
Abstract
Toxoplasma gondii is an obligate intracellular parasite that causes toxoplasmosis. It has been shown that the severity of symptoms depends on the functioning of the host immune system. Although T. gondii infection typically does not lead to severe disease in healthy people and after infection, it induces a stable immunity, but it can contribute to severe and even lethal Toxoplasmosis in immunocompromised individuals (AIDS, bone marrow transplant and neoplasia). The antigens that have been proposed to be used in vaccine candidate in various studies include surface antigens and secretory excretions that have been synthesized and evaluated in different studies. In some studies, secretory antigens play an important role in stimulating the host immune response. Various antigens such as SAG, GRA, ROP, ROM, and MAG have been from different strains of T. gondii have been synthesized and their protective effects have been evaluated in animal models in different vaccine platforms including recombinant antigens, nanoparticles, and DNA vaccine. Four bibliographic databases including Science Direct, PubMed Central (PMC), Scopus, and Google Scholar were searched for articles published up to 2020.The current review article focuses on recent studies on the use and usefulness of recombinant antigens, nanoparticles, and DNA vaccines.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Toxoplasmosis is a parasitic disease caused by the intracellular protozoan Toxoplasma gondii. The infection is transmitted through drinking of water contaminated by definitive host’s feces containing oocysts or through consumption of the definitive or intermediate hosts’ tissues containing tissue cysts [1]. The parasite life cycle includes the following steps in summary: the oocysts sporulation step in the environment that makes them infective takes 1–5 days. The next step is the infection of intermediate hosts in nature (including sheep, pigs, cattle, birds, rodents, and humans) after ingesting soil, water, or plant material contaminated with oocysts. In this step, oocysts transform into tachyzoites shortly after ingestion [2].
Toxoplasmosis can present in two forms in human. The first form is asymptomatic and occurs by latent infections due to tissue cysts consumption. The second form is severe infections that occur in immunocompromised hosts (e.g., AIDS and organ transplant recipients) or in fetus or newborn with congenital toxoplasmosis [3].
The toxoplasmosis adverse consequences are due to the ability of the parasite to destroy host cells especially cells from critical organs such as brain and eye [4]. Furthermore, the parasite is capable of crossing the placenta; therefore, it can cause fatal or severe and debilitating morbidity in the fetus and newborn [5]. Infection in pregnant women especially during the first trimester can lead to spontaneous abortion, preterm labor, or severe congenital defects such as hydrocephalus, mental retardation, and chorioretinitis [6]. Toxoplasmosis and its prevention is also an important issue in the veterinary medicine and livestock industry as the infection not only decreases the meat production by causing abortion especially in sheep and goat but also can be both a source for human infections and a reservoir for the parasite [7]. Currently, toxoplasmosis control strategies are largely based on treatment by medications in the acute phase of the infection [8].
However, the current medications have drawbacks including toxic effects, limited accessibility, and high costs. Furthermore, reinfections can occur after treatment due to the complicated life cycle of the T. gondii and the presence of its infectious life stages in the environment [6, 8, 9]. In addition, the drug is not effective on the tissue cysts and is not applicable in the primary stages when the infected individuals are asymptomatic [10, 11]. The parasite exists in three forms depending on the stage in its life cycle: tachyzoites, bradyzoites, and sporozoites that are similar in ultrastructure but with differences in intracellular organelles [12]. The parasite life cycle can be divided into two phases: (1) the intestinal or isospora stage in definitive feline hosts. (2) Extra-intestinal stage in both the definitive and intermediate hosts [13]. The prevalence of congenital toxoplasmosis dramatically varies from 1 to 100 per 10,000 live births in different countries and even in different regions or communities in one country [14]. It is estimated that approximately 30–50% of the world population is infected by the T. gondii [15]. The severity of the disease by the parasite is determined by the host resistance, the parasite’s variants and the antigenic variations. The main antigens of the T. gondii are membrane, cytoplasmic, and soluble antigens that the latter results from a combination of cytoplasmic shedding, active secretion by the parasite, and lysis due to the immune system responses [16]. The parasite secretory antigens that are produced by three parasite’s organelles microneme, rhoptry, and dense granule comprise 90% of the soluble antigens [17, 18]. The T. gondii DNA vaccine studies are mainly focused on four families of molecules. These are surface antigens (SAGs), microneme antigens (MIC), rhoptry antigens (ROP), and dense granules antigens (GRA) [19, 20]. Identification of the molecules that are crucial in pathogenesis and immune protection is a bottleneck in efficient vaccine development. Excretory/secretory antigens (ESAs) produced by T. gondii in tachyzoites and bradyzoites forms have an important role in immune system stimulation [21]. These antigens are mainly GRA that is suggested as a candidate antigen for vaccine development [22]. The microneme secretions contain cell surface adhesion molecules that are involved in the first step of parasite adhesion and invasion to the host [23, 24]. The rhoptry antigens are secreted into the expanding parasitophorous vacuoles during the parasite invasion [24]. The immune response to the T. gondii depends on the clinical presentation of the infection. The CD4 + and CD8 + cells are crucial for protection against the infection [25]. These cells are involved in protection by secretion of inflammatory cytokines such as IFN-γ, TNF-α, IL-1, and IL-6. Toxoplasmosis can induce CD8 + cytotoxic lymphocytes in both human and mice that can destroy the infected cells [26]. It has been found that all the mice strains can develop a strong T helper cell type 1 (Th1) immune response against the T. gondii. Furthermore, macrophages, natural killer (NK) cells, dendritic cells (DCs), antibodies, and other immune effector factors are involved in the prevention of the infection [27]. So far, a variety of T. gondii antigens have been identified by different methods and the molecular characteristics of many of them have been evaluated to be used in diagnostic, therapeutic, immunization, and vaccine development applications [28].
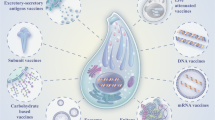
In people with a healthy immune system, the symptoms of the infection usually are similar to mild flu symptoms, while in immunocompromised patients, it can cause severe and even life-threatening complication such as encephalitis and severe ocular complications. In addition, in pregnant women, toxoplasmosis may cause abortion or congenital toxoplasmosis with manifestations such as neurological or ocular in the fetus [6, 29]. It is estimated that approximately 50% of untreated maternal infections are transmitted to the fetus in which approximately 60% are subclinical, 30% have severe damage such as hydrocephalus, intracerebral calcification, retinochoroiditis (Classical triad) and mental retardation and 9% are fatal for the fetus [30]. Generally, in immunocompetent women with one experience of T. gondii infection related abortion or fetal infection [31], the subsequent pregnancies are safe regarding T. gondii reinfection and manifestations; however, there is occasional reports implying transmission of congenital toxoplasmosis by immunocompetent women infected before conception [32, 33]. The available therapeutics for the treatment of toxoplasmosis are not completely safe and effective [6, 20]. Recently considerable progress has been made in designing toxoplasma vaccine candidates that can efficiently stimulate the immune responses [34]. In the current study, we reviewed the T. gondii candidate vaccines that include a heterogeneous collection of studies with different methodologies including recombinant antigens, micro/nanoparticles displaying the antigens and DNA vaccine.
Methods
Database search
Four bibliographic databases including Science Direct, PubMed Central (PMC), Scopus, and Google Scholar were searched for articles published up to 2020. The following MeSH (Medical Subject Headings) keywords were considered in the initial search strategy: “Toxoplasmosis,” “nanoparticles -based vaccines,” “Recombinant vaccines,” and “DNA vaccines of T. gondii” with employing the Boolean operators ‘OR’ and/or ‘AND.’
Vaccine design based on recombinant antigens
Toxoplasmosis in a healthy host that is immunocompetent stimulates a lifelong protective immunity that prevents reinfection. The immunogenic proteins that can induce these protective responses have been identified that are candidates for vaccine development for T. gondii. These immunogenic proteins include surface antigens (SAG), dense granule antigens (GRA), rhoptry proteins (ROP), and microneme proteins (MIC) [35]. The use of recombinant antigens as a candidate vaccine against toxoplasmosis has been considered since the 1990s [36]. At the beginning, the SAG 1 antigen, which is on the surface of the parasite, as well as the (GRA)1 antigen were studied [37]. Subsequently, after 2000, other secretory antigens such as GRA7, ROP2, MICs, and other proteins such as heat shock proteins considered as candidates of vaccines. In recent years, more than 10 genes have been cloned into eukaryotic and bacterial expression systems, as follows: ROP 18 [38, 39], GRA4 [40,41,42], and ROP4 [40, 41, 43, 44], ROP2 [41,42,43,44,45,46], SAG1 [41, 45, 47, 48], actin depolymerizing factor (ADF),GRA6 [49, 50], ROP5 [48], GRA2 [50, 51], SAG2 [52], Toxoplasma gondii Hsp70 (TgHSP 70) [53], Toxoplasma tissue cyst matrix protein (MAG1) [44], T. gondii serin protease inhibitor-1 (TgPI-1) [42], GRA5 [51]. Many of these antigens have been used to detect specific antibodies in the serum of mice, pigs, and cats and to assess the immune response in the host. The results of some studies have shown that immunization using these recombinant antigens responds well and effectively induce the immune responses. Table 1 shows the immunogenicity of recombinant antigens produced in different hosts such as mice, pigs, and cats.
Vaccine design based on DNA genome of antigens
The DNA vaccines are a new strategy to prevent infectious diseases to help reduce the antibiotics use and diseases spread. The DNA vaccines can be used in oral or injective forms to induce rapid immunization against a diverse range of diseases that are hard to be overcome by antibiotics or traditional vaccines. A typical DNA vaccine is composed of a plasmid with a strong viral promoter and the gene of interest that is expressed and induces specific immune responses. The advantages of DNA vaccines include stability, cost effectiveness, and safety. However, the hurdle in DNA vaccine development against parasites is the complexity of the parasitic diseases.
The GRA and SAG antigens that are parasitic secretory and surface antigens are suitable candidate antigens to design DNA vaccines for immunity against toxoplasmosis. It has been demonstrated that DNA vaccination with the sequences of GRA1, GRA7, and ROP2 proteins can induce protection against infection with different virulent T. gondii strains in C3H mice but not in BALB/c and C57BL/6 mice. Furthermore, immunization of sheep with a DNA vaccine containing the GRA1, GRA4, GRA6, and GRA7 sequences formulated in liposome showed a significant immune response against T. gondii [54]. Table 2 lists the studies of antigens used as candidate DNA vaccines. Including SAG1 antigen from ME49, VEG strains [55], SAG1, ROP16, GRA14,MIC8,ROP54, Toxoplasma gondii calcium-dependent protein kinase 2 (TgCDPK2), T. gondii Myc regulation 1 (MYR1), Perforin-like proteins (PLP)1, ROP18, GRA2, GRA5, GRA17, GRA23, GRA7, ROP2, TgHSP60, ROP21, TgHSP-40,GRA16, Rhomboid 4(ROM4), ROP35,GRA8, GRA4, GRA24, GRA25, MIC6, SAG5-D for RH [42, 55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77] and ROP35, GRA8, ROP19, GRA24, GRA25, MIC6 for PRU strain [72, 74, 75].
Vaccine design based on nanoparticles of recombinant antigens
Nanoparticles can be used to improve delivery of subunit vaccine in order to increase the immunogenicity of the pathogen proteins used in the vaccine design [78]. Furthermore, virus-like particles (VLPs) or nanoparticles have been used to design recombinant vaccines with promising safety and efficacy both in preclinical and clinical studies. VLPs display the antigens in a repetitive high-density manner similar to the proteins of viral surface proteins, which contribute to strong T-cell and B-cell immune responses against the vaccine antigens [34]. Nanoparticles can play an adjuvant role in the vaccine formulation and improve the humoral and cellular immune responses. Among different type of nanoparticles, the calcium phosphate nanoparticle (CaPN) is a well-known member that has been used for many years as a delivery system in DNA vaccines and is approved to be utilized as the adjuvant.
Much research has been done on nanoparticles to design vaccine against T. gondii. Table 3 summarized the results of several different studies on nanoparticle vaccines, including the type of particle used for the SAG1, 2 and GRA1 antigen were muramyl dipeptide (MDP) microparticle [79], for MIC16 was yeast Saccharomyces cerevisiae EBY100 strain [68], and for ROP2, ROP18, MIC8, MIC3, ROP9, SAG2, SAG1ROP18, SAG1, SAG1, and AMA1 antigens were Mycobacterium bovis, Poly (lactideco—glycolide)(PLGA), virus-like particles (derived from baculovirus + influenza matrix protein 1), recombinant adenoviruses, PLGA, virus-like particle, polymeric nanospheres, virus-like particle, and virus-like particle respectively [80,81,82,83,84,85,86,87,88].
Discussion
In recent years, progress has been made in designing a potential vaccine against T. gondii. Studies have also been performed using different types of T. gondii antigens, including recombinant vaccines, DNA vaccines, subunit vaccines, attenuated live vaccines, and nanoparticle vaccines [20]. Accordingly, significant advances have been made in characterization and isolation of antigens, gene cloning, antigen expression, and immunological methods. In addition to the prevention strategies, new options are now needed to develop effective vaccines as a way to prevent the toxoplasmosis [29]. Most of the T. gondii antigens are important for the virulence and immunogenicity of the parasite. However, future studies should focus on the quality and quantity of antigens and identify potential candidate antigens against T. gondii infection. In addition, more extensive studies are needed to identify recombinant vaccines, DNA vaccine performance, and evaluate recombinant nanoparticle vaccines. Many of the vaccine strategies against toxoplasmosis have been experiments in animal models; nevertheless, these experiments only resulted in relative protection against T. gondii infection.
Vaccines designed with recombinant antigens rely on the defined antigens to induce a host-specific immune system against pathogenic microorganisms, which can be expressed by plasmids in the bacterial and yeast hosts or delivered by viral vectors [89]. Recombinant antigen vaccines have advantages over classic methods. One of the problems with using live vaccines is that if the host’s immune system is defective, it may cause the tachyzoite to return from an attenuated form to an active invasive form [29]. Another disadvantage is the complexity of obtaining sufficient amounts of purified immunogenic components of the antigen by the classical methods. The quality and effect of vaccination against T. gondii using recombinant antigens is very important, especially in pregnant women who may be at risk for the first time [6, 18, 20]. In the preparation of recombinant antigens, when the antigen is purified well, it can have far fewer side effects than raw antigens or live vaccines [90,91,92,93].
Studies on rodent animal models have shown that DNA vaccines can effectively induce both humoral and T-cell responses against a wide variety of candidate vaccine antigens [94]. Nevertheless, for unclear reasons, in primates and human’s poor immune, responses have been observed to parasite DNA vaccines. Various adjuvants including cytokines and CpG oligonucleotides have been studied to improve the immune responses in these large animals [95].
Monomeric linear protein that can assemble into a nanoparticle is a new method for inducing immune responses against peptide epitopes of antigens from an infectious agent. As most of the infectious agents invade the host through mucosal surfaces, researchers are interested to design vaccines with the ability to mimic this aspect of the pathogens to induce an effective immune response; therefore, a better understanding of the mechanisms that the pathogens use to interact with cells and the biological fluids is required to design vaccines with adequate efficiency. Monomeric linear protein based nanoparticles vaccine against toxoplasmosis were in a study and effectively elicited T-cell-dependent cellular immune cells responses [35]. As the group SAG antigens are highly expressed on the Toxoplasma gondii tachyzoite, they are considered as one of the main candidates for toxoplasmosis vaccine design. The SAG1 is a 30 KDa beta-glycoprotein that can be extracted from tachyzoites and sporozoites of the T. gondii [96].
The SAG1 is the most immunogenic structure in tachyzoites and the first structure of the parasite that interact with the host cells. The gene encoding this protein is a single-copy gene and contains no introns. It is believed that SAG1 is the most promising candidate to develop an effective vaccine against T. gondii because it stimulates both the cellular and humoral immune responses [6, 20, 37, 79, 96]. According to the results of previous studies use of vaccines based on recombinant forms of the antigen and nanoparticle-based vaccines can induce stable specific immunity in hosts, including pregnant women (at risk of primary infection) and immunocompromised patients [79]. Previous studies indicate that the use of GRA1, GRA2, GRA6, GRA5, GRA4, and GRA7 antigens are good candidate for the design and production of DNA vaccines. For example, the GRA7 antigen is an acidic 29 KDa protein and comprise about 0.5% of all the T. gondii proteins. The Gra7 gene is composed of 1.3 Kbps and has no introns. The GRA7 antigen is present in the parasitophorous vacuole in host cells infected by tachyzoites and also in the cytoplasm of host cells infected by bradyzoites. GRA7 is expressed in all of the stages of T. gondii infection and is a considerable candidate for the vaccine design [91, 93]. It can effectively induce both cellular and humoral immune responses against the T. gondii [29]. Another member of this family is GRA4 that is a 40 KDa protein secreted into the parasitophorous vacuole by the parasite [40, 41]. This protein strongly interacts with the milk IgA and to a lesser degree with the intestinal mucosal layer IgA [24]. The amino acid sequences 297–345 in the GRA4 are called C protein and can interact with the milk and intestinal mucosal IgA and serum IgG in mice infected by T. gondii and also serum IgG in human and sheep [24]. The GRA4 stimulates the mucosal T lymphocytes in BALB/C and CBA/J mice strains. The GRA4 can induce mucosal and systemic immune responses in mice after T. gondii ingestion [24, 29, 41]. Furthermore, GRA14 is a 47 KDa protein with 409 amino acids. The gene encoding this protein is consisting of 1227 bps. The GRA14 is present in the membranes of parasitophorous vacuole and intravacuolar network. This protein has a unique topology that is not seen in other proteins [29, 58, 97]. Due to the unique topology and its long length inside the vacuole system, this protein probably is a potential strong inducer of the immune responses. ROP proteins are the largest family of T. gondii serine-threonine kinases [24]. The evaluation of previous studies demonstrates that the ROP18 is the most interesting member of the ROP family in recombinant, nanoparticle, and DNA vaccine development studies [24, 64, 98]. It is probably due to the pre-formed presence of the antigen inside the rhoptry and as it is secreted into the parasitophorous vacuole during invasion to the host cells [64, 98, 99]. This antigen is one of the key virulence factors of T. gondii that protects the parasite from the host immune responses by its kinase activity [98]. The amino acids 243 to 539 are involved in the protein kinase activity of the protein. Another effect of this kinase activity is enhancing the parasite replication inside the host cells [99]. Previous studies have shown that GRA1, SAG1, SAG2, MIC1, MAG1, ROP18, GRA6, and GRA2 antigens are highly immunogenic. In addition, these antigens have been shown to stimulate specific antibodies in the host body or cytokines in vitro in the culture medium of splenocyte cells [6, 24, 29, 92]. ROP, GRA, and SAG antigens are the strongest candidates for the vaccine because they have been shown to contain relatively long antigenic fragments and regions, especially ROP, which appears to be a more suitable candidate than the other two antigens [6, 20]. It has also been shown that this antigen can elicit a strong protective immune response. DNA immunization of BALB/c mice with homogeneous mixtures of plasmids encoding short micronemic antigen fragments has been shown to enhance protective immunity, leading to an 85% reduction in the burden of T. gondii cysts [55].
References
Nazari N, Bozorgomid A, Janbakhsh A, Bashiri F (2018) Toxoplasma gondii and human immunodeficiency virus co-infection in western Iran: a cross sectional study. Asian Pac J Trop Med 11(1):58
Gilot-Fromont E, Lélu M, Dardé M-L et al (2012) The life cycle of Toxoplasma gondii in the natural environment. Toxoplasmosis-recent advances 10:2845
Halonen SK, Weiss LM (2013) Toxoplasmosis Handbook of clinical neurology 114:125–145. https://doi.org/10.1016/b978-0-444-53490-3.00008-x
Carruthers VB, Suzuki Y (2007) Effects of Toxoplasma gondii infection on the brain 33(3):745–751
Robbins JR, Zeldovich VB, Poukchanski A et al (2012) Tissue barriers of the human placenta to infection with Toxoplasma gondii. Infect Immun 80(1):418–428. https://doi.org/10.1128/iai.05899-11
Javadi Mamaghani, A, Tabaei SJS, Ranjbar MM et al (2020) Designing diagnostic kit for Toxoplasma gondii based on GRA7, SAG1, and ROP1 Antigens: an in silico strategy. Int J Peptide Res Therapeutics 26(4):2269–2283
Stelzer S, Basso W, Silván JB et al (2019) Toxoplasma gondii infection and toxoplasmosis in farm animals: risk factors and economic impact. Food Waterborne Parasitol 15:e00037
Konstantinovic N, Guegan H, Stäjner T et al (2019) Treatment of toxoplasmosis: current options and future perspectives. Food Waterborne Parasitol 15
Robert-Gangneux F, Dardé ML (2012) Epidemiology of and diagnostic strategies for toxoplasmosis. Clin Microbiol Rev 25(2):264–296. https://doi.org/10.1128/cmr.05013-11
Alday PH, Doggett JS (2017) Drugs in development for toxoplasmosis: advances, challenges, and current status. Drug Des Dev Ther 11:273–293. https://doi.org/10.2147/dddt.S60973
Montazeri M, Mehrzadi S, Sharif M et al (2018) Activities of anti-Toxoplasma drugs and compounds against tissue cysts in the last three decades (1987 to 2017), a systematic review. Parasitol Res 117(10):3045–3057. https://doi.org/10.1007/s00436-018-6027-z
Dubey JP, Lindsay DS, Speer CA (1998) Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clin Microbiol Rev 11(2):267–299
Black MW, Boothroyd JC (2000) Lytic cycle of Toxoplasma gondii. Microbiology and molecular biology reviews. Microbiol Mol Biol Rev 64(3):607–623. https://doi.org/10.1128/mmbr.64.3.607-623.2000
El Bissati K, Levigne P, Lykins J et al (2018) Global initiative for congenital toxoplasmosis: an observational and international comparative clinical analysis. Emerg Microbes & Infect 7(1):165. https://doi.org/10.1038/s41426-018-0164-4
Flegr J, Prandota J, Sovičková M, Israili ZH (2014) Toxoplasmosis--a global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries. PloS one 9(3):e90203. https://doi.org/10.1371/journal.pone.0090203
Ahady MT, Hoghooghi-Rad N, Madani R, Esmaeili Rastaghi AR (2018) Identification of Antigenic and immunogenic proteins of Toxoplasma gondii in human and sheep by immunoproteomics. Iran J Parasitol 13(1):39–48
Ferra B, Holec-Gąsior L, Grąźlewska W (2020) Toxoplasma gondii recombinant antigens in the serodiagnosis of toxoplasmosis in domestic and farm animals. Animals 10(8):1245. https://doi.org/10.3390/ani10081245
Naghi Vishteh M, Javadi Mamaghani A, Rashidi S et al (2020) Peptide-based monoclonal antibody production against SAG1 (P30) protein of Toxoplasma gondii. Monoclonal Antibodies in Immunodiagnosis and Immunotherapy 39(2):51–56
Ghaffarifar F, Jafarimodrek M, Vazini H et al (2019) Assessment of DNA vaccine encoding Toxoplasma gondii microneme complete gene and IL-12 as adjuvant in BALB/c mice. Iran J Basic Med Sci 22(8):901–907. https://doi.org/10.22038/ijbms.2019.34872.8276
Javadi Mamaghani, A, Fathollahi A, Spotin A (2019) Candidate antigenic epitopes for vaccination and diagnosis strategies of Toxoplasma gondii infection: a review. Microb Pathog 137:103788
Wang S, Zhang Z, Wang Y et al (2017) Toxoplasma gondii excretory/secretory antigens (TgESAs) suppress pro-inflammatory cytokine secretion by inhibiting TLR-induced NF-κB activation in LPS-stimulated murine macrophages. Oncotarget 8(51):88351–88359. https://doi.org/10.18632/oncotarget.19362
Arab-Mazar Z, Fallahi S, Yadegarynia D, Javadi Mamaghani A et al (2019) Immunodiagnosis and molecular validation of Toxoplasma gondii infection among patients with end-stage renal disease undergoing haemodialysis. Parasitology 146(13):1683–1689
Soldati D, Dubremetz JF, Lebrun M (2001) Microneme proteins: structural and functional requirements to promote adhesion and invasion by the apicomplexan parasite Toxoplasma gondii. Int J Parasitol 31(12):1293–1302
Daryani A, Kalani H, Sharif M et al (2013) Toxoplasma gondii: a review of excretory secretory antigens. J Mazandaran University of Medical Sci 22(2):220–232
Khan IA, Hwang S, Moretto M (2019) Toxoplasma gondii: CD8 T Cells Cry for CD4 Help. Front Cell Infect Microbiol 9:136. https://doi.org/10.3389/fcimb.2019.00136
Denkers EY, Gazzinelli RT (1998) Regulation and function of T-cell-mediated immunity during Toxoplasma gondii infection. Clin Microbiol Rev 11(4):569–588
Dupont CD, Christian DA, Hunter CA (2012) Immune response and immunopathology during toxoplasmosis. Semin Immunopathol 34(6):793–813. https://doi.org/10.1007/s00281-012-0339-3
Liu Q, Singla LD, Zhou H (2012) Vaccines against Toxoplasma gondii: status, challenges and future directions. Hum Vaccin Immunother 8(9):1305–1308. https://doi.org/10.4161/hv.21006
Karimi M, Tabaei SJS, Ranjbar MM, Fathi F, Jalili A, Zaminet Gh, Javadi Mamaghani, A al (2020) Construction of a synthetic gene encoding the multi-epitope of Toxoplasma gondii and demonstration of the relevant recombinant protein production: a vaccine candidate. Galen Med J 9:1708
Ambroise-Thomas P, Petersen E (2000) Congenital toxoplasmosis: past, present and future. In Congenital toxoplasmosis 1–7
Chaudhry SA, Gad N, Koren G (2014) Toxoplasmosis and pregnancy Canadian family physician 60(4):334–336
Verma R, Khanna P (2013) Development of Toxoplasma gondii vaccine: a global challenge. Hum Vaccin Immunother 9(2):291–293. https://doi.org/10.4161/hv.22474
Saadatnia G (2017) Toxoplasmosis infection in pregnant women. Sarem J Med Res 2(2):127–131
Lee DH, Lee SH, Kim AR, Quan FS (2016) Virus-like nanoparticle vaccine confers protection against Toxoplasma gondii. PloS one 11(8):e0161231
El Bissati K, Zhou Y, Dasgupta D et al (2014) Effectiveness of a novel immunogenic nanoparticle platform for Toxoplasma peptide vaccine in HLA transgenic mice. Vaccine 32(26):3243–3248
Gedik Y, İz SG, Can H et al (2016) Immunogenic multistage recombinant protein vaccine confers partial protection against experimental toxoplasmosis mimicking natural infection in murine model. Trials in Vaccinology 5:15–23
Wang Y, Yin H (2014) Research progress on surface antigen 1 (SAG1) of Toxoplasma gondii. Parasit Vectors 7:180. https://doi.org/10.1186/1756-3305-7-180
Qu D, Han J, Du A (2013) Enhancement of protective immune response to recombinant Toxoplasma gondii ROP18 antigen by ginsenoside Re. Exp Parasitol 135(2):234–239
Rashid I, Moiré N, Héraut B et al (2017) Enhancement of the protective efficacy of a ROP18 vaccine against chronic toxoplasmosis by nasal route. Med Microbiol Immunol 206(1):53–62
Martin V, Supanitsky A, Echeverria PC et al (2004) Recombinant GRA4 or ROP2 protein combined with alum or the gra4 gene provides partial protection in chronic murine models of toxoplasmosis. Clin Vaccine Immunol 11(4):704–710
Dziadek B, Gatkowska J, Grzybowski M et al (2012) Toxoplasma gondii: the vaccine potential of three trivalent antigen-cocktails composed of recombinant ROP2, ROP4, GRA4 and SAG1 proteins against chronic toxoplasmosis in BALB/c mice. Parasitology 131(1):133–138
Liu M, Yuan Z, Peng G et al (2010) Toxoplasma gondii microneme protein 8 (MIC8) is a potential vaccine candidate against toxoplasmosis. Parasitol Res 106(5):1079–1084
Dziadek B, Gatkowska J, Brzostek A et al (2009) Toxoplasma gondii: the immunogenic and protective efficacy of recombinant ROP2 and ROP4 rhoptry proteins in murine experimental toxoplasmosis. Exp Parasitol 123(1):81–89
Gatkowska J, Wieczorek M, Dziadek B et al (2018) Assessment of the antigenic and neuroprotective activity of the subunit anti-Toxoplasma vaccine in T. gondii experimentally infected mice. Vet Parasitol 254:82–94
Li W-S, Chen Q-X, Ye J-X et al (2011) Comparative evaluation of immunization with recombinant protein and plasmid DNA vaccines of fusion antigen ROP2 and SAG1 from Toxoplasma gondii in mice: cellular and humoral immune responses. Parasitol Res 109(3):637–644
Zulpo DL, Igarashi M, Sammi AS et al (2017) rROP2 from Toxoplasma gondii as a potential vaccine against oocyst shedding in domestic cats. Rev Bras Parasitol Vet 26(1):67–73
Haumont M, Delhaye L, Garcia L et al (2000) Protective immunity against congenital toxoplasmosis with recombinant SAG1 protein in a guinea pig model. Infect Immun 68(9):4948–4953
Zheng B, Lu S, Tong Q et al (2013) The virulence-related rhoptry protein 5 (ROP5) of Toxoplasma gondii is a novel vaccine candidate against toxoplasmosis in mice. Vaccine 31(41):4578–4584
Huang X, Li J, Zhang G et al (2012) Toxoplasma gondii: protective immunity against toxoplasmosis with recombinant actin depolymerizing factor protein in BALB/c mice. Exp Parasitol 130(3):218–222
Golkar M, Shokrgozar M-A, Rafati S et al (2007) Evaluation of protective effect of recombinant dense granule antigens GRA2 and GRA6 formulated in monophosphoryl lipid A (MPL) adjuvant against Toxoplasma chronic infection in mice. Vaccine 25(21):4301–4311
Ching XT, Fong MY, Lau YL (2016) Evaluation of immunoprotection conferred by the subunit vaccines of GRA2 and GRA5 against acute toxoplasmosis in BALB/c mice. Front Microbiol 7:609
Lunden A, Parmley S, Bengtsson KL, Araujo FJ (1996) Use of a recombinant antigen, SAG2, expressed as a glutathione-S-transferase fusion protein to immunize mice against Toxoplasma gondii. Parasitol Res 83(1):6–9
Czarnewski P, Araújo ECB, Oliveira MC et al (2017) Recombinant TgHSP70 immunization protects against Toxoplasma gondii brain cyst formation by enhancing inducible nitric oxide expression. Front Cell Infect Microbiol 7:142. https://doi.org/10.3389/fcimb.2017.00142
Sun XM, Zou J, A AE et al (2011) DNA vaccination with a gene encoding Toxoplasma gondii GRA6 induces partial protection against toxoplasmosis in BALB/c mice. Parasit Vectors 4:213. https://doi.org/10.1186/1756-3305-4-213
Angus C, Klivington-Evans D, Dubey J, Kovacs JA (2000) Immunization with a DNA plasmid encoding the SAG1 (P30) protein of Toxoplasma gondii is immunogenic and protective in rodents. J Infect Dis 181(1):317–324
Yuan ZG, Zhang XX, He XH et al (2011) Protective immunity induced by Toxoplasma gondii rhoptry protein 16 against toxoplasmosis in mice. Clin Vaccine Immunol 18(1):119–124
Pagheh AS, Sarvi S, Gholami S et al (2019) Protective efficacy induced by DNA prime and recombinant protein boost vaccination with Toxoplasma gondii GRA14 in mice. Microb Pathog 134:103601
Ahmadpour E, Sarvi S, Soteh MBH et al (2017) Enhancing immune responses to a DNA vaccine encoding Toxoplasma gondii GRA14 by calcium phosphate nanoparticles as an adjuvant. Immunol Lett 185:40–47
Yang WB, Zhou DH, Zou Y et al (2017) Vaccination with a DNA vaccine encoding Toxoplasma gondii ROP54 induces protective immunity against toxoplasmosis in mice. Acta Trop 176:427–432
Chen K, Wang JL, Huang SY et al (2017) Immune responses and protection after DNA vaccination against Toxoplasma gondii calcium-dependent protein kinase 2 (TgCDPK2). Parasite 24
Maraghi S, Ghadiri AA, Tavalla M et al (2019) Evaluation of immunogenicity and protective effect of DNA vaccine encoding surface antigen1 (SAG1) of Toxoplasma gondii and TLR-5 ligand as a genetic adjuvant against acute toxoplasmosis in BALB/c mice. Biologicals 62:39–49
Sobati H, Dalimi A, Kazemi B, Ghaffarifar F (2019) Evaluation of anti-Toxoplasma gondii immune responses in BALB/c mice induced by DNA vaccines encoding surface antigen 1 (SAG1) and 3 (SAG3). Mol Genet Microbiol Virol 34(1):59–66
Zheng B, Ding J, Lou D et al (2019) The virulence-related MYR1 protein of Toxoplasma gondii as a novel DNA vaccine against toxoplasmosis in mice. Front Microbiol 10:734
Chen Y, Yu M, Hemandez JA et al (2018) Immuno-efficacy of DNA vaccines encoding PLP1 and ROP18 against experimental Toxoplasma gondii infection in mice. Exp Parasitol 188:73–78
Ching XT, Fong MY, Lau YL (2017) Evaluation of the protective effect of deoxyribonucleic acid vaccines encoding granule antigen 2 and 5 against acute Toxoplasmosis in BALB/c Mice. Am J Trop Med Hyg 96(6):1441
Zhu WN, Wang JL, Chen K et al (2017) Evaluation of protective immunity induced by DNA vaccination with genes encoding Toxoplasma gondii GRA17 and GRA23 against acute toxoplasmosis in mice. Exp Parasitol 179:20–27
Vazini H, Ghafarifar F, Sharifi Z, Dalimi A (2018) Evaluation of immune responses induced by GRA7 and ROP2 genes by DNA vaccine cocktails against acute toxoplasmosis in BALB/c mice. Avicenna J Med Biotechnol 10(1):2
Wang LJ, Xiao T, Xu C et al (2018) Protective immune response against Toxoplasma gondii elicited by a novel yeast-based vaccine with microneme protein 16. Vaccine 36(27):3943–3948. https://doi.org/10.1016/j.vaccine.2018.05.072
Zhang Z, Li Y, Wang M et al (2018) Immune protection of rhoptry protein 21 (ROP21) of Toxoplasma gondii as a DNA vaccine against toxoplasmosis. Front Microbiol 9:909
Hu LY, Zhang NZ, Zhang FK et al (2017) Resistance to chronic Toxoplasma gondii Infection Induced by a DNA vaccine expressing GRA16. Biomed Res Int 2017:1295038. https://doi.org/10.1155/2017/1295038
Han Y, Zhou A, Lu G et al (2017) Protection via a ROM4 DNA vaccine and peptide against Toxoplasma gondii in BALB/c mice. BMC Infect Dis 17(1):1–9
Zhang Z, Li Y, Liang Y et al (2018) Molecular characterization and protective immunity of rhoptry protein 35 (ROP35) of Toxoplasma gondii as a DNA Vaccine. Vet Parasitol 260:12–21
Chu JQ, Huang S, Ye W et al (2018) Evaluation of protective immune response induced by a DNA vaccine encoding GRA8 against acute toxoplasmosis in a murine model. Korean J Parasitol 56(4):325
Zhou J, Wang L, Lu G et al (2016) Epitope analysis and protection by a ROP19 DNA vaccine against Toxoplasma gondii. Parasite 23
Xu XP, Liu WG, Xu QM et al (2019) Evaluation of immune protection against Toxoplasma gondii infection in mice induced by a multi-antigenic DNA vaccine containing TgGRA24, TgGRA25 and TgMIC6. Parasite 26
Lu G, Zhou A, Meng M et al (2014) Alpha-galactosylceramide enhances protective immunity induced by DNA vaccine of the SAG5D gene of Toxoplasma gondii. BMC Infect Dis 14(1):1–9
Desolme B, Mévélec MN, Buzoni-Gatel D, Bout D (2000) Induction of protective immunity against toxoplasmosis in mice by DNA immunization with a plasmid encoding Toxoplasma gondii GRA4 gene. Vaccine 18(23):2512–2521
Ducournau C, Nguyen TT, Carpentier R et al (2017) Synthetic parasites: a successful mucosal nanoparticle vaccine against Toxoplasma congenital infection in mice. Future Microbiol 12(5):393–405
Oledzka G, Bo L, Hiszczynska-Sawicka E et al (2017) Toxoplasma gondii: immunological response of sheep to injections of recombinant SAG1, SAG2, GRA1 proteins coupled to the non-toxic microparticle muramyl dipeptide. Small Rumin Res 150:111–117
Wang H, Liu Q, Liu K et al (2007) Immune response induced by recombinant Mycobacterium bovis BCG expressing ROP2 gene of Toxoplasma gondii. Parasitol Int 56(4):263–268
Nabi H, Rashid I, Ahmad N et al (2017) Induction of specific humoral immune response in mice immunized with ROP18 nanospheres from Toxoplasma gondii. Parasitol Res 116(1):359–370
Karthik L, Kumar G, Keswani T et al (2014) Protease inhibitors from marine actinobacteria as a potential source for antimalarial compound. PloS one 9(3):e90972
Zhang D, Jiang N, Chen Q (2019) Vaccination with recombinant adenoviruses expressing Toxoplasma gondii MIC3, ROP9, and SAG2 provide protective immunity against acute toxoplasmosis in mice. Vaccine 37(8):1118–1125
Allahyari M, Mohabati R, Vatanara A, Golkar M (2020) In-vitro and in-vivo comparison of rSAG1-loaded PLGA prepared by encapsulation and adsorption methods as an efficient vaccine against Toxoplasma gondii. J Drug Deliv Sci Technol 55:101327
Lee SH, Kang HJ, Lee DH et al (2018) Virus-like particle vaccines expressing Toxoplasma gondii rhoptry protein 18 and microneme protein 8 provide enhanced protection. Vaccine 36(38):5692–5700
Naeem H, Sana M, Islam S et al (2018) Induction of Th1 type-oriented humoral response through intranasal immunization of mice with SAG1-Toxoplasma gondii polymeric nanospheres. Artificial Cells, Nanomedicine, and Biotechnology 46(sup2):1025–1034
Choi WH, Park JS (2020) Immunogenicity and protective effect of a virus-like particle containing the SAG1 antigen of Toxoplasma gondii as a potential vaccine candidate for toxoplasmosis. Biomedicines 8(4):91
Kim MJ, Lee SH, Kang HJ et al (2020) Virus-like particle vaccine displaying Toxoplasma gondii apical membrane antigen 1 induces protection against T. gondii ME49 infection in mice. Microb Pathog 142:104090
Nascimento IP, Leite L (2012) Recombinant vaccines and the development of new vaccine strategies. Braz J Med Biol Res 45:1102–1111
Aghamolaei S, Kazemi B, Bandehpour M et al (2020) Design and expression of polytopic construct of cathepsin-L1, SAP-2 and FhTP16. 5 proteins of Fasciola hepatica. J Helminthol 94
Arab-Mazar Z, Fallahi S, Koochaki A et al (2016) Immunodiagnosis and molecular validation of Toxoplasma gondii-recombinant dense granular (GRA) 7 protein for the detection of toxoplasmosis in patients with cancer. Microbiol Res 183:53–59
Arab-Mazar Z, Fallahi S, Koochaki A et al (2016) Cloning, expression and immunoreactivity of recombinant Toxoplasma gondii GRA5 protein. Iran J Microbiol 8(5):331
Arab-Mazar Z, Seyyed-Tabaei SJ, Mirahmadi H (2014) Cloning of dense granular (GRA) 7 gene of Toxoplasma gondii into pTZ57RT vectors for sub-cloning in prokaryotic and eukaryotic plasmids. Novelty in Biomedicine 2(4):114–119
Foroutan M, Ghaffarifar F, Sharifi Z et al (2019) Rhoptry antigens as Toxoplasma gondii vaccine target. Clin Exp Vaccine Res 8(1):4–26. https://doi.org/10.7774/cevr.2019.8.1.4
Heegaard PMH, Dedieu L, Johnson N et al (2011) Adjuvants and delivery systems in veterinary vaccinology: current state and future developments. Adv Virol 156(2):183–202. https://doi.org/10.1007/s00705-010-0863-1
Jalallou N, Bandehpour M, Khazan H et al (2012) Evaluation of recombinant SAG1 protein for detection of Toxoplasma gondii specific immunoglobulin M by ELISA test. Iran J Parasitol 7(4):17–21
Ashrafi M, Sobati H, Tabaei SJ (2018) Construction and sequencing of dense granular14 (GRA14) gene of Toxoplasma gondii (RH) in expression prokaryotic plasmid PET32a: a preliminary study in vaccine production. J Appl Biotechnol Reps 5(2):75–80
Yin K, Zhao G, Xu C et al (2019) Prediction of Toxoplasma gondii virulence factor ROP18 competitive inhibitors by virtual screening. Parasit Vectors 12(1):98. https://doi.org/10.1186/s13071-019-3341-y
El Hajj H, Lebrun M, Arold ST et al (2007) ROP18 is a rhoptry kinase controlling the intracellular proliferation of Toxoplasma gondii. PLoS Pathog 3(2)
Acknowledgements
This study was supported by the Student Research committee, Kermanshah University of Medical Sciences, Kermanshah, Iran.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval
The protocol was approved by the Kermanshah University of Medical Sciences Ethics Committee [IR.KUMS.REC.1400.070].
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Mamaghani, A.J., Fathollahi, A., Arab-Mazar, Z. et al. Toxoplasma gondii vaccine candidates: a concise review. Ir J Med Sci 192, 231–261 (2023). https://doi.org/10.1007/s11845-022-02998-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11845-022-02998-9