
Abstract
Polyamines (PAs) are related to many physiological processes, including soil drought stress. Two yellow lupin ‘Morocco 4’ (drought tolerant) and ‘Taper’ (drought sensitive) were exposed to soil drought for 2 weeks. The half of the examined plants were additionally sprayed with a solution of polyamine biosynthesis inhibitor—dl-α-difluoromethylarginine (DFMA). Yellow lupin leaves showed a 19% increase and seeds a 54% decrease in the total PA contents. The seeds contained fourfold less PAs than the leaves under drought conditions. The highest amount of spermidine and lack of agmatine were found in the leaves, while in the seeds the highest content of spermine and the presence of agmatine was confirmed. The use of DFMA under drought conditions decreased the content of spermine in ‘Morocco 4’ and ‘Taper’ (41 and 19%, respectively) and spermidine in ‘Taper’ (by 13%), as well as reduced two out of three of the yield components. More tolerant ‘Morocco 4’, after DFMA treatment was characterized by a higher spermidine and spermine content and a smaller decrease in yield components compared to the less tolerant ‘Taper’. Simultaneously subjecting plants to soil drought and DFMA treatment caused in ‘Morocco 4’ a decline in the number of pods and seeds per plant and seeds dry weight per plant (64, 50 and 54%, respectively), while in ‘Taper’ a reduction of the number of pods per plant and seeds per pod (32 and 27%, respectively) was observed. These results confirm that PAs are involved in yellow lupin tolerance and may play a protective function under soil drought conditions.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Droughts are the most complex phenomena and their causes and effects are still not well known and understood. In the last 30 years, droughts have occurred more often throughout the world, causing great social and economic damages. Droughts are becoming increasingly frequent; they are more intense and cover larger areas. Soil drought seriously limits plant growth, which leads to a significant decrease in crop yield. It can reduce crop yield even up to 50%. The availability of water is in fact one of the most important factors increasing plant yield.
A group of plants particularly sensitive to adverse weather conditions are legume plants, including yellow lupin. Legumes are known for their characteristic flower structure and ability to interact with rhizobia as well as their importance to humans (Graham and Vance 2003). For decades, legumes have been used in animal nutrition, mainly to increase the protein concentration in the fodder. Legume plants contain approximately 30% of protein, with the most valuable amino acids, essential for the proper functioning of animal organisms. Therefore, they constitute a valuable source of protein in the human diet. Unfortunately, improvement in legume crop yields is inferior to those of cereals. The reasons are unfavorable environmental conditions under which many legume species are grown. Furthermore, the problem of water shortages for legumes is getting worse along with the increasing number of areas affected by drought (Postel 2000). For legumes, the most dangerous are water shortages in critical phases, which are periods of the greatest biomass increase as well as in the phase of generative organ formation. In legumes, drought reduces yield by premature and insufficient seed filling, and as a result of flower and young pod rejections. There is a vital need to increase drought tolerance in legumes (Graham and Vance 2003).
Polyamines (PAs) are low molecular organic cations occurring in a wide range of animal and plant organisms (Hussain et al. 2011). With respect to plant growth and development, PAs are widely involved in cell division and differentiation, morphogenesis, secondary metabolism, senescence, apoptosis, DNA synthesis, gene transcription, protein translation and chromatin organization (Yang et al. 2007; Alet et al. 2012; Tavladoraki et al. 2012). In recent years, the role of PAs in plants against abiotic and biotic stresses has been thoroughly investigated (Liu et al. 2006). A generally observed fact is that PA concentration change under various types of environmental stresses including drought stress. Stress-tolerant plants are characterized by a high capacity of increasing PA biosynthesis in response to stress that can be observed as a two- to threefold increase of endogenous PA levels compared to unstressed plants (Kasukabe et al. 2004). PAs play important role in the stabilization of biologically active polyanionic compounds, such as cytoplasmic membrane phospholipids, proteins (including enzymes) or nucleic acids. These compounds change their activity by interactions with amino groups and other molecules (Hura et al. 2015). Due to their polycationic nature, PAs occur in cells in the free form, as well as soluble/insoluble conjugates (Gemperlová et al. 2006). PAs can reduce the damage of membrane phospholipids resulting from increased activity of lipoxygenase under stress (Lester 2000). PAs are responsible for scavenging reactive oxygen species (ROS) and can indirectly influence the activity of catalase, peroxidases or superoxide dismutase. PA catabolism in plants is strongly associated with the production of hydrogen peroxide, because H2O2 is the frequent product of the polyamine metabolic pathway (Moschou et al. 2012). Additionally, PA catabolism-derived H2O2 production was proven to induce plant cell death, which simultaneously is a defense response to abiotic stress (Tisi et al. 2011).
Detoxification of ROS is another important PA function. A study on tobacco and tomato plants overexpressing a synthesis enzyme, arginine decarboxylase, showed that transgenic plants have a higher drought stress tolerance and drastically reduced ROS levels (Wang et al. 2011). It was also confirmed that low levels of PAs in soy plants, especially of Put and Spd, are linked to an increased damage from stress and decreased water content (Nayyar and Chander 2004). In addition, PAs exert a positive influence on the activity of enzymes of the Halliwell–Asada pathway that controls non-enzymatic antioxidant content such as ascorbate and glutathione (Kubiś 2001).
In numerous studies, PA inhibitors were used to modify the cellular PA content to evaluate their role in various plant processes (Kaur-Sawhney et al. 2003; Takahashi and Tong 2015). However, little known about the scope of inhibitor activity, their transport and metabolism (Sobieszczuk-Nowicka and Legocka 2007). However, the results of in vivo experiments using this chemical should be estimated carefully because of their unexpected side effects (Smith 1990). Treatment with polyamine synthesis inhibitors reduces the plant’s tolerance to stress, but the simultaneous application of exogenous PAs and inhibitors can retain their tolerance (Lee 1997; He et al. 2002). Flores (1991) showed that the use of arginine and ornithine decarboxylase inhibitors before osmotic stress stimulates the accumulation of PAs. Furthermore, the use of an arginine inhibitor reduces the amount of putrescine in stressed plants (Aziz et al. 1998). Smith et al. (1985) showed that long-term treatment with DFMA restrains pea plant growth. However, Yamamoto et al. (2016) did not observe significant differences in rice plants growth with or without PA inhibitor.
The aim of the study was to examine changes in the content of polyamines under stress conditions and dl-α-difluoromethylarginine (DFMA) treatment in yellow lupin plants subjected to soil drought stress, differing in drought tolerance, and to determine how these compounds affected the yield components.
Materials and methods
Plant material
Two cultivars of yellow lupin (Lupinus luteus L.): ‘Morocco 4’, the more tolerant to soil drought, and the more sensitive cv. ‘Taper’ were tested. Estimation of tolerance to soil drought stress of the tested cultivars was performed during the previous experiments (Juzoń et al. 2013).
The seed material was received from the Poznańska Plant Breeding Ltd., Tulce, Poland. Yellow lupin seeds were sown into a 3.6 kg mixture of horticultural soil (Ziemia uniwersalna, Ekoziem, Jurków, Poland) with sand (2:1 v/v) in pots (50 cm × 20 cm × 20 cm). The plants were grown under conditions attributable to the spring–summer period (April–September), in an open-air vegetation tunnel protected from rain by gardening foil (Agrimpex Ltd., Jarosław, Poland).
When plants developed five to six leaves, watering of half of them (30 plants) was restricted for 2 weeks and the seedlings were subjected to soil drought at 25% field water capacity (FWC) for a period of 2 weeks. The rest of the plants (30 plants) were watered to 70% FWC (control). Plants after the drought period were watered to 70% FWC to allow further analysis of the yield components as well as evaluate the impact of re-watering on the physiological state of tested plants (data not shown). Water content in the soil was controlled gravimetrically, including the weight of plants growing in the pots, and by using a moisture meter MO750 (Extech Instruments Corporation, USA).
On the first day of drought, 15 plants subjected to soil drought and 15 well-watered plants (control) were fully sprayed with an aqueous solution of polyamine biosynthesis inhibitor, dl-α-difluoromethylarginine (DFMA) (2.5 mL/plant), at a concentration of 0.1 mM.
Relative water content in leaves
Relative water content (RWC) was determined in leaves on the 1st and 14th day of drought according to Eric et al. (2005) using the following formula: RWC (%) = (Wf − Wd)/(Wt − Wd) × 100, where Wf, Wd and Wt represent fresh weight, dry weight and turgid weight, respectively. Samples were collected from second, fully developed leaf from each independent plant in 15 replications.
Polyamine (PA) content
Polyamine analyses in the leaves and seeds of yellow lupin were made according to Marcińska et al. (2013). The measurements were performed on the plants’ leaves on the 1st and 14th day of drought. Three to four fully formed leaves (counting from the top) were used for the analyses. In seeds, PAs were analyzed after plants’ maturation. The samples were lyophilized and then homogenized in a ball mill (MM 400, Retsch, Kroll, Germany). 0.02 g of the lyophilized leaves and seeds was extracted in 1 mL of 5% chloric acid (VII) and sonicated for 10 min. Then the samples were centrifuged at 37,000×g for 10 min and the supernatant was collected. The extraction was repeated and the supernatants were combined and mixed. The portion (200 µL) of combined supernatants were neutralized with 10 µL saturated NaOH. After that 400 µL dansyl chloride solution (5 mg/mL in acetone) and 200 µL saturated sodium carbonate solution were added. The samples were incubated at room temperature overnight. Subsequently, proline solution (100 mg/mL in water) was added and the mixture was incubated for 30 min. Finally, dansylated PAs were extracted in 750 µL toluene. The extraction was made twice and the upper toluene layers were collected, combined and evaporated under nitrogen (TurboVap® LV, Caliper Life Sciences, USA). The dry residue was dissolved in methanol, filtered through 0.22 μm membrane and analyzed by high-performance liquid chromatography (HPLC). The HPLC system (Agilent Technologies 1100 series, USA) was equipped with a fluorescence detector and autosampler, Zorbax Eclipse XDB-C18 4.6 × 75 mm 3.5 μm column (Agilent Technologies, Santa Clara, CA, USA). The mobile phase was methanol and water under a linear gradient of 60–95% methanol from 1 to 10 min. Fluorescence detection was conducted at 365 nm excitation wavelength and 510 nm emission wavelength. The content of PAs was determined in milligram per 1 gram DW.
Yield component analyses
At the stage of plant maturity, the number of seeds per plant, number of seeds per pod, dry weight of seeds per plant, weight of 1000 seeds and the dry weight of shoot were determined.
Statistical analysis
A completely randomized design was used to perform the experiment. There were 12 pots with 60 plants in total. A replication for each treatment (well-watered plants with and without DFMA or plants subjected to drought with and without DFMA) consisted of three pots with five plants. The results presented in figures constitute the mean values ± standard error based on 15 plant as replicates. Data were analyzed using two-way ANOVA using a statistical software package STATISTICA 10.0 (Stat-Soft, USA). Duncan’s multiple range test at the 0.05 probability level was used to determine the significance of differences between cultivars, marked as different letters. Student’s t test at the 0.05 significance level was also used to compare the means for each treatment.
Results
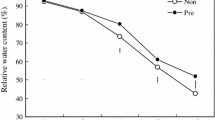
Relative water content in lupin leaves under soil drought
Relative water content significantly differed between treatments, within each genotype, which was confirmed by Duncan’s multiple range test (Fig. 1). On the 1st day of drought, cv. ‘Morocco 4’ showed a decrease of 4% of relative water content (RWC) compared to control plants. On the 14th day of drought, both cultivars showed a reduction of RWC, the largest after inhibitor treatment in ‘Morocco 4’ leaves (5%). However in cv. ‘Taper’ comparable decrease of RWC values was determined regardless of the absence or presence of DFMA (4 and 3%, respectively).

Effect of soil drought on the relative water content (RWC) (%) in the leaves of two cultivars of yellow lupin (‘Morocco 4’, ‘Taper’) on day 1 and 14 of drought; C control, 70% FWC, D drought, 25% FWC; the mean values, n = 15. Significant differences according to Duncan’s test at the 0.05 level of probability are marked with different letters
White-colored bars represent well-watered (control) plants without and with application of DFMA. Inhibitor of polyamine biosynthesis was applied exactly on the 1st day of drought. Since it is difficult to expect measurable effects of its actions on the day of application, its impact on the tested parameters was considered only after 14 days of drought.
Polyamine (PAs) content in lupin leaves and seeds under soil drought
Qualitative and quantitative analyses of PA contents were conducted on the basis of retention times (Rt) of PA standards (Sigma-Aldrich, Germany) and their peak areas (Fig. 2). In the analyzed plant material, seven PAs were identified: agmatine (Agm; Rt = 7.625 min), 1.3-diaminopentane (1.3-dPen; Rt = 8.852 min), putrescine (Put; Rt = 9.123 min), cadaverine (Kad; Rt = 9.536 min), 1.6-diaminohexane (1.6-dHex; Rt = 10.076 min), spermidine (Spd; Rt = 12.117 min) and spermine (Spm; Rt = 13.887 min). Among the measured PAs, Put, Spd and Spm were present at the highest concentrations. An increase in the total PA content was found in yellow lupin leaves under drought conditions (Table 1). PA concentrations on the 1st day of drought was 433.7 µg g−1 DW; thus, there was an increase of 7% compared to control conditions. Although yellow lupin leaves contained less PAs on day 14 of drought than on day 1, an increase of 19% compared to control plants was also observed. On the 1st day of drought, lupin leaves showed the highest amount of Put (170.9 µg g−1 DW) and Spd (184.8 µg g−1 DW), which accounted for 82% of the total PA content. On that day, an increase in Put and Spd contents (67 and 11%, respectively) was also reported. However, on the 14th day of drought, Put content did not change significantly, while Spd increased by 17%. Spm content did not show statistically significant changes both on day 1 and 14, and its content in the leaves ranged from 20.5 to 34.8 µg g−1 DW. Agm was not detected in the leaves of the tested species.

Chromatograms of polyamine pure standard mixture (a) and endogenous polyamines in yellow lupin leaves cv. Morocco 4 at 14 days of drought without DFMA (b)
The composition of PAs in the leaves was not the same as in the seeds of yellow lupin. The presence of four PAs: Agm, Put, Spd and Spm was confirmed in the seeds (Table 1). Most of all, the seeds contained 1.5-fold less PAs than the leaves under control conditions. Yellow lupin seeds obtained from plants growing under control conditions contained the highest amounts of Spm (92.3 µg g−1 DW), nearly fourfold more than in the leaves. The amount of Agm and Spd was comparable (50.7 and 43.3 µg g−1 DW, respectively), while Put was present in the smallest quantity (5.83 µg g−1 DW). Moreover, soil drought stress decreased the PA contents in the seeds, as opposed to an increase observed in the leaves. The total PA contents in the seeds declined by 54%. The highest reduction was observed in Agm and Spm (64 and 65%, respectively), while Spd showed a 27% decrease. The only exception was Put that did not show significant changes.
The influence of DFMA on the PA content and yield components of lupin
The results of DFMA polyamine biosynthesis inhibitor application are presented in Fig. 3. The two tested cultivars of yellow lupin demonstrated different reactions to soil drought and the presence of DFMA. On the 1st day, the total PA contents did not change significantly in the tolerant cv. ‘Morocco 4’, while an increase by 35% was detected in the sensitive cv. ‘Taper’. Furthermore, cv. ‘Morocco 4’ had a fivefold higher concentration of 1.3-dPen than cv. ‘Taper’ under drought conditions. On the other hand, cv. ‘Taper’ contained Put under drought, while this polyamine was not found in cv. ‘Morocco 4’. Both cultivars contained similar concentrations of 1.6-dHex, ranging from 89.45 to 138.50 µg g−1 DW, but an increase by 27% was observed in cv. ‘Taper’. Although this cultivar contained 28% less of Spd than cv. ‘Morocco 4’, it showed a 43% increase compared to control conditions. Spm concentration in both cultivars did not exhibit any significant changes.

Effect of soil drought on the total content of polyamines (μg g−1 DW) in the leaves of two cultivars of yellow lupin (‘Morocco 4’, ‘Taper’) on day 1 and 14 of drought; C control, 70% FWC, D drought, 25% FWC; the mean values, n = 9; nd not detected. Significant differences according to Duncan’s test at the 0.05 level of probability are marked with different letters
The period of 14 days of soil drought did not affect the total content of PAs in yellow lupin leaves. Cv. ‘Morocco 4’ contained 1.3-dPen under drought, while its presence in cv. ‘Taper’ was not confirmed. There was an increase in Put content in both cultivars: ‘Morocco 4’—25%, ‘Taper’—52%. Cultivar ‘Morocco 4’ was also characterized by a 42% increase of 1.6-dHex, and a 12% decrease of Spd. Cultivar ‘Taper’ did not show significant modification in the contents of these PAs. As regards the Spm content, a decrease was recorded in both cultivars tested (‘Morocco 4’—20%, ‘Taper’—40%).
The application of DFMA on day 14 of drought did not induce significant changes in the total PA contents in yellow lupin plants. However, a twofold increase in 1.3-dPen and a decrease in Spm content in cv. ‘Morocco 4’ was found (50 and 41%, respectively). Other PAs tested were not influenced by DFMA, while Put was not detected at all. Cultivar ‘Taper’ showed a decrease in the Spd and Spm content (13 and 19%, respectively). DFMA did not affect the synthesis of Put and 1.6-dHex, and 1.3-dPen was not found.
When the inhibitor was not applied, soil drought did not cause significant changes in yield components (Table 2). The use of DFMA under drought conditions led to a reduction in the number of pods per plant, number of seeds per plant and dry weight of seeds in cv. ‘Morocco 4’ (64, 50 and 54%, respectively). ‘Taper’ was characterized by a decrease in the number of pods per plant and number of seeds per pod (32 and 27%, respectively), which was nearly twofold smaller. The only exception was the weight of 1000 seeds which was increased by 21% in cv. ‘Morocco 4’.
Discussion
Approximately, one-third of the world’s cultivated land is affected by the problem of drought stress (Massacci et al. 2008). Drought as one of the major abiotic stresses limits crop yields all over the world (Sharp et al. 2004). Despite many studies on morphological, physiological, biochemical and molecular mechanisms of adaptation, plant tolerance to drought stress is not well understood, and obtaining drought-tolerant cultivars is a fairly slow process (Cabuslay et al. 2002).
In our study, we applied drought stress at the initial growth phase of lupin to check how it affects the yield of the tested cultivars. The cessation of watering was introduced when the plants had five to six leaves. We established two measurements days, 1 and 14, which meant, respectively, the beginning and the end of drought treatment. Rosales et al. (2012) studying drought stress in common bean measured RWC for the first time 29 days after sowing and the final measurement was done on the last day of drought treatment. In their studies, it was noticed that leaf RWC generally decreased with time in both well-watered and drought treatments. However, under drought, RWC of many genotypes were significantly lower compared to the control condition, and the differences became larger with time. Also, Subbarao et al. (2000) in pigeonpea measured RWC not on the first day of drought, but 10 days after beginning treatment. In our experiment, control plants throughout the experiment were watered to 70% FWC. A statistically significant decrease in RWC of control plants at 14 day compared to 1 day could be an effect of additional environmental factors, and a period of 13 days between these two measurements seems to be sufficient to observe permanent changes in RWC (Fig. 1). According to the scheme of the experiment, 14 days of drought occurred in May when higher temperature (reaching ca. 15 °C), compared to 1 day, could lead to the observed decrease. Low air saturation and accompanying wind (plants grown in an open-air vegetation tunnel) could be probably responsible for these changes. The important point is that the differences between well-watered and plants subjected to drought at 14 day were observed.
PAs are marked with great interest, due to a wide spectrum of their biological effects. Their action can vary substantially, depending on the plant species (Zapata et al. 2004). In many cases, stress is associated with an increase in the free PA content, demonstrating that its metabolism is a major component of stress tolerance mechanisms. Generally, it is observed that tolerant genotypes accumulate greater amounts of PAs than the sensitive ones (Hatmi et al. 2015). The content of free PAs depends on their biosynthesis as well as on their transport, conjugation and degradation. In our study, the total PA contents also varied in cultivars differing in tolerance to soil drought stress. The more tolerant cv. ‘Morocco 4’ had a higher amount of PAs compared with the less tolerant cv. ‘Taper’. Put, Spd and Spm are among the most prevalent PAs in plants (Marcińska et al. 2013), which was also confirmed in the present study where these three compounds constituted over 90% of the total PA contents in yellow lupin leaves and seeds. According to Hura et al. (2015), PA contents may differ between and within species and rely on plant organ and developmental stage. In our study, the absence of Put on the 1st and 14th day of drought could be explained by the fact that this polyamine is used for further synthesis of Spd and Spm, of which large amounts were recorded on these days. Thus, Put was mostly used in the Spd and Spm synthesis and the remaining amount was not sufficient to be detected. It was observed that yellow lupin plants were characterized by the highest amount of Spd in leaves when compared with Put and Spm; thus, it is presumed that this triamine can be involved in the tolerance of yellow lupin to soil drought. Furthermore, Spd seems to be more stable under drought conditions and in the presence of DFMA. Therefore, its greater involvement in the plant response to applied stress is supposed. Moreover, the tolerant cv. ‘Morocco 4’ showed nearly a twofold higher content of Spd in leaves compared to the sensitive cv. ‘Taper’. However, González de Mejía et al. (2003) showed that the concentration of Spm was higher than that of the other PAs in bean genotypes, which suggested that Spm, as opposed to Put, was not used in the biosynthesis of other PAs.
In our investigations, changes noticed in the content of PAs in the leaves and seeds of yellow lupin plants showed opposite trends. While there was an increase in the PA content in the leaves, a decrease was observed in the seeds. Furthermore, we detected the presence of Agm in yellow lupin seeds, which was not found in leaves. Spm was a polyamine that occurred in yellow lupin seeds in the largest quantities and was drastically decreased under soil drought conditions. González de Mejía et al. (2003) in tepary bean (Phaseolus acutifolius) seeds obtained similar results, where Spm concentrations were the highest compared to that of the others polyamines. Their results imply that Spm is not used, as Put is, in the biosynthesis of other polyamines. Endogenous PA levels in plants show dynamic changes under abiotic stress treatments. In the tested leaves of yellow lupin, Put was elevated on day 1 and did not show substantial changes on day 14, while Spd was decreased on day 1 and increased on day 14 of drought. Similarly, in Arabidopsis, Put increased after drought stress treatment for 12 days, while Spd declined after 7, 10 and 12 days of drought stress treatment (Alcázar et al. 2010). To date, several chemicals have been recognized as inhibitors of polyamine biosynthesis (Kaur-Sawhney et al. 2003), including dl-α-difluoromethylarginine (DFMA) and difluoromethylornithine (DFMO). The experiments conducted using exogenous PA application and/or inhibitors of enzymes involved in PA biosynthesis indicated a possible role of these compounds in plant adaptation to several environmental stresses (Alcázar et al. 2006). Despite the fact that treatments with PA biosynthesis inhibitors diminished stress tolerance, this effect was reversed by simultaneous application of exogenous PAs. However, these inhibitors are heterogeneous and possess unspecific roles in PA biosynthesis (Shi and Chan 2014). The current study showed that the DFMA treatment on the 14th day of drought decreased the total PAs and Spm content in ‘Morocco 4’ cultivar as well as Spd and Spm in ‘Taper’ cultivar, which made it difficult to clearly identify the mode of DFMA action in yellow lupin plants. Alcázar et al. (2006) reported that the stability and specificity of inhibitors were questionable.
Most studies focused on PA metabolism under stress conditions, excluding their impact on yield. However, according to Alcázar et al. (2006), manipulation of polyamine biosynthesis by using inhibitors of their biosynthesis may improve plant tolerance against multiple environmental stresses. In accordance with our research, it seems that these compounds may exert an indirect impact on yield components of yellow lupin plants (Table 2). Tolerant cultivar ‘Morocco 4’ under soil drought condition and influence of PA inhibitor showed a decline in the Spm content accompanied also by a decrease of some yield components. Under soil drought and the influence of DFMA, ‘Morocco 4’ produced less pods and seeds per plant and dry weight of these seeds was also reduced. Probably, this cultivar adapted to more severe hydration conditions according to its origin, and in more favorable conditions can function better even in the presence of DFMA. Moreover, it is presumed that the concentration of applied inhibitor was not high enough for Morocco to bring the expected effect. Sensitive cultivar ‘Taper’, under the same conditions (drought and DFMA treatments), was characterized by a decrease in pod number per plant and seed number per pod. When no inhibitor was used, both cultivars did not show a significant reduction in the values of yield components. These results are contrary to our earlier research, where most of the tested yield components decreased under drought conditions (Juzoń et al. 2013). It is assumed that this unexpected situation may be due to the weather condition, because the year of the conducted experiment was characterized by high humidity and plants could not be affected by stress efficiently enough.
Until now, the physiological role of PAs in yellow lupin tolerance to drought stress is still unclear. Therefore, it seems necessary to conduct further research that may help in understanding the mechanisms that could be used in breeding and farming practice in the future.
Author contribution statement
KJ and ES designed and carried out the experiment, analyzed the data and wrote the manuscript; IC-M and IM carried out RWC analysis; MD and PW carried out HPLC analysis. All these authors have read and approved the final manuscript.
References
Alcázar R, Marco F, Cuevas JC, Patron M, Ferrado A, Carrasco Tiburcio AF, Altabella T (2006) Involvement of polyamines in plant response to abiotic stress. Biotechnol Lett 28:1867–1876
Alcázar R, Planas J, Saxena T, Zarza X, Bortolotti C, Cuevas J, Bitrián M, Tiburcio AF, Altabella T (2010) Putrescine accumulation confers drought tolerance in transgenic Arabidopsis plants over-expressing the homologous Arginine decarboxylase 2 gene. Plant Physiol Biochem 48:547–552
Alet AI, Sánchez DH, Cuevas JC, Marina M, Carrasco P, Altabella T, Tiburcio AF, Ruiz OA (2012) New insights into the role of spermine in Arabidopsis thaliana under long-term salt stress. Plant Sci 182:94–100
Aziz A, Martin-Tanguy J, Larher F (1998) Stress-induced changes in polyamine and tyramine levels can regulate proline accumulation in tomato leaf discs treated with sodium chloride. Physiol Plant 104:195–202
Cabuslay GS, Ito O, Aleja AA (2002) Physiological evaluation of response of rice (Oryza sativa L.) to water deficit. Plant Sci 163:815–827
Eric SO, Bloa ML, Clark CJA, Royal A, Jaggard KW, Pidgeon JD (2005) Evaluation of physiological traits as indirect selection for drought tolerance in sugar beet. Field Crops Res 91:231–249
Flores HE (1991) Changes in polyamine metabolism in response to abiotic stress. In: Slocum RD, Flores HE (eds) Biochemistry and physiology of polyamines in plants. CRC Press, Boca Raton, pp 213–225
Gemperlová L, Nováková M, Vaňková R, Eder J, Cvikrová M (2006) Diurnal changes in polyamine content, arginine and ornithine decarboxylase, and diamine oxidase in tobacco leaves. J Exp Bot 57:1413–1421
González de Mejía E, Martínez-Resendiz V, Castanõ-Tostado E, Loarca-Pin G (2003) Effect of drought on polyamine metabolism, yield, protein content and in vitro protein digestibility in tepary (Phaseolus acutifolius) and common (Phaseolus vulgaris) bean seeds. J Sci Food Agric 83:1022–1030
Graham PH, Vance CP (2003) Legumes: importance and constraints to greater use. Plant Physiol 131:872–877
Hatmi S, Gruau C, Trotel-Aziz P, Villaume S, Rabenoelina F, Baillieul F et al (2015) Drought stress tolerance in grapevine involves activation of polyamine oxidation contributing to improve immune response and low susceptibility to Botrytis cinerea. J Exp Bot 66:775–787
He LX, Nada K, Kasukabe Y, Tachibana S (2002) Enhanced susceptibility of photosynthesis to low-temperature photoinhibition due to interruption of chill-induced increase of S-adenosylmethionine decarboxylase activity in leaves of spinach (Spinacia oleracea L.). Plant Cell Physiol 43:196–206
Hura T, Dziurka M, Hura K, Ostrowska A, Dziurka K (2015) Free and cell wall-bound polyamines under long-term water stress applied at different growth stages of × Triticosecale Wittm. PLoS One 10(8):e0135002. doi:10.1371/journal.pone.0135002
Hussain SS, Ali M, Ahmad M, Siddique KHM (2011) Polyamines: natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol Adv 29:300–311
Juzoń K, Skrzypek E, Czyczyło-Mysza I, Marcińska I (2013) Effect of soil drought on the yield structure, protein and phenolics content in Pisum sativum and Lupinus luteus. Acta Agron Hung 61:267–278
Kasukabe Y, He L, Nada K, Misawa S, Ihara I, Tachibana S (2004) Overexpression of spermidine synthase enhances tolerance to multiple environmental stress and up-regulates the expression of various stress-regulated genes in transgenic Arabidopsis thaliana. Plant Cell Physiol 45:712–722
Kaur-Sawhney R, Tiburcio AF, Altabella T, Galston A (2003) Polyamines in plants: an overview. J Cell Mol Biol 2:1–12
Kubiś J (2001) Polyamines and ‘scavenging system’: influence of exogenous spermidine on Halliwell-Asada pathway enzyme activity in barley leaves under water deficit. Acta Physiol Plant 23:335–341
Lee TM (1997) Polyamine regulation of growth and chilling tolerance of rice (Oryza sativa L.) roots cultured in vitro. Plant Sci 122:111–117
Lester GE (2000) Polyamines and their cellular anti-senescence properties in honey dew muskmelon fruit. Plant Sci 160:105–112
Liu JH, Nada K, Honda C, Kitashiba H, Wen XP, Pang XM, Moriguchi T (2006) Polyamine biosynthesis of apple callus under salt stress: importance of arginine decarboxylase pathway in stress response. J Exp Bot 57:2589–2599
Marcińska I, Czyczyło-Mysza I, Skrzypek E, Grzesiak MT, Janowiak F, Filek M, Dziurka M, Dziurka K, Waligórski P, Juzoń K, Cyganek K, Grzesiak S (2013) Alleviation of osmotic stress effects by exogenous application of salicylic or abscisic acid on wheat seedlings. Int J Mol Sci 14:13171–13193
Massacci A, Nabiev SM, Pietrosanti L, Nematov SK, Chernikova TN, Thor K, Leipner J (2008) Response of the photosynthetic apparatus of cotton (Gossypium hirsutum) to the onset of drought stress under field conditions studied by gas-exchange analysis and chlorophyll fluorescence imaging. Plant Physiol Biochem 46:189–195
Moschou PN, Wu J, Tavladoraki P, Angelini R, Roubelakis-Angelakis KA (2012) The polyamines and their catabolic products are significant players in the turnover of nitrogenous molecules in plants. J Exp Bot 63:5003–5015
Nayyar H, Chander S (2004) Protective effects of polyamines against oxidative stress induced by water and cold stress in chickpea. J Agron Crop Sci 190:355–365
Postel SL (2000) Entering an era of water scarcity. Ecol Appl 10:941–948
Rosales MA, Ocampo E, Rodríguez-Valentín R, Olvera-Carrillo Y, Acosta-Gallegos J, Covarrubias AA (2012) Physiological analysis of common bean (Phaseolus vulgaris L.) cultivars uncovers characteristics related to terminal drought resistance. Plant Physiol Biochem 56:24–34
Sharp RE, Porokyo V, Hejlek LG, Spollen WG, Springer GK, Bohnert HJ, Nguyen HT (2004) Root growth maintenance during water deficits: physiology to functional genomics. J Exp Bot 55:2343–2351
Shi H, Chan Z (2014) Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J Integr Plant Biol 56:11–121
Smith TA (1990) Plant polyamines-metabolism and function. In: Flores HE, Arteca RN, Shannon JE (eds) Polyamines and ethylene: biochemistry, physiology, and interactions. Am Soc Plant Physiol, pp 1–23
Smith MA, Davies PJ, Reid JB (1985) Role of polyamines in gibberellin-induced internode growth in peas. Plant Physiol 78:92–99
Sobieszczuk-Nowicka E, Legocka J (2007) Nowe podejścia w badaniach nad rolą poliamin w komórce roślinnej. Post Biol Kom 34:527–540 (in polish)
Subbarao GV, Chauhan YS, Johansen C (2000) Patterns of osmotic adjustment in pigeonpea—its importance as a mechanism of drought resistance. Eur J Agron 12:239–249
Takahashi T, Tong W (2015) Regulation and diversity of polyamine biosynthesis in plants. In: Kusano T, Suzuki H (eds) Polyamines. A universal molecular nexus for growth, survival, and specialized metabolism. Springer, Tokyo, pp 27–44
Tavladoraki P, Cona A, Federico R, Tempera G, Viceconte N, Saccoccio S, Battaglia V, Toninello A, Agostinelli E (2012) Polyamine catabolism: target for antiproliferative therapies in animals and stress tolerance strategies in plants. Amino Acids 42:411–426
Tisi A, Federico R, Moreno S, Lucretti S, Moschou PN, Roubelakis-Angelini R, Cona A (2011) Perturbation of polyamine catabolism can strongly affect root development and xylem differentiation. Plant Physiol 157:200–215
Wang BQ, Zhang QF, Liu JH, Li GH (2011) Overexpression of PtADC confers enhanced dehydration and drought tolerance in transgenic tobacco and tomato: effect on ROS elimination. Biochem Biophys Res Commun 413:10–16
Yamamoto A, Shim IS, Fujihara S (2016) Inhibition of putrescine biosynthesis enhanced salt stress sensitivity and decreased spermidine content in rice seedlings. Biol Plant. doi:10.1007/s10535-016-0676-5
Yang J, Zhang J, Liu K, Wang Z, Liu L (2007) Involvement of polyamines in the drought resistance of rice. J Exp Bot 58:1545–1555
Zapata PJ, Serrano M, Pretel MT, Amorós A, Botella MÁ (2004) Polyamines and ethylene changes during germination of different plant species under salinity. Plant Sci 167:781–788
Acknowledgements
This work was funded by the National Science Centre, Poland, Project no. 621/N-COST/09/2010/0.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by K Apostol.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Juzoń, K., Czyczyło-Mysza, I., Marcińska, I. et al. Polyamines in yellow lupin (Lupinus luteus L.) tolerance to soil drought. Acta Physiol Plant 39, 202 (2017). https://doi.org/10.1007/s11738-017-2500-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-017-2500-z