
Abstract
We assessed the effects of arbuscular mycorrhizal fungi (AMF) Rhizophagus irregularis inoculation on salt stress tolerance in roots of the drought-tolerant plant Elaeagnus angustifolia. We studied a plant growth index, spore density and hyphal length density of AMF, the Na+ contents and ultrastructure of root cells, as well as rhizosphere soil enzyme activities of mycorrhizal and non-mycorrhizal E. angustifolia seedlings under different salt stress. Under salt stress, growth of E. angustifolia with mycorrhizal inoculation was higher than that of non-inoculated treatments. The spore density and hyphal length density decreased significantly under salt stress in rhizosphere soil of mycorrhizal E. angustifolia seedlings (p < 0.05). The root cells of E. angustifolia seedlings inoculated with R. irregularis at 300 mmol L−1 salt had more organelles, greater integrity, and lower root Na+ contents than those of non-inoculated seedlings. In addition, the results showed notably higher activities of catalase, phosphatase, urease and saccharase in rhizosphere soil of the mycorrhizal seedlings in response to salinity compared to those of the non-mycorrhizal seedlings. Therefore, AMF inoculation could enhance salt stress tolerance in roots of E. angustifolia.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Salinity is one of the main abiotic factors that deleteriously affect plant development and yield (Elhindi et al. 2017; Metwally and Abdelhameed 2018). High salt concentration in the soil can limit plant biochemical processes, mainly because osmosis leads to water deficits and specific ionic effects that can lead to toxicity and nutrient imbalance (Yin et al. 2019; Heuer 2005; Zhu and Gong 2014). In addition, salt stress affects the ability of plant roots to absorb water by reducing soil water potential, which increases the likelihood of cell dehydration (Abdel-Fattah et al. 2016; Ebrahim and Saleem 2017; Metwally and Abdelhameed 2018).
Arbuscular mycorrhizal fungi (AMF) can establish mycorrhizal symbioses with most terrestrial plant species (Chang et al. 2018; Xu et al. 2016), which can lead to changes in the morphological, nutritional and physiological state of plants. Moreover, these effects of AMF symbioses can enhance plant resistance against different abiotic stresses, such as salt, drought, and cold (Alqarawi et al. 2014a, b; Hashem et al. 2015). Shekoofeh et al. (2012) proved that inoculation with AMF protects Ocimum basilicum against salt stress by improving mineral uptake, chlorophyll synthesis and water use efficiency. The enhanced growth of AMF-treated tomato plants was associated with mycorrhizal-associated host plant nutrient uptake (Hashem et al. 2015). Combining AMF and rhizobium inoculations can also enhance plant growth, nodulation, and nitrogen fixation of soybean under salt stress (Elsheikh and Wood 1995; Younesi et al. 2013). Meanwhile, the number of mycelia and vesicles, plus arbuscule formation and colonization were reduced significantly due to salt stress (Hashem et al. 2018, 2019).
AMF can promote the growth of plants in normal and stressed environments by improving soil enzyme activities (Gamalero et al. 2010; Navarro et al. 2014; Alqarawi et al. 2014a, b). Soil enzymes are derived from soil microorganisms and plant roots and are involved in many biological processes (Marx et al. 2001; Shao et al. 2019), and increasing salinity levels may reduce soil enzyme activity (Usman 2015; Zheng et al. 2017). Raiesi and Sadeghi (2019) found that in cadmium-contaminated ecosystems, high-salinity conditions have a greater negative impact on soil enzyme activity than non-saline conditions. In Zhang et al. (2014) study, the inoculation of AMF and A. spartina enhanced the activities of soil enzymes (i.e., urease, invertase, neutral and alkaline phosphatase) under salt stress.
Our research group has been studying the response of arbuscular mycorrhizal seedlings to salt stress for many years. Elaeagnus angustifolia belongs to the family Elaeagnaceae, and is the main tree species used for salt-alkaline land restoration in China because of its well-developed root system (Asgarzadeh et al. 2014; Zhang et al. 2018). Some achievements have been made in the studies of mycorrhizal resistance enzymes, photosynthetic physiology (Sun et al. 2016; Jia et al. 2018), ion distribution (Chang et al. 2018) and proteomics (Song et al. 2015a, b) of E. angustifolia seedlings, and we continue using this species here to study mycorrhizal effects on seedlings. The purpose of this study was to explore the effects of salt stress on mycorrhizal E. angustifolia seedlings and whether inoculation with AMF could alleviate the toxicity of salt stress on plants. To do so, we measured the plant growth index, the spore density and hyphal length density, the content of Na+ in root system, the ultrastructure of root cells and soil enzyme activity after treatment with different salt concentrations.
Materials and methods
Experimental design
The experiment used a randomized complete block design with two factors (non-mycorrhizal, abbreviated NM): mycorrhizal, which consisted of inoculation with Rhizophagus irregularis (previously known as Glomus intraradices, abbreviated RI), and four levels of the salinity factor: 0, 100, 200, and 300 mmol L−1 NaCl. Each of the eight treatments (NM0, NM100, NM200, NM300 and RI0, RI100, RI200, RI300) had ten replicates (Huaran et al. 2018; Chang et al. 2018). In order to avoid potential bias of pot locations, the positions of the pots were changed randomly every week.
Plant materials and soil
The seeds of E. angustifolia were provided by Heilongjiang Jinxiu Dadi Biological Engineering Co., Ltd., P. R. China, and were surface-sterilized with 0.2% KMnO4 for 10 min, washed for four times with sterile water, and grown in plastic pots. Every pot contained 5 L of substrate that was previously sterilized in an autoclave for 1 h at 121 °C for three times on alternate days. The substrate was a mixture of soil, peat, and vermiculite (soil:peat:vermiculite = 6:2:2, V/V; Na+ content 0.24%, pH 7.8) mainly from the Forest Botanical Garden of Heilongjiang Province (China, 45° 42′ 40.09′′ N 126° 38′ 22.23′′ W), and was passed through a 5-mm sieve. The pH of the original substrate was 7.2 (Water:soil = 5:1 [v/w]), organic matter content was 1.2%, available nitrogen content was 123.4 mg kg−1, available phosphorus content was 12.6 mg kg−1, and available potassium content was 76.5 mg kg−1. The conductivity of the original soil was 0.5 ds m−1.
Inoculation treatments
The RI inocula consisted of soil, spores, mycelia, and infected root fragments. It was obtained from an open pot culture (Sorghum bicolor L.) of R. irregularis isolated from an area with severe salinity problems (Zhao Yue Shan National Wetland Park, Heilongjiang Province, P. R. China, 46° 5′ 37.98′′ N and 125° 57′ 28.43′′ W). At the time of sowing, the soil was inoculated with R. irregularis. Each pot of mycorrhiza was inoculated with 10-g R. irregularis and planted 2 cm below the surface of the soil. The NM group received the same number of autoclaved mycorrhiza inoculations.
Salt stress
In order to avoid the effects of drought, water was supplied three times a week during the entire period of plant growth. The plants were subjected to salt stress after about 100-days growth sufficient for symbiotic establishment. Four levels (0, 100, 200, and 300 mmol L−1 NaCl) of saline solution were added to the soil by from a 2-mmol L−1 stock saline solution based on the amount of substrate in the pots. In order to avoid osmotic shock, the content of NaCl in the soil was gradually increased on alternate days. The desired levels of NaCl were reached after 6 days. In order to avoid salt loss, a tray was attached under the plastic pots.
Sampling
After one month of salt stress treatment, we harvested E. angustifolia plants. A part of the growing plants was randomly selected for each treatment and their roots were washed three times with distilled water for the determination of mycorrhizal colonization and Na+ content. The other part was cleaned and put in 2.5% glutaraldehyde solution and stored at 4 °C for ultrastructural observation. At the same time, all the rhizosphere soils of E. angustifolia were collected and screened by air drying for the determination of spore density, mycelium quantity, and soil enzyme activity.
Experimental methods
The height and basal diameter of E. angustifolia were measured accurately with vernier calipers. Root area and root length of E. angustifolia plants in each group were measured with a root scanner (LC-4800, Shanghai Xintian International Trade Co., Ltd.) after being washed carefully under distilled water to ensure their integrity.
Mycorrhizal colonization was evaluated by using fibrous roots as a subsample. These were rinsed with 10% potassium hydroxide at 90 °C for 15 min, soaked with 2% hydrochloric acid for 5 min, and stained with acid fuchsin (Koske and Gemma 1989). AMF spores were separated from the soil by wet sieving and 50% sucrose centrifugation (Brundrett et al. 1996). We determined hyphal length density in rhizosphere soil of E. angustifolia by Trypan blue staining (Ren et al. 2015).
The Na+ kurtosis of E. angustifolia root was determined by scanning electron microscope (TM3030, HITACHI, Japan, abbreviated SEM) and energy dispersive spectrometer (Quantax70, HITACHI, Japan, abbreviated EDS). Fibrous roots were fixed with 2.5% glutaraldehyde at 4 °C refrigerator and trimmed with sharp blades until the trimmed sample roots were about 1 cm. Then the roots were firmly adhered to the sample table with silver conductive adhesive, vacuum dried for 24 h, and sprayed with carbon. Finally, these were placed in the SEM. The analysis voltage was 20 kV and the beam spot was 7.2. SEM and was used to determine the sites of mycorrhizae in roots; quantitative analysis of Na+ was carried out by EDS. The obtained Na+ kurtosis value was the mass percentage at the range sampled point; this was not the Na+ contents per gram of root obtained in the previous experiment.
The fibrous roots of E. angustifolia under four different treatments (NM0, NM300, RI0 and RI300) were cut to a length of 1–2 mm, immersed in 2.5% glutaradehyde and then fixed at 4 °C for 4 h to prevent air bubbles from entering. Then, the samples were washed three times in phosphate buffer solution (PBS, 0.1 mol L−1, pH 7.0), post-fixed with 2% osmium tetroxide and rinsed three times again in PBS. Later, the samples were dehydrated with acetone, impregnated, and embedded in resin. Ultrathin sections were extracted on a copper grid and then secondary stained with 2.5% (w/v) uranyl acetate followed by staining with lead citrate. Finally, the samples were observed and photographed by transmission electron microscopy (H-7650, HITACHI, Japan, marked as TEM).
The phenylbiphenyl phosphate colorimetric method, 3,5-dinitrosalicylic acid colorimetric method, sodium phenol-sodium hypochlorite colorimetric method, and potassium permanganate titration were used to determine the activities of phosphatase (PHO), saccharase (SAC), urease (URE), and catalase (CAT), respectively (Guan 1986; Akhtar et al. 2018).
Statistical analysis
Data were analyzed using SPSS version 16.0. One-way analysis of variance (ANOVA) was performed to examine the significance of salt concentration on spore density, hyphal contents and Na+ contents. Two-way analysis of variance (ANOVA) was performed to examine the significance of inoculated with R. irregularis, salt, and their interactions on plants growth and soil enzyme activities. Significant differences between treatments were confirmed using Tukey’s HSD test at p < 0.05. Correlation analysis between salt concentration and each of spore density hyphal length was carried out using a two-tailed Pearson test, with p < 0.05 as the level of significance. All data were plotted using Origin 8.0 software.
Results
The plant growth index of E. angustifolia
The growth of E. angustifolia changed when salt was added. The growth of E. angustifolia inoculated with R. irregularis increased significantly under salt stress compared with non-mycorrhizal treatments (Table 1). Plant height, basal diameter, root length, and root surface area of mycorrhizal E. angustifolia were all significantly higher than those of non-mycorrhizal E. angustifolia. The height of mycorrhizal E. angustifolia seedlings increased by 9.82%, 7.04%, and 9.10%, the basal diameter increased by 10.57%, 10.35%, and 20.80%, the root length increased by 19.27%, 18.03% and 18.96%, and the root surface area increased by 46.56%, 39.27%, and 35.53%, respectively, compared with non-mycorrhizal treatment under 100, 200 and 300 mmol L−1 salt concentration. On the whole, the growth of E. angustifolia decreased with increased salt concentration. Compared with the salt-free treatment, the height of E. angustifolia plants treated with NM300 and RI300 decreased by 13.03% and 12.02%, the basal diameter decreased by 29.38% and 26.30%, the root length decreased by 26.59% and 22.49%, and the root surface area decreased by 42.70% and 35.90%, respectively, when the salt concentration was 300 mmol L−1. Salt and R. irregularis had no significant interactive effects on basal diameter, root length, and root surface area (p > 0.05, Table 1); however, there was an interactive effect on plant height (p < 0.001, Table 1). The growth of plants was hindered by salt stress, but the growth of mycorrhizal E. angustifolia was slightly less affected than that of non-inoculated seedlings, suggesting that R. irregularis could alleviate the toxicity of salt stress to plants and promote its growth.
Colonization
Plant growth (especially plant height) in seedlings inoculated with R. irregularis was higher than non-inoculated seedings under salt stress (300 mmol L−1; Fig. 1a).

a Plant growth of two treatments under 300 mmol L−1 salt stress; b. photomicrographs of structural colonization of arbuscular mycorrhizal fungi in the roots of E. angustifolia
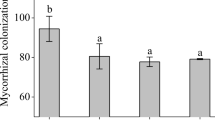
R. irregularis and E. angustifolia formed confirmed symbioses. Mycorrhizal structures associated with root colonization in E. angustifolia seedlings included internal hyphae, vesicles, and arbuscules. Mycorrhizal colonization of E. angustifolia seedlings was detected regularly in our samples using the method of Koske and Gemma (1989). A small amount of hyphae appeared in the roots of E. angustifolia seedlings approximately 2 months after inoculation, but colonization was low and no vesicles were found. Once seedlings grew for 3 months, mycorrhizal colonization of roots increased and vesicles appeared. When seedlings grew for 4 months, mycelia of R. irregularis in roots could be clearly observed under the microscope, a large number of vesicles were produced, and colonization rate reached more than 90%. No colonization was found in non-inoculated seedlings. We also confirmed mycorrhizal colonization of E. angustifolia after 1 month of salt stress and there were still over 90% colonized (Fig. 1b). Mycorrhizal colonization of E. angustifolia seedlings was not affected by salt stress.
Spore density and hyphal length density
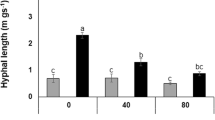
Spore density and hyphal length density of E. angustifolia rhizosphere soil were inhibited by salt stress (Fig. 2). With increasing salt concentration, the inhibitory effect was greater. The spore density in rhizosphere soil under the RI0 treatment was the highest, with an average of 27.67 ind. g−1 soil. The spore density of rhizosphere soil under RI300 treatment was the lowest, with an average content of ind. g−1 soil, and spore density increased by 33.87%, 88.63%, and 207.40% in the RI100, RI200 and RI300 treatments, respectively, compared to the RI0 treatment. The hyphal length density of soil was highest in the RI0 treatment, with an average concentration of 211.05 mm g−1. The hyphal length density of soil was lowest in the RI300 treatment, with an average concentration of 92.85 mm g−1. Hyphal length density increased by 15.54%, 34.16%, and 127.31% in the RI100, RI200 and RI300 treatments, respectively, compared to the RI0 treatment. There were significant differences in R. irregularis spore density and hyphal length density in rhizosphere soil depending on salt stress treatments (p < 0.05), and spore density (r = − 0.999, p < 0.05) and hyphal length density (r = − 0.973, p < 0.05) were both significantly negatively correlated with salt concentration.

R. irregularis spore density (a) and hyphal length density (b) in rhizosphere soil of E. angustifolia under different salt concentrations (n = 3). Error bars show the standard error. Columns with different letters indicate significant differences between treatments at p < 0.05
Root Na+ contents
The sodium ion kurtosis of the non- mycorrhizal treatments was higher than mycorrhizal treatments (Fig. 3)

Peak figure Na+ determination of E. angustifolia root spectrometer under the four treatments
The average Na+ peak value of E. angustifolia roots was 10.13% in NM0 treatment. The average Na+ peak value in RI0 treatment was 7.52%, which was 25.77% lower than that in NM0 treatment (p < 0.01). The average peak value of Na+ in NM300 treatment was 11.84%, while that in RI300 treatment was 8.04%. Compared with NM300 treatment, the average peak value of Na+ in roots decreased by 32.09% in RI300 treatment (p < 0.05; Fig. 4).

Na+ contents in fibrous roots of E. angustifolia under salt stress (300 mmol L−1). Error bars show the standard error. Columns with different letters indicate significant differences between treatments at p < 0.05
The ultrastructure of E. angustifolia root
TEM photographes of E. angustifolia root cells in NM0 treatment showed that the whole cell structure was relatively intact, including mitochondria, vacuoles and cell walls (Fig. 5). The root cells of E. angustifolia in RI0 treatment were closely arranged with complete cell structure. The complete cell walls, nuclei, nuclear membranes, nucleolus and uniformly distributed nuclear chromatin could be seen. A large number of regular round or oval mitochondria and vacuoles could be seen. The NM300-treated E. angustifolia root cells had incomplete cell structure, obvious plasmolysis, degraded organelles and serious cell damage. However, in RI300 treated root cells of E. angustifolia, the cells were still closely arranged, some organelles were degraded, the number of mitochondria was reduced, and vacuoles were still visible.

Root cell ultrastructure map of E. angustifolia. CW, cell wall; M, mitochondria; N, nucleus; Ne, nucleolus; NM, nuclear membrane; Ka, nuclear chromatin; IS, intercellular space; V, vacuoles
Rhizosphere soil enzyme activity
The activities of CAT, PHO, URE and SAC in rhizosphere soil of E. angustifolia in both treatments decreased gradually with increased salt concentration (Fig. 6). For a given salt concentration, the activities of CAT, PHO, and SAC in rhizosphere soil of E. angustifolia inoculated with R. irregularis were significantly higher than those in non-mycorrhizal treatments (p < 0.05). The activity of URE was not significantly different between NM0 and RI0 (p > 0.05), but was significantly different between mycorrhizal treatments under the other three salt concentrations (p < 0.05).

Enzyme activities in rhizosphere soil of E. angustifolia under different salt concentrations. NM, non-mycorrhizal; RI, Rhizophagus irregularis; a catalase; b phosphatase; c urease; d saccharase. Error bars show the standard error. Columns with different letters indicate significant differences between treatments at p < 0.05
At 100 mmol L−1 salt concentration, the activities of CAT, PHO, URE, and SAC in rhizosphere soil of E. angustifolia in the NM100 treatment decreased by 2.40%, 15.74%, 14.43%, and 29.01%, respectively, compared with NM0 treatment (Table 2). These differences between NM0 and NM100 treatments were significant, except for CAT (p < 0.05). Compared with RI0 treatment, the activities of CAT, PHO, URE, and SAC in the RI100 treatment decreased by 3.86%, 8.07%, 8.43%, and 28.26%, respectively, (p < 0.05 for all but URE). At 200 mmol L−1 salt concentration, the concentrations of the above four enzymes in the NM200 treatment decreased by 13.18%, 33.32%, 25.75%, and 36.71% compared with NM0 treatment, while those in RI200 treatment decreased by 9.21%, 19.48%, 16.24%, and 34.69%, respectively, compared with RI0 treatment. At 300 mmol L−1 salt concentration, the above four enzymes of NM300 treatment decreased by 31.50%, 38.18%, 49.96%, and 39.85%, respectively, compared with NM0 treatment, while those of RI300 treatment decreased by 14.80%, 40.68%, 37.21%, and 38.99%, respectively, compared with RI0 treatment. At 200 and 300 mmol L−1 salt concentration, the above indexes of non-mycorrhizal and mycorrhizal treatments were significantly different from those of non-salt treatments (p < 0.05). In addition, the interaction between R. irregularis and salt concentration on URE and SAC were not significant (p > 0.05, Table 2), but it was significant for CAT and PHO (p < 0.001, Table 2). The above results showed that salt stress can inhibit the activity of soil enzymes in E. angustifolia rhizosphere; the activity of soil enzymes decreased with increasing salt concentration. With increased salt concentration, the effect of mycorrhizal symbiosis between R. irregularis and E. angustifolia was clear; the mycorrhizae slowed the decrease of soil enzyme activity under salt stress.
Discussion
Effects of AMF on the growth of E. angustifolia under different salt concentrations
Symbiosis between AMF and host plants can not only promote the growth and development of plants (Hashem et al. 2019), but also enhance the resistance of host plants against different abiotic stresses (Smith and Read 2008; Hashem et al. 2018). AMF can reduce the inhibition of plant functions brought on by salt stress, and increase plant height, basal diameter, and growth, in general. AMF can increase the biomass of both above-ground and underground parts of plants under salt stress (Abd_Allah et al. 2015). The higher the salt concentration in the soil, the lower the biomass of plants (Babu et al. 2012), but inoculation of AMF can promote the growth of host plants under salt stress (Abd_Allah et al. 2015). Bheemareddy and Lakshman (2011) demonstrated that reduction in plant height, leaf area, and number of leaves per plants were observed and inoculation of AMF increased these attributes and also mitigated the salinity-induced deleterious effects. These conclusions are consistent with the results of our experiment.
In this experiment, we measured the growth of E. angustifolia height, basal diameter, root length, and root surface area under salt stress. The height and basal diameter of E. angustifolia seedlings with R. irregularis inoculation were significantly higher than those of non-mycorrhizal plants. Salt stress reduced the height and basal diameter of E. angustifolia plants, but for a given salt concentration, the mycorrhizal E. angustifolia grew better than non-mycorrhizal plants did. The total length and the root system of host plants increased following mycorrhizal treatment (Buerkert et al. 2000). The same conclusion has been obtained when measuring root length and root surface area in this experiment. However, under a given salt concentration, the roots of non-mycorrhizal seedlings were not easy to obtain completely, broke easily and were more fragile, while the roots of mycorrhizal E. angustifolia were relatively easy to measure. The analysis showed that the root system of E. angustifolia was more vulnerable to salt stress, and R. irregularis could enhance the salt tolerance of E. angustifolia roots and enhance the strength of root system.
Effects of salt stress on R. irregularis
Prior research shows that salt stress can reduce AMF spore density (Ouziad et al. 2006), slow down the growth of mycelia (Hammer and Rillig 2011), and inhibit the growth of mycelia and vesicles in roots, thus slowing down colonization of AMF (Navarro et al. 2014). But not much is known about the mechanism of interaction between salt stress and AMF. In this study, salt stress was induced in seedlings of E. angustifolia which had formed mycorrhizal symbioses. Even under high salt stress, the colonization of mycorrhizal of E. angustifolia seedlings was not affected. The colonization was still over 90% and the colonization intensity was high (Fig. 1b). The reason for the above results may be that the salt stress time is too short (one month), so it is necessary to continue to trace the mycorrhizal colonization rate of E. angustifolia under salt stress. In addition, E. angustifolia may have a better symbiotic relationship with R. irregularis under salt stress (Song et al. 2015a, b). However, Aroca et al. (2013) demonstrated that after 5 weeks, G. intraradices colonization decreased by 17% and 30% following 40 and 80 mmol L−1 NaCl-treatment of lettuce plants compared with non-treated plants (0 mmol L−1). Spore density and hyphal length density decreased significantly in rhizosphere soil with increased of salt concentration. Thus, salt stress inhibited R. irregularis spores and mycelia in soil, and the higher the salt concentration, the greater the inhibition (Kumar et al. 2014).
Effect of salt stress on Na+ contents of E. angustifolia roots in different treatments
Na+ in soil infiltrates into plant root cells with water, which decreases intracellular water potential. When Na+ enters the plant cells in large quantities, it can disrupt the ion balance in the cells, dehydrating root cells and slowing normal plant growth. We found that the Na+ contents in roots of E. angustifolia treated with R. irregularis was lower than that of non-mycorrhizal treatments, which indicated that the root Na+ contents of E. angustifolia was reduced by R. irregularis. Chang et al. (2018) suggested that inoculation with AMF promotes Na+, K+, Ca2+, and Mg2+ accumulation in the roots of E. angustifolia during treatment with salt. When the salt concentration was 300 mmol L−1, the Na+ kurtosis value of NM300 in the roots of E. angustifolia increased significantly. However, the trend of Na+ increase in RI300 roots was not obvious. The results showed that the root system of E. angustifolia had restrained the intake of Na+ in a certain range, thus slowing down the salt damage to the root system. The degree of damage by Na+ to plasma membranes and enzyme activity was reduced, and salt tolerance of E. angustifolia was enhanced. However, how mycorrhizal symbiosis reduce root uptake of Na+ remains to be further studied.
Effect of salt stress on root ultrastructure of E. angustifolia under different treatments
Roots are the most important nutrient absorption organs in the underground part of plants and are also the plant parts first to experience and most sensitive to salt stress. Salt stress can also cause damage to the structure of root cells (Liang et al. 2018). Changes in the internal structure of root cells can affect the normal physiological growth of plants (Läuchli and Grattan 2014). Previous studies have shown that when plants are exposed to salt stress, root cells will undergo nuclear disintegration (Smith et al. 1982) and plasmolysis, and cell walls becomes irregular (Kumar and Kumar 2014).
In this study, the internal structure of E. angustifolia root cells under severe salt stress had more organelles and intact cytoplasmic membranes and no plasmic wall separation when treated with mycorrhiza symbionts. The results showed that mycorrhizae increased the salt tolerance of E. angustifolia and reduced the salt stress injury to the root cells of plants. The reason may be that under severe salt stress, R. irregularis limits plant absorption of sodium ions, thus avoiding plant toxicity (Goussi et al. 2018). Moreover, the root cells of E. angustifolia were damaged by a large amount of Na+ in the absence of mycorrhizae. In this study, we also found that the roots of seedlings inoculated with R. irregularis could inhibit a large amount of Na+ intake, so the root cells were less damaged. There are few studies on the ultrastructure of E. angustifolia roots inoculated with AMF under salt stress. Fan et al. (2018) reported that inoculation with F. mosseae partially maintains the grana stacking under atrazine stress. However, the mechanism of salt tolerance conferred by AMF to E. angustifolia needs to be further studied by analysis of ultrastructure and molecular physiological changes.
Effects of salt stress on enzyme activities in E. angustifolia seedlings rhizosphere soil under different treatments
Soil enzymes are secreted by soil microbes (Black 1982). They act as catalysts in soil biochemical reactions and take part in many important biochemical processes in soil (Ai et al. 2015) such as plant absorption of water and nutrients (Yuhui et al. 2017), redox reactions, decomposition of humus (Eivazi et al. 2018), and the transformation of organic compounds and microbial morphology (Qi et al. 2016). In other words, soil enzymes are involved in the development and change of soil, and they are important indicators of soil activity (Du et al. 2018). The activities of SAC, PHO, URE, and CAT in soil can indicate the cycling of soil nutrients such as nitrogen, carbon and phosphorus (Acosta-Martinez et al. 2018). We show that with increasing salt concentration, the activity of soil enzymes showed a downward trend, indicating that salt stress inhibited the activity of the soil enzymes SAC, PHO, URE, and CAT. This trend is consistent with Raiesi and Sadeghi’s (2019) reports. However, under a given level of salt stress, the activity of soil enzymes in the rhizosphere of mycorrhizal E. angustifolia seedlings was higher than that of non-mycorrhizal seedlings. In particular, for SAC, URE, and CAT, the higher the concentration of salt stress, the greater the difference between enzyme activity in non-mycorrhizal and mycorrhizal treatments. The results showed that R. irregularis can increase soil enzyme activity, enhance soil vigor, slow down the toxic effects of salt stress and provide better conditions for the growth of host plants. This result also supports other related research (Ye et al. 2015; Chen et al. 2018).
Conclusion
The results of the present investigation indicate that R. irregularis enhances the salt tolerance of E. angustifolia by enhancing soil enzyme activity, which ameliorates the soil environment and reduces sodium ion content (and therefore toxicity) in roots. Alleviated salinity stress in E. angustifolia leads to greater plant height, basal diameter, root length, and root surface area. Hence, it is conceivable to conclude that the right combination of AMF and E. angustifolia can partially or completely alleviate the stress of salinity.
References
Abd_Allah EF, Hashem A, Alqarawi AA, Bahkali AH, Alwhibi MS (2015) Enhancing growth performance and systemic acquired resistance of medicinal plant Sesbania sesban (L.) Merr using arbuscular mycorrhizal fungi under salt stress. Saudi J Biol Sci 22(3):274–283
Abdel-Fattah GM, Rabie GH, Sh Lamis D, Rabab AM (2016) The impact of arbuscular mycorrhizal fungi on growth and physiological parameters of cowpea plants grown under salt stress conditions. Int J Appl Sci Biotechnol 4:372–379
Acosta-Martinez V, Cano A, Johnson J (2018) Simultaneous determination of multiple soil enzyme activities for soil health-biogeochemical indices. Appl Soil Ecol 126:121–128
Ai C, Liang G, Sun J, He P, Tang S, Yang S, Zhou W, Wang X (2015) The alleviation of acid soil stress in rice by inorganic or organic ameliorants is associated with changes in soil enzyme activity and microbial community composition. Biol Fertil Soils 51:465–477
Akhtar K, Wang W, Ren G, Khan A, Feng Y, Yang G (2018) Changes in soil enzymes, soil properties, and maize crop productivity under wheat straw mulching in Guanzhong, China. Soil Tillage Res 182:94–102
Alqarawi AA, Abd_Allah EF, Hashem A (2014a) Alleviation of salt-induced adverse impact via mycorrhizal fungi in Ephedra aphylla Forssk. J Plant Interact 9(1):802–810
Alqarawi AA, Hashem A, Abd_Allah EF, Alshahrani TS, Huqail AA (2014b) Effect of salinity on moisture content, pigment system, and lipid composition in Ephedra alata Decne. Acta Biol Hung 65(1):61–71
Aroca R, Ruizlozano JM, Angel MZ, José AP, José MG, María JP et al (2013) Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants. J Plant Physiol 170(1):47–55
Asgarzadeh M, Vahdati K, Lotfi M, Arab M, Babaei A, Naderi F, Pir Soufi M, Rouhani G (2014) Plant selection method for urban landscapes of semi-arid cities (a case study of Tehran). Urban For Urban Green 13(3):450–458
Babu MA, Singh D, Gothandam KM (2012) The effect of salinity on growth, hormones and mineral elements in leaf and fruit of tomato cultivar PKM1. J Anim Plant Sci 22:159–164
Bheemareddy VS, Lakshman HC (2011) Effect of salt and acid stress on Triticum aestivum inoculated with Glomus fasciculatum. J Anim Plant Sci 7:945–956
Black CA (1982) Methods of soil analysis. Part 2. Chemical and microbiological properties. American Society of Agronomy, Madison
Brundrett M, Bougher N, Dell B, Grove T, Malajczuk N (1996) Working with mycorrhizas in forestry and agriculture. RePEc
Buerkert AB, Bagayoko M, George E, Romheld V (2000) Effects of mycorrhizae and phosphorus on growth and nutrient uptake of millet, cowpea and sorghum on a West African soil. J Agric Sci 135:399–407
Chang W, Sui X, Fan X, Jia T, Song F (2018) Arbuscular mycorrhizal symbiosis modulates antioxidant response and ion distribution in salt-stressed Elaeagnus angustifolia seedlings. Front Microbiol 9:652
Chen X, Ding Z, Tang M, Zhu B (2018) Greater variations of rhizosphere effects within mycorrhizal group than between mycorrhizal group in a temperate forest. Soil Biol Biochem 126:237–246
Du Z, Zhu Y, Zhu L, Zhang J, Li B, Wang J, Wang J, Zhang C, Cheng C (2018) Effects of the herbicide mesotrione on soil enzyme activity and microbial communities. Ecotoxicol Environ Saf 164:571–578
Ebrahim MKH, Saleem A (2017) Alleviating salt stress in tomato inoculated with mycorrhizae: photosynthetic performance and enzymatic antioxidants. J Taibah Univ Sci 11(6):850–860
Eivazi F, Afrasiabi Z, Jose E (2018) Effects of silver nanoparticles on the activities of soil enzymes involved in carbon and nutrient cycling. Pedosphere 28(2):209–214
Elhindi KM, Sharaf El-Din A, Elgorban AM (2017) The impact of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects in sweet basil (Ocimum basilicum L.). Saudi J Biol Sci 24:170–179
Elsheikh EAE, Wood M (1995) Nodulation and N2 fixation by soybean inoculated with salt-tolerant rhizobia or salt-sensitive bradyrhizobia in saline soil. Soil Biol Biochem 27(4):657–661
Fan X, Chang W, Feng F, Song F (2018) Responses of photosynthesis-related parameters and chloroplast ultrastructure to atrazine in alfalfa (Medicago sativa L.) inoculated with arbuscular mycorrhizal fungi. Ecotoxicol Environ Saf 166:102–108
Gamalero E, Berta G, Massa N, Glick BR, Lingua G (2010) Interactions between Pseudomonas putida UW4 and Gigaspora rosea BEG9 and their consequences for the growth of cucumber under salt-stress conditions. J Appl Microbiol 108(1):236–245
Goussi R, Manaa A, Derbali W, Cantamessa S, Abdelly C, Barbato R (2018) Comparative analysis of salt stress, duration and intensity, on the chloroplast ultrastructure and photosynthetic apparatus in Thellungiella salsuginea. J Photochem Photobiol B Biol 183:275–287
Guan SY (1986) Soil enzymes and their research methods. Agricultural Press, Beijing
Hammer EC, Rillig MC (2011) The influence of different stresses on glomalin levels in an arbuscular mycorrhizal fungus—salinity increases glomalin content. PLoS ONE 6(12):e28426
Hashem A, Abd_Allah EF, Alqarawi AA, Aldubise A, Egamberdieva D (2015) Arbuscular mycorrhizal fungi enhances salinity tolerance of Panicum turgidum Forssk by altering photosynthetic and antioxidant pathways. J Plant Interact 10:230–242
Hashem A, Alqarawi AA, Radhakrishnan R, Al-Arjani ABF, Aldehaish HA, Egamberdieva D et al (2018) Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J Biol Sci 25(6):1102–1114
Hashem A, Fathi Abd_Allah E, Alqarawi AA, Wirth S, Egamberdieva D (2019) Comparing symbiotic performance and physiological responses of two soybean cultivars to arbuscular mycorrhizal fungi under salt stress. Saudi J Biol Sci 26(1):38–48
Heuer B (2005) Photosynthetic carbon metabolism of crops under salt stress. In: Pessarakli M (ed) Handbook of photosynthesis, 2nd edn. Taylor & Francis Group, Boca Raton, pp 776–789
Huaran H, Hao L, Feihu L (2018) Seed germination of hemp (Cannabis sativa L.) cultivars responds differently to the stress of salt type and concentration. Ind Crops Prod 123:254–261
Jia TT, Chang W, Fan XX, Song FQ (2018) Effects of arbuscular mycorrhizal fungi on photosynthetic and chlorophyll fluorescence characteristics in Elaeagnus angustifolia seedlings under salt stress. Acta Ecol Sin 38:1337–1347
Koske RE, Gemma JN (1989) A modified procedure for staining roots to detect VA mycorrhizas. Mycol Res 92(4):486–488
Kumar SP, Kumar CV (2014) Impact of cinnamic acid on physiological and anatomical changes in Maize Plants (Zea mays L.) grown under salinity stress. J Stress Physiol Biochem 10(2):44–54
Kumar A, Dames JF, Gupta A, Sharma S, Gilbert JA, Ahmad P (2014) Current developments in arbuscular mycorrhizal fungi research and its role in salinity stress alleviation: a biotechnological perspective. Crit Rev Biotechnol 35:461–474
Läuchli A, Grattan SR (2014) Plant abiotic stress: salt. In: Encyclopedia of agriculture and food systems. pp 313–329
Liang W, Ma X, Wan P, Liu L (2018) Plant salt-tolerance mechanism: a review. Biochem Biophys Res Commun 495(1):286–291
Marx MC, Wood M, Jarvis SC (2001) A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol Biochem 33(12):1633–1640
Metwally RA, Abdelhameed RE (2018) Synergistic effect of arbuscular mycorrhizal fungi on growth and physiology of salt-stressed Trigonella foenum-graecum plants. Biocatal Agric Biotechnol 16:538–544
Navarro JM, Pérez-Tornero O, Morte A (2014) Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the rootstock salt tolerance. J Plant Physiol 171(1):76–85
Ouziad F, Wilde P, Schmelzer E, Hildebrandt U, Bothe H (2006) Analysis of expression of aquaporins and Na+/H+ transporters in tomato colonized by arbuscular mycorrhizal fungi and affected by salt stress. Environ Exp Bot 57(1):177–186
Qi R, Li J, Lin Z, Li Z, Li Y, Yang X et al (2016) Temperature effects on soil organic carbon, soil labile organic carbon fractions, and soil enzyme activities under long-term fertilization regimes. Appl Soil Ecol 102:36–45
Raiesi F, Sadeghi E (2019) Interactive effect of salinity and cadmium toxicity on soil microbial properties and enzyme activities. Ecotoxicol Environ Saf 168:221–229
Ren BH, Wu SL, Chen BD, Wu ZX, Zhang X (2015) Cr stable isotope fractionation in arbuscular mycorrhizal dandelion and Cr uptake by extraradical mycelium. Pedosphere 25(2):186–191
Shao T, Gua X, Zhua T et al (2019) Industrial crop Jerusalem artichoke restored coastal saline soil quality by reducing salt and increasing diversity of bacterial community. Appl Soil Ecol 138:195–206
Shekoofeh E, Sepideh H, Roya R (2012) Role of mycorrhizal fungi and salicylic acid in salinity tolerance of Ocimum basilicum resistance to salinity. Afr J Biotechnol 11(9):2223–2235
Smith SE, Read DJ (2008) Mycorrhizal symbiosis, 3rd edn. Academic Press, London
Smith MM, Hodson MJ, Öpik H, Wainwright SJ (1982) Salt-induced ultrastructural damage to mitochondria in root tips of a salt-sensitive ecotype of Agrostis stolonifera. J Exp Bot 33(136):886–895
Song F, Qi D, Liu X, Kong X, Gao Y, Zhou Z, Wu Q (2015a) Proteomic analysis of symbiotic proteins of Glomus mosseae and Amorpha fruticosa. Sci Rep 5:18031
Song F, Sun YX, Fan XX, Chang W (2015b) An arbuscular mycorrhizal fungus improving salt-alkali resistance of Elaeagnus angustifolia. Chinese patent: CN104560730A
Sun YF, Song FQ, Chang W, Fan XX (2016) Effect of arbuscular mycorrhizal fungi on growth and physiology of Elaeagnus angustifolia seedlings subjected to salinity stress. Sci Silvae Sin 52:18–27
Usman ARA (2015) Influence of NaCl-induced salinity and Cd toxicity on respiration activity and Cd availability to barley plants in farmyard manure-amended soil. Environ Soil Sci Appl. https://doi.org/10.1155/2015/483836
Xu Z, Ban Y, Jiang Y, Zhang X, Liu X (2016) Arbuscular mycorrhizal fungi in wetland habitats and their application in constructed wetland: a review. Pedosphere 26(5):592–617
Ye S, Yang Y, Xin G, Wang Y, Ruan L, Ye G (2015) Studies of the Italian ryegrass–rice rotation system in southern China: arbuscular mycorrhizal symbiosis affects soil microorganisms and enzyme activities in the Lolium mutiflorum L. rhizosphere. Appl Soil Ecol 90:26–34
Yin J, Jia J, Lian Z et al (2019) Silicon enhances the salt tolerance of cucumber through increasing polyamine accumulation and decreasing oxidative damage. Ecotoxicol Environ Saf 169:8–17
Younesi O, Moradi A, Namdari A (2013) Influence of arbuscular mycorrhiza on osmotic adjustment compounds and antioxidant enzyme activity in nodules of salt-stressed soybean (Glycine max). Acta Agric Slov 101(2):219–230
Yuhui W, Denghua Y, Junfeng W, Yi D, Xinshan S (2017) Effects of elevated CO2 and drought on plant physiology, soil carbon and soil enzyme activities. Pedosphere 27(5):846–855
Zhang H, Qin P, Zhang W (2014) Effects of inoculation of arbuscular mycorrhizal fungus and apophysomyces spartina on P-uptake of castor oil plant (Ricinus communis L.) and rhizosphere soil enzyme activities under salt stress. Agric Sci Technol 15(4):659–664
Zhang X, Li G, Du S (2018) Simulating the potential distribution of Elaeagnus angustifolia L. based on climatic constraints in China. Ecol Eng 113:27–34
Zheng L, Zhang M, Xiao R, Chen J, Yu F (2017) Impact of salinity and Pb on enzyme activities of a saline soil from the Yellow River delta: a microcosm study. Phys Chem 97:77–87
Zhu Y, Gong H (2014) Beneficial effects of silicon on salt and drought tolerance in plants. Agron Sustain Dev 34:455–472
Acknowledgements
This work was supported by Key Laboratory of Microbiology, College of Heilongjiang Province.We would also like to thank Elizabeth Tokarz at the Yale University for her assistance with English language and grammatical editing of the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: This research was financially supported by the Special Fund for Forest Scientific Research in the Public Welfare (201504409), the National Natural Science Foundation of China (31570635), Study on Microbial Control Techniques of Agricultural Non-point Source Pollution in Heilongjiang Province (TD2019C002), and National Sugar Crops Technical System “Sugarbeet Planting System” in China (CARS-170209).
The online version is available at http://www.springerlink.com
Corresponding editor: Yu Lei.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
He, W., Fan, X., Zhou, Z. et al. The effect of Rhizophagus irregularis on salt stress tolerance of Elaeagnus angustifolia roots. J. For. Res. 31, 2063–2073 (2020). https://doi.org/10.1007/s11676-019-01053-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-019-01053-1