation for experiments, we also repeated our experiments with an immortalized cell line from human Achilles tendon. For generation of a tendon-derived stable cell line, tendon-derived cells were obtained from human Achilles tendon biopsy according to methods described by Kohler et al. [16]. Lentivirus, expressing telomerase reverse transcriptase (TERT), was produced as previously described [6] and according to the manufacturer’s instructions in the ViraPowerTM lentiviral expression system (Invitrogen, Karlsruhe, Germany). The primary Achilles tendon cells were infected at passage 4 with hTERT lentivirus (multiplicity of infection, 5 × 104) in the presence of 6 µg/mL polybrene (Sigma, Munich, Germany). After a selection with 10 µg/mL blasticidin (Invitrogen) for 7 days, the transgene expression was validated by hTERT PCR as described previously [6]. This new hTERT-immortalized Achilles tendon cell line then was used for various analyses. Both cell types were maintained in α-MEM at pH 7.6 supplemented with 1% penicillin and streptomycin and 10% FBS (Life Technologies, Carlsbad, CA, USA).
Experimental Design
Cells were seeded on tissue culture well plates at a density of 5 × 103 cells/cm2 plate and allowed to adhere overnight before treating with 0.024% (v/v) of P2G and dextrose. This concentration was selected based on prior in vitro work with prolotherapy [10]. After 6, 12, and 24 hours of treatment, cell populations were evaluated using assays as described subsequently. All comparisons are made between an untreated control group and experimental groups (P2G and dextrose). For conditioned medium studies, after 24 hours of treatment, the conditioned medium was collected and stored at −80° C for further analysis of cell-secreted products. All experiments were performed with biologic replicates (n = 3) and each experiment was repeated at least twice.
Cellular Metabolic Activity
Cell metabolic activity was evaluated using the fluorometric PrestoBlue® assay (Life Technologies). At each time, each well was emptied and 10% (v/v) PrestoBlue reagent in fresh cell culture medium was added to each well. The plates were incubated for 1 hour followed by measurements of fluorescence in technical duplicates at 560 nm excitation and 590 nm emission with a spectrophotometer (Tecan Schweiz AG, Mannedorf, Switzerland). Fluorescent values for treatment groups were normalized to control groups.
Real-time Quantitative Reverse Transcription-PCR Analysis
For qPCR studies, total RNA was extracted with the RNeasy® Mini Kit (Qiagen, Valencia, CA, USA) according to the vendor’s protocol. Isolated RNA was normalized, then converted to cDNA using the Reverse Transcription System (Promega Corporation, Madison, WI, USA). After reverse transcription, the cDNA was combined with SYBR® Green Master Mix (Life Technologies), and the primers for markers (Table 1) of interest then were subjected to qPCR. Experiments were conducted with glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the housekeeping gene. Samples and reagents were subjected to qPCR using the PikoRealTM PCR System (Thermo Fisher, St Louis, MO, USA).
ELISA
Conditioned medium collected from hamstring tenocyte cultures was analyzed for presence of two soluble factors. ELISA kits for total TGF-β1 (Biolegend, San Diego, CA, USA) and PGE2 (Cayman Chemical Co, Ann Arbor, MI, USA) were obtained and used to assess levels of soluble protein secreted by hamstring tenocytes after treatment with proliferants. Assays were performed in technical duplicate according to the manufacturer’s directions and absorbance was quantified using a Molecular Devices EMax® microplate reader (Molecular Devices, Sunnyvale, CA, USA).
TGF-β Bioactivity Assay
We used a TGF-β reporter cell line to screen for antiinflammatory effects of the proliferants. Transformed mink lung cells, a kind gift from Daniel Rifkin PhD (Department of Cell Biology, New York University, New York, NY, USA), are cells that have been modified to produce luciferase under activation of a TGF-β-responsive plasminogen activator inhibitor-1 promoter [1, 34]. The cells were seeded in triplicate at 8 × 103/cm2 and allowed to attach overnight in basic DMEM. Next, cells were exposed to proliferants for 16 hours. At this point, the experiment was stopped and cells were washed with PBS, lysed, and then analyzed for luciferase activity using a standard luciferin substrate kit (Promega Corporation) and luminometer (Tecan Schweiz AG).
Scratch Wound Healing Assay
Cell migration was assessed using the standard scratch wound-healing assay. Briefly, 1 × 104 cells/cm2 human Achilles tenocytes were seeded and allowed to reach confluence. A scratch was induced across the cell monolayer using a sterile 1-mL pipette tip. Afterward, culture medium was replaced with fresh medium and cell populations were treated with either P2G or dextrose. At various times, the migration of cells while closing the “wound” was monitored using light microscopy. For each sample, a total of three images of the “wound” were taken in each of the three regions (top, middle, bottom) of the well. Images were analyzed using Image J (NIH, Bethesda, MD, USA). Briefly, the area created by the “wound” was quantified by measuring the number of corresponding pixels for each image.
Statistical Analysis
All quantitative data are reported as mean ± SD. A one- or two-way ANOVA was performed with a Bonferroni’s multiple comparison post hoc test to compare proliferant groups. A Student’s t-test was used for comparison of control groups with each treatment group in the mRNA expression experiments (GraphPad Prism Software 5.0; GraphPad, La Jolla, CA, USA). Unless otherwise stated, differences were reported as statistically significant when p values were less than 0.05.
Results
Proliferant Effects on Cellular Metabolic Activity
In human hamstring and Achilles tenocyte cell lines, P2G consistently produced greater reductions in cell metabolic activity than did dextrose. For all times, the metabolic activity of hamstring tenocyte populations treated with P2G was lower compared with populations treated with dextrose (Fig. 2A). The mean metabolic activity of hamstring tenocyte populations reached lowest levels after 12 hours for P2G at 64% (SD, ± 4) and dextrose at 85% (SD, ± 2) of control levels (mean difference, 0.206; 95% CI, 0.144–0.268; p < 0.001). Additionally, Achilles tenocyte populations showed lower metabolic activity when treated with P2G, and after 24 hours showed no signs of recovery after treatment with P2G (Fig. 2B). The mean difference between the control and P2G-treated groups at 24 hours was 0.2348 (95% CI, 0.1355–0.3342; p < 0.001).

Prolotherapy decreases metabolic activity in human tenocytes. The normalized cell metabolic activity of (A) hamstring tendon tenocytes and (B) Achilles tenocytes during 24 hours of treatment with proliferants was determined. Fluorescent values were normalized to nontreated control populations (dotted line at 1.0) (*p < 0.05, †p < 0.01, ‡p < 0.001).
Proliferant Effects on Tenocyte mRNA Expression
Within 6 hours, P2G treatment induced decreases in the mRNA expression of collagen type I in hamstring tendon cells, whereas neither proliferant had any detectable effects on the expression of collagen type III (Fig. 3A). For collagen type I mRNA in hamstring tenocytes, the mean relative expression of P2G and dextrose-treated cells were 0.48- (SD, ± 0.18) and 0.44-fold (SD, ± 0.04), respectively. The mean difference between the control and P2G groups was 0.532-fold (95% CI, 0.197–0.868-fold; p < 0.05). Additionally, in Achilles tenocyte populations, P2G and dextrose treatment led to decreases in collagen type I mRNA expression, whereas only dextrose led to a decrease in collagen type III mRNA expression (Fig. 3B). For collagen type I expression in Achilles tenocytes, the mean relative expression of P2G and dextrose-treated cells were 0.51-(SD, ± 0.09) and 0.40- (SD, ± 0.04), respectively. The mean difference between the control and P2G groups was 0.497-fold (95% CI, 0.221- 0.773-fold; p < 0.05). The mean difference between the control and dextrose groups was 0.611-fold (95% CI, 0.335–0.887-fold; p < 0.01). For collagen type III expression in Achilles tenocytes, the mean relative expression of P2G and dextrose-treated cells were 0.76-fold (SD, ± 0.12) and 0.37-fold (SD, ± 0.03), respectively. The mean difference between the control and dextrose-treated groups was −0.637-fold (95% CI, −0.821 to 0.455-fold; p < 0.001).

Prolotherapy decreases mRNA expression of collagen. The mRNA expression of (A) hamstring tendon and (B) Achilles tenocytes was determined after 6 hours of proliferant exposure (*p < 0.05, †p < 0.01, ‡p < 0.001).
Furthermore, within 6 hours in the dextrose-treated populations there was a decrease in mean relative IL-8 and NFκB-1 mRNA expression (0.29 ± 0.13- and 0.60 ± 0.01-fold, respectively) of hamstring tenocytes, whereas P2G treatment increased the relative mRNA expression of COX-2 and IL-8 by 18.56 ± 6.69- and 22.46 ± 91-fold, respectively (Fig. 4A). The mean differences between the control and dextrose-treated groups for IL-8 and NFκB-1 were −0.399-fold (95% CI, −0.7835 to −0.0151-fold; p < 0.05) and −0.732-fold (95% CI, −1.24 to −0.225-fold; p < 0.05), respectively. The mean difference between the control and P2G-treated groups in relative expression of COX-2 mRNA was 17.5-fold (95% CI, 1.02–33.98; p < 0.05). The mean difference in this comparison for IL-8 expression was 21.45-fold (95% CI, −1.18 to 44.09; p < 0.01). In Achilles tenocyte populations, dextrose and P2G caused a decrease in the mRNA expression of NFκB-1 (0.16 ± 0.13- and 0.48 ± 0.08-fold, respectively). The mean difference between the control and dextrose-treated groups was −0.846-fold (95% CI, −1.14 to −0.552-fold; p < 0.01). The mean difference between the control and P2G-treated groups was −0.525-fold (95% CI, −0.766 to −0.283-fold; p < 0.01). In addition, there was an upregulation of COX-2, IL-6, and IL-8 after treatment with dextrose (5.33 ± 0.86-, 2.13 ± 0.33-, and 3.83 ± 0.97-fold, respectively), whereas P2G-upregulated COX-2 and IL-8 (7.06 ± 2.40- and 14.52 ± 4.99- fold, respectively) (Fig. 4B). The mean difference between the control and dextrose-treated groups in the relative expression of COX-2 mRNA was 4.34-fold (95% CI, 2.22- 6.457-fold; p < 0.001). The mean difference for this comparison for IL-6 was 1.11-fold (95% CI, 0.437 −1.78-fold; p < 0.05) and for IL-8 was 2.83-fold (95% CI, 1.22–4.44-fold; p < 0.01). The mean difference between the control and P2G-treated groups in the relative expression of COX-2 was 6.06-fold (95% CI, 2.21- 9.91-fold; p < 0.05) and for IL-8 was 13.5-fold (95% CI, 5.52–21.5-fold; p < 0.01).

Prolotherapy increases mRNA expression of proinflammatory markers. The mRNA expression of inflammatory markers was determined in (A) hamstring tendon and (B) Achilles tenocytes after 6 hours of proliferant exposure (*p < 0.05, †p < 0.01, ‡p < 0.001).
Proliferant Effects on Tenocyte Secretion
Evaluation of soluble levels of PGE2 showed that P2G-treated hamstring tendon cells produced higher levels of PGE2 compared with dextrose-treated and control hamstring tenocyte populations (76.18 ± 24.65 versus 81.28 ± 15.15 versus 401.21 ± 93.60 pg/mL, respectively; p < 0.01) (Fig. 5A). Mean differences for comparing the P2G-treated group with the control and dextrose-treated groups were −325 pg/mL (95% CI, −466.7 to −183.3 pg/mL; p < 0.01) and −319.9 pg/mL (95% CI, −461.6 to −178.2 pg/mL; p < 0.01), respectively. After assessment of total TGF-β1, results did not show a considerable change in protein levels after treatment with proliferants (Fig. 5B). We then directly assessed the potential of treatments on directly inducing TGF-β activity. Additional screening with a TGF-β reporter cell line revealed that after 16 hours, both proliferants induced decreased luminescence after treatment. More specifically, P2G-treated cells exhibited the lowest luminescence in comparison to control cells and dextrose-treated populations (Fig. 5C). Relative luminescent unit (RLU) values for control, dextrose-, and P2G-treated cell populations were 17.8 ± 3.60, 15.7 ± 3.20, and 9.14 ± 3.61 RLU, respectively. The mean difference for comparing the control and P2G-treated groups was 8.63 RLU (95% CI, 3.42–13.8 RLU; p < 0.01). The mean difference for comparing the dextrose and P2G-treated groups was 6.51 RLU (95% CI, 1.30–11.7 RLU; p < 0.05).

Prolotherapy increases prostaglandin E2 (PGE2) and decresases TGF-β1 secretion. The amounts of soluble (A) PGE2 and (B) total transforming growth factor-beta 1 (TGF-β1) secreted from hamstring tendon tenocytes after 1 day of treatment was determined using ELISA. (C) The normalized TGF-β bioactivity measured in relative luminescent units (RLU) of TGF-β reporter cells after a 16-hour treatment with proliferants (*p < 0.05, †p < 0.01).
Proliferant Effects on Tenocyte Migration
In human hamstring and Achilles tendon cell populations, prolotherapy treatment with P2G and dextrose led to a decrease in cell migration (Fig. 6A, B). After 24 hours of treatment, the scratch assay revealed differences in residual scratch width among the untreated control, P2G, and dextrose groups (345 ± 109 versus 886 ± 196 versus 523 ± 171 μm, respectively; p < 0.001). Mean differences for comparing the P2G-treated group with the control and dextrose groups were 540.3 μm (95% CI, 330.7–749.9 μm) and −361.8 μm (95% CI, −488.2 to −235.4 μm), respectively. It appeared as if cell populations treated with dextrose began to reclose the wound, whereas cells treated with P2G showed less cell migration and a larger wound area. Additionally, after 6 hours of treatment with both proliferants, cells had mean cell migration rates of 73 ±9 μm/hour for control cells, 18 ± 17 μm/hour for dextrose- treated cells, and 21 ± 37 μm/hour for P2G-treated cells (p < 0.01) (Fig. 6C). Mean differences for comparing the untreated control group with the dextrose and P2G-treated groups were 54.66 μm/hour (95% CI, 18.46–90.86 μm) and 51.6 μm/hour (95% CI, 15.4–87.8 μm), respectively.

Prolotherapy decreases tendon cell migration. (A) Brightfield imaging of Achilles tenocytes during wound closure (white vertical lines) during treatment with proliferants, (B) average scratch width of the Achilles tenocyte monolayer wound closure during treatment with proliferants, and (C) average cell migration rate of the Achilles tenocytes during treatment with proliferants are shown (†p< 0.01).
Discussion
Prolotherapy is a commonly used, but controversial alternative treatment for connective tissue injury. Prior investigations of the treatment’s efficacy and mechanisms have been lacking. To begin to understand prolotherapy and its clinical utility, we can start to elucidate the immediate response of prolotherapy at the cellular level by studying the effect of commonly used proliferant agents. Our results revealed that both proliferant agents tested (P2G and dextrose) led to immediate decreases in the metabolic activity of cultured human tenocytes. In addition, treatment led to strong decreases in the mRNA expression of extracellular matrix components collagen types I and III. Concurrently, we observed differential mRNA expression of COX-2, IL-6, and IL-8. Further screening with a TGF-β reporter cell line revealed strong decreases in TGF-β bioactivity after proliferant treatment. Assessment for levels of soluble markers revealed elevated levels of PGE2, the enzymatic product of COX-2 signaling, after treatment with P2G. Cell migration analysis revealed that prolotherapy decreased tenocyte migratory capabilities.
There are several limitations to this study. First, it is an in vitro study of cultured tenocytes. How live tendons function in a live patient with a tendon injury is likely different. It is possible that the in vivo response would differ because of biologic factors present in the cellular microenvironment and the tissue operating under physiologic conditions (eg, tissue biomechanics and fluid or blood flow). However, we believe that studying how the cells react in vitro is a key initial step to determining how these treatments might work in the clinical setting. Another limitation is that our experiments were performed on only two different cell types. Albeit, different tendon types (and even other cell types from surrounding tissues) may behave in a different fashion. In fact, some of our results highlight this at the cellular level. Although we have not studied every possible tendon type, we offer important preliminary evidence with these two tissue types at the in vitro cellular level. Moreover, some of the more-dramatic responses seen (downregulation of collagen type I mRNA, upregulation of COX-2 mRNA after P2G treatment) were consistent among both cell types studied. Furthermore, our in vitro model focuses on the resident tissue cells (tendon cells) responding to prolotherapy. To fully characterize the prolotherapy-induced reponse in the tissue, additional work regarding the effect of the cellular products generated by prolotherapy-treated tendon cells on activating cultured immune cells is necessary. These data would provide important information regarding the type of localized inflammatory response that would be expected in vivo.
Our first research question inquired about the effect of prolotherapy on the metabolic activity of tendon cells. In agreement with the central hypothesis behind prolotherapy, both proliferants resulted in decreased cell activity levels during treatment. Results showed that P2G induced much lower cell activity than dextrose at every time assessed for the hamstring tenocytes (Fig. 2A) and a similar trend was seen for the Achilles tenocytes at 6 and 24 hours of treatment (Fig. 2B). These results suggest that P2G may be more effective at inducing the initial cell death required to initiate the wound-healing cascade. Proliferant drugs are classified in three categories: irritants, osmotics, and chemotactics. Each class is named after the suggested mechanism of action for initiating the localized inflammatory response [24, 28]. Dextrose is considered an osmotic that functions through the dehydration and subsequent necrosis of cells after injection. Other studies have explored the utility of hypertonic dextrose solutions for inducing fibrosis and the mechanisms by which this occurs [25, 35]. The most-active ingredient in P2G, phenol, works as an irritant, which oxidizes into quinine groups leading to cellular damage. Treatment with both molecules ultimately leads to localized cell death that initiates the body’s wound-healing cascade.
We next asked about the effect of prolotherapy on the mRNA expression of relevant tendon developmental and inflammatory markers. Gene expresssion analysis of hamstring and Achilles tenocytes at 6 hours of treatment showed that both proliferant treatments resulted in decreased mRNA transcripts of collagen types I and III mRNA in Achilles tenocytes and only decreased collagen I mRNA in hamstring tenocytes. Furthermore, analysis of a panel of proinflammatory markers showed that P2G elevated mRNA transcripts of COX-2 (also known as prostaglandin-endoperoxide synthase 2) and IL-8. These results suggest that prolotherapy with dextrose and P2G, most notably P2G, induced elevated mRNA expresssion of specific inflammatory markers. COX-2 has been implicated in many proinflammatory responses seen in tendon disorders and wound repair [21, 33]. Additionally, it is recognized as the precursor to and involved in the production of PGE2, a potent inducer and mediator of tissue inflammation [4]. In addition, the decreased expression of NFκB-1 (and varied expression of IL-6 and IL-8) suggests that cells may have been experiencing a specific expression pattern during treatment. These results warrant further investigation into the upstream cell-signaling mechanisms involved.
Our next question inquired about the secretion of key factors from tendon cells after prolotherapy treatment. Using ELISA to measure soluble levels of TGF-β1 and PGE2, we observed a notable increase in PGE2 levels after treatment with P2G (Fig. 5A). Using a reporter cell line to screen for TGF-β bioactivity, we saw that dextrose and P2G decreased TGF-β bioactivity after 24 hours of treatment (Fig. 5C). TGF-β has been implicated in the development of several musculoskeletal tissues and is a well-known antiinflammory factor known to induce collagen and extracelluar matrix production during wound healing [20, 30]. It is believed by some that prolotherapy induces cellular death and the subsequent release of proinflammatory and chemotactic factors, which initiate the body’s wound healing cascade [2, 14, 15]. Results from our experiments directly support aspects of this theory, but also present additional evidence for prolotherapy as a negative regulator of TGF-β signaling. However, more work is needed to further determine the exact mechanism and to predict the clinical effectiveness of prolotherapy. Previous work involving the utility of proliferant molecules in vitro for orthopaedic applications is extremely limited and available studies span several different applications [5, 19, 26]. Nonetheless, the active molecules in each proliferant in our study have been studied previously. Investigations involving dextrose have explored the effect of prolonged exposure to elevated glucose levels for the study of diabetic conditions and the effect on various cell responses. For instance, in a study with human mesangial cells, Clarkson et al. [5] showed that sustained exposure to elevated extracellular glucose levels led to the differential expression of 200 genes that primarily were involved in the regulation of extracellular matrix production, cell growth, and cell cytoskeletal maintenance. Additionally, in a study of elevated glucose levels on the effect of human renal fibroblasts, Lam et al. [19] showed that connective tissue growth factor and insulin-like growth factor were heavily implicated in glucose-induced collagen deposition during fibrosis. Phenol is commonly used in the clinic as a neurolytic agent for nerve blocks and as a local anesthetic [13]. Although much of the prolotherapy response in vivo will be dependent on the volume and concentration used, it appears that P2G may have a more-pronounced inflammatory reponse in tendon owing to its mechanism of action.
Finally, we assessed the effect of each proliferant on directly modulating cellular migration, another critical component of cell function. The wound-healing assay is a common model to study cellular migration [7, 22]. This parameter provides useful supplemental information about the remaining cell population after the initial cell death from prolotherapy. Results showed that dextrose and P2G, most notably P2G, induced decreased cellular migration in Achilles tenocytes during the 24 hours of treatment (Fig. 6). Interestingly, it appeared that cell populations treated with dextrose started to show recovery by beginning to close the gap at 24 hours. These results suggest that prolotherapy induces important changes in tendon cells that are accompanied by strong decreases in the cells’ ability to migrate. They also support the notion that prolotherapy does not directly enhance tendon cell migration after treatment. Any increases in cell migration after prolotherapy may be an indirect result of the treatment (eg, downstream cellular signaling from immune cells).
The results from our experiments showed that human tenocytes treated with dextrose and P2G experienced considerable decreases in cellular activity. Furthermore, gene expression studies confirmed the upregulation of proinflammatory markers COX-2 and IL-8, whereas the growth-related marker collagen type I was downregulated. P2G led to a strong increase in PGE2 secretion, and further screening with a reporter cell line confirmed a strong decrease in TGF-β bioactivity after treatment with both proliferants. Finally, we showed that both proliferants, most notably P2G, led to decreased cellular migration. These preliminary findings will require further rigorous in vivo study in a live animal model and ultimately humans, but they provide initial insight into the potential mechanisms behind prolotherapy with P2G proving to be a more-potent inducer of inflammation in vitro.
References
Abe M, Harpel J, Metz CN, Nunes I, Loskutoff DJ, Rifkin DB. An assay for transforming growth factor-beta using cells transfected with a plasminogen activator inhibitor-1 promoter-luciferase construct. Anal Biochem. 1994;216:276–284.
Banks A. A rationale for prolotherapy. J Orthop Med. 1991;13:54–59.
Chou R, Atlas SJ, Stanos SP, Rosenquist RW. Nonsurgical interventional therapies for low back pain: a review of the evidence for an American Pain Society clinical practice guideline. Spine (Phila Pa 1976). 2009;34:1078–1093.
Cilli F, Khan M, Fu F, Wang JH. Prostaglandin E2 affects proliferation and collagen synthesis by human patellar tendon fibroblasts. Clin J Sport Med. 2004;14:232–236.
Clarkson MR, Murphy M, Gupta S, Lambe T, Mackenzie HS, Godson C, Martin F, Brady HR. High glucose-altered gene expression in mesangial cells: actin-regulatory protein gene expression is triggered by oxidative stress and cytoskeletal disassembly. J Biol Chem. 2002;277:9707–9712.
Docheva D, Padula D, Popov C, Weishaupt P, Pragert M, Miosge N, Hickel R, Bocker W, Clausen-Schaumann H, Schieker M. Establishment of immortalized periodontal ligament progenitor cell line and its behavioural analysis on smooth and rough titanium surface. Eur Cell Mater. 2010;19:228–241.
Doorn J, Fernandes HA, Le BQ, van de Peppel J, van Leeuwen JP, De Vries MR, Aref Z, Quax PH, Myklebost O, Saris DB, van Blitterswijk CA, de Boer J. A small molecule approach to engineering vascularized tissue. Biomaterials. 2013;34:3053–3063.
Empson YM, Ekwueme EC, Hong JK, Paynter DM, Kwansa AL, Brown C, Pekkanen AM, Roman M, Rylander NM, Brolinson GP, Freeman JW. High elastic modulus nanoparticles: a novel tool for subfailure connective tissue matrix damage. Transl Res. 2014;164:244–257.
Florida Academy of Pain Medicine. Regenerative injection therapy (RIT): effectiveness and appropriate usage. Pain Clinic Magazine.2002;4:38–45. Available at: http://www.gracermedicalgroup.com/resources/articles/rf_file_0025.pdf. Accessed April 10, 2017.
Freeman JW, Empson YM, Ekwueme EC, Paynter DM, Brolinson PG. Effect of prolotherapy on cellular proliferation and collagen deposition in MC3T3-E1 and patellar tendon fibroblast populations. Transl Res. 2011;158:132–139.
Hackett GS. Ligament and Tendon Relaxation (Skeletal Disability): Treated by Prolotherapy, (Fibro-osseous Proliferation). Springfield, IL: Charles C. Thomas; 1958.
Hackett GS, Henderson DG. Joint stabilization; an experimental, histologic study with comments on the clinical application in ligament proliferation. Am J Surg. 1955;89:968–973.
Jarrett L, Nandi P, Thompson AJ. Managing severe lower limb spasticity in multiple sclerosis: does intrathecal phenol have a role? J Neurol Neurosurg Psychiatry. 2002;73:705–709.
Jensen KT, Rabago DP, Best TM, Patterson JJ, Vanderby R Jr. Response of knee ligaments to prolotherapy in a rat injury model. Am J Sports Med. 2008;36:1347–1357.
Jensen KT, Rabago DP, Best TM, Patterson JJ, Vanderby R Jr. Early inflammatory response of knee ligaments to prolotherapy in a rat model. J Orthop Res. 2008;26:816–823.
Kohler J, Popov C, Klotz B, Alberton P, Prall WC, Haasters F, Muller-Deubert S, Ebert R, Klein-Hitpass L, Jakob F, Schieker M, Docheva D. Uncovering the cellular and molecular changes in tendon stem/progenitor cells attributed to tendon aging and degeneration. Aging Cell. 2013;12:988–999.
Krogh TP, Bartels EM, Ellingsen T, Stengaard-Pedersen K, Buchbinder R, Fredberg U, Bliddal H, Christensen R. Comparative effectiveness of injection therapies in lateral epicondylitis: a systematic review and network meta-analysis of randomized controlled trials. Am J Sports Med. 2013;41:1435–1446.
Kwansa AL, Empson YM, Ekwueme EC, Walters VI, Freeman JW, Laurencin CT. Novel matrix based anterior cruciate ligament (ACL) regeneration. Soft Matter. 2010;6:5016–5025.
Lam S, van der Geest RN, Verhagen NA, van Nieuwenhoven FA, Blom IE, Aten J, Goldschmeding R, Daha M, van Kooten C. Connective tissue growth factor and IGF-1 are produced by human renal fibroblasts and cooperate in the induction of collagen production by high glucose. Diabetes. 2003;52:2975–2983.
Leask A, Abraham DJ. TGF-beta signaling and the fibrotic response. FASEB J. 2004;18:816–827.
Legerlotz K, Jones ER, Screen HR, Riley GP. Increased expression of IL-6 family members in tendon pathology. Rheumatology (Oxford). 2012;51:1161–1165.
Liang CC, Park AY, Guan JL. In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nat Protoc. 2007;2:329–333.
Lynch SA, Renstrom PA. Treatment of acute lateral ankle ligament rupture in the athlete: conservative versus surgical treatment. Sports Med. 1999;27:61–71.
Nair LS. Prolotherapy for tissue repair. Transl Res. 2011;158:129–131.
Natarajan R, Gonzales N, Xu L, Nadler J. Vascular smooth muscle cells exhibit increased growth in response to elevated glucose. Biochem Biophys Res Commun. 1992;187:552–560.
Pace D, Elliot A. Effects of acetone and phenol on established cell lines cultivated in vitro. Cancer Res. 1962;22:107–112.
Provenzano PP, Alejandro-Osorio AL, Valhmu WB, Jensen KT, Vanderby R Jr. Intrinsic fibroblast-mediated remodeling of damaged collagenous matrices in vivo. Matrix Biol. 2005;23:543–555.
Rabago D, Slattengren A, Zgierska A. Prolotherapy in primary care practice. Prim Care. 2010;37:65–80.
Sanderson LM, Bryant A. Effectiveness and safety of prolotherapy injections for management of lower limb tendinopathy and fasciopathy: a systematic review. J Foot Ankle Res. 2015;8:57.
Schiller M, Javelaud D, Mauviel A. TGF-beta-induced SMAD signaling and gene regulation: consequences for extracellular matrix remodeling and wound healing. J Dermatol Sci. 2004;35:83–92.
Sit RW, Chung VCh, Reeves KD, Rabago D, Chan KK, Chan DC, Wu X, Ho RS, Wong SY. Hypertonic dextrose injections (prolotherapy) in the treatment of symptomatic knee osteoarthritis: a systematic review and meta-analysis. Sci Rep. 2016;6:25247.
Voleti PB, Buckley MR, Soslowsky LJ. Tendon healing: repair and regeneration. Annu Rev Biomed Eng. 2012;14:47–71.
Wang JH, Jia F, Yang G, Yang S, Campbell BH, Stone D, Woo SL. Cyclic mechanical stretching of human tendon fibroblasts increases the production of prostaglandin E2 and levels of cyclooxygenase expression: a novel in vitro model study. Connect Tissue Res. 2003;44:128–133.
Wipff PJ, Rifkin DB, Meister JJ, Hinz B. Myofibroblast contraction activates latent TGF-beta1 from the extracellular matrix. J Cell Biol. 2007;179:1311–1323.
Yoshii Y, Zhao C, Schmelzer JD, Low PA, An KN, Amadio PC. Effects of hypertonic dextrose injections in the rabbit carpal tunnel. J Orthop Res. 2011;29:1022–1027.
Author information
Authors and Affiliations
Corresponding author
Additional information
The institutions of one or more of the authors have received funding from the National Science Foundation (NSF CBET 1243144 [JWF], NSF CBET 1034026 [JWF], and NSF DGE 0801620 [ECE]) and Smith & Nephew (HAMF).
All ICMJE Conflict of Interest Forms for authors and Clinical Orthopaedics and Related Research ® editors and board members are on file with the publication and can be viewed on request.
Clinical Orthopaedics and Related Research ® neither advocates nor endorses the use of any treatment, drug, or device. Readers are encouraged to always seek additional information, including FDA-approval status, of any drug or device prior to clinical use.
Ludwig-Maximilians-University (Munich, Germany), Utrecht Medical Center (Utrecht, The Netherlands), and Rutgers Environmental Health and Safety Office (Piscataway, NJ, USA) approved or waived approval for the human protocol for this investigation, and each author certifies that all investigations were conducted in conformity with ethical principles of research.
This work was performed at Rutgers University, Piscataway, NJ, USA.
About this article

Cite this article
Ekwueme, E.C., Mohiuddin, M., Yarborough, J.A. et al. Prolotherapy Induces an Inflammatory Response in Human Tenocytes In Vitro. Clin Orthop Relat Res 475, 2117–2127 (2017). https://doi.org/10.1007/s11999-017-5370-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11999-017-5370-1