
Abstract
We analyzed the contribution of genetic factors on the association between puberty timing and body mass index (BMI) using longitudinal data and two approaches: (i) genetic twin design and (ii) polygenic scores (PGS) of obesity indices. Our data were derived from Finnish cohorts: 9080 twins had information on puberty timing and BMI and 2468 twins also had genetic data. Early puberty timing was moderately associated with higher BMI in childhood in both boys and girls; in adulthood these correlations were weaker and largely disappeared after adjusting for childhood BMI. The largest proportion of these correlations was attributable to genetic factors. The higher PGSs of BMI and waist circumference were associated with earlier timing of puberty in girls, whereas weaker associations were found in boys. Early puberty is not an independent risk factor for adult obesity but rather reflects the association between puberty timing and childhood BMI contributed by genetic predisposition.
Similar content being viewed by others


Introduction
The timing of puberty is influenced by both genetic and environmental factors. Previous twin studies have shown that from 50% to more than 90% of individual variation of puberty timing is accounted for by inter-individual genetic differences using menarche age in girls (Anderson et al. 2007) and voice break (Hur 2020) and pubertal growth spurt in boys (Silventoinen et al. 2008) as puberty indicators. Further, the role of genetic factors is supported by the associations of parental pubertal timing with different indicators of puberty in offspring (Busch et al. 2020). The importance of genetic factors is also shown in genome-wide-association (GWA) studies on age of menarche in girls (Day et al. 2017; Sarnowski et al. 2021) and voice break in boys (Day et al. 2015). However, there is clear evidence that the timing of puberty has gotten earlier in girls at the same time when general nutrition has improved, showing that factors other than genetic ones are also involved (Papadimitriou 2016; Eckert-Lind et al. 2020). In boys, the evidence is less clear, but a likely reason is that measuring puberty is more difficult in boys than in girls, thus historical data are sparse (Euling et al. 2008).
The secular trend toward the earlier onset of puberty has raised public health concerns because early puberty is associated with an increased risk of metabolic disorders such as type 2 diabetes (Elks et al. 2013) and increased levels of several cardiometabolic risk factors (Widén et al. 2012) as well as an increased breast cancer risk (Collaborative Group on Hormonal Factors in Breast Cancer 2012). However, there is clear evidence that higher childhood body mass index (BMI) and other obesity indices are associated with earlier timing of puberty. The evidence is strongest for girls (Kaplowitz 2008), but there are studies suggesting that high BMI is associated with earlier pubertal timing in boys as well (Busch et al. 2019; Hur 2020; Oehme et al. 2021; Silventoinen et al. 2008). Thus, the associations between puberty timing and cardio-metabolic health in adulthood can also reflect childhood BMI, which is associated with adult BMI leading to metabolic disorders (Lloyd et al. 2012).
Results based on both twin (Silventoinen et al. 20162017) and GWA studies (Locke et al. 2015) have shown that genetic factors importantly affect the variation of BMI. The genetic variants associated with BMI are found to be expressed in the brain tissue in particular (Turcot et al. 2018), suggesting the role of behavioral factors as mediators from genes to BMI. Thus, genetic factors can be important when explaining the association between puberty timing and BMI. This is supported by twin studies which have found genetic correlations between BMI and age at voice break (− 0.18) (Hur 2020) and pubertal growth spurt in boys (− 0.16) (Silventoinen et al. 2008). Further, early GWA studies already identified common loci between pubertal timing and BMI in males and females (Cousminer et al. 20132014). In more recent studies, the genetic correlation of BMI based on all common genetic variants in the UK Biobank was found to be -0.33 with age at menarche in girls and -0.35 with age at voice break in boys (UK Biobank 2021). A similar genetic correlation between childhood BMI and age at menarche (− 0.42) was found in a large GWA meta-analysis of European-origin populations (Vogelezang et al. 2020).
In this study, we aimed to analyze how the timing of puberty is associated with childhood BMI and BMI in adulthood using longitudinal twin datasets having puberty measures for both males and females. Our specific research aims are (i) to estimate the genetic correlations between puberty indices and BMI measured in adolescence and adulthood using a classical genetic twin design, (ii) to analyze whether the timing of puberty is associated with adult BMI when adjusted for BMI in adolescence, and (iii) to analyze how the genetic liability to obesity measured as polygenic scores (PGS) is associated with the timing of puberty in adolescence. The strength of the data is that the two analytical approaches—the classical twin design utilizing the genetic similarity of twins and the PGS design utilizing measured information on genetic polymorphisms through the whole genome—make different theoretical assumptions thus complementing each other. Briefly, the classical twin design is based on the different genetic relatedness of monozygotic (MZ) and dizygotic (DZ) twins, whereas the PGS design is based on observed associations between genetic polymorphisms and the trait under study (Friedman et al. 2021). Both approaches allows us to calculate shared genetic variation between the timing of puberty and BMI.
Data and methods
Study participants
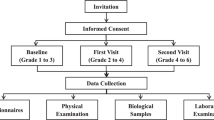
The data were derived from the FinnTwin12 and FinnTwin16 studies, two independent longitudinal cohort studies with the target population of all Finnish twins born in 1983–1987 and 1974–1979, respectively. In the FinnTwin12 study, the baseline questionnaire was sent to 6272 twin individuals at 11–12 years of age and 5184 responded. Three follow-up questionnaires were sent at 14, 17 and, on average 24 years of age (Rose et al. 2019). In the FinnTwin16 study, the baseline questionnaire was sent to 6430 twin individuals at 16 years of age and 5778 responded. Four follow-up questionnaires were sent at 17, 18, 25 and 35 years of age (Kaidesoja et al. 2019). The numbers of valid BMI measures at each age are available in Table 1. In both studies, a sub-cohort of twins were invited to laboratory examinations where they were asked to give a DNA sample (blood, buccal cells or saliva). DNA samples used in the molecular genetic analyses were available for 1438 twins in the FinnTwin12 study and 1030 twins in the FinnTwin16 study. Zygosity was based on measured genotypes for the twins we had a DNA sample. For the rest of the twins, zygosity was based on questions of physical similarity in the baseline questionnaire. This method was validated in the FinnTwin12 study using 395 same-sex twin pairs and in these twins 97% of questionnaire assignment of zygosity was confirmed by a DNA test showing high reliability (Jelenkovic et al. 2011).
Measures and genotyping
BMI (kg/m2) was calculated from self-reported values, which showed high reliability in these cohorts (Saarni et al. 2009). Because of the skewness of BMI distributions at all ages (skewness parameters 1.11–2.36), we log-transformed BMI, producing roughly normal distributions (skewness parameters 0.57–1.08). Sexual maturation in the FinnTwin12 study was measured at 12 and 14 years of age by using the Pubertal Development Scale (PDS), described in detail elsewhere (Dick et al. 2001). The PDS questionnaire includes five items on the timing of secondary sexual characteristics: body hair, growth spurt, skin change, voice change (boys)/breast change (girls), and facial hair (boys)/menarche (girls). The development stage was assessed in a 4-point scale ranging from 1 (no development) to 4 (complete development) except for menarche (1 = has not occurred and 4 = has occurred). At 12 years of age, the fourth response option was not used since only a few children were expected to have complete development. The mean of these ratings was then used as the indicator of puberty; we interpreted that advantaged maturation, i.e., larger PDS scores, indicates the earlier timing of puberty. In the FinnTwin16 study, pubertal age (PA) was asked retrospectively on the age of menarche in girls and voice break in boys at 17 years of age. For those who reported to have not yet experienced menarche/voice break, PA was coded as 17 years of age. BMI and puberty indices were adjusted for age at the time of survey separately in males and females to take into account the age differences between twin pairs within the waves.
The technical details of genotyping, imputation and PGS calculations have been described elsewhere (Kujala et al. 2020). The PGSs were derived using GWA summary statistics of BMI (Yengo et al. 2018), waist-to-hip ratio adjusted for BMI (WHR-BMI) (Pulit et al. 2019) and waist circumference (WC) (Broad Institute 2021). The total number of single nucleotide polymorphisms (SNP) used for the PGS calculations were 996,919 for BMI, 1,148,565 for WHR-BMI and 1,146,408 for WC. The number of individuals in the GWA studies were 692,578 for BMI, 484,563 for WHR-BMI and 419,807 for WC.
Statistical methods
We started the analyses using classical twin modeling based on the comparisons of similarity between MZ and DZ twins (Posthuma et al. 2003). MZ twins are virtually identical at the gene sequence level, whereas DZ twins share half of genetic variance as in ordinary siblings. Based on this principle, the trait variation can be decomposed to additive genetic variation (A) including all main effects of the loci affecting the trait (correlations of 1.0 within MZ and 0.5 within DZ pairs); dominance genetic variation (D) caused by interactions between alleles in the same locus (correlations of 1.0 within MZ and 0.25 within DZ pairs); shared environment (C) including the effect of all environmental factors that make twins in a pair similar to each other (correlations of 1.0 within both MZ and DZ twins); and unique environment (E) including environmental factors specific to each twin individual as well as any measurement error (correlations of 0 within both MZ and DZ twins). Because we have only twins reared together in our data, we were unable to estimate D and C effects simultaneously. The genetic twin models were fitted using the OpenMx package, version 3.0.2, of R statistical software (Neale et al. 2016).
We estimated the genetic and environmental correlations of PDS with BMI at different ages by using bivariate Cholesky decomposition, a method decomposing all variance and covariance in the data into uncorrelated factors. Further, we estimated these correlations between PDS at 12 and 14 years of age to analyze how much they reflect the same genetic factors. Thus, separate bivariate models were conducted at each age. Since puberty is later in boys than in girls and thus the puberty indices are not necessarily comparable between sexes, we used only same-sex pairs in these analyses. The number of complete pairs (MZ and same-sex DZ) in each bivariate model varied between 822 and 1475 for PDS (FinnTwin12 study) and between 930 and 978 for PA (FinnTwin16 study) (Supplementary Table 1). We then analyzed the associations of PDS and PA with BMI in adulthood after adjusting for the baseline BMI using the linear regression model. If independent associations between the puberty indices and post-puberty BMI existed, we decomposed the regression residuals to additive genetic and unique environmental components.
The heritability estimates of puberty indices and BMI have previously been reported in these study cohorts, suggesting the presence of dominance genetic factors for PA (Kaprio et al. 1995) and shared environment for PDS in females (Wehkalampi et al. 2008) and shared environment for BMI until 14 years of age in both males and females (Lajunen et al. 2009). At later ages, an additive genetic/unique environment (AE) model was the best fitting model for BMI (Lajunen et al. 2009; Ortega-Alonso et al. 2012). Thus, we selected the additive genetic/shared environment/unique environment (ACE) model as the starting point of genetic modeling. The model fit statistics are presented in Supplementary Table 1. We did not find evidence that there would be shared environmental correlations affecting the associations between PDS or PA and BMI. This correlation was only statistically significant at conventional level in the model for PDS and BMI at 12 years of age in males (p = 0.0171). However, this may be a spurious finding because of multiple testing. When we compared the AE models to the ACE models, for 6 models we found a statistically significant difference at the conventional level (p = 0.004–0.0400), but they were not statistically significant after the Bonferroni correction of multiple testing (significance level p = 0.002 for 24 tests). We decided to use the AE model in the statistical analyses; if the shared environmental variance component is estimated without simultaneously estimating the shared environmental correlation, it would have artificially increased the additive genetic correlation.
After these genetic twin models, we continued the analyses by studying how the PGSs of obesity indices were associated with the puberty indices. In these analyses, we used linear regression models with puberty indices as the dependent and PGS as an independent variable. The population stratification was adjusted for by including ten principal components of the genome into the model as independent variables. We calculated standardized β-coefficients presenting the change of 1 standard deviation (SD) of PDS or PA per the change of 1 SD of each PGS of obesity indices. The PGS analyses, as well as all statistical testing in the descriptive analyses, were performed using linear regression models by the Stata/SE 16.1 for Windows statistical software (StataCorp, College Station, TX, USA). The cluster option was used to correct the standard errors and confidence intervals (CI) for the lack of statistical independence of twins sampled as twin pairs (Williams 2000).
Results
Table 1 presents the descriptive statistics of our study cohort. Boys developed later than girls in their sexual maturation measured by PDS at 12 and 14 years of age or retrospectively asked PA at 17 years of age. Boys had slightly higher BMI than girls at 12 years of age. This difference disappeared by 14 years of age but emerged again at 17 years of age. The sex difference in BMI increased during adulthood and was largest at 35 years of age when men had 1.79 (95% CI 1.52–2.05) kg/m2 higher BMI than women.
We started the analyses by studying the contributions of genetic and environmental factors on the correlations between PDS, PA and BMI at different ages (Table 2). Separate bivariate Cholesky decomposition was used at each age to decompose the trait correlation into additive genetic correlation and unique environmental correlation. We also calculated the proportions of the trait correlations explained by additive genetic and unique environmental factors. A modest correlation was observed between PDSs at ages 12 and 14 (r = 0.36 in boys and 0.51 in girls), which was nearly totally explained by genetic factors: the genetic correlations showed that 24% of genetic variation in boys and 38% in girls affecting sexual maturation was shared between these ages. BMI was positively correlated with PDS at both ages: the correlations varied from 0.14 in boys at 12 years of age to 0.28 in girls at 14 years of age. Very similar correlations (-0.15 in boys and -0.28 in girls) were also found when studying PA and BMI at the age of 16. For the most part (84% or more), these correlations were explained by genetic factors. The genetic correlations showed that from 3 to 11% of genetic variation of adolescence BMI and puberty indices was explained by the same genetic factors. When we studied the correlations between the puberty indices and post-puberty BMI, they systematically decreased over age. However, these correlations were again mainly explained by shared genetic factors (64% or more).
Next, we analyzed how the puberty indices were associated with post-pubertal BMI after adjusting the results for baseline BMI (Table 3). However, only weak and mainly statistically non-significant associations were found. The only statistically significant associations were found between PA and BMI at 17 and 18 years of age in females, but these associations were also small in size (0.03 and 0.06 SD of BMI per 1 SD change in PA) and faded with later age. Thus, we did not further decompose these associations into genetic and environmental factors.
Finally we analyzed how the PGSs of different obesity indices were associated with puberty timing (Table 4). In girls, the high PGSs of both BMI and WC were associated with more advanced maturation measured by PDS both at age 12 and 14. However, at 12 years of age, this association was only marginally statistically significant for the PGS of WC. The strongest association was found at 14 years of age when a 1 SD increase of the PGS of WC increased the PDS by 0.13 SD. In boys, similar associations were found, but they were weaker than in girls and not statistically significant. When using retrospectively reported PA, the associations were weak and not statistically significant. We did not find systematic associations between the PGS of WHR-BMI and the puberty indices.
Discussion
In our study based on two independent longitudinal cohorts of twins, we found clear evidence that there is shared genetic overlap between the timing of puberty and BMI being the strongest in childhood but still existing in adulthood. When using classical twin modeling, the results were very systematic for males and females: the genetic correlations were moderate between puberty timing and childhood BMI and decreased when studying BMI in adulthood. When studying the associations between the PGSs of adiposity indices and puberty timing, the associations were stronger in females than in males. However, the confidence intervals were also wide and the results were not statistically significant for males. Thus, larger data sets are needed to analyze the question of sex differences in detail. However, in general, our results correspond well with previous twin studies (Hur 2020; Silventoinen et al. 2008) and GWA studies (Cousminer et al. 2013; Cousminer et al. 2014; UK Biobank 2021; Vogelezang et al. 2020) that also show shared genetic variation between the timing of puberty and BMI. Further, previous studies have also shown that the PGSs of puberty indices are associated with BMI in childhood and adulthood, providing further evidence on the shared genetic background (Bell et al. 2018; Gill et al. 2018). Our results confirm that a large proportion, more than 90% at most ages, of the correlations between the timing of puberty and BMI is due to common genetic factors. Together, these results provide strong evidence that the same genetic factors affecting early puberty and childhood adiposity explain the association between the timing of puberty and BMI in adulthood. However, it is noteworthy that at maximum only 11% of the genetic variation is shared between puberty timing and adolescence BMI, thus most of the genetic variation is specific for these two traits.
When studying the longitudinal associations, we found only a little evidence that the timing of puberty is associated with adult BMI independently of pre-pubertal BMI. We found that the early timing of puberty was associated in males and females with higher adult BMI, but the correlations were weaker than with childhood BMI and largely disappeared when adjusted for childhood BMI. These results are largely similar to those found previously in population-level studies, but it is noteworthy that only a few studies so far have studied these association in males (Prentice and Viner 2013). The conclusion that pre-pubertal fat mass in particular is behind the association between the earlier timing of puberty and later adiposity in males and females is supported by a UK study having longitudinal data of growth and direct measures of fat mass based on dual-energy X-ray absorptiometry (DEXA) scans from 9 to 18 years (O'Keeffe et al. 2020).
Our findings are consistent with Mendelian randomization analyses from the UK Biobank finding that high early BMI was associated with earlier timing of puberty whereas adult BMI has a negligible effect after controlling for early BMI (Richardson et al. 2020). This same study also found that prepubertal BMI and the timing of puberty act through adult BMI to affect the risk of coronary heart disease and type 2 diabetes. However, for breast cancer the associations were different and the study showed strong evidence for a direct causal effect of earlier age of menarche on higher risk of breast cancer independent of early life or adult body size (Richardson et al 2020). The evidence of a causal effect of early puberty on breast cancer risk was also found in twin studies where earlier puberty was associated with earlier diagnosis of breast cancer in MZ twin pairs concordant for breast cancer (Hamilton & Mack 2003) and with higher breast cancer risk in twin pairs discordant for early-onset breast cancer (Swerdlow et al 2002). This evidence suggests that even when early puberty is associated with many diseases, the causal pathways between early BMI, puberty timing, adult BMI and disease risk can vary according to the specific disease.
The mechanisms behind the found associations are still unclear. The shared genetic background of the timing of puberty and obesity can be due to a pleiotropic effect but also causality. If higher childhood adiposity affects the earlier timing of puberty, it can create a genetic correlation between these traits. Further, our results do not exclude the possibility that environmental factors can contribute to this association. BMI has globally increased during the recent decades (NCD Risk Factor Collaboration 2016), but the heritability estimates of BMI have been largely constant since both genetic and environmental variations have increased at the same time (Silventoinen et al. 2017). In a similar way, environmental factors may affect more children having genetic susceptibility to adiposity, and increased fat mass can then affect the earlier timing of puberty. Regardless of the mechanisms behind this association, the genetic background between the timing of puberty and BMI supports the conclusion that early puberty is not an independent risk factor of adult obesity, but rather that the origin of this association is in childhood.
Our data have both strengths and weaknesses. Our main strength is that we have longitudinal information on puberty timing and BMI in genetically informative data in both males and females. Having puberty measures for males is a major advantage, since most of the previous studies have used menarche age as a puberty indicator and thus include only females. Additionally, BMI and the timing of puberty were measured in adolescence thus decreasing recall bias which is a problem especially in males. However, it is noteworthy that PA was measured retrospectively at age 17 and was only based on one question, which may have increased measurement error. This may explain why our results for PA were generally weaker than for PDS. We were able to use two genetic designs—classical genetic twin study and molecular genetic data—each making different theoretical assumptions and thus increasing the robustness of the results. Our main weakness is that we needed to rely on self-reported data on measuring both puberty and obesity. Having the assessment of puberty stage done by trained healthcare professionals and measures of fat mass by magnetic resonance imaging or DEXA would have increased the preciseness of our results. Further, our study size was somewhat underpowered for the analyses using the PGSs of obesity indices, especially when testing sex differences.
In conclusion, our results indicate that early puberty is not an independent risk factor of adult obesity but rather reflects the genetic predisposition to obesity. Creating less obesogenic environments for children would be especially important for those children having genetic susceptibility to adiposity and thus a higher risk for early puberty.
Data Availability
Because of the consent given by study participants and the high degree of identifability, data cannot be made publicly available. Data are available through the Institute for Molecular Medicine Finland (FIMM) Data Access Committee (DAC) for authorized researchers who have IRB/ethics approval and an institutionally approved study plan. For more details, please contact the FIMM DAC (fimm‐dac@helsinki.f).
Code Availability
Computer codes are available from the corresponding author.
References
Anderson CA, Duffy DL, Martin NG, Visscher PM (2007) Estimation of variance components for age at menarche in twin families. Behav Genet 37:668–677
Bell JA, Carslake D, Wade KH, Richmond RC, Langdon RJ, Vincent EE, Holmes MV, Timpson NJ, Davey Smith G (2018) Influence of puberty timing on adiposity and cardiometabolic traits: A Mendelian randomisation study. PLoS Med 15:1002641
Broad Institute (2021) Pan-UK biobank: Pan-ancestry genetic analysis of the UK biobank. Retrieved from https://pan.ukbb.broadinstitute.org/
Busch AS, Hagen CP, Juul A (2020) Heritability of pubertal timing: detailed evaluation of specific milestones in healthy boys and girls. Eur J Endocrinol 183:13–20
Busch AS, Hollis B, Day FR, Sørensen K, Aksglaede L, Perry JRB, Ong KK, Juul A, Hagen CP (2019) Voice break in boys-temporal relations with other pubertal milestones and likely causal effects of BMI. Hum Reprod 34:1514–1522
Collaborative Group on Hormonal Factors in Breast Cancer (2012) Menarche, menopause, and breast cancer risk: individual participant meta-analysis, including 118 964 women with breast cancer from 117 epidemiological studies. Lancet Oncol 13:1141–1151
Cousminer DL, Berry DJ, Timpson NJ, Ang W, Thiering E, Byrne EM et al (2013) Genome-wide association and longitudinal analyses reveal genetic loci linking pubertal height growth, pubertal timing and childhood adiposity. Hum Mol Genet 22:2735–2747
Cousminer DL, Stergiakouli E, Berry DJ, Ang W, Groen-Blokhuis MM, Körner A et al (2014) Genome-wide association study of sexual maturation in males and females highlights a role for body mass and menarche loci in male puberty. Hum Mol Genet 23:4452–4464
Day FR, Bulik-Sullivan B, Hinds DA, Finucane HK, Murabito JM, Tung JY, Ong KK, Perry JRB (2015) Shared genetic aetiology of puberty timing between sexes and with health-related outcomes. Nat Commun 6:8842
Day FR, Thompson DJ, Helgason H, Chasman DI, Finucane H, Sulem P et al (2017) Genomic analyses identify hundreds of variants associated with age at menarche and support a role for puberty timing in cancer risk. Nat Genet 49:834–841
Dick DM, Rose RJ, Pulkkinen L, Kaprio J (2001) Measuring puberty and understanding its impact: a longitudinal study of adolescents twins. J Youth Adolesc 30:385–399
Eckert-Lind C, Busch AS, Petersen JH, Biro FM, Butler G, Bräuner EV, Juul A (2020) Worldwide secular trends in age at pubertal onset assessed by breast development among girls: A systematic review and meta-analysis. JAMA Pediatr 174:e195881
Elks CE, Ong KK, Scott RA, van der Schouw YT, Brand JS, Wark PA et al (2013) Age at menarche and type 2 diabetes risk: the EPIC-InterAct study. Diabetes Care 36:3526–3534
Euling SY, Herman-Giddens ME, Lee PA, Selevan SG, Juul A, Sørensen TIA, Dunkel L, Himes JH, Teilmann G, Swan SH (2008) Examination of US puberty-timing data from 1940 to 1994 for secular trends: panel findings. Pediatrics 121:S172–S191
Friedman NP, Banich MT, Keller MC (2021) Twin studies to GWAS: there and back again. Trends Cogn Sci 25:866–869
Gill D, Brewer CF, Del Greco MF, Sivakumaran P, Bowden J, Sheehan NA, Minelli C (2018) Age at menarche and adult body mass index: a Mendelian randomization study. Int J Obes 42:1574–1581
Hamilton AS, Mack TM (2003) Puberty and genetic susceptibility to breast cancer in a case-control study in twins. N Engl J Med 348:2313–2322
Hur YM (2020) Common genetic influences on age at pubertal voice change and BMI in male twins. Twin Res Hum Genet 23:235–240
Jelenkovic A, Ortega-Alonso A, Rose RJ, Kaprio J, Rebato E, Silventoinen K (2011) Genetic and environmental influences on growth from late childhood to adulthood: a longitudinal study of two Finnish twin cohorts. Am J Hum Biol 23:764–773
Kaidesoja M, Aaltonen S, Bogl LH, Heikkilä K, Kaartinen S, Kujala UM et al (2019) FinnTwin16: a longitudinal study from age 16 of a population-based Finnish twin cohort. Twin Res Hum Genet 22:530–539
Kaplowitz PB (2008) Link between body fat and the timing of puberty. Pediatrics 121:S208–S217
Kaprio J, Rimpelä A, Winter T, Viken RJ, Rimpelä M, Rose RJ (1995) Common genetic influences on BMI and age at menarche. Hum Biol 67:739–753
Kujala UM, Palviainen T, Pesonen P, Waller K, Sillanpää E, Niemelä M et al (2020) Polygenic risk scores and physical activity. Med Sci Sports Exerc 52:1518–1524
Lajunen HR, Kaprio J, Keski-Rahkonen A, Rose RJ, Pulkkinen L, Rissanen A, Silventoinen K (2009) Genetic and environmental effects on body mass index during adolescence: a prospective study among Finnish twins. Int J Obes 33:559–567
Lloyd LJ, Langley-Evans SC, McMullen S (2012) Childhood obesity and risk of the adult metabolic syndrome: a systematic review. Int J Obes 36:1–11
Locke AE, Kahali B, Berndt SI, Justice AE, Pers TH, Day FR et al (2015) Genetic studies of body mass index yield new insights for obesity biology. Nature 518:197–206
NCD Risk Factor Collaboration (2016) Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1698 population-based measurement studies with 19·2 million participants. Lancet 387:1377–1396
Neale MC, Hunter MD, Pritikin JN, Zahery M, Brick TR, Kirkpatrick RM et al (2016) OpenMx 2.0: extended structural equation and statistical modeling. Psychometrika 81:535–549
Oehme NHB, Roelants M, Bruserud IS, Madsen A, Bjerknes R, Rosendahl K, Juliusson PB (2021) Low BMI, but not high BMI, influences the timing of puberty in boys. Andrology 9:837–845
O’Keeffe LM, Frysz M, Bell JA, Howe LD, Fraser A (2020) Puberty timing and adiposity change across childhood and adolescence: disentangling cause and consequence. Hum Reprod 35:2784–2792
Ortega-Alonso A, Pietiläinen KH, Silventoinen K, Saarni SE, Kaprio J (2012) Genetic and environmental factors influencing BMI development from adolescence to young adulthood. Behav Genet 42:73–85
Papadimitriou A (2016) The evolution of the age at menarche from prehistorical to modern times. J Pediatr Adolesc Gynecol 29:527–530
Posthuma D, Beem AL, de Geus EJ, van Baal GC, von Hjelmborg JB, Iachine I, Boomsma DI (2003) Theory and practice in quantitative genetics. Twin Res 6:361–376
Prentice P, Viner RM (2013) Pubertal timing and adult obesity and cardiometabolic risk in women and men: a systematic review and meta-analysis. Int J Obes 37:1036–1043
Pulit SL, Stoneman C, Morris AP, Wood AR, Glastonbury CA, Tyrrell J et al (2019) Meta-analysis of genome-wide association studies for body fat distribution in 694 649 individuals of European ancestry. Hum Mol Genet 28:166–174
Richardson TG, Sanderson E, Elsworth B, Tilling K, Davey Smith G (2020) Use of genetic variation to separate the effects of early and later life adiposity on disease risk: mendelian randomisation study. BMJ 369:1203
Rose RJ, Salvatore JE, Aaltonen S, Barr PB, Bogl LH, Byers HA et al (2019) FinnTwin12 cohort: an updated review. Twin Res Hum Genet 22:302–311
Saarni SE, Pietiläinen K, Kantonen S, Rissanen A, Kaprio J (2009) Association of smoking in adolescence with abdominal obesity in adulthood: a follow-up study of 5 birth cohorts of Finnish twins. Am J Public Health 99:348–354
Sarnowski C, Cousminer DL, Franceschini N, Raffield LM, Jia G, Fernández-Rhodes L et al (2021) Large trans-ethnic meta-analysis identifies AKR1C4 as a novel gene associated with age at menarche. Hum Reprod 36:1999–2010
Silventoinen K, Haukka J, Dunkel L, Tynelius P, Rasmussen F (2008) Genetics of pubertal timing and its associations with relative weight in childhood and adult height: the Swedish Young Male Twins Study. Pediatrics 121:e885–e891
Silventoinen K, Jelenkovic A, Sund R, Hur Y-M, Yokoyama Y, Honda C et al (2016) Genetic and environmental effects on body mass index from infancy to the onset of adulthood: an individual-based pooled analysis of 45 twin cohorts participating in the collaborative project of Development of Anthropometrical measures in Twins (CODATwins) study. Am J Clin Nutr 104:371–379
Silventoinen K, Jelenkovic A, Sund R, Yokoyama Y, Hur YM, Cozen W et al (2017) Differences in genetic and environmental variation in adult body mass index by sex, age, time period, and region: an individual-based pooled analysis of 40 twin cohorts. Am J Clin Nutr 106:457–466
Swerdlow AJ, de Stavola BL, Floderus B, Holm NV, Kaprio J, Verkasalo PK, Mack T (2002) Risk factors for breast cancer at young ages in twins: an international population-based study. J Natl Cancer Inst 21(94):1238–1246
Turcot V, Lu Y, Highland HM, Schurmann C, Justice AE, Fine RS et al (2018) Protein-altering variants associated with body mass index implicate pathways that control energy intake and expenditure in obesity. Nat Genet 50:26–41
UK Biobank (2021) Genetic correlation between traits and disorders in the UK biobank. Retrieved from https://ukbb-rg.hail.is/rg_browser
Vogelezang S, Bradfield JP, Ahluwalia TS, Curtin JA, Lakka TA, Grarup N et al (2020) Novel loci for childhood body mass index and shared heritability with adult cardiometabolic traits. PLoS Genet 16:e1008718
Wehkalampi K, Silventoinen K, Kaprio J, Dick DM, Rose RJ, Pulkkinen L, Dunkel L (2008) Genetic and environmental influences on pubertal timing assessed by height growth. Am J Human Biol 20:417–423. https://doi.org/10.1002/ajhb.20748
Widén E, Silventoinen K, Sovio U, Ripatti S, Cousminer DL, Hartikainen AL, Laitinen J, Pouta A, Kaprio J, Järvelin M-R, Peltonen L, Palotie A (2012) Pubertal timing and growth influences cardiometabolic risk factors in adult males and females. Diabetes Care 35:850–856
Williams R (2000) A note on robust variance estimation for cluster-correlated data. Biometrics 56:645–646
Yengo L, Sidorenko J, Kemper KE, Zheng Z, Wood AR, Weedon MN et al (2018) Meta-analysis of genome-wide association studies for height and body mass index in ∼700000 individuals of European ancestry. Hum Mol Genet 27:3641–3649
Funding
Open Access funding provided by University of Helsinki including Helsinki University Central Hospital. Data collection has been supported by National Institute of Alcohol Abuse and Alcoholism (Grants AA-12502, AA-00145, AA-09203, AA15416 and AA018755) and the Academy of Finland (Grants 100499, 118555, 141054, 264146, 308248 and 312073).
Author information
Authors and Affiliations
Contributions
KS conducted the analyses and drafted the manuscript. TP created the polygenic risk scores. JK collected the data. AJ, TP, LD and JK contributed to the interpretation of data and critically revised the manuscript. All authors approve the version to be published and agree to be accountable for all aspects of the work.
Corresponding author
Ethics declarations
Conflict of interest
Karri Silventoinen, Aline Jelenkovic, Teemu Palviainen, Leo Dunkel, Jaakko Kaprio have declare that they have no conflict of interest.
Ethical Approval
The ethics committee of the Department of Public Health of the University of Helsinki (Helsinki, Finland), the ethics committee of the Helsinki University Central Hospital District (Helsinki, Finland), and the Institutional Review Board of Indiana University (Bloomington, IN, USA) approved the study protocols.
Informed Consent
Parents or legal guardians provided informed consent for the twins at ages 12 and 14, and the twins themselves provided informed consent at age 17 and later ages.
Consent for Participate
Written informed consent was obtained from all participants in the study.
Consent of Publication
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling Editor: Yoon-Mi Hur, PhD
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Silventoinen, K., Jelenkovic, A., Palviainen, T. et al. The Association Between Puberty Timing and Body Mass Index in a Longitudinal Setting: The Contribution of Genetic Factors. Behav Genet 52, 186–194 (2022). https://doi.org/10.1007/s10519-022-10100-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10519-022-10100-3