
Abstract
The evolution of photosynthesis from primitive photosynthetic bacteria to higher plants has been driven by the need to adapt to a wide range of environmental conditions. The red alga Cyanidioschyzon merolae is a primitive organism, which is capable of performing photosynthesis in extreme acidic and hot environments. The study of its photosynthetic machinery may provide new insight on the evolutionary path of photosynthesis and on light harvesting and its regulation in eukaryotes. With that aim, the structural and functional properties of the PSI complex were investigated by biochemical characterization, mass spectrometry, and X-ray crystallography. PSI was purified from cells grown at 25 and 42 °C, crystallized and its crystal structure was solved at 4 Å resolution. The structure of C. merolae reveals a core complex with a crescent-shaped structure, formed by antenna proteins. In addition, the structural model shows the position of PsaO and PsaM. PsaG and PsaH are present in plant complex and are missing from the C. merolae model as expected. This paper sheds new light onto the evolution of photosynthesis, which gives a strong indication for the chimerical properties of red algae PSI. The subunit composition of the PSI core from C. merolae and its associated light-harvesting antennae suggests that it is an evolutionary and functional intermediate between cyanobacteria and plants.







Similar content being viewed by others

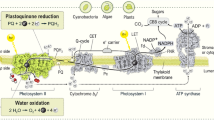
Abbreviations
- PSI:
-
Photosystem I
- PSII:
-
Photosystem II
- LHC:
-
Light-harvesting complex
References
Adams PD et al (2010) PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr D 66:213–221
Ago H, Adachi H, Umena Y et al (2016) Novel features of eukaryotic photosystem II revealed by its crystal structure analysis from a red alga. J Biol Chem 291:5676–5687
Alboresi A, Le Quiniou C, Yadav SK, Scholz M, Meneghesso A, Gerotto C, Simionato D, Hippler M, Boekema EJ, Croce R, Morosinotto T (2017) Conservation of core complex subunits shaped the structure and function of photosystem I in the secondary endosymbiont alga Nannochloropsis gaditana. New Phytol 213, 714–726
Allen MB (1959) Studies with Cyanidium caldarium, an anomalously pigmented chlorophyte. Arch Mikrobiol 32:270–277
Amunts A, Drory O, Nelson N (2007) The structure of a plant photosystem I supercomplex at 3.4 A resolution. Nature 447:58–63
Amunts A, Toporik H, Borovikova A, Nelson N (2010) Structure determination and improved model of plant photosystem I. J Biol Chem 285:3478–3486
Barber J (2004) Engine of life and big bang of evolution: a personal perspective. Photosynth Res 80:137–155
Bengis C, Nelson N (1977) Subunit structure of chloroplast photosystem I reaction center. J Biol Chem 252:4564–4569
Ben-Shem A, Frolow F, Nelson N (2003) Crystal structure of plant photosystem I. Nature 426:630–635
Ben-Shem A, Frolow F, Nelson N (2004) Evolution of photosystem I—from symmetry through pseudosymmetry to asymmetry. FEBS Lett 564:274–280
Busch A, Hippler M (2011) The structure and function of eukaryotic photosystem I. Biochim Biophys Acta 1807:864–877
Busch A, Nield J, Hippler M (2010) The composition and structure of photosystem I-associated antenna from Cyanidioschyzon merolae. Plant J 62:886–897
Chan CX, Bhattacharya D (2013) Analysis of horizontal genetic transfer in red algae in the post-genomics age. Mob Genet Elem 3:e27669
Chitnis PR, Purvis D, Nelson N (1991) Molecular cloning and targeted mutagenesis of the gene psaF encoding subunit III of photosystem I from the cyanobacterium Synechocystis sp. PCC 6803. J Biol Chem 266:20146–20151
Croce R, van Amerongen H (2013) Light-harvesting in photosystem I. Photosynth Res 116:153–166
DeLano WL (2002) Pymol: an open-source molecular graphics tool. Scientific, San Carlos
Drop B, Webber-Birungi M, Fusetti F, Kouril R, Redding KE, Boekema EJ, Croce R (2011) Photosystem I of Chlamydomonas reinhardtii contains nine light-harvesting complexes (Lhca) located on one side of the core. J Biol Chem 286:44878–44887
Drop B, Yadav KNS, Boekema EJ, Croce R (2014) Consequences of state transitions on the structural and functional organization of photosystem I in the green alga Chlamydomonas reinhardtii. Plant J 78:181–191
Elrad D, Grossman AR (2004) A genome’s-eye view of the light-harvesting polypeptides of Chlamydomonas reinhardtii. Curr Genet 45:61–75
Emsley P, Cowtan K, Coot (2004) Model-building tools for molecular graphics. Acta Crystallogr Sect D 60:2126–2132
Germano M, Yakushevska AE, Keegstra W, van Gorkom HJ, Dekker JP, Boekema EJ (2002) Supramolecular organization of photosystem I and light-harvesting complex I in Chlamydomonas reinhardtii. FEBS Lett 525:121–125
Haniewicz P, Abram M, Nosek L, Kirkpatrick J, El-Mohsnawy E, Janna Olmos JD, Kouril R, Kargul JM. (2017) Molecular mechanisms of photoadaptation of photosystem I supercomplex of in an evolutionary cyanobacterial/algal intermediate. Plant Physiol 01022
Hippler M, Reichert J, Sutter M et al (1996) The plastocyanin binding domain of photosystem I. EMBO J 15:6374–6384
Hippler M, Drepper F, Rochaix JD, Mühlenhoff U (1999) Insertion of the N-terminal part of PsaF from Chlamydomonas reinhardtii into photosystem I from Synechococcus elongatus enables efficient binding of algal plastocyanin and cytochrome c6. J Biol Chem 274:4180–4188
Jensen PE et al (2007) Structure, function and regulation of plant photosystem I. Biochim Biophys Acta 1767:335–352
Jordan P, Fromme P, Witt HT, Klukas O, Saenger W, Krauss N (2001) Three-dimensional structure of cyanobacterial photosystem I at 2.5 A resolution. Nature 411:909–917
Kabsch W (2010) Xds. Acta Crystallogr Sect D 66:125–132
Kargul J, Nield J, Barber J (2003) Three-dimensional reconstruction of a light-harvesting complex I-photosystem I (LHCI-PSI) supercomplex from the green alga Chlamydomonas reinhardtii – insights into light harvesting for PSI. J Biol Chem 278:16135–16141
Kargul J, Janna Olmos JD, Krupnik T (2012) Structure and function of photosystem I and its application in biomimetic solar-to-fuel systems. J Plant Physiol 169:1639–1653
Keeling PJ (2004) Diversity and evolutionary history of plastids and their hosts. Am J Bot 91:1481–1493
Keeling PJ, Palmer JD (2008) Horizontal gene transfer in eukaryotic evolution. Nat Rev Genet 9:605–618
Kim E, Graham LE (2008) EEF2 analysis challenges the monophyly of archaeplastida and chromalveolata. PLoS ONE 3(7):e2621
Liguori N, Roy LM, Opacic M, Durand G, Croce R (2013) Regulation of light harvesting in the green alga Chlamydomonas reinhardtii: the C-terminus of LHCSR is the knob of a dimmer switch. J Am Chem Soc 135:18339–18342
Malavath T, Caspy I, Netzer-El SY et al (2018) Structure and function of wild-type and subunit-depleted photosystem I in Synechocystis. Biochim Biophys Acta 0–1
Matsuzaki M et al (2004) Genome sequence of the ultrasmall unicellular red alga Cyanidioschyzon merolae 10D. Nature 428:653–657
Mazor Y, Nataf D, Toporik H, Nelson N (2014) Crystal structures of virus-like photosystem I complexes from the mesophilic cyanobacterium Synechocystis PCC 6803. Elife 3:e01496
Mazor Y, Borovikova A, Nelson N (2015) The structure of plant photosystem i super-complex at 2.8 Å resolution. Elife 4:1–18
Mazor Y, Borovikova A, Caspy I, Nelson N (2017a) Structure of the plant photosystem i supercomplex at 2.6 Å resolution. Nat Plants 3:1–9
Mazor Y, Borovikova A, Caspy I, Nelson N (2017b) Structure of the plant photosystem I supercomplex at 2.6Å resolution. Nat Plants 3:17014
Merchant SS, Prochnik SE, Vallon O et al (2007) The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 318:245–250
Michel H, Ostermeier C Crystallization of membrane proteins. Biophys Methods 697–700 (1997)
Minoda A, Sakagami R, Yagisawa F, Kuroiwa T, Tanaka K (2004) Improvement of culture conditions and evidence for nuclear transformation by homologous recombination in a red alga, Cyanidioschyzon merolae 10D. Plant Cell Physiol 45:667–671
Moreira D, Guyader HL, Phillippe H (2000) The origin of red algae and the evolution of chloroplasts. Nature 405:69–72
Naithani S, Hou JM, Chitnis PR (2000) Targeted inactivation of the psaK1, psaK2 and psaM genes encoding subunits of photosystem I in the cyanobacterium Synechocystis sp. PCC 6803. Photosynth Res 63:225–236
Nechushtai R, Nelson N (1981) Purification properties and biogenesis of Chlamydomonas reinhardii photosystem I reaction center. J Biol Chem 256:11624–11628
Nelson N (2011) Photosystems and global effects of oxygenic photosynthesis. Biochim Biophys Acta 1807:856–863
Nelson N, Ben-Shem A (2004) The complex architecture of oxygenic photosynthesis. Nat Rev Mol Cell Biol 5:971–982
Nelson N, Junge W (2015) Structure and energy transfer in photosystems of oxygenic photosynthesis. Annu Rev Biochem 84:659–683
Nikolova D, Weber D, Scholz M, Bald T, Scharsack JP, Hippler M (2017) Temperature-induced remodeling of the photosynthetic machinery tunes photosynthesis in the thermophilic alga Cyanidioschyzon merolae. Plant Physiol 174:35–46
Nozaki H, Matsuzaki M, Takahara M et al (2003) The phylogenetic position of red algae revealed by multiple nuclear genes from mitochondria-containing eukaryotes and an alternative hypothesis on the origin of plastids. J Mol Evol 56:485–497
Otwinowski Z, Minor W (1997) Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol 276:307–326
Qin X, Suga M, Kuang T, Shen JR (2015) Structural basis for energy transfer pathways in the plant PSI-LHCI supercomplex. Science 348:989–995
Reeb V, Bhattacharya D (2010) The thermo-acidophilic Cyanidiophyceae (Cyanidiales). In: Seckbach J, Chapman DJ (eds) Red algae in the genomic age. Springer Netherlands, Dordrecht, pp 409–426
Sharon I, Alperovitch A, Rohwer F, Haynes M, Glaser F, Atamaa-Ismaeel N, Pinter RY, Partensky F, Koonin EV, Wolf YI, Nelson N, Oded Béjà O (2009) Photosystem I gene cassettes are present in marine virus genomes. Nature 461:258–262
Sommer F, Drepper F, Haehnell W, Hippler M (2004) The hydrophobic recognition site formed by residues PsaA-Trp651 and PsaB-Trp627 of photosystem I in Chlamydomonas reinhardtii confers distinct selectivity for binding of plastocyanin and cytochrome c6. J Biol Chem 279:20009–20017
Stauber EJ, Busch A, Naumann B, Svatos A, Hippler M (2009) Proteotypic profiling of LHCI from Chlamydomonas reinhardtii provides new insights into structure and function of the complex. Proteomics 9:398–408
Strong M, Sawaya MR, Wang S et al (2006) Toward the structural genomics of complexes: crystal structure of a PE/PPE protein complex from Mycobacterium tuberculosis. Proc Natl Acad Sci USA 103:8060–8065
Tan S, Ducret a, Aebersold R, Gantt E (1997) Red algal LHC I genes have similarities with both Chl a/b- and a/c-binding proteins: a 21 kDa polypeptide encoded by LhcaR2 is one of the six LHC I polypeptides. Photosynth Res 53:129–140
Thangaraj B, Jolley CC, Sarrou I et al (2011) Efficient light harvesting in a dark, hot, acidic environment: the structure and function of PSI-LHCI from Galdieria sulphuraria. Biophys J 100:135–143
Tian L, Liu Z, Wang F, Shen L, Chen J, Chang L, Zhao S, Han G, Wang W, Kuang T, Qin X, Shen JR (2017) Isolation and characterization of PSI-LHCI super-complex and their sub-complexes from a red alga Cyanidioschyzon merolae. Photosynth Res 133:201–214
Umena Y, Kawakami K, Shen J-R, Kamiya N (2011) Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 A ̊. Nature 473:55–60
Vagin AA et al (2004) REFMAC5 dictionary: organization of prior chemical knowledge and guidelines for its use. Acta Crystallogr Sect D 60:2184–2195
Vaguine AA, Richelle J, Wodak SJ (1999) SFCHECK: a unified set of procedures for evaluating the quality of macromolecular structure-factor data and their agreement with the atomic model. Acta Crystallogr Sect D 55:191–205
Vanselow C, Weber APM, Krause K, Fromme P (2009) Genetic analysis of the photosystem I subunits from the red alga, Galdieria sulphuraria. Biochim Biophys Acta 1787:46
Zhang H, Kurisu G, Smith JL, Cramer WA (2003) A defined protein-detergent-lipid complex for crystallization of integral membrane proteins: the cytochrome b6f complex of oxygenic photosynthesis. Proc Natl Acad Sci USA 100, 5160–5163
Acknowledgements
The authors would like to thank the ESRF, SLS, and BESSYII synchrotrons for beam time and the staff scientists for excellent guidance and assistance. This work was supported by a grant (No. 293579 – HOPSEP) from the European Research Council, by The Israel Science Foundation (Grant No. 569/17), and by the I-CORE Program of the Planning and Budgeting Committee and The Israel Science Foundation (Grant No. 1775/12). M.H acknowledges funding by the German Science Foundation (DFG HI 739/13.1).
Author information
Authors and Affiliations
Corresponding author
Additional information
Maya Antoshvili and Ido Caspy equally contributed.
Rights and permissions
About this article

Cite this article
Antoshvili, M., Caspy, I., Hippler, M. et al. Structure and function of photosystem I in Cyanidioschyzon merolae. Photosynth Res 139, 499–508 (2019). https://doi.org/10.1007/s11120-018-0501-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11120-018-0501-4