
Abstract
Since the outbreak of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) that caused the coronavirus disease-19 (COVID-19), in December 2019, the infection has spread around the globe. Some of the risk factors include social distancing, mask wearing, hand washing with soap, obesity, diabetes, hypertension, asthma, cardiovascular disease, and dysbiosis. Evidence has shown the incidence of total infection and death rates to be lower in sub-Saharan Africa when compared with North Africa, Europe and North America and many other parts of the world. The higher the metabolic syndrome rate, the higher the risk of SARS-CoV-2 infection. Africa has a lower rate of metabolic syndrome risk than many other continents. This paradox has puzzled several in the biomedical and scientific communities. Published results of research have demonstrated the exciting correlation that the combination of young age of the population coupled with their native plant-based diet has lowered their risk factors. The plant-based diet include whole grains (millet, sorghum), legumes (black-eye peas, dry beans, soybean), vegetables, potato, sweet potato, yams, squash, banana, pumpkin seeds, and moringa leaves, and lower consumption of meat. The plant-based diet results in a different gut microbiota than of most of the rest of the world. This has a significant impact on the survival rate of other populations. The “plant-based diet” results in lower rates of obesity, diabetes and dysbiosis, which could contribute to lower and less severe infections. However, these hypotheses need to be supported by more clinical and biostatistics data.
Similar content being viewed by others


Introduction
Since the initial December 2019 outbreak and spread of the severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) and COVID-19 worldwide pandemic, the incidence during the first wave of SARS-CoV-2 infection and COVID-19 remained lower in sub-Saharan Africa than many parts of the world (Fig. 1, Table 1). In both waves, with the exception of South Africa, the incidence remained remarkably low in sub-Saharan African countries compared to Europe, North America, South America, and India. In most sub-Saharan African countries, the incidence of SARS-CoV-2 infection and COVID-19 is confined to major metropolitan areas like Lagos, Kinshasa, Abidjan, Nairobi, Addis Ababa, Dakar, Accra, or Bamako. However, despite the dense populations in Lagos (14,368,000), Kinshasa (14,340,000), Abidjan (5,203,000), Nairobi (4,735,000), Addis Ababa (4,794,000), or Dakar (3,140,000), SARS-CoV-2 infectivity is lower than Sao Paolo with a population of 22,000,000, New York with a population of 18,804,000, Rio with a population of 13,458,075, London with a population of 9,304,000, or New Delhi with a population of 30,291,000 [https://coronavirus.jhu.edu/map.html] (Fig. 1 and Table 1). With the exception of South Africa, other sub-Saharan African countries have some of the most poorly equipped healthcare systems and testing infrastructure for the infection in the world. Public health experts and the media have tried to explain the puzzling low SARS-CoV-2 infection and COVID-19 pandemic in most sub-Saharan Africa during the first wave of the pandemic [1,2,3,4] [https://www.washingtonpost.com/world/africa/africa-coronavirus-low-cases-deaths/2020/12/10/e907a1c2-3899-11eb-aad9-8959227280c4_story.html]. Others predicted that the mortality and morbidity will be higher from SARS-CoV-2 in sub-Saharan Africa than other parts of the world because of influx of 2 million Chinese nationals now living and working in Africa [5]. This is in combination with the widespread of communicable diseases in the continent [5]. COVID-19-induced mortality in sub-Saharan Africa was less than 1% of the first global COVID-19-induced mortality [https://coronavirus.jhu.edu/map.html]. By 2021, the continent was estimated to have less than 5% of the world cases [6]. These observations are puzzling the scientific community around the world. It has been suggested that the low rate of COVID-19 related symptoms and fatality in sub-Saharan Africa may be associated with a number of factors including host genetics, previous exposures to viral infections, lack of testing, misdiagnosis, relatively high temperatures on the continent, and the young age distributions of the populations in the region [4, 7].

SARS-CoV-2 incidence or total cases (TC), COVID-19 related total death (TD), and death per million population (DM) in sub-Saharan African countries
We hypothesize that the combination of younger age distribution, plant-based diet, and gut microbiota will attenuate the infectivity in the region. South Africa appears to be an outlier, or the exception to the hypothesis and will be explained later in this review. To support this hypothesis, we evaluated current literature on (1) The mechanism of SARS-CoV-2 infection and potential dietary inhibitors, (2) The younger age distribution of the sub-Saharan African population compared to the Americas, Europe, and India as the major risk factor for SARS-CoV-2 infection and COVID-19 onset, (3) Diet and gut microbiota in sub-Saharan Africa versus dysbiosis as a risk factor for SARS-CoV-2 infection and COVID-19 onset worldwide, and (4) The association of major risk factors and comorbidities for SARS-CoV-2 and COVID-19 recently observed in sub-Saharan Africa compared to the rest of the world.
The Mechanism of SARS-CoV-2 Infection and Potential Dietary Inhibitors
SARS-CoV-2 infection occurs when SARS-CoV-2 laden-droplets from an infected person are transmitted to another individual. SARS-CoV-2 uses serine proteases to prime the virus and allow the virus to attach to the receptor angiotensin converting enzyme-2 (ACE2). The ACE2 receptor is expressed at high levels in nasopharynx, alveolar type II cells in the lungs, as well as on myocardial cells, esophagus keratinocytes, intestinal (ileum and colon) epithelial cells, kidney, pancreas, bladder urothelial cells, nervous system, and skin epithelial cells. Inhibiting the binding of the virus to the cell receptor prevents development of the infection. If the virus is successful in attaching to the cell receptor after being primed/activated or after endocytosis the virus is able to penetrate the cell, replicate, resulting in the cascade of events, cause COVID-19. Trypsin-like proteases are commonly found in the respiratory tract [8]. High levels of ACE2 receptors are found in the respiratory tract which combined with serine proteases facilitate priming and binding of SARS-CoV-2 to the tissues. The severity of COVID-19 can be exacerbated by old age, obesity, diabetes, hypertension, cardiovascular disease, kidney disease, asthma, allergies, compromised immune system, and dysbiosis. Individuals with these conditions have impaired endothelial cells, dysbiosis, pre-existing systemic low-grade inflammation, increased expression of SARS-CoV-2 entry receptors including ACE2 and CD147, and elevated levels of serum proteases. For example, transmembrane protease serine 2 (TMPRSS2) that can catalyze SARS-CoV-2 activation [9]. Strategies to inhibit proteases that prime the SARS-CoV-2 virus for entry into the host cells are targets for nonvaccine or antiviral treatment for COVID-19 until newly introduced vaccines prove their efficacies [10]. These priming proteases may be inactivated by bioactive compounds found in foods that most sub-Saharan Africans consume. Below is the analysis of the association of SARS-CoV-2 infection and COVID-19 risk factors and infection incidence, disease onset, and mortality in sub-Saharan Africa.
The Age Distribution of the Sub-Saharan Africa Population as Major Risk Factor for SARS-CoV-2 Infection and COVID-19
SARS-CoV-2 severely infects elderly people (age 55 years and older), and manifestations of the COVID-19 infection is often more severe in elderly population regardless of race. Immunosenescence or impaired immune response is observed in the elderly more than young people. Japan has an older population distribution compared to the rest of the world. Throughout the first wave of the COVID-19, Japan where masks have been extensively used, reported lower infection and death rate among its old population than similar old populations in Asian countries. The Japanese populations used face masks more extensively than other Asian countries and Western nations where masks were not used before the pandemic [11]. Twenty-three percent of the Italian population is 65 years old or higher and Italy has one of the highest COVID-19 infection rate since the spring of 2020 [12]. Old age is a major risk factor for SARS-CoV-2 infection and severe COVID-19 because dysbiosis and reduced microbiota diversity are frequent in old people and predisposes these populations to COVID-19 severity. Older people in general throughout the world are at higher risk of SARS-CoV-2 infection and COVID-19 than younger people. The population distribution in sub-Saharan Africa does not have as many elderly people as the countries in the rest of the world. The median age of the population in sub-Saharan Africa is estimated at 19.7 years [13]. The median age of the population in South Africa, Laos, Italy, Belgium, Brazil, and Canada, is 28, 24, 47, 41, 33, and 41 years, respectively https://www.indexmundi.com/. As seen in Fig. 1 and Table 1, the death rate in the Democratic Republic of Congo, Uganda, Rwanda, or Burundi is much lower than in Italy, Brazil, or Mexico. Young people have stronger immune system than older adults, may develop symptomatic or mild infection that may be undetected until resolved [13]. SARS-CoV-2 infection in young adults is not associated with the loss of ACE anti-inflammatory and anti-oxidative activity. As a result, young patients do not present with acute respiratory distress syndrome (ARDS) as do old people.
Diet and Dysbiosis in Sub-Saharan Africa vs Dysbiosis as a Major Risk Factor for SARS-CoV-2 Infection and COVID-19
In sub-Saharan Africa, the staple diet is rich in plant-based diets including whole grains (millet, sorghum), legumes (black-eye peas, dry beans, soybean), vegetables, potato, sweet potato, yams, squash, banana, pumpkin seeds, and moringa leaves, and lower consumption of meat [14]. The plant foods are excellent sources of protease inhibitors including the trypsin and trypsin-chymotrypsin inhibitors, that are bioavailable and can prevent SARS-CoV-2 from attaching to the host cells [15]. Some of these plant foods such as the squash are also excellent sources of protease inhibitors that can help prevent blood clotting, a death causing factor in COVID-19 [16]. Major bean (Phaseolus) consuming countries in the world include Rwanda, Uganda, Tanzania, Burundi, and Costa Rica. As of May 22, 2021, the corresponding populations, confirmed cases of COVID-19, and COVID-19 related deaths were 12.3 million; 26,602; 349 in Rwanda, 46.13 million; 43,370; 350 in Uganda, 59.7million; 509; 21 in Tanzania, 11.9 million; 4,451; 6 in Burundi, and 5.0 million; 299,219; 3,765 in Costa Rica, respectively [https://coronavirus.jhu.edu/map.html]. In Uganda, Rwanda, and Burundi, colored beans are eaten three times a day in almost every household. The health benefits of cooked beans cannot be overstated. The protease inhibitors found in dry beans are also present in soybean and soy products. Major soymilk consuming countries include Hong Kong, Singapore, Thailand, China, and Malaysia. As of May 22, 2021, Hong Kong with 14 million population had 11,830 confirmed COVID-19 cases, and 210 COVID-19 related deaths. Singapore with 5.7 million people had 61,770 confirmed cases and 32 deaths. Thailand with 69.8 million people had 126,118 confirmed cases, and 759 deaths. China with 1.43 billion people, had 90,954 confirmed cases and 4,636 deaths. Malaysia with 32.5 million people, 498,796 confirmed cases, and 2,149 deaths [https://coronavirus.jhu.edu/map.htm]. Major tofu consuming countries include China, Japan, South Korea, Vietnam, and Philippines. China with a populations, of 1.43 billion had 90,954 confirmed cases, and three deaths per million. Japan had 126.3 million; people, 704,159 confirmed cases and 96 deaths per million. South Korea had 51.7 million people, 135,344 confirmed cases and 38 deaths per million. Vietnam had 97.5 million, 41 confirmed cases, and 0.4 deaths per million. The Philippines had 108,900 million people, 1,171,403 confirmed cases and 178 deaths per million [https://www.worldometers.info/coronavirus/?utm_campaign=instagramcoach1]. Laos with a population of 7.3 million had 1,763 confirmed COVID-19 cases and 2 COVID-19 related death. Cambodia has a population of 16.8 million, 24,157 cases, and 165 COVID-19 related death. TMPRSS2 gene expression is reduced by the isoflavone genistein which is prevalent in soy products including soymilk, tofu, miso, and soy flour [17]. TMPRSS2 gene is more highly expressed in Caucasians than Asians or African men [18]. In summary, countries with high per capita consumption of dry beans or soy products have shown low infection and death rate.
Banana (Musa species) is a major staple in Uganda, Rwanda, and many other countries. Africans consume 21 kg of banana and plantain per capita while the world per capita is about 11.7 kg. In Uganda, the per capita for banana and plantain is 191 kg [https://www.iita.org/]. As of November 14, 2020, Uganda had 46.13 million people, 15,402 confirmed cases and 144 deaths. Rwanda had 12.3 million people, 5,362 confirmed cases and 42 deaths. Burundi had 11.9 million people, 624 confirmed cases and 1 death. Unripened or ripened bananas contain an inhibitor of trypsin, chymotrypsin, and papain, and a heat-stable inhibitor of ACE [19, 20]. Banana is a whole-fruit which provides prebiotics and in vitro data show that banana powder can produce anti-inflammatory postbiotics short chain fatty acids (SCFA) including acetate, propionate, and butyrate [21].
Sweet potato (Ipomoea batatas) is heavily consumed in Nigeria, Tanzania, Ethiopia, and Angola. As of May 22, 2021, populations, confirmed cases, and deaths in these countries were respectively 207.5 million; 165,944; 2,067, 58.73 million; 509; 21, 114.96 million; 268,520; 4,060, and 32.86 million;31,909; 709 [https://www.worldometers.info/coronavirus/?utm_campaign=instagramcoach1?].
Sporamin A and B are the major storage proteins in sweet potato and endowed with a defense role as a Kunitz trypsin inhibitor [22]. Sweet potato also contains anti-coagulants coumarins, scopoletin, aesculetin, and umbelliferone. Purple sweet potato is a good source of pelargonidin, an anticoagulant phenolic compound [23].
Yam (Dioscorea) is mostly consumed in West, Central, and East Africa. Dioscorin is the major yam storage protein and is a low temperature stable inhibitor of trypsin [24]. Yam is a good source of dietary fiber for SCFA production in the gut [25]. The low infectivity and death incidence in African countries including Uganda, the Democratic Republic of Congo, and Ivory Coast may be explained in part by high consumption of combination of yams, beans, melon and pumpkin seeds, and sorghum. All these plant foods are very rich in trypsin inhibitors and when cooked at temperature below 100 ℃, these foods preserve the trypsin inhibitory activity.
When minimally processed, the foods mentioned above can be good sources of bioactives including the Kunitz trypsin inhibitor, cystatin, and cathepsin inhibitors that can inhibit SARS-CoV-2 infection [15, 16]. The plant bioactives mentioned above can stand high temperature of 100 ℃ for 5–10 min but are heat stable at lower temperatures. In Africa, most foods are boiled and not canned or retorted at 121 ℃. The bioactive proteins in the legumes are anti-inflammatory and health-enhancing [26]. Beans are excellent sources of all the proteases mentioned early. Colored beans are also good sources of pelargonidin known for its in vivo antithrombic and antiplatelet properties [27]. Polyphenols in colored Phaseolus beans inhibit DPP4, α-amylase, α-glucosidase, and reactive oxygen species making colored beans a good food for diabetics who are at risk of SARS-CoV-2 infection [28]. Plant protease inhibitors prevent obesity [29] and inhibit enzymes associated with chronic diseases invasion and proliferation [30]. These plant-based foods are good sources of protease inhibitors and anticoagulants.
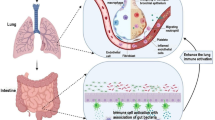
Dysbiosis is the imbalance in favor of maladaptive and pathogenic bacteria in the host gut microbiota. Dysbiosis is significantly associated with all the identified major risk factors for COVID-19 including old age, obesity, diabetes, hypertension, cardiovascular disease, and compromised immune system [31]. Dysbiosis contributes to the course of COVID-19 because 70 to 80% of the body immune system is in the gastrointestinal tract and the immunological health of the gut that is reflected by the gut microbiota influences lung health through gut-lung axis [32]. Dysbiosis results in uncontrolled intestinal inflammation, a compromised immune system which exacerbates COVID-19. Dysbiosis increases gut permeability and the leakage of pro-inflammatory lipopolysaccharides and pro-inflammatory cytokines in the blood that can exacerbate the cytokine storm and organ failure. Short or long-term consumption of diet that consists of high-fat, high meat fat and proteins, high reducing sugars, and low plant-based proteins and fiber, rapidly and reproducibly alters the human gut microbiome, causes the disruption of gut microbiota homeostasis, and develops dysbiosis [33]. The distribution and dominance of microbiota in the diet are rapidly altered by changes in diet. Dysbiosis significantly affects severe COVID-19 patients. Dysbiosis is cause and consequence of COVID-19. The GI tract is exposed to any pathogen or antigen that is introduced into the respiratory system suggesting that SARS-CoV-2 virus that enters through the nose can also move to the gastrointestinal tract [34]. Both the lung and gastrointestinal tract are major sites of ACE2 receptors. In the lungs, following attachment of SARS-CoV-2 to ACE2-rich cells and endocytosis, if viral clearance is impaired, the immune system progresses towards cytokine release and cytokine storm. Enterobacteriaceae, a family of pathogenic bacteria found in gut dysbiosis were identified in the lungs from diseased COVID-19 patients [35]. This suggests that cytokines released by these bacteria in the lungs are similarly released in the gut and the gut-lung axis exacerbates disease severity in individuals with gut dysbiosis and co-morbidities.
To support the plant-based diet and gut microbiota hypothesis of low SARS-CoV-2 infectivity in sub-Saharan Africa, recent findings on gut microbiota of sub-Saharan Africans may hold some explanation. De Filippo et al. (2010) investigated and compared human intestinal and fecal microbiota from European children from urban area of Florence, Italy and age-matched children from a rural African village of Burkina Faso [36]. The children from rural Burkina Faso somehow represented the children in most rural settings in sub-Saharan Africa while the children from Florence represented those in most Western countries’ settings. By using high throughput 16 rDNA sequencing and other biochemical analyses, De Filippo et al. found high levels of Bacteroidetes and very low levels of Firmicutes and abundance of Prevotella and Xylanibacter bacteria and more anti-inflammatory short chain fatty acids in the gut of Burkina Faso children than the Italian children [36]. Gut microbiome composition of people living on plant-based diet in sub-Saharan Africa and profiled through 16SrRNA gene analysis using 454- pyrosequencing of stool analyses revealed a higher gut microbiota diversity dominated by Actinobacteria (Bifidobacterium); Bacteroidetes (Bacteroides–Prevotella); Firmicutes (C. histolyticum, Eubacterium, Oscillibacter, Butyricicoccus and Sporobacter); Proteobacteria (Succinivibrio, Shigella and Escherichia) and Spirochaetes (Treponema) and depleted in only Actinobacteria (Bifidobacterium catenulatum), Firmicutes (Clostridium difficile and Akkermansia muciniphila [37, 38]. Meanwhile the gut microbiota of people in western countries showed abundance of Firmicutes (Blautia, Dorea, Roseburia, Faecalibacterium, Ruminococcus, Oscillospira, C. perfringens, C. difficile and S. aureus); Bacteroidetes (Bacteroides), Actinobacteria (B. adolescentis and B. catenulatum) and Verrucomicrobia (A. muciniphila)[38]. The presence of a highly diverse microbiota in sub-Saharan African populations who consume plant-based diet was confirmed by Vipperla and O'Keefe who showed the prevalence of Prevotella in the gut microbiome of the Africans compared to Africa- American whose gut microbiota was dominated by Bacteroides [39].
Diet has a major effect on the composition of the gut microbiota. The diet in Burkina Faso and many other sub-Saharan African countries is mostly plant-based using whole grain (millet, sorghum), legumes (black-eye peas, dry beans, soybean), vegetables, and little meat. The plant-based diet is rich in protease inhibitors, fiber, prebiotics, phenolic compounds, minerals, all of which improve gut health. The fiber, prebiotics, and phenolics are fermented in the colon to produce anti-inflammatory short chain fatty acids acetate, butyrate, and propionate, and urolithins. In Florence, the children consume typical western diet associated with high consumption of animal protein and fat, refined starch, reducing sugars, and low in plant-based ingredients. In recent work, De Filippo et al. showed that when Africans move from rural areas to major cities and adopt the Western type of diet, the gut microbiota profile changes with progressive reduction in anti-inflammatory short chain fatty acids such as butyrate and propionate [40]. The rate of dysbiosis and COVID-19 is higher in major sub-Saharan African urban areas than rural areas. SARS-CoV-2 infectivity and COVID-19 is high in African- Americans and low in sub-Saharan Africans. The high infection and fatality rates in South Africa can in part be explained by the high levels of processed foods including sweets and animal products and less unprocessed plant-based diet consumed in the country unlike other sub-Saharan African countries [41]. As a result, obesity, diabetes, hypertension, and other noncommunicable chronic degenerative diseases are higher in South Africa compared to the rest of the sub-Saharan Africa. The higher the metabolic syndrome rate, the higher the risk of SARS-CoV-2 infection and COVID-19 development. Regular consumption of properly processed plant-based diet will improve gut microbiota and attenuate SARS-CoV-2 infection.
The Association of Major Risk Factors for SARS-CoV-2 Infection and COVID-19 and Current field Results in Sub-Saharan Africa
Social Distancing, Mask Wearing, and Hand Washings with Soap
Social distancing, quarantine, mask wearing, and frequent hand washings with soap are the best practices to reduce the spread of SARS-CoV-2 infection. When these protective measures fail, COVID-19 has an increased likelihood of onset and progression. Densely populated areas are risk factors for rapid and uncontrolled SARS-CoV-2 infection dissemination and COVID-19 onset and propagation. Lagos, Kinshasa, Abidjan, Nairobi, Abuja, or Dakar are densely populated, yet these cities have lower rate of infectivity than comparable cities across South America, Europe, North America, and Asia with similar or high/dense populations. The second wave of COVID-19 that started in November 2020 has been reported to be associated with an increase in the rate of infection, yet the total infection and death rates in Africa are insignificant compared to total infection and death rates in Western Europe and North America (Fig. 1 and Table 1). Comparisons of results during the first wave and the second wave show that Sub-Saharan Africa still has a low rate of SARS-CoV-2 infection and COVID-19 related death (Fig. 1 and Table 1). In most African major cities, people are not abiding to social distancing, quarantine, mask wearing, and frequent hand washings with soap because most people work in informal business and cannot sustain a day without work [42,43,44,45].
Obesity
Obesity is a major risk factor for severe SARS-CoV-2 infection and COVID-19. SARS-CoV-2 severely infects obese subjects more than lean individuals [46, 47]. Diet-induced metabolic alterations including obesity play a significant role in the incidence and severity of COVID-19[48,49,50]. Adipose tissue is a key endocrine organ that secretes a plethora of factors including adipokines, chemokines, and cytokines that profoundly impact metabolism and immune system. Acute inflammation from COVID-19 may amplify existing chronic inflammation secondary to obesity and lead to more severe disease and poorer outcomes [51]. Furthermore, metabolic changes associated with obesity such as insulin and leptin resistance negatively impact immune cell function. All these changes substantially influence the immune cell growth and proliferation and affect glucose metabolism and activation which ultimately results in impairment of host immune defense.
Although lung ACE2 is a major target for SARS-CoV-2 infection, the higher expression of ACE2 gene expression in human visceral and subcutaneous adipose tissue than human lung makes obese individuals more vulnerable to the infection and disease than lean individuals [52]. Obese individuals have higher levels of interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), ACE2 on adipocytes, leptin, lower concentration of adiponectin, hyperactivated mechanistic target of rapamycin (mTOR), natural killer cells polarized to non-cytotoxic, all of which can increase the duration of SARS-CoV-2 shedding [47]. SARS-CoV-2 virion from obese individuals is higher than virion from old people than virion from individuals with cardiovascular disease than virion from individuals with lung disease than virion from individuals with diabetes than virion from individuals with hypertension than virion from smokers than virion from healthy individuals and the size of virion is associated with the odds of developing severe viral infection [53]. The BMI of severe COVID-19 patients median have been reported to be higher than the BMI of non-severe patients. Obese individuals are prone to excessive oxidative stress, diminished airway flow, and severe obese individuals are at risk of heart failure. Like the rest of the world, Sub-Saharan Africa is experiencing a rise in obesity because of increased urbanization, westernization of lifestyle, decreased physical activity, increased sedentary lifestyle, and less consumption of fruits and vegetables but high overindulgence on energy-dense processed foods in urban areas [54]. Descriptive data analysis of obesity prevalence by country showed higher adult obesity values in the Americas and Europe and lower values in Asia and Africa [55]. Several studies have observed the higher incidence of adult overweight and obesity in urban areas compared to rural regions of sub-Saharan Africa. [56]. While the incidence of obesity is increasing in major African cities, it is low (< 5% of its population) compared to the West or South East Asia. https://worldpopulationreview.com/country-rankings/most-obese-countries. With 1.038 billion people, sub-Saharan African has less than 30 million obese compared to Brazil which has more than 70 million obese people (33.5% of its population of more than 211 million people). The prevalence of obesity in US was 42.7% in 2017–2018 [www.cdc.gov] and about 28% in Canada in 2017 [https://obesitycanada.ca/guidelines/epidemiology. Accessed 1/26/2021]. India alone with 1.353 billion people has 5 million obese. China with 1,393 billion people has about 70 million obese. SARS-CoV-2 infectivity spreading, and fatality is higher in Europe, North America, Brazil, UK, and Mexico where obesity with median BMI > 25 kg/m2 is also highly prevalent [47]. Obesity is a major risk factor for severe SARS-CoV-2 infection and COVID-19 even in young people. SARS-CoV-2 infectivity and COVID-19 incidence are higher in major African cities than in the rural areas in part because processed diet-induced obesity is prevalent in urban areas than rural [41]. Mean BMI in Africa is < 22 kg/m2 [57]. The level of physical activity is higher in rural sub-Saharan African villages than in metropolitan African cities [58]. The interpersonal contacts are higher between dwellers of major sub-Saharan African cities and populations from countries with high infection rates than between rural African populations and populations external to the African region.
Diabetes
Diabetes is a major comorbidity risk factor for severe SARS-CoV-2 infection. Diabetic patients, especially those with uncontrolled blood glucose, are at greater risk for developing severe COVID-19 disease than healthy individuals. About 98% of adults with type 2 diabetes carry at least one or two comorbidities including hypertension, overweight/obesity, chronic kidney disease, or cardiovascular disease [59]. Each of these comorbidities is by itself a risk factor for severe COVID-19 outcome. The hyperglycemic environment of diabetic blood increases glucose secretion within the airway and promotes immune dysfunction by damaging neutrophil function, repressing the protective antioxidant/anti-inflammatory system and the humoral immunity, causing micro- and macro-angiopathies, neuropathy, and reducing the antibacterial activity of urine. Diabetes triggers a pro-inflammatory cytokine environment in the body associated with high levels of IL-6 and C-reactive protein. ACE2 is expressed at lower levels in the pancreas, kidneys, vascular system, and lungs of individuals with diabetes. Consequently, these individuals have reduced anti-inflammatory and anti-oxidative capabilities. All these alterations including compromised glycemic status, innate immune system, and humoral immunity inhibit resistance to any infection including SARS-CoV-2 infection. The interaction of SARS-CoV-2 with pancreatic ACE2 may lead to β-cell dysfunction-induced hyperglycemic state causing acute diabetes state [60]. Ex-vivo study has shown that lung monocytes and macrophages of COVID-19 infected people are highly glycolytic, facilitate SARS-CoV-2 replication and cytokine production. These changes initiate mitochondrial reactive oxygen species production and stabilizing the hypoxia-inducible factor-1 α (HIF-1α) [61]. HIF-1α-induced metabolic alterations in COVID-19 infected monocyte increase glycolysis and cytokine such as IL-1β production which drives T cell dysfunction and lung epithelial cell death. Increased circulating glucose levels in CoV-2 infected monocyte can promote T cell dysfunction and lung epithelial cell apoptosis in a mitochondrial-ROS/HIF-1α-dependent manner [61]. In sub-Saharan Africa, the prevalence of type 2 diabetes in adults older than 55 years is about 13% and higher in urban than rural settings [62]. While individuals with diabetes are at increased risk arising from severe complications of SARS-CoV-2, there are less people with diabetes in Africa (an estimated 15.5 million out of 1.2 billion people) [63] than China (about 110 million patients out of 1.4 billion people) than India (an estimated 120 million people out of 1.35 billion people) [64]. There are about 34 million people with diabetes in the US (about 10% of the population) [www.cdc.gov].
Hypertension
SARS-CoV-2 severely infects individuals with high blood pressure who display more than three-fold higher mortality and short-term mortality rate than normotensive individuals [65]. One potential mechanism of the contribution of hypertension to severe COVID-19 is as follows. Plasminogen activator inhibitor-1 (PAI-1) levels are high in hypertension, old age, obesity, diabetes, and cardiovascular disease [66]. Severe COVID-19 develops hypoxia in the lungs. Hypoxia, through hypoxia-inducible factor-1 alpha (HIF-1α), stimulates the production of PAI-1 in the lung [67]. Injury in the lungs of SARS-CoV-2 infected individuals leads to overproduction of hyaluronan and the latter also leads to production of more PAI-1. Excessive PAI-1 binds to toll-like receptor 4 (TLR4) on macrophages, activates NF-κB which in turn induces the secretion of pro-inflammatory cytokines and chemokines in the lungs of severely ill patients with COVID-19. The up-regulation of PAI-1 leads to coagulopathy characterized by intravascular thrombi [66]. The incidence of sequelae of uncontrolled hypertension, specifically stroke and hypertension heart disease, is greater in Africa than in many other parts of the world [68]. Hypertension afflicts about 46% of African adults aged 25 years and older [69]. There were 130 million African adults with hypertension in 2010 and the number is expected to increase to 220 million by 2030 [69]. Awareness for hypertension is < 30%, treatment < 20%, and control rate < 10% [69]. However, the rate of SARS-CoV-2 infection is low in this part of the world. SARS-CoV-2 infectivity and COVID-19 is low in Africa. In sub-Saharan Africa, COVID-19 severity and fatality have shown discordant association with the disease comorbidity risk factors including hypertension, diabetes, and cardiovascular disease whereas in North African countries such as Algeria and Egypt a positive association was established between the disease severity and the risk factors [70]. COVID-19 outcome in Africa may be associated with the modulation of the renin–angiotensin–aldosterone system (RAAS) axis, which controls ACE/ACE2 balance, in the region [70].
Cardiovascular
Pre-existing cardiovascular disease such as coronary heart disease is a comorbidity factor for severe COVID-19 outcomes. Globally speaking, cardiovascular disease disproportionately affects sub-Saharan Africans where the affected are younger, female, hypertensive and are chronic smokers [68]. Despite high prevalence of hypertension and smoking, SARS-CoV-2 infectivity and COVID-19 pandemic are low in the observed sub-Saharan African population. SARS-CoV-2 infected patients with pre-existing cardiovascular disease are a greater cardiac failure than healthy individuals. There is a discordant pattern between the high prevalence of cardiovascular disease in sub-Sahara Africa and COVID-19 severity and fatality [70].
Asthma in Sub-Saharan Africa, SARS-CoV-2 Infection, and COVID-19
Asthma is a risk factor for severe COVID-19 outcomes because asthmatics have deficient and delayed innate antiviral immune responses, and deficient and delayed lung cell interferon (IFN)‐α, IFN‐β, and IFN‐λ responses. SARS-CoV-2 infection, like asthma, induces an attenuated IFN-I and INF-III signature and therefore SARS-CoV-2 and the resultant unopposed TH2 response exacerbates asthma [71]. Asthma has an estimated high prevalence of 12% in Africa [72]. Gut and lung dysbiosis are increasingly being associated with the incidence and severity of asthma [73]. In sub-Saharan Africa, gut dysbiosis is more prevalent in urban populations than rural ones. Similarly, asthma is more prevalent in urban populations than rural populations. A positive dose–response association between consumption of fast foods three times or more in a week and asthma and allergies was found in urban areas of South Africa [74]. However, SARS-CoV-2 infectivity and COVID-19 are low in sub-Saharan Africa.
Conclusion
The correlation between the administration of the Bacille Calmette-Guérin (BCG) vaccine for tuberculosis and SARS-CoV-2 infection and COVID-19 severity is striking [75]. It is not known whether exposure to BCG directly reduces SARS-CoV-2 infectivity or not, however several studies have shown relatively low SARS-CoV-2 infections in countries that implemented universal BCG vaccinations (including most sub-Saharan African countries, Japan, India, Afghanistan, Nepal, Bhutan, Bangladesh, Israel, and others) and high infections and mortalities in countries that have BCG immunization programs only for high-risk communities (USA, Canada, Italy, Spain, France, Germany, South Korea, Iran) [75]. However, American Indians, Alaskan natives, India, and Brazil’s BCG adherence does not seem to support the high SARS-CoV-2 infection and COVID-19 mortality rate in these communities [75] [https://www.worldometers.info/coronavirus/]. The prevalence of co-morbidities including diabetes and obesity in American Indians and Alaskan natives, diabetes in India, and obesity and diabetes in Brazil may explain the inefficacy of the BCG vaccine against these SARS-CoV-2 co-morbidities in these populations. Others have suggested that the nonspecific effect of the BCG vaccine over the immune system is unknown and needs to be tested appropriately [76]. COVID-19 can have serious post-disease side effects, which underline the benefits of preventive efforts over aggressive management of the disease. Plant-based diet rich in inhibitors of SARS-CoV-2 infection may reduce the severity of infection and be a bridge to an effective vaccine. The strength of the hypothesis put herein includes the association between plant-based diet and reduced dysbiosis and the young age in the region, which together can help reduce infectivity in the event of contact with populations from regions with high infection rate. However, this hypothesis needs to be reported and then tested through well designed clinical studies and biochemical analyses to determine the potential correlation between youth and plant protease inhibitor-rich diet and the low SARS-CoV-2 infectivity and COVID-19 in sub-Saharan Africa. The landscape of COVID-19 is evolving very fast to the point that current efforts are merely and mostly invested into managing clinical outcomes. Later, during the post-COVID-19 vaccine era, there will be a need in due course, for reflection for consolidating food science and nutrition potential contributions to the prevention and management of SARS-CoV-2 infection and COVID-19. The COVID-19 pandemic is a public health concern that affects and requires the contribution of food science and nutrition to this medical problem. The landscape of the disease risk factors underscores the importance of bringing food scientists and nutritionists onboard with the medical community to find effective solutions for preventing and managing the pandemic.
This hypothesis has limitations. The data analyzed were from publicly available literature database including PubMed. The hypothesis assumes homogenous rural and urban populations and behavior across the sub-Saharan African region throughout the pandemic state although each country has socio-economic characteristics which may not equally overlap with other countries in the region. The discovery of new variants of the virus in the United Kingdom, South Africa, and Brazil which has already been detected in Canada, the US, and South Korea may bring about different response from the populations that have been resistant to the first wave of the virus. However, continuous use of social distancing, hand washings with soap, face masks, and plant-based rich diet will help prevent and reduce the spread of the infection.
Abbreviations
- SARS-CoV-2:
-
Severe acute respiratory syndrome coronavirus 2
- COVID-19:
-
Coronavirus -19
- ACE2:
-
Angiotensin converting enzyme-2
- ARDS:
-
Acute respiratory distress syndrome
- BCG:
-
Bacille Calmette-Guerin
- BMI:
-
Body mass index
- CD147:
-
Cluster of differentiation 147
- DM:
-
Death per million
- DPP4:
-
Dipeptidyl peptidase 4
- IFN-:
-
Interferon
- IL-:
-
Interleukin
- mTOR:
-
Mechanistic target of rapamycin
- NF-κB:
-
Nuclear factor-kappa B
- PAI-1:
-
Plasminogen activator inhibitor-1
- RAAS:
-
Renin–angiotensin–aldosterone system
- SCFA:
-
Short chain fatty acid
- TC:
-
Total cases
- TD:
-
Total deaths
- TLR4:
-
Toll-like receptor 4
- TMPRSS2:
-
Transmembrane protease serine 2
- TNF-α:
-
Tumor necrosis factor-alpha
References
Nordling L (2020) Africa’s pandemic puzzle: why so few cases and deaths? Science 369:756–757
Mbow M, Lell B, Jochems SP et al (2020) COVID-19 in Africa: dampening the storm? Science 369:624–626
Njenga MK, Dawa J, Nanyingi M et al (2020) Why is there low morbidity and mortality of COVID-19 in Africa? Am J Trop Med Hyg 103:564–569
Maeda JM, Nkengasong JN (2021) The puzzle of the COVID-19 pandemic in Africa. Science 371:27–28
Quaresima V, Naldini MM, Cirillo DM (2020) The prospects for the SARS-CoV-2 pandemic in Africa. EMBO Mol Med 12:e12488
Hardy ÉJL, Flori P (2021) Epidemiological specificities of COVID-19 in Africa: current or future public health concern? Ann Pharm Fr 79:216–226
Musa HH, Musa TH, Musa IH et al (2020) Addressing Africa’s pandemic puzzle: perspectives on COVID-19 transmission and mortality in sub-Saharan Africa. Int J Infect Dis 102:483–488
Lukassen S, Chua RL, Trefzer T et al (2020) SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. Embo j 39:e105114
Sardu C, Gambardella J, Morelli MB et al (2020) Hypertension, thrombosis, kidney failure, and diabetes: is COVID-19 an endothelial disease? A comprehensive evaluation of clinical and basic evidence. J Clin Med 9:1417
Tanne JH (2020) Covid-19: FDA panel votes to approve Pfizer BioNTech vaccine. BMJ 371:m4799
Iwasaki A, Grubaugh ND (2020) Why does Japan have so few cases of COVID-19? EMBO Mol Med 12:e12481
Volpato S, Landi F, Incalzi RA (2020) A frail health care system for an old population: lesson form the COVID-19 outbreak in Italy. J Gerontol A Biol Sci Med Sci 75:e126–e127
Diop BZ, Ngom M, Pougué Biyong C, Pougué Biyong JN (2020) The relatively young and rural population may limit the spread and severity of COVID-19 in Africa: a modelling study. BMJ Glob Health 5:e002699. https://doi.org/10.1136/bmjgh-2020-002699
O’Keefe SJD (2019) Plant-based foods and the microbiome in the preservation of health and prevention of disease. Am J Clin Nutr 110:265–266
Srikanth S, Chen Z (2016) Plant protease inhibitors in therapeutics-focus on cancer therapy. Front Pharmacol 7:470
Otlewski J, Krowarsch D (1996) Squash inhibitor family of serine proteinases. Acta Biochim Pol 43:431–444
Akiyama T, Ishida J, Nakagawa S et al (1987) Genistein, a specific inhibitor of tyrosine-specific protein kinases. J Biol Chem 262:5592–5595
Zhou CK, Young D, Yeboah ED et al (2017) TMPRSS2:ERG gene fusions in prostate cancer of West African men and a meta-analysis of racial differences. Am J Epidemiol 186:1352–1361
Rao NM (1991) Protease inhibitors from ripened and unripened bananas. Biochem Int 24:13–22
Rao NM, Prasad KVSRG, Pai KSR (1999) Angiotensin converting enzyme inhibitors from ripened and unripened bananas. Curr Sci 76:86–88. https://www.jstor.org/stable/24101334
Tian DD, Xu XQ, Peng Q et al (2020) Effects of banana powder (Musa acuminata Colla) on the composition of human fecal microbiota and metabolic output using in vitro fermentation. J Food Sci 85:2554–2564
Yeh KW, Chen JC, Lin MI et al (1997) Functional activity of sporamin from sweet potato (Ipomoea batatas Lam.): a tuber storage protein with trypsin inhibitory activity. Plant Mol Biol 33:565–570
Tian Q, Konczak I, Schwartz SJ (2005) Probing anthocyanin profiles in purple sweet potato cell line (Ipomoea batatas L. Cv. Ayamurasaki) by high-performance liquid chromatography and electrospray ionization tandem mass spectrometry. J Agric Food Chem 53:6503–6509
Hou WC, Liu JS, Chen HJ et al (1999) Dioscorin, the major tuber storage protein of yam (Dioscorea batatas Decne) with carbonic anhydrase and trypsin inhibitor activities. J Agric Food Chem 47:2168–2172
Faustina Dufie WM, Oduro I, Ellis WO et al (2013) Potential health benefits of water yam (Dioscorea alata). Food Funct 4:1496–1501
Kennedy AR (1998) The Bowman-Birk inhibitor from soybeans as an anticarcinogenic agent. Am J Clin Nutr 68:1406s–1412s
Ku SK, Yoon EK, Lee W et al (2016) Antithrombotic and antiplatelet activities of pelargonidin in vivo and in vitro. Arch Pharm Res 39:398–408
Mojica L, Berhow M, Gonzalez de Mejia E (2017) Black bean anthocyanin-rich extracts as food colorants: physicochemical stability and antidiabetes potential. Food Chem 229:628–639
Hill AJ, Peikin SR, Ryan CA, Blundell JE (1990) Oral administration of proteinase inhibitor II from potatoes reduces energy intake in man. Physiol Behav 48:241–246
Bawadi HA, Antunes TM, Shih F, Losso JN (2004) In vitro inhibition of the activation of Pro-matrix metalloproteinase 1 (Pro-MMP-1) and Pro-matrix metalloproteinase 9 (Pro-MMP-9) by rice and soybean Bowman-Birk inhibitors. J Agric Food Chem 52:4730–4736
Ferreira C, Viana SD, Reis F (2020) Gut microbiota dysbiosis immune hyperresponse-inflammation triad in coronavirus disease 2019 (COVID-19): impact of pharmacological and nutraceutical approaches. Microorganisms 8:1514
Redinbo MR (2014) The microbiota, chemical symbiosis, and human disease. J Mol Biol 426:3877–3891
David LA, Maurice CF, Carmody RN et al (2014) Diet rapidly and reproducibly alters the human gut microbiome. Nature 505:559–563
Samuelson DR, Welsh DA, Shellito JE (2015) Regulation of lung immunity and host defense by the intestinal microbiota. Front Microbiol 6:1085
Fan J, Li X, Gao Y et al (2020) The lung tissue microbiota features of 20 deceased patients with COVID-19. J Infect 81:e64–e67
De Filippo C, Cavalieri D, Di Paola M et al (2010) Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA 107:14691–14696
Gomez A, Petrzelkova KJ, Burns MB et al (2016) Gut microbiome of coexisting BaAka Pygmies and Bantu reflects gradients of traditional subsistence patterns. Cell Rep 14:2142–2153
Precup G, Vodnar DC (2019) Gut Prevotella as a possible biomarker of diet and its eubiotic versus dysbiotic roles: a comprehensive literature review. Br J Nutr 122:131–140
Vipperla K, O’Keefe SJ (2016) Diet, microbiota, and dysbiosis: a “recipe” for colorectal cancer. Food Funct 7:1731–1740
De Filippo C, Di Paola M, Ramazzotti M et al (2017) Diet, environments, and gut microbiota. A preliminary investigation in children living in rural and urban Burkina Faso and Italy. Front Microbiol 8:1979
Holmes MD, Dalal S, Sewram V et al (2018) Consumption of processed food dietary patterns in four African populations. Public Health Nutr 21:1529–1537
Mackworth-Young CR, Chingono R, Mavodza C et al (2021) Community perspectives on the COVID-19 response. Zimbabwe Bull World Health Organ 99:85–91
Kollamparambil U, Oyenubi A (2021) Behavioural response to the Covid-19 pandemic in South Africa. PLoS One 16:e0250269
Adebowale OO, Adenubi OT, Adesokan HK et al (2021) SARS-CoV-2 (COVID-19 pandemic) in Nigeria: multi-institutional survey of knowledge, practices and perception amongst undergraduate veterinary medical students. PLoS One 16:e0248189
Pinchoff J, Kraus-Perrotta C, Austrian K et al (2021) Mobility patterns during COVID-19 travel restrictions in Nairobi urban informal settlements: who is leaving home and why. J Urban Health 98:211–221
Yang J, Hu J, Zhu C (2020) Obesity aggravates COVID-19: a systematic review and meta-analysis. J Med Virol 93:257–261
Caci G, Albini A, Malerba M et al (2020) COVID-19 and obesity: dangerous liaisons. J Clin Med 9:2511
Cava E, Neri B, Carbonelli MG et al (2021) Obesity pandemic during COVID-19 outbreak: narrative review and future considerations. Clin Nutr 40:1637–1643
Demasi M (2021) COVID-19 and metabolic syndrome: could diet be the key? BMJ Evid Based Med 26:1–2
van Zelst CM, Janssen ML, Pouw N et al (2020) Analyses of abdominal adiposity and metabolic syndrome as risk factors for respiratory distress in COVID-19. BMJ Open Respir Res 7:e000792
Zhang W, Zhao Y, Zhang F et al (2020) The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): the perspectives of clinical immunologists from China. Clin Immunol 214:108393
Al-Benna S (2020) Association of high level gene expression of ACE2 in adipose tissue with mortality of COVID-19 infection in obese patients. Obes Med 19:100283
Kwok S, Adam S, Ho JH et al (2020) Obesity: a critical risk factor in the COVID-19 pandemic. Clin Obes 10:e12403
Biadgilign S, Mgutshini T, Haile D et al (2017) Epidemiology of obesity and overweight in sub-Saharan Africa: a protocol for a systematic review and meta-analysis. BMJ Open 7:e017666
Souza SA, Silva AB, Cavalcante UMB et al (2018) Adult obesity in different countries: an analysis via beta regression models. Cad Saude Publica 34:e00161417
NCD-RisC - Africa Working Group (2017) Trends in obesity and diabetes across Africa from 1980 to 2014: an analysis of pooled population-based studies. Int J Epidemiol 46:1421–1432
NCD-RisC (2016) Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1698 population-based measurement studies with 19·2 million participants. Lancet 387:1377–1396
Otang-Mbeng W, Otunola GA, Afolayan AJ (2017) Lifestyle factors and co-morbidities associated with obesity and overweight in Nkonkobe municipality of the Eastern Cape. J Health Popul Nutr 36:22
Erener S (2020) Diabetes, infection risk and COVID-19. Mol Metab 39:101044
Yang JK, Lin SS, Ji XJ, Guo LM (2010) Binding of SARS coronavirus to its receptor damages islets and causes acute diabetes. Acta Diabetol 47:193–199
Codo AC, Davanzo GG, Monteiro LB et al (2020) Elevated glucose levels favor SARS-CoV-2 infection and monocyte response through a HIF-1α/glycolysis-dependent axis. Cell Metab 32:437–446.e435
Werfalli M, Engel ME, Musekiwa A et al (2016) The prevalence of type 2 diabetes among older people in Africa: a systematic review. Lancet Diabetes Endocrinol 4:72–84
Mapa-Tassou C, Katte JC, Mba Maadjhou C, Mbanya JC (2019) economic impact of diabetes in Africa. Curr Diab Rep 19:5
Subramani SK, Yadav D, Mishra M et al (2019) Prevalence of type 2 diabetes and prediabetes in the Gwalior-Chambal region of Central India. Int J Environ Res Public Health 16:4708
Zheng Z, Peng F, Xu B et al (2020) Risk factors of critical & mortal COVID-19 cases: a systematic literature review and meta-analysis. J Infect 81:e16–e25
Matsuyama T, Kubli SP, Yoshinaga SK et al (2020) An aberrant STAT pathway is central to COVID-19. Cell Death Differ 27:3209–3225
Kietzmann T, Roth U, Jungermann K (1999) Induction of the plasminogen activator inhibitor-1 gene expression by mild hypoxia via a hypoxia response element binding the hypoxia-inducible factor-1 in rat hepatocytes. Blood 94:4177–4185
Keates AK, Mocumbi AO, Ntsekhe M et al (2017) Cardiovascular disease in Africa: epidemiological profile and challenges. Nat Rev Cardiol 14:273–293
Wong ND, Moran A, Narula J (2018) Hypertension control in Africa: a call to action. Glob Heart 13:1–2
Chakafana G, Mutithu D, Hoevelmann J et al (2020) Interplay of COVID-19 and cardiovascular diseases in Africa: an observational snapshot. Clin Res Cardiol 109:1460–1468
Hosoki K, Chakraborty A, Sur S (2020) Molecular mechanisms and epidemiology of COVID-19 from an allergist’s perspective. J Allergy Clin Immunol 146:285–299
Kwizera R, Musaazi J, Meya DB et al (2019) Burden of fungal asthma in Africa: a systematic review and meta-analysis. PLoS One 14:e0216568
Sokolowska M, Frei R, Lunjani N et al (2018) Microbiome and asthma. Asthma Res Pract 4:1
Nkosi V, Rathogwa-Takalani F, Voyi K (2020) The frequency of fast food consumption in relation to wheeze and asthma among adolescents in Gauteng and North West provinces, South Africa. Int J Environ Res Public Health 17:1994
Sharma AR, Batra G, Kumar M et al (2020) BCG as a game-changer to prevent the infection and severity of COVID-19 pandemic? Allergol Immunopathol (Madr) 48:507–517
Moorlag S, Arts RJW, van Crevel R, Netea MG (2019) Non-specific effects of BCG vaccine on viral infections. Clin Microbiol Infect 25:1473–1478
Acknowledgements
The authors acknowledge Robert M. Levine, MD, Henry Ford Medical Center, Dearborn, MI for reviewing the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Losso, J.N., Losso, M.N., Toc, M. et al. The Young Age and Plant-Based Diet Hypothesis for Low SARS-CoV-2 Infection and COVID-19 Pandemic in Sub-Saharan Africa. Plant Foods Hum Nutr 76, 270–280 (2021). https://doi.org/10.1007/s11130-021-00907-6
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11130-021-00907-6