
Abstract
Song complexity has been identified as one song characteristic important in female choice, whereas its role in male-male interactions is less clear and not much is known which song characteristics are specifically important for territorial defense. In Acrocephalus warblers, males seem to increase song complexity towards females but reduce it during territorial disputes. One possibility to reduce song complexity could be achieved by repeating individual syllables. In this context here we examined the importance of males repeating syllables in male-male interactions. We hypothesis that repeating syllables signals a males general fighting ability, aggressive status or willingness to attack. In a playback experiment we investigated the behavioral response of unmated, territorial Eurasian reed warbler (Acrocephalus scirpaceus) males towards two simultaneously singing intruders whereby the song of the two simulated intruders differs in the degree of syllable repetitions. The response of the territory owner was determined by using several behavioral parameters. Our results revealed that males approached faster and stayed significantly longer near the song with few syllable repetitions. However, the proportion of males approaching the song with high or low syllable repetitions first, as well as the minimal distance to which males approach to the two song types did not differ as well. Thus, the weaker response towards the high repetitive song playback suggests territorial males to be more intimidated by the aggressive nature of that song type.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
One important aspect in communication is song, which is used in intrasexual competition and mate choice across the animal kingdom including insects, frogs, birds and mammals (Bennet-Clark 1975; Kelley 2004; Catchpole and Slater 2008; Hoffmann et al. 2012; Tuschhoff and Wiens 2023). Both processes are important evolutionary forces related to sexual selection and known to influence the development and appearance of acoustic signals. In this context, a song may have a dual function, serving both, to attract females as well as to deter male competitors (Collins 2004; Leisler and Schulze-Hagen 2011; Baker et al. 2019; Tuschhoff and Wiens 2023).
In some bird species however, males use one song type to address females and a different one to address males (Catchpole 1973, 1983). This implies, that the requirements may differ for these two types of receivers. In many bird species males produce very complex songs, which are usually achieved by increasing the number of syllable types resulting in large syllable repertoires (Nowicki et al. 2000; Forstmeier et al. 2006; Kipper et al. 2006). Song complexity is suggested to be driven by female choice and somehow represents an acoustic peacock tail (Catchpole 1987). For example, in several Acrocephalus warbler species, males use very complex and diverse songs to attract females and simpler and shorter songs in male-male interactions. To attract females, male great reed warblers Acrocephalus arundinaceus (Linnaeus, 1758) produce a so-called “long song” which consists of several hundred syllables (Catchpole 1983; Bensch and Hasselquist 1992). In contrast, the “short song” is addressed towards males and consists of fewer, about 60 to 70 syllables. In the moustached warbler Acrocephalus melanopogon (Temminck, 1823) males have only one continuous song type but this song is usually introduced by a short and simple whistle part, which has been shown to be specifically addressed towards males (Fessl and Hoi 1996, 2000). Also, in the reed warbler Acrocephalus scirpaceus (Hermann, 1804), song is long and continuous (Catchpole 1973; Krištofík et al. 2014), but without clear structurally differentiated parts, which would indicate a possible separate sex specific function. Their song is very complex (Leisler and Schulze-Hagen 2011), but besides of that, males sing at a variable syllable rate (Hoi et al. 2023) and frequently repeat one syllable type before they switch to the next (Heuwinkel 1978; Leisler and Schulze-Hagen 2011).
Thus, the question arises, how important syllable repetitions are and in which context? One way to investigate the importance of syllable repetitions in intrasexual context is to examine the defense response of territorial males faced to intruders producing song with variable syllable repetitions. Therefore, we performed a playback experiment simulating two intruders with low and high syllable repetitions, presented at the same time. Assuming that varying levels of syllable repetitions signal different individual quality aspects of the sender (Vallet et al. 1998; Searcy and Beecher 2009) the two intruders may represent a different challenge and we expect a different response depending on the proportion of repeated syllables. Catchpole (1983) suggested in fact, that a weaker response can be expected, when the territory owner is intimidated by an aggressive song signal. In this context Catchpole (1983) could show that territorial males of the great reed warbler respond weaker towards male song signaling a higher aggression intensity. Thus, if an increase in syllable repetitions is mediated by an increase in the aggressive state of an individual, one may predict territorial males to be more intimidated and hence reveal a weaker response towards the high repetitive song playback.
Here we broadcast the two stimuli, significantly different in the degree of syllable repetitions, in close distance to one another, within the territory of an unmated male. In contrast to the single-speaker design, which is regularly used in acoustic challenges, in our two-speaker design the focal birds are simultaneously faced with two different intruders on their territory. Therefore, we assumed that the response is influenced by the degree of the threat both intruders represent. In this context we focused on behavioural parameters reflecting the differences in the response towards the two speakers. Given that male reed warblers defend relatively small territories and are frequently faced with several competitors simultaneously, the design resembles natural circumstances.
Materials and methods
Study species and study site
For the challenging experiments we used only unmated territorial Eurasian reed warbler (Acrocephalus scirpaceus) males. In this species, song plays an important role in female attraction but also male-male interactions and territorial defense (Catchpole 1973; Leisler and Schulze-Hagen 2011; Hoi et al. 2013). Furthermore, in an earlier study, we found evidence for individual-specific variation in male syllable rate and its influence on female egg investment (Krištofík et al. 2014). Eurasian reed warblers were investigated at fishponds near Veľké Blahovo, West Slovakia (48.052433 N, 17.598156 E) in the breeding season 2020. The area covers approximately 70 ha including three fishponds with reed beds mainly comprising Phragmites australis and partly Typha angustifolia, T. latifolia, and Carex spp.
Design of the playback experiments
At the beginning of April, male reed warblers arrive at our study site and start immediately to sing their continuous and complex song to signal territory ownership. Therefore, from mid of April to beginning of June, 2020 playback experiments were performed in the vicinity of singing territory owners but only during days with favorable weather conditions (no rain or heavy wind) and until 11:00 a.m. Once singing males were sighted, observations for a period of 30 min were done to determine male territories, based on their song posts, and their mating status, based on female presence and male song output. Only unmated territorial males were used for the experiments. Reed stands are often very dense and therefore have low visibility. For this reason, we used territories at reed /water edges, which allow a better observation of the birds during the experiments. Subsequently, we placed two black playback speakers (Technaxx Mini MusicMan, 5 cm x 5 cm) at a height of 1.5 m fixed on poles, within the territory in a distance of about 6–8 m from each other and facing parallel towards the territory owner. Two minutes after setting up the playback speakers both playbacks automatically started simultaneously as well as the behavioral observations. Therefore in order to improve the quality of the observations the same two observers hid at a distance of about 15 m (depending on vegetation cover and visibility) and documented the behavioral response of the focal territorial males. Observations are quietly dictated into one common voice recorder, and they were later transcribed. To document the behavioral response of the focal males 30 s interval sampling was used, because it is the most practical way to manage the amount of observational data and 30 s is the length where all the different behaviors (e.g. movements towards the two speakers, distance to the speakers, other behaviors and parameters) could be noticed. This method was already successfully used in former experiments (e.g. Hoi et al. 2013; Darolová et al. 2020).
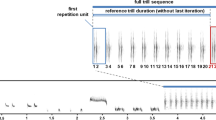
To produce the playbacks, we used a pool of song recordings of 74 reed warbler males, which were recorded at the same locality and already analyzed in previous years. For the experiments we selected recordings of four males showing the highest and four males showing the lowest syllable repetitions. We presented all four males with the highest and lowest syllable repetitions combined into a single playback rather than presenting single male song playbacks which has pros and cons. Playbacks of single males would increase the variance in the behavioral response due to the quality differences between randomly assigned song playbacks and the quality of the focal male confronted. Thus, an advantage of the multi-male design is that all focal males are confronted with standardized song set including a broader spectrum of song features. A disadvantage of this design is that in a short period the focal male experiences song of several males from the same place. The playback time was in total four min in length and each male was represented by 60 s of song. In our breeding area territorial males are confronted with several, up to seven unmated males singing simultaneously, in close distance (personal observation). Thus, to advertise four males in a row is in the range of a natural situation. In the present study, stimuli songs were unmanipulated whereas some playback studies used artificially manipulated songs. One drawback of our design could be that there are other correlated parameters being of importance as well. On the other hand, creating a song artificially also carries some risks of producing song patterns (e.g. sequences of syllables) which may be odd or of uncertain importance and therefore result in false interpretations. For this reason, we decided to use the more natural approach with its weaknesses. For the high syllable repetition playback type, the selected males, repeated on average a third less than in the low syllable repetition playback type (Table 1) with no overlap in its range and the male with the most song repetitions in the low repetition group still repeated 30 syllables less than the male with the least repetitions in the high repetition group. In contrast parameters describing song complexity did not vary and overlapped between groups. For example, parameters describing syllable diversity like the number of syllable switches (determined as the sum of changes from one syllable type to another during 30 s of song; for details see Krištofík et al. 2014), did not differ between the playback types and was for the high syllable repetition playback type (Table 1) Also, the number of new syllable types (determined as the sum of new syllable types appearing every six seconds, over a period of 30 s; Krištofík et al. 2014) is similar between the high syllable repetition playback type (Table 1). However, the average number of new syllables a male produces / 30 s slightly differs between the high syllable repetition playback type as the range distribution touches each other (Table 1). This complexity variable is a mixture between an innovative and a frequency aspect and is the sum of new syllable types multiplied by their frequency at every six-seconds over 30 s of song (see Kristofik et al. 2014). Moreover, single correlations between syllable repetitions and the three diversity parameters of the eight males used in the playbacks revealed a significant correlation only between syllable repetitions and the number of new syllables (r = 0.89, p = 0.003, n = 8), but not for the two other variables (for both p > 0.3). Given the low sample size the positive relationship must be treated with care but indicate, that opposite to the prediction, repetitions and the number of new elements increase together.
To determine the territorial response of the challenged males, the following behavioral parameters were collected: (i) to which playback type each territory owner approached first, defined as the first movement of the territory owner by at least 1 m towards one speaker, while at the same time increasing the distance to the other speaker, (ii) latency of the first approach, given as the 30 s interval (ranging from 1 to 8) in which the territory owner approached the speaker chosen, (iii) distance (m) of the focal male to both speakers for each 30 s interval, which was used to derive the nearest approach to each speaker, and (iv) the time (number of 30 s intervals) the focal male spent nearer to one of the speakers.
We removed experiments from the analyses when (i) males did not show a side preference to either of the two speakers, e.g. when a male remained at the same distance from speakers throughout the whole experiment (n = 2), or (ii) when the response of the focal male was disturbed by territory neighbors (n = 3), approaching females, or great reed warblers (Acrocephalus arundinaceus). Thus, of the 52 experiments (males tested), five (9.62%) experiments were excluded from the analyses.
Statistical analyses
We examined the birds’ response to the two speakers in terms of which speaker they chose to approach first, latency in approach, nearest approach distance and time spent near each speaker. Since we could not expect a general pattern, we considered these as four independent response variables. We checked each of the variables for stationarity during the season using a simple linear model with time as the independent variable (always insignificant, R package mgcv, Wood 2011). A binomial regression analyses was used (R package mgcv, Wood 2011) to determine the difference in the number (%) of males approaching, the latency of approach and the time spent near the two playback types (here individual was introduced as a random effect). To determine the difference regarding the order of response to either playback type a χ2 -test, and to determine the differences in the approach distance, a paired t-test was used. Furthermore, a Breusch-Pagan-test (R package olsrr, Hebbali 2020) applied to determine whether there is a seasonal change of the variance in male response towards playback types. For the other variable visual inspection did not indicate a change in the variance over time. The statistical software package R (R Core Team 2023) and IBM SPSS Statistics 27 has been used for the analyses.
Results
Of the 46 males, 28 (60.9%), approached the low repetition playback first, and 18 (39.1%) the high repetition playback (Fig. 1a). Based on a binomial regression analysis this difference is not significant (estimate = -0.44, SE = 0.30, p = 0.14).

a First approach: Percentage (± 95% confidence intervals) of males which either first approached the song type with high (n = 18) or low (n = 28) repetitions, and b latency of approach: The average ± SE time in terms of the number of 30 s intervals it took until males approached the song type with high (n = 18) or low (n = 28) repetitions for the first time
Of the 28 males first approaching the low repetition playback, nine (32.1%) also later moved towards the high repetition playback and from the 18 males which approached the high repetition playback first, six (33.33%) subsequently also approached the speaker playing song with low syllable repetitions. This difference regarding the order of response to either playback type is not significant (χ2 -test: χ2 = 0.024, df = 45, p = 0.874).
Regarding the time it took focal males to approach a speaker for the first time (latency of approach) our results revealed a clear deviation from an equal distribution (χ2 = 119.5, df = 7,45, p < 0.0001). Most males reacted very quickly, namely in the first 30 s interval, whereby the proportion was 78.6% (22/28) for the low and 44.4% (8/18) for the high repetition playback. Thus male response towards playbacks seemed to be faster to the low, than the high repetition playback (see Fig. 1b), although this difference was only marginal (estimate = 0.36, SE = 0.19, p = 0.06, df = 45).
Regarding the nearest approach towards the different playback types the nearest distance at which focal males have been observed from the two playback speakers did not significantly differ (pairwise t-tests: t = -1.19, p > 0.24, df = 36, Fig. 2a), though, males tended to, approach closer to the low repetition playback than the high repetition playback.

a Nearest distance males approach in relation to playback type and b time males spent near each playback type. Given are the average (± SE) nearest distance (m) the focal male approached the song type with high (n = 18) or low (n = 20) repetitions during the four minutes of playback presentation and the average (+ SE) time in terms of number of 30 s intervals males spent closer to the song type with high (n = 18) or low (n = 28) repetitions, during the four minutes of playback presentation
When examining the time (number of 30-sec intervals) that territory owners spent near the two playback speakers respectively, the length of stay close to the two playback types is negatively correlated (T-test: r = -0.65, t = -5.67, df = 45, p < 0.0001,) and males spent significantly longer near the low than the high repetition playback (Estimate = 1.41, SE = 0.60, p < 0.020, linear model with individuals as random effects, Fig. 2b).
However, there was a seasonal change of the variance in male response towards playback types (Breusch Pagan test): χ2 = 4.78, p = 0.028, df = 1,45): at the beginning of the season the difference of male response between low and high repetition playback is bigger namely males approached nearer to one than to the other speaker (in both directions) but this difference decreases with the season and even do not discriminate playbacks at all (Fig. 3).

Difference in the nearest approach of focal males (n = 37) towards the high and the low syllable repetition playback. For each male the difference is calculated as the nearest distance to the high– the nearest distance to the low syllable repetition playback in m
Discussion
There are a variety of studies investigated the importance of song characteristics in relation to sexual selection (Ritchie 1996; Briefer et al. 2010; Charlton and Reby 2016; Wiens and Tuschhoff 2020; Davranoglou et al. 2023). In birds these include e.g. pitch, frequency range, song amplitude, or parameters describing song complexity like syllable type switching, song rate or syllable repertoire size which seem specifically important in mate choice and male-male competition (for a review see Searcy and Beecher 2009; Linhart et al. 2013). In particular, parameters reflecting vocal complexity have been shown to be important in mate choice and in this context also reflect aspects of individual quality (Catchpole 1980), but there is less evidence for the importance of song simplicity, e.g. reflected in high syllable repetitions for male-male interactions (Spencer et al. 2003; Nowicki and Searcy 2004). In Acrocephalus warbler males seem to sing less complex during territorial disputes (Catchpole 1983) and reduce song output as well as complexity soon after mating (own unpublished observations). Therefore we would expect more simple and repetitive song to signal aggressive motivation and to be preferentially used in territorial disputes and males to respond accordingly. One possibility to reduce song complexity could be achieved by repeating individual syllables and in line with these songs used for the two playback types mainly differ in syllable repetitions. Intuitively one may expect that with increasing syllable repetitions syllable diversity would decrease and the other way round, but this did not seem to be the case (see methods). Parameters describing syllable diversity including new syllable types, as well as syllable switches (for details see methods), did not obviously differ between playback types but the number of new syllables revealed also being different between the high and low repetitive song types. Furthermore, we found a positive correlation between syllable repetitions and number of new syllables. These results have to be treated with care because they are only representative for the song of males selected for the playbacks. Intuitively one may predict that with increasing repetitions syllable diversity should decrease but this negative correlation is not necessary an implication. The variable “number of new syllables” is a composite, namely how often new syllables occur and how often they are repeated. Thus, the number of new syllables can increase and at the same time also the repetition rate. Furthermore, also song speed and syllable duration can influence the direction of that relationship. Thus, the prediction is not necessarily violated by a positive relation between the two variables. The two real diversity parameters do not seem to differ between the playback groups. Thus, syllable diversity could also influence a male´ s response but given that we selected the most extreme songs regarding syllable repetitions it is still very likely that differences in the behavioral response of territorial males are mainly due to differences in syllable repetitions.
In chaffinches (Fringilla coelebs Linnaeus, 1758) song-type switching rate seems an agonistic signal. Territorial males exhibit a stronger response towards rivals that repeat song-types more frequently rather than switch to a different (Deoniziak and Osiejuk 2020). Also in our study we found evidence for the importance of syllable repetitions in relation to the response of male European reed warblers to song playbacks of conspecific intruders. Our results show that territorial males differently respond when faced to low and high repetitive song which is mainly revealed by the difference, in the time males remained near playbacks. Males spent significantly longer near the low than the high repetition playback (Fig. 2b), a similar result was also found by Vehrencamp et al. (2007) in banded wrens (Thryothorus pleurostictus Sclater, 1870). They stayed longer close to a simulated intruder singing less repetitive song. In conclusion, male European reed warblers responded stronger when the territory intruder sings in a less repetitive way. This is in line with findings by Catchpole (1983) on the Great reed warbler. He suggests that a weaker response represents a stronger territorial threat, because the territory owner is intimidated by an aggressive vocal signal. In this context, the weaker response of our territorial European reed warblers towards songs with higher syllable repetitions may be interpreted as an indication that the highly repetitive song is more intimidating and higher syllable repetitions may play a role in intrasexual contest and territory defense (Catchpole 1973). However, there might be plasticity in the territorial response to playbacks. One explanation for the variable response of the territorial owners could be related to season, e.g., the response could change with the time of the season (Dittami 1981). In line with this we found an influence of season on the difference in the response towards the two playback types (Fig. 3). It seems that with the course of the breeding season male approach less close to the speaker which suggest territorial response is getting weaker with the season (Fig. 3). This seasonal effect, however, does not influence the general interpretation of our results. It may partly explain why the difference in some of the response parameters, is not or only marginally significant. Including later experiments and the response of males later in the season may partly reduce the effect of playback type. The difference in the response (Fig. 3) also indicates that, the strength of the response is independent of the playback type. Thus, each male seems to react either to the one or the other playback type. Furthermore, one reason as to why more but not significantly more males approach the low repetition playback first, assuming it reflects a weaker opponent, could be also a strategic one (Hoi et al. 2023). Whom to approach (repel) first might not be a fixed rule but may depend on the quality of the opponents (Dabelsteen and McGregor 1996; Temeles and Kress 2010), e.g. the quality of the territory owner in comparison to the quality of the two intruders, whereby quality in the reed warblers is signaled by specific song features like syllable repetitions or the appearance of new syllables. This may explain why not all males respond in the same manner. E.g. male approach to the low and high repetition playback type is ambiguous, as only slightly more males approached the low repetition playback first and there is no difference in approach distance between song types (Fig. 2a). Furthermore, after approaching one intruder (playback type) a proportion of males subsequently moved towards the second intruder (other playback type) (Fig. 1a). Thus, the question remains, which factors influence this decision-making process?
Given songs selected for the playbacks are at the extreme ends (with many or few syllable repetitions), song structure of most territorial males is probably in between and males will experience them as extremes.
Regarding their response, males in principle have two options: (1) to first respond towards the weaker opponent, or (2) to first expel a more serious competitor (Searcy and Beecher 2009). Thus, the assumption would be that a male producing a relatively high number of syllable repetitions may dare to conquer the high repetition playback type, and a high proportion may then subsequently move towards the weaker (low repetition playback type) intruder. In contrast, most males singing in a less repetitive way may first move towards the low repetition playback but may be less keen to subsequently approach the high repetition playback type. However, our results do not support this assumption. The proportion of males moving in a second step to the alternative playback type did not significantly differ. The proportion of males which first approached the low repetition playback (about 61.7%) and later also moved towards the high repetition playback (19.1%) did not differ (see results) from the proportion of males approaching first the high repetition playback (38.3%) and subsequently moved to the speaker producing low repetition playback (33.3%).
Examining the nearest approach distance, which is frequently used as an indicator for the strength of the response (Krebs et al. 1978; Yasukawa 1981; Osiejuk et al. 2004; Hutfluss et al. 2021) of male territorial reed warblers to the two playback types revealed no difference between low and high repetition playback type (Fig. 2a). This may imply that a song with high syllable repetitions is not more intimidating than a low repetition playback and the threat to the territory owner may not significantly differ between intruders producing a low and high repetition song.
However, an alternative not mutual exclusive explanation could be, that broadcasting conditions e.g. background noise influence whether syllables are more or less frequently repeated (Wickler and Seibt 1979). Broadcasting conditions, as a consequence of vegetation density and structure are in general less favorable in marsh habitats (Cosens and Falls 1984) and hence such a function is not unlikely but may not explain the differences found in our experiment although could be the origin for a sexual selected trait. In particular to broadcast information to male competitors and ensure that the information reaches the recipient may be more important than to lose syllables when displaying for females.
In conclusion our main finding is that territorial male reed warblers respond in general more strongly to the less than high repetitive and more complex song. Although males are able to clearly discriminate between the two song types, are the results far from being clearcut. The direction of the behavioral response of territorial males, based on the different parameters was in general consistent, but the differences appear rather weak, given the extreme traits presented and the extreme resulting differences. The results suggest that response is not gradually changing between males they rather strongly respond either to the low or the high repetitive playback type. Thus, a condition dependent strategy is likely the reason for that result. The intrinsic quality (dominance) of the receiving, relative to that of the signaling individual might be responsible for the decision whom to approach first. In line with this, future studies would be necessary to clarify the importance of individual quality of territorial owners. Only one response parameter, namely the time males spent near the two playback types significantly differed between the two playback types. Given that result one may alternatively conclude that the motivation to join the less repetitive song is not aggressively motivated. In line with this, it would be also interesting for future studies to explore the motivation driving male´s searching the neighborhood of a conspecific producing one or the other song type.
References
Baker CA, Clemens J, Murthy M (2019) Acoustic pattern recognition and courtship songs: insights from insects. Annu Rev Neurosci 42:129–147. https://doi.org/10.1146/annurev-neuro-080317-061839
Bennet-Clark HC (1975) Sound production in insects. Sci Prog 62:263–283
Bensch S, Hasselquist D (1992) Evidence for active female choice in a polygynous warbler. Anim Behav 44:301–311. https://doi.org/10.1016/0003-3472(92)90036-9
Briefer E, Vannoni E, McElligott AG (2010) Quality prevails over identity in the sexually selected vocalisations of an ageing mammal. BMC Biol 8:35. https://doi.org/10.1186/1741-7007-8-35
Catchpole CK (1973) The functions of advertising song in the sedge warbler (Acrocephalus schoenobaenus) and the reed warbler (A. scirpaceus). Behaviour 46:300–320. https://doi.org/10.1163/156853973X00067
Catchpole CK (1980) Sexual section and the evolution of complex songs among European warblers of the genus Acrocephalus. Behaviour 74:147–166. https://doi.org/10.1163/156853980X00366
Catchpole CK (1983) Variation in the song of the great reed warbler Acrocephalus arundinaceus in relation to mate attraction and territorial defence. Anim Behav 31:1217–1225. https://doi.org/10.1016/S0003-3472(83)80028-1
Catchpole CK (1987) Bird song, sexual selection and female choice. Trends Ecol Evol 2:94–97. https://doi.org/10.1016/0169-5347(87)90165-0
Catchpole CK, Slater PJB (2008) Bird song: Biological themes and variations, 2nd edn. University, Cambridge. https://doi.org/10.1017/CBO9780511754791
Charlton B, Reby D (2016) The evolution of acoustic size exaggeration in terrestrial mammals. Nat Commun 7:12739. https://doi.org/10.1038/ncomms12739
Collins SA (2004) Vocal fighting the functions of birdsong. In: Marler P, Slabbekoorn H (eds) Nature’s music. Elsevier Academic, San Diego, pp 39–79
Cosens SE, Falls JB (1984) A comparison of sound propagation and song frequency in temperate marsh and grassland habitats. Behav Ecol Sociobiol 15:161–170. https://doi.org/10.1007/BF00292970
Dabelsteen T, McGregor PK (1996) Dynamic acoustic communication and interactive playback. In: Kroodsma DE, Miller EH (eds) Ecology and Evolution of Acoustic Communication in Birds. Cornell University Press, New York, pp 398–408
Darolová A, Krištofík J, Knauer F, Hoi H (2020) Behavioural response of Eurasian blackcaps to acoustically simulated conspecific and heterospecific male intruders. J Ornithol 161:447–458. https://doi.org/10.1007/s10336-019-01743-x
Davranoglou L-R, Taylor GK, Mortimer B (2023) Sexual selection and predation drive therepeated evolution of stridulation in Heteroptera and other arthropods. Biol Rev 98:942–981. https://doi.org/10.1111/brv.12939
Deoniziak K, Osiejuk TS (2020) Song-type switching rate in the chaffinch carries a message during simulated intrusion. Behav Ecol Sociobiol 74:e47. https://doi.org/10.1007/s00265-020-2825-2
Dittami J (1981) Seasonal changes in the behavior and plasma titers of various hormones in barheaded geese, Anser indicus. Z Tierpsychol 55:289–324. https://doi.org/10.1111/j.1439-0310.1981.tb01274.x
Fessl B, Hoi H (1996) The significance of a two part song in the moustached warbler (Acrocephalus melanopogon). Ethol Ecol Evol 8:265–278. https://doi.org/10.1080/08927014.1996.9522917
Fessl B, Hoi H (2000) Song complexity and song structure in the Moustached Warbler Acrocephalus melanopogon. J Avian Biol 31:144–150. https://doi.org/10.1034/j.1600-048x.2000.310206.x
Forstmeier W, Hasselquist D, Bensch S, Leisler B (2006) Does song reflect age and viability? A comparison between two populations of the great reed warbler Acrocephalus arundinaceus. Behav Ecol Sociobiol 59:634–643. https://doi.org/10.1007/s00265-005-0090-z
Hebbali A (2020) Olsrr: tools for building OLS regression models. Version 0.5.3. https://CRAN.R-project.org/package=olsrr
Heuwinkel H (1978) Der Gesang des Teichrohrsängers (Acrocephalus scirpaceus) unter besonderer Berücksichtigung der Schalldruckpegel-(Lautstärke-) Verhältnisse. J Ornithol 119:450–461. https://doi.org/10.1007/BF01643136
Hoffmann F, Musolf K, Penn DJ (2012) Ultrasonic courtship vocalizations in wild house mice: spectrographic analyses. J Ethol 30:173–180. https://doi.org/10.1007/s10164-011-0312-y
Hoi H, Krištofík J, Darolová A (2013) Experimentally simulating paternity uncertainty: immediate and long-term responses of male and female Reed Warblers Acrocephalus scirpaceus. PLoS ONE 8:e62541. https://doi.org/10.1371/journal.pone.0062541
Hoi H, Darolová A, Krištofík J (2023) Slow song syllable rates provoke stronger male territorial responses in Eurasian Reed warblers (Acrocephalus scirpaceus). J Ornithol 164:193–202. https://doi.org/10.1007/s10336-022-02021-z
Hutfluss A, Rohr VA, Scheidt S, Steinbichl L, Bermúdez-Cuamatzin E, Slabbekoorn H, Dingemanse NJ (2021) Does song overlap signal aggressiveness? An experimental study with repeated measures in free-ranging great tits. Anim Behav 179:199–211. https://doi.org/10.1016/j.anbehav.2021.07.006
Kelley DB (2004) Vocal communication in frogs. Curr Opin Neurobiol 14:751–757. https://doi.org/10.1016/j.conb.2004.10.015
Kipper S, Mundry R, Sommer C, Hultsch H, Todt D (2006) Song repertoire size is correlated with body measures and arrival date in common nightingales, Luscinia megarhynchos. Anim Behav 71:211–217. https://doi.org/10.1016/j.anbehav.2005.04.011
Krebs JR, Ashcroft R, Webber M (1978) Song repertoires territory defense in the great tit. Nature 271:539–542. https://doi.org/10.1038/271539a0
Krištofík J, Darolová A, Majtan J, Okuliarová M, Zeman M, Hoi H (2014) Do females invest more into eggs when males sing more attractively? Postmating sexual selection strategies in a monogamous reed passerine. Ecol Evol 4:1328–1339. https://doi.org/10.1002/ece3.1034
Leisler B, Schulze-Hagen K (2011) The reed warblers. Diversity in a uniform bird family. KNNV Publishing, Zeist
Linhart P, Jaška P, Petrusková T, Petrusek A, Fuchs R (2013) Being angry, singing fast? Signalling of aggressive motivation by syllable rate in a songbird with slow song. Behav Processes 100:139–145. https://doi.org/10.1016/j.beproc.2013.06.012
Nowicki S, Searcy WA (2004) Song function and the evolution of female preferences: why birds sing, why brains matter. Ann N Y Acad Sci 1016:704–723. https://doi.org/10.1196/annals.1298.012
Nowicki S, Hasselquist D, Bensch S, Peters S (2000) Nestling growth and song repertoire size in great reed warblers: evidence for song learning as an indicator mechanism in mate choice. Proc R Soc B 267:2419–2424. https://doi.org/10.1098/rspb.2000.1300
Osiejuk TS, Ratyńska K, Cygan JP (2004) Signal value of alternating and overlapping singing in the Yellowhammer Emberiza citrinella. J Ethol 22:55–61. https://doi.org/10.1007/s10164-003-0099-6
Ritchie MG (1996) The shape of female mating preferences. Proc Natl Acad Sci USA 93:14628–14631. https://doi.org/10.1073/pnas.93.25.14628
Searcy WA, Beecher MD (2009) Song as an aggressive signal in songbirds. Anim Behav 78:1281–1292. https://doi.org/10.1016/j.anbehav.2009.08.011
Spencer KA, Buchanan KL, Goldsmith AR, Catchpole CK (2003) Song as an honest signal of developmental stress in the zebra finch (Taeniopygia guttata). Horm Behav 44:132–139. https://doi.org/10.1016/S0018-506X(03)00124-7
Temeles EJ, Kress WJ (2010) Mate choice and mate competition by a tropical hummingbird at a floral resource. Proc Roy Soc B: Biol Sci 277:1607–1613. https://doi.org/10.1098/rspb.2009.2188
Tuschhoff E, Wiens JJ (2023) Evolution of sexually selected traits across animals. Front Ecol Evol 11:1042747. https://doi.org/10.3389/fevo.2023.1042747
Vallet E, Beme I, Kreutzer M (1998) Two-note syllables in canary songs elicit high levels of sexual display. Anim Behav 55:291–297. https://doi.org/10.1006/anbe.1997.0631
Vehrencamp SL, Hall ML, Bohman ER, Depeine CD, Dalziell AH (2007) Song matching, overlapping, and switching in the banded wren: the sender’s perspective. Behav Ecol 18:849–859. https://doi.org/10.1093/beheco/arm054
Wickler W, Seibt U (1979) Duetting: a daily routine of Laniarius funebris, the Slate-coloured Boubou (Aves, Laniidae). Z Tierpsychol 51:153–157. https://doi.org/10.1111/j.1439-0310.1979.tb00679.x
Wiens JJ, Tuschhoff E (2020) Songs versus colours versus horns: what explains the diversity of sexually selected traits? Biol Rev 95:847–864. https://doi.org/10.1111/brv.12593
Yasukawa K (1981) Song repertoires in the red-winged blackbird (Agelaius phoeniceus): a test of the Beau Geste hypothesis. Anim Behav 29:114–125. https://doi.org/10.1016/S0003-3472(81)80158-3
Acknowledgements
This work was supported by the VEGA Scientific Grant Agency of the Ministry of Education, Science, Research and Sport of the Slovak Republic and the Slovak Academy of Sciences under Grant 02/0065/20 (AD, JK, LR).
Funding
Open access funding provided by The Ministry of Education, Science, Research and Sport of the Slovak Republic in cooperation with Centre for Scientific and Technical Information of the Slovak Republic
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest. Experiments comply with the current laws of Slovak Republic.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Darolová, A., Krištofík, J., Rubáčová, L. et al. Is syllable repetitions a song parameter important for male-male interactions in Eurasian reed warbler (Acrocephalus scirpaceus)?. Biologia 79, 1451–1459 (2024). https://doi.org/10.1007/s11756-024-01636-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11756-024-01636-2