
Abstract
A detailed facies study of Early Permian strata within NE Svalbard reveals a fundamental change of the depositional setting, from a restricted-marine, warm-water carbonate platform to an open-marine, temperate-water, mixed siliciclastic-carbonate ramp. The uppermost strata of the Gipshuken Formation (Templet and Sørfonna members; Sakmarian–early Artinskian?) consist of microbialites (algal mats), mudstones, bioclastic/peloidal limestones, carbonate breccias and Microcodium facies reflecting peritidal platform areas and supratidal sabkhas. A mixed heterozoan/reduced photozoan assemblage indicates temperate-water conditions within neighboring deeper, open-marine mid-platform areas, while warm-water conditions still prevailed within inner platform zones. In contrast, the lowermost strata of the overlying Kapp Starostin Formation (Vøringen Member; late Artinskian?–Kungurian) show a fully heterozoan biotic assemblage reflecting temperate water conditions within open-marine, storm-dominated, nearshore to transitional offshore areas of a mixed carbonate-siliciclastic ramp. The Vøringen Member comprises three facies associations, which form a shallowing-upward sequence subsequent to an initial transgression. The sediments reflect bryozoan bioherms in most distal areas, followed by stacked tempestites of sandy brachiopodal shell banks and Skolithos piperocks, grading into broad sand flats in most proximal areas of the inner ramp. The above environmental change is regarded as a regional event taken place across the entire shelf along the northern margin of Pangea and is attributed to paleoclimatic, paleoceanographic, as well as paleogeographic changes, possibly related to the overall northwards drift of the supercontinent. An abrupt increase in terrigenous input coinciding with this change is ascribed to the uplift of a new local source area, probably to the north or east of the investigation area.
Similar content being viewed by others

Introduction
The Arctic archipelago of Svalbard, located at the north-western corner of the Barents Sea Shelf, reveals outstanding exposures of Late Paleozoic bedrock. While minor outcrops are present on Barentsøya and Edgeøya, the bulk of Late Paleozoic exposures are located on the main island Spitsbergen and on Nordaustlandet (Fig. 1).

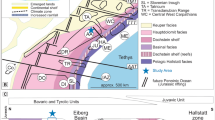
Geological overview of the Svalbard archipelago (a) and close-up of the investigation area (b) including the locations of the established sections in NE Spitsbergen (sections H1, E1) and within SW Nordaustlandet (section S1). LAFZ Lomfjorden-Agardhbukta fault zone, BFZ Billefjorden fault zone
Transitional strata of the Gipshuken Formation (Gipsdalen Group) and overlying Kapp Starostin Formation (Tempelfjorden Group) show completely different facies and hence record two fundamentally different depositional settings, which prevailed within the depositional area of Svalbard during the Permian. While evaporites and carbonates of the Gipshuken Formation are generally regarded as warm-water deposits of a peritidal carbonate platform, the sedimentary succession of the overlying Kapp Starostin Formation is interpreted to represent shallow to deeper marine environments of a mixed siliciclastic-carbonate ramp under temperate- to cold-water conditions.
The profound change from warm-water to cool- and cold-water environments has been documented in various circum-Arctic outcrop areas of Early Permian strata (offshore Barents Sea, eastern North Greenland, Arctic Canada, and Russia) and is attributed to the onset of the Permian Chert Event (Beauchamp and Baud 2002), a prolonged time period, during which massive cherts accumulated and cool- to cold-water conditions prevailed within the oceans of the northern hemisphere. From around the Sakmarian-Artinskian until the Late Permian, warm-water carbonate factories completely disappeared and were replaced by temperate and cold-water carbonate systems, while biogenic silica factories were preserved and progressively expanded from distal deep-water areas onto the shallow shelves along the western and northern margins of Pangea.
The Gipshuken and Kapp Starostin formations and their subunits are described and defined in earlier studies mainly from central and western Spitsbergen. While a number of publications provide detailed sedimentologic, paleontologic and biostratigraphic work on the Vøringen Member of the Kapp Starostin Formation, literature on the Templet and Sørfonna members of the Gipshuken Formation is scarce. The latter, first named “Gypsum beds” by Nathorst (1910) and later defined by Forbes et al. (1958) as “Upper Gypsiferous Series”, was first published under the name Gipshuken Formation by Cutbill and Challinor (1965) in a revised lithostratigraphic scheme for central and western Spitsbergen. The authors distinguished two horizons with extensive gypsum/anhydrite development (Upper and Lower Gypsum zones) and extensive solution breccias in the lower part. This general lithologic subdivision is still valid within the actual revised scheme of Svalbard (Dallmann et al. 1999), where a number of yet informal members are additionally introduced, based on different local lithologies. The uppermost part of two of these informal members, the Templet member within eastern Spitsbergen and the Sørfonna member of Nordaustlandet, are investigated within this study.
The overlying Kapp Starostin Formation and its lowermost subunit, the Vøringen Member were first mentioned by Nathorst (1910) as “Productus-bearing cherts” or “Spirifer Limestone”. Later, both units were included in a lithostratigraphical scheme as “Brachiopod Cherts” and “Limestone A” by Gee et al. (1952) and Forbes et al. (1958) and finally termed Kapp Starostin Formation and Vøringen Member by Cutbill and Challinor (1965). Publications on the Kapp Starostin Formation including the Vøringen Member mainly comprise paleontological studies focusing on taxonomy, biostratigraphy, and ecology of the varied faunal assemblage. Besides corals (Tidten 1972; Ezaki and Kawamura 1992; Ezaki 1997; Nowinski 1997; Chwieduk 2007), conodonts (Nakrem 1991; Szaniawski and Malkowski 1979) and ostracodes (Olempska and Blaszyk 1996), the bulk of publications concentrate on abundantly occurring brachiopods (Birkenmajer and Czarniecki 1960; Gobbett 1963; Biernat and Birkenmajer 1981; Nakamura et al. 1987; Malkowski 1988; Stemmerik 1988; Shen et al. 2005) and bryozoans (Nakrem 1988, 1994a, b; Nakrem and Spjeldnæs 1995), which have also been used for paleoenvironmental interpretations. In connection with the possible potential as hydrocarbon reservoir, a number of publications involve the Late Paleozoic strata of Svalbard. Cecchi et al. (1995), Samuelsberg and Pickard (1999) and Ehrenberg et al. (2001) discuss sedimentary environments, the regional sequence stratigraphic arrangement and deliver a supraregional correlation with coeval lithostratigraphic units in the Barents Sea, Arctic Canada, and Russia.
Within this article, a comprehensive facies study of the Templet and Sørfonna members of the Gipshuken Formation (Gipsdalen Group) and the overlying Vøringen Member of the Kapp Starostin Formation (Tempelfjorden Group) within NE Svalbard is provided.
Outcrop observations are combined with detailed microfacies studies comprising the description and interpretation of the sedimentary facies as well as the occurrence and composition of the biota. The investigations form the basis for the reconstruction of two fundamentally different depositional settings, comprising the transition from warm-water to temperate-water conditions in Svalbard. Possible causes for the environmental change within the Early Permian, including the interplay of a number of controlling factors, are discussed.
Geological setting
Late Paleozoic paleogeography
During the Late Paleozoic, Svalbard was located at the northern margin of Pangea, closely related to parts of the Finnmark Platform and Stappen High (Barents Sea), the Wandel Sea Basin (eastern North Greenland), the Sverdrup Basin (Arctic Canada) and the Timan-Pechora Basin (Russia). The extensive, epicontinental shelf area, arranged into a series of intracratonic platforms and basins, gradually drifted from approximately 25°N in the Late Carboniferous to around 45° in the Late Permian, with Svalbard around the 35th°N during the Artinskian (Scotese and Langford 1995; Ziegler et al. 1997; Golonka 2002).
During mid-Carboniferous times (Serpukhovian–Moscovian) Svalbard was tectonically active. Extensional rift tectonics along major, pre-existing structural lineaments (e.g., Billefjorden Fault Zone, Lomfjorden Fault Zone) led to the development of a pronounced horst-and-graben setting, comprising a series of roughly NNW-SSE-trending, elongated, narrow rift basins (e.g., Billefjorden Trough, St. Jonsfjorden Trough, Lomfjorden Trough) and adjacent highs (e.g., Ny Friesland High, Nordfjorden High, Sørkapp-Hornsund High; Steel and Worsley 1984; Worsley et al. 1986). Under arid to semi-arid climatic conditions, initial terrestrial red beds of the Gipsdalen Group were deposited and in the course of a marine transgression replaced by restricted marine platform deposits forming an intricate interplay caused by the diversified paleogeography (Steel and Worsley 1984; Pickard et al. 1996). From the Moscovian, active rifting ceased and a stable carbonate shelf marked by more uniform subsidence developed. Structural highs (except of the Sørkapp-Hornsund High) were progressively transgressed (Johannessen and Steel 1992; Hüneke et al. 2001) and carbonate platforms nucleated on the blocks (e.g., Pyefjellet beds on Ny Friesland High, Kapitol Member on Nordfjorden High), resulting in the widespread deposition of cyclic, highly diverse, fossiliferous, warm-water carbonates reflecting open- to restricted-marine depositional environments (Blomeier et al. 2009).
A following regression during the Early Permian (Sakmarian–Artinskian) resulted in the prolonged subaerial exposure, intense karstification and collapse of elevated platform areas, while restricted marine carbonates (dolomites) and evaporites formed within peritidal, depositional basins. Within the Artinskian, the regression culminated and a major hiatus developed all over Svalbard, marking the upper boundary of the Gipshuken Formation (Gipsdalen Group; Fig. 2).

With the subsequent transgressive onset of sedimentation during the late Artinskian, an unconformity formed at the base of the overlying Tempelfjorden Group (Artinskian–Kazanian?/Tatarian). By then, the depositional area was probably marked by a more subdued paleorelief, resulting from the former karstification (emersion), subsequent reworking (transgression) and a general uniform subsidence. Thus, a temperate, mixed siliciclastic-carbonate ramp marked by sedimentation of cherts, sandstones, shales and partly silicified, highly fossiliferous limestones developed over nearly entire Svalbard. Only the southernmost parts of Spitsbergen, comprising the Sørkapp-Hornsund High, remained subaerially emerged until the end of the Permian.
Permian sedimentary record of Svalbard
The Late Paleozoic sedimentary record of Svalbard comprises two major lithostratigraphic units, the Early Carboniferous to Early Permian Gipsdalen Group and the Early to Late Permian Tempelfjorden Group (Fig. 2).
Early Permian (Asselian–early Artinskian) deposits of the Gipsdalen Group comprise the upper part of the Wordiekammen Formation (Tyrrellfjellet Member in Spitsbergen; Idunfjellet Member in Nordaustlandet) and the overlying Gipshuken Formation, which is arranged into a number of local, yet informal members (Dallmann et al. 1999). The latter formation (late Sakmarian–early Artinskian) displays a regressive sequence, during which extended platform areas emerged and a prolonged hiatus developed at the top. The deposits, mainly composed of stacked evaporite/carbonate cycles and carbonate breccias, display restricted, peritidal platform areas and extended, supratidal sabkhas, on which the marginal marine platform strata were subaerially exposed (Lauritzen 1981a, b). Hence, the lower part of the up to 280-m-thick Gipshuken Formation is characterized by massive carbonate breccias in western Spitsbergen (Kloten member) and eastern Svalbard (Zeipelodden member), interfingering with rhythmically bedded, evaporite-dominated successions (Vengeberget member) in central Spitsbergen. Above, the dolomite-dominated Skansdalen member represents a warm-water marginal-marine area marked by restricted peritidal flats, lagoons and supratidal sabkha environments in western and central Spitsbergen. To the east, the sediments interfinger with the 100- to 135-m-thick Templet member, which is suggested to represent more open-marine areas. On Nordaustlandet, the sedimentary succession of the contemporaneous Sørfonna member consists mainly of carbonates (limestone, dolomite) with thin coaly interbeds, reflecting near-coastal environments (Dallmann et al. 1999). However, the environmental interpretation, stratigraphic age determinations, lateral transitions and boundary definitions of these informal members are still uncertain, partly due to an excessive dolomitization and intensive karstification of the strata.
The overlying Tempelfjorden Group is separated by a major disconformity comprising the Early Permian (Artinskian) hiatus on top of the Gipsdalen Group (Fig. 2). On nearly entire Svalbard, the Tempelfjorden Group is comprised solely of the Kapp Starostin Formation, which wedges out towards the Sørkapp-Hornsund High in south-western Spitsbergen.
The basal part of the up to 460-m-thick Kapp Starostin Formation is defined as Vøringen Member, which occurs within most areas of Svalbard (apart from Brøggerhalvøya in western Spitsbergen). The up to 40-m-thick succession consists of partly strongly silicified, sandy, fossiliferous limestones to allochemic sandstones, which formed on a transgressive, stable marine shelf during the late Artinskian to Kungurian. The Vøringen Member features a diverse fauna consisting of brachiopods, bryozoans, echinoderms, siliceous sponges and various trace fossils. Coarse-grained bioclastic limestones have been interpreted as nearshore deposits, formed during the transgression of the restricted marine platform and sabkha strata of the Gipshuken Formation below (Hellem 1980; Steel and Worsley 1984). While the lower boundary of the member is thus clearly defined by the occurrence of an unconformity and an abrupt facies change, the definition of the upper boundary is still vague (Dallmann et al. 1999). Herein it is defined with the occurrence of a distinct black shale horizon capping the entire succession of the member below in all investigated sections.
Above the Vøringen Member, the strata of the Kapp Starostin Formation are characterized by recurrent successions consisting of basal shales and dark to light-colored chert horizons, developing into partly glauconitic sandstones and strongly silicified, fossiliferous limestones, all interpreted to reflect shoreface to deeper shelf environments (Ehrenberg et al. 2001; Hüneke et al. 2001). So far, informal, local members have been defined within different areas on Spitsbergen. While the Stensiöfjellet and Revtanna members are present in central and western Spitsbergen, respectively (Dallmann et al. 1999), the Hovtinden and Svenskeegga members occur in central and southern Spitsbergen (Cutbill and Challinor 1965). On Nordaustlandet, parts of the strata above the Vøringen Member are arranged into the Palanderbukta member (Lauritzen 1981b) and Selanderneset member (Burov et al. 1965). In field, the current definition and facies description of the different members (Dallmann et al. 1999) are often hard to comprehend. In addition, neither the stratigraphic relations of the informal members nor the overall lithostratigraphic development of the Kapp Starostin Formation are fully understood yet. Future investigations will hopefully result in an improved internal arrangement and stratigraphic resolution of the formation.
The lithological, upper boundary of the Tempelfjorden Group (Kapp Starostin Formation) to the overlying, Early to Middle Triassic Sassendalen Group is sharp and distinct due to an abrupt onset of a monotonous, siliciclastic succession of mainly dark shales and siltstones with minor sandstones accompanied by the termination of biotic deposits such as cherts or bioclastic limestones (Ehrenberg et al. 2001). Contrary to the distinct facies change, age constraints of the upper boundary are rather unclear and inconsistent as different opinions exist as to the exact position of the Permian–Triassic boundary (Wignall et al. 1998; Mangerud and Konieczny 1993) as well as regarding the overall upper stratigraphic reach of the Tempelfjorden Group and whether there is a gap in the sedimentary record or conformable sedimentation into the Triassic (Mørk et al. 1989; Stemmerik 1997; Ehrenberg et al. 2000; Fig. 2).
Age determinations of the Gipshuken and Kapp Starostin formations are generally based on biostratigraphy. Within the Gipshuken Formation, age datings rely on brachiopods, bryozoans, rare conodonts, foraminifers and palynomorphs, performed by Sosipatrova (1967), Mangerud and Konieczny (1991), Nakrem et al. (1992), Nakrem (1994a, b) and Buggisch et al. (2001). The authors indicate a late Sakmarian? to early Artinskian age of the formation (Fig. 2).
The age of the Kapp Starostin Formation (late Artinskian–Kazanian/earliest Tatarian?) has been established by Szaniawski and Malkowski (1979), Biernat and Birkenmajer (1981), Nakamura et al. (1987), Stemmerik (1988), Nakrem (1988, 1994a, b), Nakrem et al. (1992), Buggisch et al. (2001) and Chwieduk (2007) by means of bryozoans, brachiopods, corals, conodonts and palynomorphs. Nakrem et al. (1992) and Nakrem (1994a) indicate a late Artinskian to early Kungurian age of the Vøringen Member based on conodonts, bryozoan zones, and former determinations of brachiopods (Nakamura et al. 1987), small foraminifers, and palynomorphs. However as cool-water faunas from Arctic Canada, the Barents Sea and Russia differ substantially from warm-water faunas of North America and the Tethys, the exact correlation of both the Gipshuken and Kapp Starostin formations with the Middle/Late Permian Standard Global Stages is still problematic.
Methods
The data base of this paper originates from three vertical sections (sections H1, E1 in E-Spitsbergen; section S1 in SW-Nordaustlandet; Fig. 1), which were established in selected key localities in connection with ongoing geological mapping of Late Paleozoic bed rocks in eastern Svalbard during several weeks from July to August in 2005 and 2006. From these sections, only the transitional strata of the Gipshuken and overlying Kapp Starostin formations are displayed within this publication.
Outcrop investigations are combined with detailed microfacies studies. A total number of 71 (section E1: 19; section H1: 36; section S1: 16) over-sized thin sections (7.5 × 10 cm to 5 × 5 cm) were used to investigate the compositional variation of the sediments of the vertical sections.
The deposits show strongly varying contents of terrigenous material (mainly detrital quartz grains) and thus range from pure carbonates, mixed siliciclastic-carbonate deposits to pure siliciclastic sediments. Carbonates with a proportion of less than 10% detrital quartz grains of the components are described according to the classification schemes of Dunham (1962) and Folk (1959). The description of the mixed siliciclastic-carbonate deposits follows the classification of Mount (1985), based on the ratio of siliciclastic versus carbonate material. In addition, characteristic fabrics and main component categories have been taken into account for the name of the different microfacies types. Siliciclastic sediments, nearly entirely consisting of detrital quartz grains (>90%) were classified after Wentworth (1922). On this basis, a semi-quantitative facies analysis has been applied and altogether ten microfacies types are described (Table 1).
Facies analyses
Field data
Within this chapter, outcrop observations including colors, lithologies, textures, structures, and macrofossils of the sedimentary strata within each vertical section are described.
Eremitten section (E1, Fig. 3)
At least 22 m of the uppermost part of the Templet member (Gipshuken Formation) are exposed at the locality Eremitten (Figs. 1, 3), forming the lower, partly scree-covered part of a coastal cliff wall. The exposed strata of the member are formed of light grey to brownish, thick- to intermediate-bedded, massive carbonate beds (limestones and dolostones), which are arranged into recurrent, 1 to 3-m-thick sediment packages, separated by up to several-centimeter-thick, dark-grey or brownish claystone horizons. The carbonate-bed sets form fining-upward, respectively, thinning-upward sequences. These are often marked by sharp, erosive subsurfaces and basal, coarse-grained lag horizons characterized by ruditic intraclasts, mud chips, and coaly plant fragments. Cross-bedding is a common feature mainly in the lower and middle part of the sets. Bioturbation displayed as tubular, vertical burrows is often visible in the middle part, while horizontal lamination is present mainly in the upper part of the limestones (microbial limestones, MFT 1; Fig. 4a) and the claystone interbeds.

Sedimentary succession at Eremitten, comprising lithologies, colors, fabric, fossils, and the microfacies types of the uppermost part of the Templet member and Vøringen Member. The location of the section (E1) is marked in Fig. 1b

a Location Eremitten, Templet member: algal mats often show a distinct horizontal to wavy lamination, which can be recognized due to the alternation of light and darker grey, micritic laminae. Diameter of lens cap is 3 cm. b Section H1/bed 24, Templet member: the thick- to medium-bedded algal mats are often broken and strongly fragmented. Irregularly voids between the ruditic fragments are filled with white-translucent or clear sparite (arrow) and/or internal sediment. Diameter of lens cap is 3 cm. c Location Hódbreen, Templet member: local accumulations of up to a few centimeters large, coaly plant fragments (A) within a dolomite bed. The plant remains occur together with smaller, whitish gastropod? Shells (arrows). Diameter of lens cap is 3 cm. d Section S1/bed 1, Sørfonna member: this light grey, massive dolomite breccias contain various lithoclasts as only components. The multicolored, darker grey or green, poorly sorted, and angular to edge-rounded components reach sizes of up to a few centimeters. e Section H1, boundary Templet/Vøringen members: up to 50-cm-high and 1-m-wide trepostome bryozoan bioherms (A) occur at the base of the Vøringen Member. The boundary to the underlying mudstones of the Gipshuken Formation is marked by a hardground (discontinuity surface, arrow), which served as growth substrate for the colonies. F Section H1, boundary Templet/Vøringen members: Close-up of the hardground of Fig. 5. The surface is characterized by abundant, vertical oriented, ledge-shaped borings, which penetrate the muddy sediments of the Templet member (A) and are filled with arenitic, skeletal material of the Vøringen Member (B). The length of the displayed detail is ca. 9 cm. G Section S1, boundary Sørfonna/Vøringen members: Horizontal bedded, dark-weathering carbonates of the Gipshuken Formation (A) are separated by a sharp, slightly irregularly boundary from light-grey, mixed siliciclastic-carbonate deposits of the overlying Vøringen Member (B). The latter are marked by trough- and planar-cross bedding (arrow, bedding planes emphasized). Hammer for scale. h Section H1/unit 30: the occurrence of distinct sedimentary couplets is a prominent feature of the Vøringen Member. The couplets generally consist of greyish or brownish colored, coarse-grained, thick- to medium-bedded, sandy, skeletal limestones (MFTs 7, 8) at the base (beds 1a, 2a). The upper, medium-bedded horizons are formed of lighter colored, generally finer-grained, sandy limestones to skeletal sandstones (MFTs 9, 10), which are intensely bioturbated (beds 1b, 2b)
At the top of the Gipshuken Formation an approximately 80-cm-thick, multicolored dolomite breccia is overlain by a centimeter-thick, rusty-weathering, dark limestone horizon, which marks the sharp boundary to the overlying, ca. 18-m-thick Vøringen Member of the Kapp Starostin Formation (Fig. 3).
The lower, ca. 10-m-thick part of the Vøringen Member is characterized by coarse-grained, thick- to medium-bedded, allochemic sandstones to sandy, allochemic limestones showing grey or brownish weathering colors, rarely with a greenish or rusty tinge.
Sandy, allochemic limestones at the base are characterized by ruditic fragments of brachiopods, bryozoans, crinoids, and more rarely gastropods, as well as locally occurring, brownish lithoclasts up to a few centimeters in size (mixed-bioclastic, sandy allochem limestones, MFT 7). Further up in the strata, the diversity of the components generally decreases and coarse-grained brachiopod deposits (brachiopodal, sandy allochem limestones to allochemic sandstones, MFT 8) containing abundant to frequent bioclasts and biogens of mainly spiriferid and productid brachiopods occur. These bioclasts are either enriched in certain horizons within the single beds or show a general decrease in size and proportion upwards (graded bedding). Quartz sand is generally present in varying amounts, forming the arenitic matrix. The bedding planes are wavy to slightly wavy and continuous. Fe-staining and cross-bedding is occasionally a prominent feature. The coarse-grained brachiopod deposits are commonly overlain by medium-bedded, fine-grained, sandy allochem limestones to allochemic sandstones (MFT 9) and calcareous sandstones (MFT 10) forming the upper part of stacked sediment couplets. The generally thinner horizons are marked by intensive bioturbation due to abundant, simple tubular Skolithos burrows oriented perpendicular to the bedding planes, 0.5–1.0 cm in diameter and up to 25 cm long (Fig. 5a). The intense bioturbation has apparently led to local silicification marked by whitish discolorations of the otherwise light grey to light brownish sediments. With upwards decreasing thickness of the sediment couplets, the preservation of Skolithos degrades and sediments of this type often appear as discontinuous, white horizons or lenses.

a Section E1/bed 33, Vøringen Member: close-up of an upper horizon of a sediment couplet marked by abundant, simple tubular, perpendicular to the bedding plane oriented Skolithos burrows. The whitish color is due to silicification. Diameter of lens cap is 3 cm. b Section S1, Vøringen Member: the upper part of the member consists of a thick succession of lateral discontinuous-bedded and highly bioturbated, sandy allochem limestones to allochemic sandstones (A), overlain by a distinct green (glauconitic) allochemic sandstone horizon (B), followed by light-colored cherts, which in turn are capped by a distinct black shale horizon (C), bounding the entire lithostratigraphic unit. c Section S1/bed 13, Vøringen Member: burrows (A) within the mixed siliciclastic-carbonate sediments in the upper part of the member are often outlined by irregular, whitish discolorations reflecting a preferred silicification of the neighboring host sediments. Scale is 10 cm. d Section S1/bed 13, Vøringen Member: an intense bioturbation of the mixed siliciclastic-carbonate sediments of the Vøringen Member is often displayed by vertical oriented Zoophycos burrows extending from discontinuous bedding planes into the host sediment. Note the accumulation of coarser, partly rusty components on the bed surface (arrow). e Section E1, Vøringen Member: the mixed siliciclastic-carbonate succession of the Vøringen Member is capped by a dark claystone horizon (arrow) overlain by bedded dark grey cherts. The abrupt facies change at the upper boundary is often accompanied by rusty discolorations implying longer periods of reduced sedimentation rates. Hammer for scale. f Section H1/sample 24.1b, Templet member: algal mats (microbialites, laminated bindstones, MFT 1) are commonly characterized by an undulating, fine-horizontal lamination. The latter is due to alternating densities of micritic laminae and abundant, elongated open-space structures filled by blocky spar. g Section E1/sample 9.1, Templet member: local fragmentation of the algal mats (MFT 1) is displayed by poorly sorted, angular to edge-rounded components (A) embedded in a lighter microsparitic matrix or surrounded by blocky sparite. The darker bindstone intraclasts often show a primary, internal lamination (arrow) resulting from microbially induced carbonate precipitation. h Section E1/sample 7.1, Templet member: a sandy mudstone (MFT 2) shows the nest-like accumulation of well-rounded, ovoid, homogenous peloids (fecal pellets, arrows) and a plant fragment (A). The latter is characterized by a black color due to advanced coalification of the organic matter and an internal cellular fabric. Frequent silt to very fine sand-sized quartz grains reflect enhanced terrestrial input within restricted areas
The entire series gradually passes into a more uniform succession of partly glauconitic, calcareous sandstones (MFT 10) with thin intercalated brachiopod and whitish chert horizons, comprising the entire upper part of the Vøringen Member (Fig. 5e). The medium-bedded, light brownish to greenish sediments show discontinuous, slightly wavy bedding planes and intense bioturbation, dominated by Zoophycos. At the top of the Vøringen Member, rusty weathering colors occur. The upper boundary is characterized by an abrupt facies change and clearly marked by the occurrence of a several-centimeter-thick, black shale horizon, overlain by a several-meter-thick, dark chert succession.
Hódbreen section (H1, Fig. 6)
At this location (Fig. 1), a several-meter-thick dolomite breccia, the Zeipelodden member (Gipshuken Formation; Fig. 2) locally crops out at the base of the cliff wall.

The lower part of the overlying strata of the Templet member is mostly covered by scree, but light brownish to greyish, strongly cavernous, thick- to medium-bedded dolomite beds, locally containing accumulations of coaly plant fragments (Fig. 4c) and large lithoclasts occur in a number of smaller outcrops. At the very top of the member, an at least ca. 5-m-thick succession of mainly thick- to medium-bedded limestones is present, displaying the lowermost strata in the section H1. These limestones mainly consist of thick algal mats and mudstones (MFTs 1, 2; Fig. 4b), which are partly strongly fragmented, brecciated, and reworked.
The abrupt boundary between the Templet member and the overlying, ca. 21-m-thick Vøringen Member (Kapp Starostin Formation) is reflected by a distinct hardground. Intensely Fe-stained, highly bioturbated, silty mudstones of the Templet member are penetrated by borings and desiccation cracks, partly backfilled with arenitic, bioclastic material from the Vøringen Member above (Figs. 4e, f, 8a).
Situated on this hardground, bioherms formed of branching, trepostome bryozoans up to 50 cm high and 1 m wide are abundantly present and preserved in situ (Fig. 4e). The bioherms appear on different levels within the lower 2 m of the Vøringen Member and are embedded and covered by a series of medium-bedded, coarse-grained, partly sandy, mixed-bioclastic limestones (MFT 7). These comprise the coarse debris of the bioherms as well as arenitic to ruditic fragments of brachiopods, crinoids, and occasional, up to a few-centimeters-large chaetetid colonies. The bioclastic limestones are associated with calcareous sandstones (MFT 10), reflecting a high terrigenous input due to the local accumulation of sand-sized quartz grains.
Upwards in strata, the biotic assemblage of the allochthonous material changes from a mixed-bioclastic (mainly bryozoans, echinoderms, brachiopods) into a brachiopod-dominated association (MFT 8). Coevally, bioturbation increases, reflected by a variety of trace fossils (e.g., Skolithos, Zoophycos), which often favor silicification, resulting in whitish discolorations around preferably vertically oriented burrows. The sedimentary beds are often arranged into stacked sediment couplets (Fig. 4h), mainly formed of coarse-grained, brachiopodal, sandy limestones to sandy allochem sandstones (MFT 8), overlain by generally fine-grained, sandy allochem limestones to calcareous sandstones (MFTs 9, 10). Particularly, the latter show abundant Skolithos burrows, which are often strongly silicified.
In the upper part of the section, the sediments gradually change into a more homogenous succession formed of strongly silicified, thin brachiopodal limestones and thicker calcareous sandstones (MFT 10), which are often marked by greenish colors due to the presence of glauconite. The upper boundary of the Vøringen Member is marked by the occurrence of a 5 to 10-cm-thick, black shale horizon, capping the entire succession.
Selanderneset section (S1, Fig. 7)
In the south-western part of Nordaustlandet (Gustav Adolf Land), the Kapp Starostin Formation crops out in a ca. 20-km-long coastal exposure (Stupstranda), forming the eastern shores of the Hinlopenstretet (Fig. 1). Here, the sharp boundary between the Gipshuken Formation and the overlying Kapp Starostin Formation is accessible in a number of localities at the base of the cliff wall (Fig. 4g).

At the section location, the uppermost meters of the Sørfonna member (Gipshuken Formation) are exposed. The sediments consist of grey to light grey, thick- to medium-bedded, massive carbonates (limestones and dolomites) and up to several-centimeters-thick, multicolored or grey, partly marly claystone horizons, intercalated in irregular intervals. They often show a fine internal horizontal lamination and separate the carbonates on slightly wavy, but continuous bedding planes. Besides heavily fragmented algal mats (MFT 1), the uppermost carbonate beds also contain poorly sorted dolomite breccias (MFT 6) marked by angular to edge-rounded lithoclasts, which are embedded within a fine-grained matrix (matrix-supported fabric). The greenish, greyish, or brownish colored lithoclasts are up to a few centimeters large and display the single ruditic component type (Fig. 4d). The sharp boundary between the Sørfonna member and the overlying Vøringen Member is formed by a several-centimeter-thick, dark grey claystone horizon marked by rusty stains.
The Vøringen Member constitutes a roughly 11-m-thick succession, which is bounded at the top by a distinct, ca. 20 cm thick, dark shale horizon (Fig. 5b).
Similar to the exposure at Hódbreen, in situ trepostome bryozoan buildups, up to 30 cm high, are the key feature within the lowermost limestone beds. The bioherms, which occur on different levels within the lower 1.5 m of the Vøringen Member, are embedded in partly cross-bedded, coarse-grained, mixed-bioclastic, sandy allochem limestones (MFT 7). Besides bryozoan bioherms, rounded chaetetid colonies of up to a few centimeters occur occasionally.
Higher upwards, brachiopodal deposits are only scarcely developed and an arrangement in sediment couplets is only vague. Instead, calcareous sandstones (MFT 10), containing medium- to coarse sand-sized quartz grains, form the bulk of the strata within this location. Bioturbation (mainly Zoophycos) within the sandstones is distinct, in places intensely developed and outlined by whitish decolorizations (Figs. 5c, d). Due to the latter, the medium-bedded sandstones show laterally discontinuous bedding planes and amalgamate with each other. Whitish chert nodules and lenses occur occasionally. An increase in glauconite content is displayed by a gradual color change from brownish to distinct greenish upwards.
Microfacies analysis
In addition to the outcrop observations, detailed microfacies studies are applied, comprising the description, occurrence and environmental interpretation of ten microfacies types (MFTs; Table 1).
Each MFT is defined by a specific composition and texture and thus can be linked to specific depositional processes and environments. While MFTs 1–6 occur within the investigated sediments of the Gipshuken Formation, MFTs 7–10 are restricted to the strata of the Kapp Starostin Formation. An exception is MFT 3 (Spiculites), which appears in both formations although only to a minor extent in the investigated Gipshuken strata.
MFT 1: microbial limestones (Figs. 5f, g)
Description: This MFT comprises various deposits originating from microbial (algal) carbonate precipitation. The autochthonous to par-autochthonous deposits form thick- to medium-bedded, greyish, partly brownish limestone beds, mainly composed of laminated algal mats. The bindstones consist of a micritic groundmass (without organic microstructures), which shows a distinct to diffuse horizontal to wavy lamination due to the changing density of micrite and abundant open-space structures between and within single laminae (stromatolites after Riding 1999). These conspicuous, fenestral, blocky spar-filled voids generally have elongated shapes with lengths of up to some centimeters and occasionally show micritic internal sediments at the base (stromatactis). Secondary cracks and intense fragmentation are common features, cutting through the primary fabric of the microbialites. In places, large, irregular voids filled by multiple generations of calcite cements and/or poorly sorted internal sediment consisting of a fine micritic to microsparitic groundmass and up to several-centimeter-large, angular to edge-rounded algal intraclasts, are present. Clotted fabrics marked by the occurrence of irregularly shaped and -sized, rounded aggregates and peloids of dense micrite within lighter microsparite occur rarely. Other components such as arenitic skeletal fragments (mainly echinoderms) and silt to fine sand-sized detrital quartz grains are generally scarce. Microcodium crystals are locally present either as single crystals, incorporated within the micrite or as composite, thin crusts within some elongated cavities.
Interpretation: Microbial limestones have been described from various settings including marine, marginal-marine and non-marine depositional environments (Flügel 2004). Here, well-developed algal mats formed within marginal-marine, peritidal flats of a warm-water carbonate platform. Within the intertidal area, sea-level fluctuations are presumed to be responsible for the lamination of the microbialites themselves, as they controlled the episodic trapping, binding and precipitation of fine-grained carbonate. Longer exposure times probably led to the occurrence of desiccation cracks and local fragmentation of the algal mats, displaying the disruption and collapse of individual horizons of the stromatolites. The occurrence of single, allochthonous Microcodium crystals and the association with the Microcodium facies, point to the vicinity of a supratidal sabkha in a landward direction. This MFT is comparable with the laminated facies (dolo-laminites and microbial structures) of Hüneke et al. (2001), who state that the microbial origin is clearly obvious and dolomitization is due to a later diagenetic overprint.
MFT 2: mudstones (Figs. 5h, 8a)
Description: Mudstones (micrites, dismicrites) and more rarely wackestones nearly entirely consist of grey or reddish, partly silty to sandy micrite/microsparite. The fine-grained, generally component-poor deposits show scattered Microcodium and dolomite crystals, peloids formed of dense micrite, siliceous sponge spiculae, recrystallized bioclasts and/or coaly plant fragments. In places, well-sorted, spherical fecal pellets and/or sand-sized quartz grains are enriched in nests or lenses. Occasionally, the mudstones show opaque mineral separations, pyrite crystals or Fe-staining and are characterized by irregularly shaped cavities, which are filled with blocky sparite and/or internal sediment consisting of skeletal, peloidal or micritic/microsparitic material. Locally, intense bioturbation is obvious.

a Section H1/sample 25.1, Templet member: this reddish, intensely Fe-stained mudstone (MFT 2) from the top of the Gipshuken Formation shows an intensive bioturbation. The latter is reflected by irregularly shaped cavities (A), interpreted as the remains of partly collapsed burrows, filled with light-grey, internal sediment, which in turn is strongly altered and recrystallized. b Section H1/sample 23.1, Templet member: microspiculites (MFT 3) are characterized by a low-diverse component composition, mainly consisting of abundant monazon microscleres embedded in a micritic matrix, forming packed wackestones to packstones. c Section H1/sample 42.1, Vøringen Member: abundant, densely packed, monazon megascleres, which are completely silicified, form a megaspiculite (MFT 3) within the shelter of a marginally dissolved brachiopod shell (A). d Section H1/sample 42.1, Vøringen Member: close-up of completely silicified, monazon megascleres showing various longitudinal, transversal, and cross sections. The central canals of the needles are occasionally filled with distinct green, authigenic glauconite minerals (arrows). e Location H1/sample 20.3, Templet member: this bioclastic, poorly sorted, and washed rudstone (MFT 4) is characterized by a heterozoan assemblage composed of trepostome bryozoans (A) and crinoids (B) embedded in a micritic matrix. Blocky and syntaxial sparite cements are present in well-washed areas. f Location E1/sample 11.1, Templet member: this well-sorted, bioclastic, peloidal packstone (MFT 4) shows photozoan elements comprising bioclasts of the dasycladacean algae Velebitella simplex (longitudinal sections, arrows). Frequent peloids (A) probably originate from algal mats or other microbial, encrusting organisms. g Section H1/sample 24.17, Templet member: Microcodium facies (MFT 5) is characterized by the abundant occurrence of single, light-brownish crystals forming low-diverse, packed wackestones to packstones. Note the desiccation cracks (arrow) around intensely stained carbonate nodules (A). h Section S1/sample 7.2: autochthonous Microcodium deposits (MFT 5) are characterized by composite layers or crusts, consisting of single, parallel oriented crystals. Here, Microcodium (A) occurs in a dismicrite replacing blocky spar and dense, probably microbial micrite
Interpretation: Mudstones generally originate from the accumulation of fine-grained, suspended sediment particles under quiet-water conditions. In the marginal marine platform setting of the Gipshuken Formation, the occurrence of mudstones is associated with the presence of algal mats reflecting protected areas, in which suspended matter accumulated. This interpretation is in accordance to the formation of the dismicrite facies of Hüneke et al. (2001). The local presence of abundant, sand-sized quartz grains and scattered plant fragments within the mudstones of the Gipshuken Formation points to restricted nearshore areas, in which terrestrial material accumulated.
Intense bioturbation and the precipitation of various minerals shown by opaque separations and intense Fe-staining of mudstones at the very top of the Gipshuken Formation imply sediment-starved conditions. The latter resulted in the formation of hardgrounds marked by condensed sediments, which were intensely burrowed, bored, and mineralized due to longer time available for pore-fluid migration.
MFT 3: spiculites (Figs. 8b–d, 9g)
Description: Spiculites (mainly spiculitic packstones, minor wackestones) are characterized by local, laterally restricted accumulations of siliceous sponge spicules embedded in often dense (microbial?) micrite, which is occasionally replaced by microquartz. The strongly silicified spiculites are composed of commonly to frequently occurring monazon mega- and/or microspiculae (microspiculites, megaspiculites). While microscleres show maximum lengths of 0.5 mm and a diameter of around 50 μm, macroscleres have length of up to 2 mm and a diameter of around 0.3 mm. The latter possess a central canal, which is filled with micrite, brownish mineral separations or greenish, authigenic glauconite minerals. Silt- to sand-sized, well-sorted quartz grains, fine arenitic to ruditic skeletal fragments (often strongly recrystallized and miciritized filaments, brachiopod shells) and peloids commonly occur as minor component categories within this MFT. The deposits are strongly affected by silicification to varying degrees, including the complete or partial replacement of the original matrix and/or the siliceous sponge spicules. In these areas, megascleres are often recognizable only due to earlier mineral linings.

a Section E1/sample 9.2: the edge-rounded to rounded intraclasts of this poorly sorted, monomict carbonate breccia (MFT 6) are completely recrystallized, forming a component-supported fabric. Interparticle spaces are cemented with blocky sparite or filled with dense, partly intensely Fe-stained micrite (microbial origin? Arrow). b Section S1/sample 8.1: this carbonate breccia (MFT 6) is marked by angular to edge-rounded intraclasts with an internal Microcodium facies. Voids of the intraclastic rudstone are filled with blocky sparite, lighter grey microsparite, and smaller intraclasts (peloids). c Section H1/sample 25.2: this poorly sorted, mixed-bioclastic rudstone (MFT 7) shows a heterozoan biotic assemblage consisting of coarse-grained fragments of trepostome (A) and fenestrate (B) bryozoans and echinoderms (C), enclosed by blocky sparite. Note the syntaxial cement around some echinoderm fragments (arrows). d Section E1/sample 37.3, Vøringen Member: main component categories of this coarse-grained, brachiopodal, sandy allochem limestone (MFT 8) are frequently occurring, well-rounded brachiopod bioclasts (light brownish colors due to complete silicification) and very coarse, sand-sized, single and composite quartz clasts (A). Note an intensely stained lithoclast (B) and a smaller fragment of a trepostome bryozoan (arrow). e Section H1/sample: 36.3, Vøringen Member: this brachiopodal, allochemic sandstone (MFT 8) displays a transitional microfacies into fine-grained, sandy allochem limestones to allochemic sandstones (MFT 9). It is characterized by a high proportion of sand-sized, edge-rounded quartz clasts (A), smaller brachiopodal bioclasts (B) and peloids. f Section H1/sample 32.3, Vøringen Member: smaller bioclasts of brachiopods (A), quartz grains (B) and peloids (C) form the main component types of well-sorted, fine-grained, sandy allochem limestones to allochemic sandstones (MFT 9). The matrix is completely silicified. g Section H1/sample 30.1: this picture shows an abrupt transition from a fine-grained, sandy allochem limestone (A, MFT 9), characterized by sand-sized quartz grains, peloids, and bioclasts, to a microspiculite (B, MFT 3) marked by abundant monazon microscleres embedded in a micritic matrix. h Section E1/sample 20.3: calcareous sandstones (MFT 10) occasionally to rarely show ruditic bioclasts (brachiopod A, bryozoan B) embedded in a sandy matrix. Note the cross section through a Skolithos burrow (C), in which peloids and smaller bioclasts are enriched
Interpretation: The occurrence of spiculites is usually connected to cooler, deeper marine environments, as cold water favors the preservation of biogenic opal and quiet-water conditions below the fair weather wave base (FWWB) allows the in-place deposition of the spicules. The bulk of monazon siliceous sponge spicules commonly originate from sponges of the group Demospongiae, which after their decay are often deposited in-place due to the disintegration of the skeleton. However, Gammon and James (2001) suggest from recent Demospongiae that fossil sponges also most likely have been formed in shallow-water, near-coastal environments. This assumption is confirmed by the local accumulations of sponge needles within the Gipshuken Formation, interpreted to reflect minor occurrences of siliceous sponges within nearshore, quiet-water areas of peritidal flats of the carbonate platform. The components are interpreted as heterozoan elements of a mixed heterozoan-reduced photozoan assemblage reflecting sub-tropical environmental conditions during the sedimentation of the upper part of the Gipshuken Formation.
Spiculites within the Vøringen Member correspond to the proximal subfacies of the spiculite facies of Hüneke et al. (2001) and to the light-colored spiculites of Ehrenberg et al. (2001). The authors interpret the facies to represent low energy, open shelf areas generally below the storm weather wave base (SWWB). As this interpretation mainly refers to occurrences of massive spiculites from overlying chert horizons of the Kapp Starostin Formation, we see the rare occurrence of spiculitic mudstones in the Vøringen Member as the proximal reaches of this facies into shallow submarine, near-coastal areas. The parallel orientation of the megascleres and their restricted occurrence within small, protected intergranular spaces (burrows) or intragranular voids (umbrella shelter) implies current or wave activity and generally points to higher energy conditions across the open-marine ramp areas.
MFT 4: bioclastic, peloidal wacke- to packstones (Fig. 8e, f)
Description: This MFT comprises poorly to moderately sorted, packed wackestones, packstones and more seldom grainstones (float- to rudstones), forming up to ca. 20-cm-thick beds, discontinuous horizons or lenses, which are present within isolated outcrops several meters below the sections at Eremitten and Hódbreen. The MFT contains various bioclasts and peloids as the main grain categories. The components, which show no preferred orientation, mainly consist of fragments of echinoderms, gastropods, brachiopods, Tubiphytes, partly ruditic debris of trepostome and fenestrate bryozoans and peloids. Staffellid (Pseudoendothyra sp.) foraminifers, chaetetids, poorly preserved dasycladaceans (Velebitella simplex Kochansky-Devidè 1964) and ostracodes occur only locally in minor amounts. Commonly occurring peloids are irregularly shaped and variably sized, edge-rounded to rounded and formed of dense micrite, often without any internal fabric, thus most likely originating from the disintegration and abrasion of algal mats (algal peloids). Rarely, single Microcodium and dolomite crystals occur. The matrix is mainly composed of grey, partly reddish or brownish colored micrite/microsparite, with blocky sparite in some well-washed areas.
Interpretation: This MFT is characterized by a partly diverse component composition, comprising both heterozoan (crinoids, brachiopods, bryozoans) as well as photozoan elements (Tubiphytes, dasycladacean algae). A similar biotic composition has been observed within the Canadian Arctic (Sverdrup Basin) by Reid et al. (2007) and Bensing (2007) within the late Sakmarian to earliest Artinskian Raanes Formation, representing inner- to mid-ramp areas. Here, bryozoans, brachiopods and siliceous sponge spicules are interpreted as mid-ramp elements immigrating into inner shelf areas due to an underlying cooling from warm to subtropical (warm temperate) conditions. The occurrence of this MFT probably reflects the same development in Svalbard. Bioclasts from more open-marine and probably cooler environments further offshore were imported into intertidal to shallow-subtidal areas of the platform interior, which was still warm-water to this point of time. Under moderate energy conditions (tides, waves) the components were washed together into lateral restricted accumulations, which combine both photozoan warm-water elements from the inner platform and heterozoan biota from the mid-platform.
MFT 5: Microcodium facies (Fig. 8g, h)
Description: This microfacies comprises both allochthonous and autochthonous Microcodium deposits. Both types are characterized by the occurrence of equant to bladed, sparry calcite crystals, which commonly show light brownish to yellowish colors and have subhedral shapes.
Within allochthonous Microcodium deposits, edge-rounded crystals mainly occur as isolated grains, rarely to occasionally constituting smaller composite radial-structured, petal-like aggregates. The irregularly distributed components are commonly embedded in grey or brownish micritic/microsparitic matrixes, forming low diverse, well-sorted wackestones to packstones. Ruditic, often intensely Fe-stained carbonate nodules occasionally occur within this microfacies type. These roundish to irregularly shaped nodules also often show an internal Microcodium facies, comprising abundant Microcodium crystals embedded in a micritic matrix. They are often partly surrounded by desiccation cracks filled with blocky sparite.
More rare, autochthonous Microcodium deposits are characterized by the occurrence of smaller round to half-round aggregates and larger composite crusts consisting of single to multiple stacked layers of crystals, which are connected along their longitudinal axis. The crusts are commonly embedded in a micritic groundmass or occur within microbial mats, replacing the host sediment.
Interpretation: The occurrence of Microcodium has been frequently described within Late Paleozoic, marine carbonates (Kabanov et al. 2008). Although the specific formation is still under discussion (calcified mycorrhizae of root cells (after Klappa 1980) versus saprotrophic fungi and actinobacteria (after Kabanov et al. 2008), its non-marine, terrestrial nature is widely accepted. Thus, the occurrence of Microcodium clearly signals the cessation of marine conditions connected with a prolonged subaerial exposure of marine deposits and the development of immature paleosols in a terrestrial setting. During this time, the deposits in the emerged platform areas were pedogenically neoformed, reflected by the formation of carbonate nodules and the growth of autochthonous Microcodium aggregates. Single crystals, the allochthonous disintegration products of the Microcodium mats, were coevally redeposited within adjacent, inter- and shallow subtidal areas of the carbonate platform.
MFT 6: carbonate breccias (Fig. 9a, b)
Description: This MFT forms massive, medium- to thick-bedded, multicolored beds with sharp, partly strongly wavy to irregular bedding planes. It also occurs within other limestones (mudstones and microbial limestones) in laterally restricted areas. Carbonate breccias are characterized by abundantly occurring, mm to several cm-sized intraformational lithoclasts displaying the only component type of poorly sorted, intraclastic rud- to floatstones. The angular to rounded and irregularly to roundish-shaped lithoclasts mainly show greyish and yellowish to brownish but also reddish and greenish colors. The components are generally strongly altered, i.e., stained, recrystallized, dolomitized and/or marginally dissolved and thus primary, internal fabrics corresponding to mudstones, microbial limestones or Microcodium facies are mostly obliterated. The intraclasts are irregularly distributed, resulting in component- as well as matrix-supported fabrics. The matrix consists of often intensely stained, dolomitized or recrystallized, fine-grained sediment or of several generations of blocky sparite. Locally, smaller interparticle voids are also filled by multiple generations of reddish micrite coatings.
Interpretation: Intraclastic carbonate breccias within the uppermost part of the Gipshuken Formation originate from the prolonged emersion of the platform strata during periods of low sea level. Subjected to a supratidal sabkha environment, already lithified peritidal platform deposits developed into carbonate breccias due to erosion, reworking and redeposition. Time of exposure, alteration and transport length of the components varies strongly; locally, in-place brecciation and only little transport indicated by the good fitting of angular components resulted in the formation of component-supported collapse breccias. In other places, rounded intraclasts show different colors and multiple generations of fragmentation, recrystallization and dolomitization, implying longer transport and intense subaerial alteration and karstification. Various models, such as the seepage-reflux model or the evaporation-drawdown model (Flügel 2004) propose dolomitization connected to the emersion of marine limestones, which seems to be most likely, considering the depositional setting at the top of the Gipshuken Formation.
MFT 7: coarse-grained, mixed-bioclastic, sandy allochem limestones (Fig. 9c)
Description: This MFT comprises grey to slightly brownish, thick- to medium-bedded, partly sandy allochemic limestones (after Mount 1985), which are coarse-grained, moderately to poorly sorted and characterized by a typical heterozoan biotic assemblage. The main constituents of these skeletal rud- to floatstones are formed of frequently to commonly occurring fragments of brachiopods (mainly spiriferid and productide shells and spines), bryozoans (trepostome and fenestrate) and echinoderms (mainly crinoids). Up to several-cm-large chaetetid colonies, gastropod and bivalve shells are locally present in minor amounts as well as brownish, strongly altered carbonate lithoclasts (extraclasts) and smaller peloids with internal mudstone fabrics. Locally, abundant sponge spicules are present within smaller, poorly washed pockets or umbrella-structures of brachiopod valves. Authigenic glauconite minerals rarely occur within central canals of megaspiculae and other intraskeletal cavities. The loosely to densely packed components have no preferred orientation and are irregularly distributed within the sediments, which locally show cross-bedding. In poorly washed areas, the matrix is composed of partly stained micrite/microsparite as well as sand-sized, edge-rounded detrital quartz grains or accumulations of unidentifiable, fine-grained skeletal fragments (filaments). However, most of the inter- and intra-particle spaces are cemented by blocky sparite or by the syntaxial overgrowth of echinoderm fragments. Particularly brachiopod shells are often silicified. Low-relief stylolites occur commonly on the sharp, slightly wavy bedding planes.
The allochthonous sediments locally embed frequently occurring, in situ bioherms formed of ramose, trepostome bryozoans (sections S1, H1; Fig. 4e). The erect, up to 50-cm-high and up to 1-m-wide, rigid contiguous colonies show bush-like growth forms due to very frequent bifurcations in three dimensions of up to 2 cm thick, laterally unconnected branches (bryozoan growth habit classification of Hageman et al. 1998).
Interpretation: The facies corresponds best to the bryozoan limestone facies (FA-2) of Ehrenberg et al. (2001) and the crinoid-bryozoan facies of Hüneke et al. (2001). The highly fossiliferous deposits comprise a fully heterozoan (bryonoderm) biotic assemblage, represented by brachiopods, bryozoans, echinoderms and minor siliceous sponges and thus is interpreted to represent temperate marine conditions marked by cooler water temperatures and probably increased nutrient levels. The coarse-grained character of the deposits, cross-bedding and the common presence of blocky sparite within inter- and intra-particle voids suggest sedimentation under high-energy conditions probably due to storm events on an open-marine, shallow shelf. The composition of the non-biotic components reveals two different source areas. Occasional carbonate extraclasts originate from pre-existing, lithified platform strata of the underlying Gipshuken Formation and reflect erosion and reworking of emerged carbonate platform areas, described from central Spitsbergen (Ehrenberg et al. 2001; Groen 2010). Coevally, high amounts of detrital, sand-sized quartz grains reflect increased terrigenous input from a terrestrial source area, supposedly in the vicinity of the investigation area.
MFT 8: coarse-grained, brachiopodal, sandy allochem limestones to allochemic sandstones (Fig. 9d, e)
Description: This MFT is characterized by massive, medium- to thick-bedded, coarse-grained, well- to poorly sorted shell accumulations or pavements mainly consisting of whole biogens and bioclasts of thick-shelled, punctate and impunctate brachiopods (commonly spiriferids and productids, more seldom terebratulids, arthyrids, rhynchonellids, strophomenids). The abundantly to frequently occurring skeletal fragments are edge-rounded to well-rounded, and often show a more or less parallel orientation and well-preserved internal microstructures due to their primary low-Mg calcite skeletal substance. Fragments of thin-shelled, punctate brachiopods and spines are also present in minor amounts, as well as occasional to rarely occurring bioclasts of bivalves, gastropods, trepostome bryozoans and echinoderms (mainly crinoids). Greenish to brownish, strongly altered and Fe-stained lithoclasts with an internal mudstone fabric are generally rare and occur only locally. Within poorly washed areas, the components are embedded in matrixes composed of various materials, consisting of medium to coarse sand-sized, edge-rounded to rounded quartz grains, fine-grained skeletal fragments (filaments from brachiopods and ostracodes) or abundant megaspiculae of siliceous sponges embedded in micrite/microsparite (spiculites). The remaining pore spaces are commonly cemented by one generation of blocky or syntaxial cements. Skeletal fragments and the matrix are locally strongly silicified and comprise opaque mineral separations. Authigenic glauconite minerals are occasionally present mainly within intraskeletal voids (central canal of megascleres, brachiopod biogens). The sharp and slightly wavy bedding planes are locally marked by low-relief stylolites and occasional show scours on the subsurfaces. Graded bedding is locally developed.
Interpretation: This MFT corresponds to the brachiopod facies of Hüneke et al. (2001) and the brachiopod limestone facies (FA-1) of Ehrenberg et al. (2001). It reflects extensive, sandy brachiopod shell banks (lumachelles, coquinas) within open-marine, nearshore to transitional offshore areas of the inner, mixed siliciclastic-carbonate ramp. High-energy conditions are reflected by the mainly sparitic matrix of the deposits, coarse grain-size and high degree of abrasion (roundness) of the skeletal fragments. Partly erosional subsurfaces, graded bedding and the association with fine-grained, sandy allochem limestones to allochemic sandstones (MFT 9) suggest that the deposits represent proximal storm beds (tempestites; Aigner 1985), comprising amalgamated and composite shell layers. Forming the lower, thick-bedded, bioclast-dominated and coarse-grained part of recurrent sediment couplets, the brachiopodal deposits record periodic storm events, during which the skeletal material was reworked on sandy flats between the FWWB and SWWB.
MFT 9: fine-grained, sandy allochem limestones to allochemic sandstones (Fig. 9e–g)
Description: This MFT comprises medium-bedded, massive, mixed siliciclastic-carbonate deposits with packstone and grainstone fabrics. The generally well-sorted sediments are intensely bioturbated, showing abundant Skolithos burrows oriented perpendicular to the bedding planes (piperock; Droser 1991). They include rather loosely packed, fine- to medium-arenitic skeletal fragments (filaments), peloids and angular to edge-rounded, fine-arenitic quartz grains in more or less equal amounts as main component categories. The elongated filaments show either a primary calcitic, fibrous microstructure or are strongly micritized. They probably represent the fine-grained debris of brachiopods, from which larger bioclasts are also occasionally present. Occasional to rarely occurring echinoderm, bryozoan (trepostome and fenestrate) and other completely recrystallized skeletal fragments form minor component types. The equally distributed components show no preferred orientation and are preferably enclosed by one generation of blocky sparite. In poorly washed areas and inside the abundantly occurring Skolithos burrows, a microsparitic, and more seldom a microspiculitic matrix is present. Rhombic pyrite crystals of up to 2 cm in size as well as brownish mineral separations and greenish glauconite minerals occur occasionally to rarely. Whitish colors in these otherwise commonly greyish to greenish sediments reflect intense silicification.
Interpretation: Skolithos burrows have been interpreted as a feature of shallow submarine, nearshore areas in moderate- to high-energy environments (Buatois et al. 2002; Palma et al. 2007), which is in agreement with the general depositional setting of the Vøringen Member. In connection with tempestites, piperocks are mentioned by Vossler and Pemberton (1988) and Droser (1991), comprising nearshore sandstones with abundant Skolithos burrows. According to the authors, the inchnofacies formed due to opportunistic sea-bottom feeders, which colonized the sand subsequent to storm events. As the upper unit of the characteristic sediment couplets, the deposits are interpreted to constitute the normal background sedimentation during time periods without storm activity. The packstone and grainstone fabrics reflect agitated to moderate water conditions between the FWWB and SWWB, in which the sand-sized particles (quartz grains, peloids and bioclasts) were constantly winnowed and well-sorted due to waves or/and currents.
MFT 10: calcareous sandstones (Fig. 9h)
Description: This MFT comprises massive, generally well-washed and -sorted, densely packed and partly highly porous sandstones. Commonly to rarely occurring ruditic skeletal fragments and biogens are embedded in a matrix of very well-sorted, edge-rounded to rounded, single crystal, sand-sized quartz grains with occasional to rare peloids. The biota mainly comprises thin- and thick-shelled brachiopods, and trepostome bryozoans and crinoids to a minor extent. Elongated bioclasts and whole shells show a preferred orientation parallel to discontinuous, slightly wavy bedding planes. Ruditic bioclasts and sand-sized quartz grains are cemented by one generation of blocky sparite. Microsparite occurs mainly in frequently occurring burrows, dominated by the spreiten structure Zoophycos but also comprising mainly vertical Skolithos burrows. The sediments show varying degrees of silicification affecting skeletal fragments as well as calcite cements. Mineral separations and glauconite minerals occur occasionally to rarely.
Interpretation: Calcareous sandstones correspond to glauconitic sandstones (FA-7) of Ehrenberg et al. (2001). The facies imply a high siliciclastic input from an adjacent terrestrial source area into the shallow-marine realm and are interpreted to represent widespread sand flats within the nearshore areas of the inner ramp. Constant reworking and winnowing due to tidal currents and/or wave action above the FWWB led to the well sorting and selective concentration of quartz grains. An intense bioturbation by bottom feeders is reflected by abundant burrows, dominated by Zoophycos. Within the Mesozoic and Cainozoic, Zoophycos is generally considered a characteristic trace fossil for bathyal to abyssal settings. In the Late Paleozoic Kapp Starostin Formation, however, the occurrence of Zoophycos has been interpreted as indication of shallow-water deposition above SWWB (Bromley et al. 1989) and of increased trophic resources (Reid et al. 2007).
Characteristic greenish colors of the calcareous sandstones most likely result from an increased proportion of glauconite, whose formation is usually connected to low sedimentation rates and deeper marine, quiet-water conditions. As a synsedimentary formation within the suggested nearshore depositional environment seems to be inappropriate, we suggest a post-sedimentary origin of glauconite during the sedimentation of the overlying dark shale horizon capping the entire Vøringen Member. In our view, the deposition of the black shales indicates the termination of the shallow-marine, inner ramp sedimentation and reflects outer ramp sedimentation characterized by water depths far below the SWWB. During this period, low sedimentation rates and quiet water conditions prevailed within the depositional area, suitable for the growth of glauconite within the open pore spaces of the sandstones.
Facies pattern of the investigated strata
Both the strata of the Gipshuken Formation (Templet and Sørfonna members) and the Kapp Starostin Formation (Vøringen Member) show a certain pattern of the different MFTs, forming characteristic facies associations and sequences, which in turn reflect a combination of processes within specific depositional settings and environments (Nichols 1999).
The carbonates of the Gipshuken Formation comprise two main microfacies associations. Microbial limestones (MFT 1) are often associated with mudstones (MFT 2) and rarely occurring spiculites (MFT 3) due to their formation in a peritidal platform environment. In addition, Microcodium facies (MFT 5) are commonly associated with carbonate breccias (MFT 6) caused by the subaerial exposure and alteration of the marine strata in a supratidal sabkha environment. However, as the supratidal facies association emanate from the peritidal sediments, the MFTs of both associations often occur together, reflecting multiple exposure events during sedimentation in a marginal marine depositional setting.
The MFTs of the Vøringen Member are arranged into three characteristic associations. At the base, allochthonous, coarse-grained, mixed-bioclastic, sandy allochem limestones (MFT 7) and calcareous sandstones (MFT 10) embed locally occurring, in situ bryozoan bioherms, which used consolidated hardgrounds and condensed surfaces as growth substrate. Upwards in strata, autochthonous buildups are absent and the proportion of bryozoan debris gradually decreases within the allochthonous sediments. These commonly form distinct sediment couples consisting of coarse-grained brachiopodal, sandy limestones as lower member (MFT 8) and fine-grained, sandy allochem limestones to calcareous sandstones (MFTs 9, 10) as upper member. The stacked sediment couplets are interpreted as tempestites reflecting a storm-dominated, shallow-marine depositional environment. With the further decrease of bioclasts in the upper part of the Vøringen Member, the strata is characterized by the third association, mainly formed of calcareous sandstones (MFT 10), interbedded with occasional occurring, sandy brachiopodal limestones. All three facies associations are arranged in a specific order thus forming a facies sequence, which reflects a temporal change of the depositional processes and environments, discussed in the next chapter.
Discussion
Depositional environments
Combining outcrop information with the detailed microfacies studies reveals two completely different depositional settings, which prevailed during the sedimentation of the Templet and Sørfonna members (Gipshuken Formation) and the overlying Vøringen Member (Kapp Starostin Formation).
Gipshuken Formation
The facies of the investigated strata of the Templet and Sørfonna members display a typical marginal marine setting of a carbonate platform (Fig. 10a).

Reconstruction of the depositional environments during the sedimentation of the Templet and Sørfonna members (a) and the Vøringen Member (b). The occurrences of the specified microfacies types (MFTs 1–10) are displayed in both subfigures. Legend in Fig. 3
Within supratidal sabkhas, subaerially emerged, marine limestones were affected in varying degrees by desiccation (cracks and fragmentation), meteoric alteration (staining, micritization, recrystallization, dissolution), dolomitization, karstification, erosion, reworking and pedogenesis. Carbonate breccias (MFT 6) formed due to the in situ fragmentation of subaerially exposed carbonates or after transport due to redeposition of strongly altered lithoclasts. Weakly developed paleosols and vegetation are reflected by Microcodium facies (MFT 5) and the occurrence of coaly plant fragments.
In a seaward direction, peritidal to shallow subtidal areas of an inner carbonate platform are reflected by a variety of facies types. Restricted marine, possibly hypersaline conditions within these areas are displayed by a low diversity of the biota. The latter is mainly represented by abundantly occurring, thick algal mats (MFT 1), originating from microbial organisms which populated intertidal and shallow subtidal platform zones. Within these areas, low-energy conditions led to the accumulation of fine-grained suspended matter, resulting in the formation of mudstones (MFT 2). The fine-grained sediments locally contain silt- to sand-sized detrital quartz grains, reflecting the proximity to a terrestrial, siliciclastic source area, from which generally sparse siliciclastic material was imported into the depositional area. Local accumulations of siliceous spiculae (MFT 3) and fecal pellets reflect the rare presence of siliceous sponges and bottom feeding organisms within these nearshore quiet-water areas. Laterally restricted accumulations of algal peloids, Tubiphytes, ostracodes, staffellid foraminifers and fragments of dasycladacean algae, gastropods, chaetetids, brachiopods, echinoderms and bryozoans, partly imported from more open-marine platform areas further offshore, led to the formation of bioclastic, peloidal wacke- to packstones (float- to rudstones, MFT 4). Within rare, agitated-water areas, grainstones were formed from these components.
Gipshuken/Kapp Starostin boundary
Evidence for a prolonged hiatus due to the subaerial exposure of the Gipshuken carbonate platform during the Artinskian is apparent and displayed by a number of facies and features within the uppermost strata of the formation across Svalbard. Keilen (1992) and Groen (2010) describe the occurrence of caliche, root traces and Microcodium in central Spitsbergen, while accumulations of plant fragments in certain horizons of the Templet member in NE Spitsbergen and coaly interbeds from the Sørfonna member on Nordaustlandet are reported in Dallmann et al. (1999). All these features suggest the development of paleosols, which require a prolonged hiatus in sedimentation and a sufficient period of geomorphic stability. Carbonate breccias and low-relief, karstic exposure surfaces described from the uppermost part of the Gipshuken Formation in central Spitsbergen (Lauritzen 1981a; Samuelsberg and Pickard 1999; Groen 2010) and from NE Svalbard (MFT 6; this study) also reflect significant breaks in sedimentation.
In addition to a hiatus connected to subaerial exposure, an unconformity caused by erosion of the uppermost strata of the Gipshuken Formation with the transgressive onset of the Vøringen Member sedimentation during the late Artinskian developed across extended parts of Svalbard (Dallmann et al. 1999). The latter is documented by the occurrence of strongly altered carbonate lithoclasts (extraclasts from the Gipshuken Formation) described from the Vøringen Member in central Spitsbergen (Ehrenberg et al. 2001; Groen 2010) and to a minor extend from NE Svalbard (MFTs 7, 8; this study).
In sections H1 and S1 however, the boundary itself is formed by an intensely Fe-stained and bored hardground, respectively black laminated claystone horizon, which were subsequently used as growth substrate by local bryozoan colonies. The horizons imply a pre-existing, already lithified platform surface, reflecting local sediment-starved conditions and condensation with the onset of the Vøringen Member transgression. Due to the preservation of the transitional strata, the boundary in this area is interpreted as an unconformity (paraconformity) formed by non-deposition instead of erosion. Submarine exposure in the Artinskian is also reported by Ehrenberg et al. (2001) in coeval strata from the southern Barents Sea (Bjarmeland Group; Larssen et al. 2005). Here, pyritized, glauconitic surfaces at the top of the Unit L-8, which comprises the Polarev and/or Isbjorn formations, may indicate contemporaneous non-deposition and condensation in a general deeper environmental setting. However, insufficient age constrains prevent a precise correlation of these condensed horizons with the Gipshuken/Kapp Starostin formations boundary of Svalbard.
Ehrenberg et al. (2001) assume that the hiatus at the Gipshuken/Kapp Starostin boundary comprise a time period of uncertain magnitude from less than 1 to maximum 15 Ma within the Early Permian. According to age datings at the top of the Gipshuken Formation (Sakmarian–early Artinskian?) and at the base of the Vøringen Member (late Artinskian–early Kungurian), the gap in the sedimentary record is generally restricted to the Artinskian, reducing the maximum time span to roughly 9 Ma (Fig. 2). In consideration of the period needed to develop a hardground shown in section H1, a much shorter hiatus (far below 1 Ma years) based solely on non-deposition and condensation is suggested. However, due to the erosional removal of the uppermost Gipshuken strata in other areas of Svalbard, the gap in the sedimentary record probably varies substantially and comprises much longer time periods, locally.
Kapp Starostin Formation
The sedimentary record of the Vøringen Member represents a fundamentally different depositional setting than the underlying Templet and Sørfonna members. The facies reflect open, shallow-submarine, nearshore to transitional offshore areas of an open-marine, storm-dominated ramp, characterized by mixed siliciclastic-carbonate sediments and a heterozoan biotic association (Fig. 10b).
In all three sections, the strata of the Vøringen Member allow an arrangement into three facies associations, which in turn form a facies sequence reflecting an overall shallowing-upward of the depositional area subsequent to an initial transgression.
At the base, consolidated hardgrounds and condensed surfaces (sections H1, S1) reflect sediment-starved conditions with the beginning of the Vøringen Member sedimentation in this particular area of Svalbard. The horizons comprise a certain time period prior to the start-up of the bryozoan buildups, during which the necessary requirements (e.g., water depth, temperature, nutrient level) developed. With the reach of favorable conditions, bryozoan bioherms (see MFT 7) locally appeared. Although the occurrence of bioclasts from trepostome bryozoans is commonly reported in the lowermost beds of the Vøringen Member from different localities in Spitsbergen (Winsnes 1966; Fredriksen 1988; Nakrem 1988, 1994a, b; Ehrenberg et al. 2001), it is the first time that in situ occurrences are documented on Svalbard. We suggest that these colonies formed local “reef flats or mounds” at the offshore transition from the inner to the mid ramp around the SWWB (Fig. 10b).
Coeval occurrences of bryozoan facies have been described from the Barents Sea (Finnmark Platform), North Greenland and Arctic Canada by Bruce and Toomey (1992), Ehrenberg et al. (2001; bryozoan limestone, FA-2), Stemmerik (1997; crinoid-bryozoan grainstone, F 3 and bryozoan rudstones, F 4), Beauchamp (1994) and Beauchamp and Desrochers (1997; bryonoderm carbonates), indicating a period of important reef growth within the shelf areas along the northern margin of Pangea. The authors describe coarse-grained, allochthonous deposits and isolated or composite bryozoan build-ups with partly significant vertical thickness, forming either offshore shallow-water banks or mounds or shoreline-attached platform margins at the transition to deeper basins.
The colonies are embedded by allochthonous coarse-grained, mixed-bioclastic, sandy limestones (MFT 7) mainly comprising fragments of various bryozoans, brachiopods and echinoderms, mixed together with strongly changing amounts of detrital quartz and rare extraclasts from the underlying Gipshuken strata. The coarse-grained character, poor sorting and mostly sparitic matrix of the embedding, allochthonous deposits as well as the robust growth form of the sediment bafflers themselves (Nakrem 1994a) imply high-energy conditions, probably to repeatedly occurring storm events. The latter are recorded by two separate bioherm horizons within the individual sections, reflecting repeatedly changing conditions from sediment-starved, low energy enabling the formation of hard substrates, to high-energy with a high terrestrial input during storm events.
The basal sediments are overlain by a succession of coarse-grained, brachiopodal, sandy allochem limestones to allochemic sandstones (MFT 8), interbedding with intensely bioturbated, fine-grained, sandy allochem limestones to allochemic sandstones (MFT 9) and calcareous sandstones (MFT 10). These deposits form stacked sediment couplets, which are interpreted to reflect proximal, composite and/or amalgamated tempestites due to repeatedly occurring storm events within open-marine, nearshore to transitional offshore areas of the inner ramp between the FWWB and SWWB (Fig. 10b). The latter were marked by low-relief, shallow submarine sand flats and sandy brachiopodal shell banks (coquinas, lumachelles) following the bryozoan reef zone in a general more proximal, landward position.
The lower, coarse-grained member of the couplets are interpreted to reflect storm events, during which unconsolidated sediment material and pre-existing, already lithified carbonates of the underlying Gipshuken Formation were eroded, reworked and redeposited under high-energy conditions. Storm-induced deposits are discussed by Aigner (1985) and Immenhauser (2009) describing similar features such as erosive subsurfaces, graded bedding and poor sorting.
In addition to sedimentary features directly connected to storms, Vossler and Pemberton (1988) and Droser (1991) indicate increased biotic activity of burrowing organisms in the aftermath of individual storm events. The latter is reflected by the occurrence of so-called piperocks, i.e., nearshore sandstones characterized by abundant Skolithos burrows on top the actual storm layers. This storm-related feature is in perfect agreement with the upper finer member of the couplets, supposed to reflect intense bioturbation and common background sedimentation under moderate energy conditions during periods without storm activity.
Further upwards in strata, the recurrent sediment couplets gradually pass into a more uniform, thick succession of glauconitic, calcareous sandstones (MFT 10) with only occasionally interbedded, thin brachiopod shell horizons. In our opinion, this facies association presumably represents the most proximal depositional areas, namely broad sand flats of nearshore to shoreface zones around the FWWB. Here, mainly detrital sediment material (quartz grains) originating from a nearby terrestrial source area was constantly winnowed and reworked due to the activity of waves and tides and exported onto more distal ramp areas during storm events.
All three sections show a high siliciclastic input with the onset of the Vøringen Member sedimentation, reflected by the occurrence of sandy limestones (MFTs 7–9) and calcareous sandstones (MFT 10; Figs. 3, 6, 7). In contrast, limestones of the Vøringen Member in central Spitsbergen (Billefjorden, Dicksonfjorden; Groen 2010) contain only very little detrital quartz and calcareous sandstones don’t occur at all. The latter suggests the uplift of a terrestrial source area to the N or E of the investigation area in NE Svalbard. Correlating the individual sections, S1 (Fig. 7) shows the smallest thickness of the Vøringen Member (ca. 11 m; H1: ca. 21 m; E1: ca. 18 m) combined with the thickest proportion of the calcareous sandstone facies, implying that this section might have been closest to a source area, possessing the least accommodation space (reflected in the smallest thickness) and the biggest proportion of nearshore sand shoals.
Environmental change within the Early Permian
Sediments of the Gipshuken Formation (late Sakmarian–early Artinskian) in central Spitsbergen reflect a rhythmic, probably sea-level driven deposition of stacked evaporite/carbonate cycles within extended, low-relief, inner platform areas, comprising supratidal sabkhas and peritidal, restricted marine lagoons (Lauritzen 1981a). Some features such as the decrease of evaporites in the carbonate-dominated upper part in central Spitsbergen (Skansdalen member) and the occurrence of plant fragments, paleosols and coaly interbeds in the Templet and Sørfonna members in NE Svalbard, imply a progressive change of the continental climate from arid or semi-arid to more humid subtropical conditions (Keilen 1992). The latter also may account for a low but first-time-recognizable siliciclastic input in the Templet member within the investigation area, resulting from an increased, possibly seasonal, monsoonal-like precipitation combined with an intensified weathering of the terrestrial hinterland.
A progressive environmental change within the marine realm during the sedimentation of the Gipshuken Formation (Templet and Sørfonna members) and overlying Kapp Starostin Formation (Vøringen Member) is revealed by correlating the biota of equivalent strata from central Spitsbergen, NE Svalbard and the southern Barents Sea under consideration of their environmental setting.
In central Spitsbergen, a reduced photozoan biotic association with no heterozoan elements have been described from the uppermost Gipshuken strata (Groen 2010). During this time period, a marginal marine platform setting, marked by restricted, inner platform areas and subaerial sabkhas prevailed (Lauritzen 1981a, b). Contemporaneously, more open-marine conditions existed within NE Svalbard, reflected by a diverse, composite biofacies, characterized by reduced photozoan as well as heterozoan elements (Keilen 1992; MFTs 4, 3, this work). Equivalent strata from the southern Barents Sea (Bjarmeland Group; Larssen et al. 2005) reflect a general deeper, open-marine, outer platform setting marked by a typical temperate-water, heterozoan biotic association consisting mainly of crinoids, bryozoans, brachiopods and siliceous sponges (Stemmerik 1997). The lower part of the Bjarmeland Group (Polarrev and Ulv formations, mid Sakmarian–early? Artinskian) is supposed to correlate with the Gipshuken Formation on Svalbard and shows a fully heterozoan association characterized by thick bryozoan- and bryozoan/Tubiphytes-dominated bioherms (Polarrev Formation) on top or along local highs around the SWWB.
The distribution pattern of the biota shows that while warm-water conditions still prevailed in inner platform areas in central Spitsbergen, temperate-water conditions were already present in deeper depositional settings in the southern Barents Sea, with NE Spitsbergen representing a transitional position during this period. This interpretation is in accordance with Bensing (2007) and Reid et al. (2007), who describe similar biotic assemblages within coeval deposits in the Canadian Arctic (Sverdrup Basin, upper Raanes Formation), interpreted as transitional from warm-water to slightly cooler, subtropical marine conditions. The authors assume that while a warm-water environment might still have prevailed within the inner peritidal platform zones, reflected by thick algal mats and the presence of Tubiphytes, dasycladaceans and staffellid foraminifers, cooler conditions represented by crinoids, brachiopods and bryozoans might have already been present within deeper submarine areas of the mid-platform.
With the transgressive onset of the Vøringen Member sedimentation, photozoan elements have disappeared completely, also in the innermost platform areas in central Spitsbergen, and are replaced by an entirely heterozoan biotic association (bryonoderm; Hüneke et al. 2001). In combination with highly diverse bryozoan assemblages, classified as boreal or northern according to Nakrem (1994a, b), and the occurrence of authigenic glauconite in sandy deposits, temperate climatic conditions within all ramp zones are reflected in the Svalbard area by now. Similar conditions prevailed within the southern Barents Sea. The Isbjørn Formation (mid? Sakmarian–late Artinskian) forming the upper part of the Bjarmeland Group is seen as the lateral equivalent of the Vøringen Member (Larssen et al. 2005) and is interpreted to represent inner areas of cool-water carbonate platforms (Stemmerik 1997). The strata also contain a fully heterozoan biotic assemblage comprised mainly of bedded limestones dominated by crinoids and bryozoan fragments.
The change from warm to temperate and eventually cold marine conditions on the shelves along the northern margin of Pangea during the Permian have been linked to climatic, paleoceanographic and regional paleogeographic changes connected to the overall northwards drift of the supercontinent (Steel and Worsley 1984; Worsley et al. 1986; Stemmerik and Worsley 1989; Beauchamp 1994; Nakrem 1994a; Cecchi et al. 1995; Beauchamp and Desrochers 1997; Stemmerik 1997; Ehrenberg et al. 2001; Hüneke et al. 2001; Beauchamp and Baud 2002; Bensing 2007; Reid et al. 2007).
Beauchamp and Baud (2002) assume that the decrease of the water temperature was mainly caused by changes in the oceanic circulation pattern (Fig. 11). According to the authors, this mechanism was a major factor for the onset of the Permian Chert Event (PCE), during which cold, nutrient-rich deep waters, connected with the seasonal melting of possible sea ice within northern high latitudes, were brought via oceanic upwelling onto the western and northern shallow shelf areas of Pangea. These paleoceanographic changes were possibly emphasized by regional paleogeographic changes in the course of the final consolidation of Pangea. The latter comprises the closure of the Uralian seaway (Fig. 11), which is assumed to have taken place latest in Kungurian times (Cecchi et al. 1995). The shutting down of such a major intercontinental oceanic gateway certainly would have caused fundamental environmental changes within the adjacent shallow seas, as warm-water masses from the Tethys were progressively cut off and the shelf areas along the northern margin of Pangea were increasingly influenced by northern, cold-water systems of Panthalassa.

Global (a) and regional paleogeographic (b) maps showing the continent/ocean configuration during the Early Permian. a The situation during the deposition of the upper part of the Gipshuken Formation (early Artinskian). Extended parts of Svalbard (encircled in red) are emerged and the Uralian seaway is wide open (source Ron Blakey: http://jan.ucc.nau.edu/~rcb7/paleogeographic.html). b A detailed reconstruction of the northern margin of Pangea during the sedimentation of the Vøringen Member (late Artinskian). Nearly entire Svalbard is flooded and the Uralian seaway is closed (modified after Reid et al. 2007). Note the newly emerged land to the NE of the investigation area. SB Sverdrup Basin, C Crocker Land
The mixed siliciclastic-carbonate deposits of the Vøringen Member are seen as the result of both a generally lower carbonate production, typical for such temperate-water carbonate systems (Schlager 2000), but also result from an escalating input of terrestrial, siliciclastic material into the depositional area. The remarkable increase in detrital quartz with the onset of the sedimentation of the Vøringen Member points to changes in the local paleogeography of Svalbard, in particular to the development of a new siliciclastic source area in the vicinity of the investigation area. In connection with more temperate and humid continental conditions, increased weathering, precipitation and run-off provide a plausible explanation for the sudden increase in continental silicia within the Vøringen Member strata in NE Svalbard. Additionally to an increased siliciclastic input, more nutrients would have been imported into the marine realm. Combined with the proposed upwelling of cold, nutrient-rich deep waters onto the northern margin of Pangea, these processes might have led to elevated nutrient levels. Terrestrial input, water temperature as well as nutrient level are considered as the major controlling factors for the environmental change and biotic turnover along the northern margin of Pangea within the Early Permian (Bensing 2007; Reid et al. 2007).
Conclusions
Within NE Svalbard, the upper strata of the Gipshuken Formation (Templet and Sørfonna members, Sakmarian–early Artinskian?) represent a typical marginal marine, low-energy carbonate-platform setting. Sediments mainly consist of thick microbial limestones (algal mats, MFT 1) and mudstones (MFT 2) reflecting restricted-marine, peritidal areas of an inner carbonate platform. Commonly to abundantly occurring Microcodium facies (MFT 5), carbonate breccias (MFT 6) and local accumulations of coaly plant fragments represent supratidal sabkhas, on which the marine strata were affected by desiccation, meteoric alteration, karstification, and pedogenesis during subaerial exposure. However, rarely occurring fossiliferous wacke- to grainstones (MFTs 3, 4), comprising a mixed heterozoan/reduced photozoan assemblage, indicate the import of heterozoan biota from deeper, open-marine, mid-platform areas in the vicinity of the investigation area. These platform areas most likely were marked by temperate water conditions already.
With the onset of the sedimentation of the Vøringen Member (late Artinskian?–Kungurian), a basal unconformity formed due to the previous erosion/condensation of the uppermost Gipshuken strata.
The sedimentary succession of the member is marked by a fully heterozoan biotic association reflecting temperate water conditions within open-marine, storm-dominated, mixed carbonate-siliciclastic, inner ramp areas by now. Three main facies associations are pointed out, forming a shallowing-upward sequence, subsequent to an initial transgression.
At the base, locally occurring, in situ bryozoan bioherms, which used consolidated hardgrounds as growth substrate, represent transitional offshore areas to the mid-ramp around the SWWB. While the hard grounds imply submarine exposure and sediment-starved conditions probably below the SWWB prior to the bryozoan growth, coarse-grained, poorly sorted, mixed-bioclastic, sandy limestones (MFT 7) embedding the bioherms, reflect rapid, storm-dominated sedimentation.
Above, coarse-grained brachiopodal, sandy limestones to allochemic sandstones (MFT 8) and fine-grained, sandy allochem limestones to allochemic sandstones (MFTs 9, 10) form characteristic sediment couplets, repeatedly deposited onto each other. These sediments are interpreted to represent proximal tempestites deposited on open-marine, nearshore to offshore, inner ramp areas between the FWWB and SWWB, comprising broad sand flats and sandy brachiopodal shell banks.
Calcareous sandstones (MFT 10) with occasional interbedded brachiopodal, sandy limestones are present in the upper part of the Vøringen Member, representing proximal sand shoals within nearshore areas around the FWWB.
The environmental change recorded within the transitional strata of the Gipshuken and Kapp Starostin Formations in Svalbard has been documented in various other locations comprising coeval shelf strata along the northern margin of Pangea (e.g., southern Barents Sea, eastern North Greenland, Arctic Canada, Russia). Thus, the environmental development is regarded as a regional phenomenon, which shows a successive progression of temperate- and cold-water conditions across the shallow seas. The change has been attributed to a number of controlling factors such as water temperature, respectively depth, nutrient and energy levels as well as siliciclastic input, which in turn are linked to large-scale climatic, paleoceanographic and paleogeographic changes connected to the northwards drift of Pangea. Decreasing water temperatures are probably caused by the upwelling of cold, nutrient-rich deep waters of Panthalassa, supported by the closure of the Uralian gateway, cutting off warm-water masses from the Tethys. Higher nutrient and energy levels as well as increased siliciclastic input are probably due to more humid climatic conditions marked by increased precipitation and storm activity combined with the uplift of a local siliciclastic source area to the N or E of the investigation area.
References
Aigner T (1985) Storm depositional systems. Lecture Notes in Earth-Science 3. Stuttgart, Germany
Beauchamp B (1994) Permian climatic cooling in the Canadian Arctic. In: Klein G (ed) Pangaea: paleoclimate, tectonics, and sedimentation during accretion, zenith, and breakup of a supercontinent. Geol Soc Am Spec Paper 288, pp 229–246
Beauchamp B, Baud A (2002) Growth and demise of Permian biogenic chert along northwest Pangaea: evidence for end-Permian collapse of thermohaline circulation. Palaeogeogr Palaeoclimatol Palaeoecol 184:37–63
Beauchamp B, Desrochers A (1997) Permian warm- to very cold-water carbonates and cherts in Northwest Pangaea. In: James NP, Clarke JAD (eds) Cool-water carbonates, SEPM Spec Publ 56. Tulsa, Oklahoma, USA, pp 327–347
Bensing J (2007) An Early Permian subtropical carbonate system: sedimentology and diagenesis of the Raanes and Great Bear Cape Formations, Sverdrup Basin, Arctic Canada. MSc thesis, Queen’s University, Kingston, Ontario, Canada
Biernat G, Birkenmajer K (1981) Permian brachiopods from the base of the Kapp Starostin Formation at Polakkfjellet, Spitsbergen. Studia Geologica Polonica 73:7–24
Birkenmajer K, Czarniecki S (1960) Stratigraphy of marine Carboniferous and Permian deposits in Hornsund (Vestspitsbergen), based on brachiopods. Bull Pol Acad Sci Earth Sci 8:203–209
Blomeier D, Scheibner C, Forke H (2009) Facies arrangement and cyclostratigraphic architecture of a shallow-marine, warm-water carbonate platform: the Late Carboniferous Ny Friesland Platform in eastern Spitsbergen (Pyefjellet Beds, Wordiekammen Formation, Gipsdalen Group). Facies 55:291–324
Bromley R, Hanken N-M, Asgård U, Fredriksen K, Henriksen L (1989) Shallow-water Zoophycos assemblages, Upper Permian, Svalbard. Int Geol Congr 28:204–205
Bruce JR, Toomey DF (1992) Late Palaeozoic bioherms occurrences of the Finnmark Shelf, Norwegian Barents Sea: analogues and regional significance. In: Vorren TO, Bergsager E, Dahl-Stamnes ØA, Holter E, Johansen B, Lie E, Lund TB (eds) Arctic geology and petroleum potential, NPF special publication Nr 2. Elsevier, Amsterdam, pp 377–392
Buatois LA, Mangano MG, Alissa A, Carr TR (2002) Sequence stratigraphic and sedimentologic significance of biogenic structures from a Late Paleozoic marginal- to open-marine reservoir, Morrow Sandstone, subsurface of southwest Kansas, USA. Sed Geol 152:99–132
Buggisch W, Joachimski M, Lützner H, Thiedig F, Hüneke H (2001) Conodont stratigraphy of the Carboniferous and Permian strata from Brøggerhalvøya and the Billefjorden Trough (Svalbard). Geol Jb B91:637–689
Burov JP, Gavrilov BP, Klubov BA, Pavlov AV, Ustrickij VI (1965) Novye dannye o verchne-permskich otloženijach Špicbergena. (New data on Upper Permian deposits of Spitsbergen). In: Sokolov VN (ed) Materialy po geologii Špicbergena. Leningrad, Russia
Cecchi C, Markello J, Waite L (1995) Sequence stratigraphy architecture of Carboniferous-Permian sedimentary systems of the Norwegian Barents Sea with comparison to coeval systems of the USA. In: Steel RJ, Felt VL, Johannesen EP, Mathieu C (eds) Sequence stratigraphy on the Northwest European Margin, special publication Nr 5. Elsevier, Amsterdam, pp 545–569
Chwieduk E (2007) Middle Permian rugose corals from the Kapp Starostin Formation, South Spitsbergen (Treskelen Peninsula). Acta Geol Pol 57:281–304
Cutbill JL, Challinor A (1965) Revision of the stratigraphical scheme for the Carboniferous and Permian rocks of Spitsbergen and Bjørnøya. Geol Mag 102:418–439
Dallmann W, Gjelberg JG, Harland WB, Johannessen EP, Keilen HB, Lønøy A, Nilsson I, Worsley D (1999) The upper palaeozoic lithostratigraphy. In: Dallmann W (ed) Lithostratigraphic lexicon of Svalbard. Norsk Polarinstitutt, Tromsø, pp 25–126
Droser MJ (1991) Ichnofabric of the paleozoic Skolithos Ichnofacies and the nature and distribution of Skolithos piperock. Palaios 6:316–325
Dunham RJ (1962) Classification of carbonate rocks according to depositional texture. In: Ham WE (ed) Classification of carbonate rocks. A symposium. AAPG Memoir, vol 1, pp 108–171
Ehrenberg SN, Pickard NAH, Svånå TA, Nilsson I, Davydov VI (2000) Sequence stratigraphy of the inner Finnmark carbonate platform, Barents Sea: correlation between well 7128/6-1 and the shallow IKU wells: Norsk Geologisk Tidsskrift, vol 80, pp 129–161
Ehrenberg SN, Pickard NAH, Henriksen LB, Svånå TA, Gutteridge P, Macdonald D (2001) A depositional and sequence stratigraphic model for cold-water, spiculitic strata based on the Kapp Starostin Formation (Permian) of Spitsbergen and equivalent deposits from the Barents Sea. AAPG Bull 85:2061–2087
Ezaki Y (1997) Cold-water Permian Rugosa and their extinction in Spitsbergen. Bol R Soc Esp Hist Nat (Sec Geol) 92:381–388
Ezaki Y, Kawamura T (1992) Carboniferous-Permian corals from Skansen and Festningen, central Spitsbergen: their faunal characteristics. In: Nakamura K (ed) Investigations on the Upper Carboniferous—Upper Permian succession of west Spitsbergen 1989–1991 by Japanese-Norwegian Research Group. Hokkaido University, Faculty of Science, Department of Geology and Mineralogy, Sapporo, pp 1–134
Flügel E (2004) Microfacies of carbonate rocks. Springer, Berlin Heidelberg New York
Folk RL (1959) Practical classification of limestones. AAPG Bull 43:1–38
Forbes CL, Harland WB, Hughes NF (1958) Palaeontological evidence for the age of the Carboniferous and Permian rocks of Central Vestspitsbergen. Geol Mag 95:463–490
Fredriksen K (1988) Sedimentologiske og diagenetiske undersøkelser av Kapp Starostin Formasjonen på Akseløya og Mariaholmen, Bellsund, Svalbard. MSc thesis, University of Tromsø, Norway
Gammon PR, James NP (2001) Palaeogeographical influence on Late Eocene biosiliceous sponge-rich sedimentation, southern Western Australia. Sedimentology 48:559–584
Gee ER, Harland WB, McWhae JRH (1952) Geology of central Vestspitsbergen. Part I: review of the geology of Spitsbergen with special reference to central Vestspitsbergen. Trans R Soc Edinb 52:299–356
Gobbett D (1963) Carboniferous and Permian brachiopods of Svalbard. Norsk Polarinstitutt Skrifter 127:1–201
Golonka J (2002) Plate-tectonic maps of the Phanerozoic. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM Spec Publ 72:21–70
Groen RD (2010) From a restricted carbonate platform to a temperate, storm-dominated ramp: the onset of the Permian Chert Event in central Spitsbergen. MSc thesis, VU University, Amsterdam, The Netherlands
Hageman SJ, Bock PE, Bone Y, McGrowran B (1998) Bryozoan growth habits: classification and analysis. J Paleontol 72:418–436
Hellem T (1980) En sedimentologisk og diagenetisk undersøkelse av utvalgte profiler fra Tempelfjordgruppen (Perm) i Isfjordområdet, Spitsbergen. MSc thesis, University of Oslo, Norway
Hüneke H, Joachimski M, Buggisch W, Lützner H (2001) Marine carbonate facies in response to climate and nutrient level: the Upper Carboniferous and Permian of Central Spitsbergen (Svalbard). Facies 45:93–136
Immenhauser A (2009) Estimating palaeo-water depth from the physical rock record. Earth Sci Rev 96:107–139
Johannessen EP, Steel RJ (1992) Mid-Carboniferous extension and rift-infill sequences in the Billefjorden Trough, Svalbard. Nor Geol J 72:35–48
Kabanov P, Anadon P, Krumbein WE (2008) Microcodium: an extensive review and a proposed non-rhizogenic biologically induced origin for its formation. Sed Geol 205:79–99
Keilen HB (1992) Lower Permian sedimentary sequences in Central Spitsbergen, Svalbard. In: Nakamura K (ed) Investigations on the Upper Carboniferous-Upper Permian succession of West Spitsbergen. Department of Geology and Mineralogy, Faculty of Sciences, Hokkaido University, Japan, pp 127–134
Klappa CF (1980) Rhizoliths in terrestrial carbonates: classification, genesis and significance. Sedimentology 27:613–629
Larssen GB, Elvebakk G, Henriksen LB, Kristensen S-E, Nilsson I, Samuelsberg TJ, Svånå TA, Stemmerik L, Worsley D (2005) Upper Palaeozoic lithostratigraphy of the southern part of the Norwegian Barents Sea. NGU Bull 444:3–43
Lauritzen Ø (1981a) Investigations of Carboniferous and Permian sediments in Svalbard I. The development of the Gipshuken Formation (Lower Permian) at Trollfuglfjella in Central Spitsbergen, Svalbard. Norsk Polarinstitutt Skrifter 176:1–22
Lauritzen Ø (1981b) Investigations of Carboniferous and Permian sediments in Svalbard II. The Carboniferous and Permian stratigraphy of the Wahlenbergfjorden area, Nordaustlandet, Svalbard. Norsk Polarinstitutt Skrifter 176:23–44
Malkowski K (1988) Paleoecology of Productacea (Brachiopoda) from the Permian Kapp Starostin Formation, Spitsbergen. Pol Polar Res 9:3–60
Mangerud G, Konieczny RM (1991) Palynological investigations of Permian rocks from Nordaustlandet, Svalbard. Polar Res 9:155–167
Mangerud G, Konieczny RM (1993) Palynology of the Permian succession of Spitsbergen, Svalbard. Polar Res 12:65–93
Mørk A, Embry AF, Weitschat W (1989) Triassic transgressive-regressive cycles in the Sverdrup Basin, Svalbard, and the Barents Shelf. In: Collinson JD (ed) Correlation in hydrocarbon exploration. Graham and Trotman, for the Norwegian Petroleum Society, London, pp 113–130
Mount J (1985) Mixed siliciclastic and carbonate sediments: a proposed first-order textural and compositional classification. Sedimentology 32:435–442
Nakamura K, Kimura G, Winsnes TS (1987) Brachiopod zonation and age of the Permian Kapp Starostin Formation (Central Spitsbergen). Polar Res 5:207–219
Nakrem H (1988) Permian bryozoans from southern Spitsbergen and Bjørnøya. A review of bryozoans described by J. Malecki (1968, 1977). Polar Res 6:113–121
Nakrem H (1991) Distribution of conodonts through the Permian succession of Svalbard. Geonytt 1:38–39
Nakrem H (1994a) Bryozoans from the Lower Permian Vøringen Member (Kapp Starostin Formation), Spitsbergen, Svalbard. Norsk Polarinstitutt Skrifter 196:1–93
Nakrem H (1994b) Middle Carboniferous to Early Permian bryozoans from Spitsbergen. Acta Geol Pol 39:45–116
Nakrem H, Spjeldnæs N (1995) Ramipora hochstetteri Toula, 1875 (Bryozoa, Cystoporata) from the Permian of Svalbard. J Paleontol 69:831–838
Nakrem H, Nilsson I, Mangerud G (1992) Permian biostratigraphy of Svalbard (Arctic Norway)—a review. Int Geol Rev 34:933–959
Nathorst AG (1910) Beiträge zur Geologie der Bäreninsel, Spitzbergens und des König-Karl-Landes. Bull Geologiska Institutionen Universitetet i Upps 10:261–416
Nichols G (1999) Sedimentology and stratigraphy. Blackwell Science, London
Nowinski A (1997) A new trachypsammiid cnidarian from the Late Permian of Spitsbergen. Pol Polar Res 18:159–169
Olempska E, Blaszyk J (1996) Ostracods from the Permian of Spitsbergen. Pol Polar Res 17:3–60
Palma RM, Lopez-Gomez J, Piethe RD (2007) Oxfordian ramp system (La Manga Formation) in the Bardas Blancas area (Mendoza Province) Neuquen Basin, Argentina: facies and depositional sequences. Sed Geol 195:113–134
Pickard NAH, Eilertsen F, Hanken N-M, Johansen TA, Lønøy A, Nakrem HA, Nilsson I, Samuelsberg TG, Somerville ID (1996) Stratigraphic framework of Upper Carboniferous (Moscovian-Kasimovian) strata in Bünsow Land, central Spitsbergen: palaeogeographic implications. Nor Geol J 76:169–185
Reid CM, James NP, Beauchamp B, Kyser TK (2007) Faunal turnover and changing oceanography: Late Palaeozoic warm-to-cool water carbonates, Sverdrup Basin, Canadian Arctic Archipelago. Palaeogeogr Palaeoclimatol Palaeoecol 249:128–159
Riding RE (1999) The term stromatolites: towards an essential definition. Lethaia 32:321–330
Samuelsberg T, Pickard N (1999) Upper Carboniferous to Lower Permian transgressive-regressive sequences of central Spitsbergen, Arctic Norway. Geol J 34:393–411
Schlager W (2000) Sedimentation rates and growth potential of tropical, cool-water and mud-mound carbonate systems. Geol Soc Lond Spec Publ 178:217–227
Scotese CR, Langford RP (1995) Pangaea and the paleogeography of the Permian. In: Scholle PA, Peryt TM, Ulmer-Scholle DS (eds) The Permian of northern Pangaea. Paleogeography, paleoclimates, stratigraphy, vol I. Springer, Berlin Heidelberg New York, pp 3–19
Shen S, Tazawa J, Shi G (2005) Carboniferous and Permian Rugosochonetidae (Brachiopoda) from West Spitsbergen. Alcheringa 29:241–256
Sosipatrova GP (1967) Verchnepaleozojskie foraminifery Špicbergena. (Upper Paleozoic foraminifers of Spitsbergen.). Materialy po stratigrafii Špicbergena. NIIGA, pp 94–120
Steel RJ, Worsley D (1984) Svalbard’s post-Caledonian strata—an atlas of sedimentational patterns and palaeogeographic evolution. In: Spencer AM, Holter E, Johnson SO, Mørk A, Nysæther E, Songstad P, Spinnangr Å (eds) Petroleum geology of the North European Margin. Graham & Trotman Ltd, London, pp 109–135
Stemmerik L (1988) Discussion. Brachiopod zonation and age of the Permian Kapp Starostin Formation (Central Spitsbergen). Polar Res 6:179–180
Stemmerik L (1997) Permian (Artinskian-Kazanian) cool-water carbonates in North Greenland, Svalbard and the western Barents Sea. In: James NP, Clarke JAD (eds) Cool-water carbonates. SEPM Spec Publ 56:349–364
Stemmerik L, Worsley D (1989) Late Palaeozoic sequence correlations, North Greenland, Svalbard and the Barents Shelf. In: Collinson JD (ed) Correlation in hydrocarbon exploration. Norwegian Petroleum Society, Graham & Trotman, London, pp 99–111
Szaniawski H, Malkowski K (1979) Conodonts from the Kapp Starostin Formation (Permian) of Spitsbergen. Acta Geol Pol 24:231–264
Tidten G (1972) Morphologisch-ontogenetische Untersuchungen an Pterocorallia aus dem Permo-Karbon von Spitzbergen. Palaeontogr A 139:1–63
Vossler SM, Pemberton SG (1988) Skolithos in the Upper Cretaceous Cardium Formation—an ichnofossil example of opportunistic ecology. Lethaia 21:351–362
Wardlaw BR, Davydov V, Gradstein FM (2004) The Permian period. In: Gradstein F, Ogg J, Smith A (eds) A geologic Time Scale 2004. Cambridge University Press, Cambridge, pp 249–270
Wentworth CK (1922) A scale of grade and class terms for clastic sediments. J Geol 30:377–392
Wignall PB, Morante R, Newton R (1998) The Permo-Triassic transition in Spitsbergen: δ13C chemostratigraphy, Fe and S geochemistry, facies, fauna and trace fossils. Geol Mag 135:47–62
Winsnes T (1966) Observations on the Carboniferous and Permian rocks of Vestspitsbergen. Norsk Polarinstitutt Årbok 1964:7–29
Worsley D, Aga OJ, Dalland A, Elverhøi A, Thon A (1986) The geological history of Svalbard, evolution of an Arctic archipelago. Statoil, Stavanger
Ziegler AM, Gibbs MT, Hulver ML (1997) A mini-atlas of oceanic water masses in the Permian Period. In: Shi GR, Archbold NW, Grover M (eds) The Permian System: stratigraphy, palaeogeography and resources. Proc Strzelecki Symp, pp 323–343
Acknowledgments
We are grateful to Winfried Dallmann for initiating the investigations as a part of the Norwegian Polar Institute’s geological mapping program GEOKART. Steve Ehrenberg is acknowledged for a review with valuable suggestions, considerably improving an earlier manuscript. The organization and logistics department of the Norwegian Polar Institute and involved staff from Airlift are thanked for organizational and technical support during the field campaigns on Svalbard. The geological department of the University Centre in Svalbard (UNIS) is acknowledged for providing microscope and camera equipment for the facies documentation. Ralf Bätzel, technician at the University of Bremen, is thanked for the excellent preparation of numerous thin-sections forming the basis of the microfacies investigations of this publication.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Blomeier, D., Dustira, A., Forke, H. et al. Environmental change in the Early Permian of NE Svalbard: from a warm-water carbonate platform (Gipshuken Formation) to a temperate, mixed siliciclastic-carbonate ramp (Kapp Starostin Formation). Facies 57, 493–523 (2011). https://doi.org/10.1007/s10347-010-0243-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-010-0243-z